
Investigating the Potential Influence of TAS2R16 Genetic Variants and Protein Levels on Multiple Sclerosis Development

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. DNA Extraction and Genotyping
2.3. Sample Preparation for Serum Levels Measurement
2.4. Enzyme Immunoassay
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Frischer, J.M.; Bramow, S.; Dal-Bianco, A.; Lucchinetti, C.F.; Rauschka, H.; Schmidbauer, M.; Laursen, H.; Sorensen, P.S.; Lassmann, H. The relation between inflammation and neurodegeneration in multiple sclerosis brains. Brain 2009, 132, 1175–1189. [Google Scholar] [CrossRef]
- Kamel, F. Factors involved in relapse of multiple sclerosis. J. Microsc. Ultrastruct. 2019, 7, 103. [Google Scholar] [CrossRef]
- Walton, C.; King, R.; Rechtman, L.; Kaye, W.; Leray, E.; Marrie, R.A.; Robertson, N.; La Rocca, N.; Uitdehaag, B.; Van Der Mei, I.; et al. Rising prevalence of multiple sclerosis worldwide: Insights from the Atlas of MS, third edition. Mult. Scler. J. 2020, 26, 1816–1821. [Google Scholar] [CrossRef] [PubMed]
- Nourbakhsh, B.; Mowry, E.M. Multiple Sclerosis Risk Factors and Pathogenesis. Contin. Lifelong Learn. Neurol. 2019, 25, 596–610. [Google Scholar] [CrossRef]
- Brynedal, B.; Duvefelt, K.; Jonasdottir, G.; Roos, I.M.; Akesson, E.; Palmgren, J.; Hillert, J. HLA-A confers an HLA-DRB1 independent influence on the risk of multiple sclerosis. PLoS ONE 2007, 2, e664. [Google Scholar] [CrossRef] [PubMed]
- Agliardi, C.; Alfredsson, L.; Alizadeh, M.; Anderson, C.; Andrews, R.; Søndergaard, H.B.; Baker, A.; Band, G.; Baranzini, S.E. Analysis of immune-related loci identifies 48 new susceptibility variants for multiple sclerosis. Nat. Genet. 2013, 45, 1353–1360. [Google Scholar]
- The International Multiple Sclerosis Genetics Consortium; The Wellcome Trust Case Control Consortium 2. Genetic risk and a primary role for cell-mediated immune mechanisms in multiple sclerosis. Nature 2011, 476, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Database, G.; TAS2R16 Gene-GeneCards|T2R16 Protein|T2R16 Antibody. Genecards.org. 2022. Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=TAS2R16 (accessed on 5 September 2023).
- National Center for Biotechnology Information. Database SNP, rs860170. 2022. Available online: https://www.ncbi.nlm.nih.gov/snp/rs860170 (accessed on 30 May 2023).
- National Center for Biotechnology Information. Database SNP, rs978739. 2022. Available online: https://www.ncbi.nlm.nih.gov/snp/rs978739 (accessed on 30 May 2023).
- National Center for Biotechnology Information. Database SNP, rs1357949. 2022. Available online: https://www.ncbi.nlm.nih.gov/snp/rs1357949 (accessed on 30 May 2023).
- Barontini, J.; Antinucci, M.; Tofanelli, S.; Cammalleri, M.; Monte, M.D.; Gemignani, F.; Vodicka, P.; Marangoni, R.; Vodickova, L.; Kupcinskas, J.; et al. Association between polymorphisms of TAS2R16 and susceptibility to colorectal cancer. BMC Gastroenterol. 2017, 17, 104. [Google Scholar] [CrossRef]
- Tuzim, K.; Korolczuk, A. An update on extra-oral bitter taste receptors. J. Transl. Med. 2021, 19, 440. [Google Scholar] [CrossRef]
- Welcome, M.O.; Mastorakis, N.E. The taste of neuroinflammation: Molecular mechanisms linking taste sensing to neuroinflammatory responses. Pharmacol. Res. 2021, 167, 105557. [Google Scholar] [CrossRef]
- Duarte, A.C.; Costa, A.R.; Gonçalves, I.; Quintela, T.; Preissner, R.; Santos, C.R.A. The druggability of bitter taste receptors for the treatment of neurodegenerative disorders. Biochem. Pharmacol. 2022, 197, 114915. [Google Scholar] [CrossRef] [PubMed]
- Carey, R.M.; Workman, A.D.; Yan, C.H.; Chen, B.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; Lee, R.J.; Cohen, N.A. Sinonasal T2R-mediated nitric oxide production in response to Bacillus cereus. Am. J. Rhinol. Allergy 2017, 31, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.H.; Hahn, S.; McMahon, D.; Bonislawski, D.; Kennedy, D.W.; Adappa, N.D.; Palmer, J.N.; Jiang, P.; Lee, R.J.; Cohen, N.A. Nitric oxide production is stimulated by bitter taste receptors ubiquitously expressed in the sinonasal cavity. Am. J. Rhinol. Allergy 2017, 31, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Dehkordi, O.; Rose, J.E.; Fatemi, M.; Allard, J.S.; Balan, K.V.; Young, J.K.; Fatima, S.; Millis, R.M.; Jayam-Trouth, A. Neuronal expression of bitter taste receptors and downstream signaling molecules in the rat brainstem. Brain 2012, 1475, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Voigt, A.; Bojahr, J.; Narukawa, M.; Hübner, S.; Boehm, U.; Meyerhof, W. Transsynaptic Tracing from Taste Receptor Cells Reveals Local Taste Receptor Gene Expression in Gustatory Ganglia and Brain. J. Neurosci. 2015, 35, 9717–9729. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.J.; Park, J.H.; Choi, J.S.; Chun, M.H.; Moon, Y.W.; Lee, M.Y. Enhanced expression of the sweet taste receptors and alpha-gustducin in reactive astrocytes of the rat hippocampus following ischemic injury. Neurochem. Res. 2010, 35, 1628–1634. [Google Scholar] [CrossRef] [PubMed]
- Liman, E.R.; Zhang, Y.V.; Montell, C. Peripheral coding of taste. Neuron 2014, 81, 984–1000. [Google Scholar] [CrossRef]
- Singh, N.; Vrontakis, M.; Parkinson, F.; Chelikani, P. Functional bitter taste receptors are expressed in brain cells. Biochem. Biophys. Res. Commun. 2011, 406, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Aliaga, E.S.; Barkhof, F. MRI mimics of multiple sclerosis. Handb. Clin. Neurol. 2014, 122, 291–316. [Google Scholar]
- Campa, D.; De Rango, F.; Carrai, M.; Crocco, P.; Montesanto, A.; Canzian, F.; Rose, G.; Rizzato, C.; Passarino, G.; Barale, R. Bitter Taste Receptor Polymorphisms and Human Aging. PLoS ONE 2012, 7, e45232. [Google Scholar] [CrossRef]
- Malovini, A.; Accardi, G.; Aiello, A.; Bellazzi, R.; Candore, G.; Caruso, C.; Ligotti, M.E.; Maciag, A.; Villa, F.; Puca, A.A. Taste receptors, innate immunity and longevity: The case of TAS2R16 gene. Immun. Ageing 2019, 16, 5. [Google Scholar] [CrossRef]
- Carrai, M.; Steinke, V.; Vodicka, P.; Pardini, B.; Rahner, N.; Holinski-Feder, E.; Morak, M.; Schackert, H.K.; Görgens, H.; Stemmler, S.; et al. Association between TAS2R38 gene polymorphisms and colorectal cancer risk: A case-control study in two independent populations of Caucasian origin. PLoS ONE 2011, 6, e20464. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Lee, J.; Choi, I.J.; Kim, Y.W.; Ryu, K.W.; Kim, J. Variations in TAS1R taste receptor gene family modify food intake and gastric cancer risk in a Korean population. Mol. Nutr. Food Res. 2016, 60, 2433–2445. [Google Scholar] [CrossRef]
- Choi, J.H.; Lee, J.; Choi, I.J.; Kim, Y.W.; Ryu, K.W.; Kim, J. Genetic Variation in the TAS2R38 Bitter Taste Receptor and Gastric Cancer Risk in Koreans. Sci. Rep. 2016, 6, 26904. [Google Scholar] [CrossRef]
- Bufe, B.; Hofmann, T.; Krautwurst, D.; Raguse, J.D.; Meyerhof, W. The human TAS2R16 receptor mediates bitter taste in response to beta-glucopyranosides. Nat. Genet. 2002, 32, 397–401. [Google Scholar] [CrossRef]
- Vane, J.R. The fight against rheumatism: From willow bark to COX-1 sparing drugs. J. Physiol. Pharmacol. 2000, 51 Pt 1, 573–586. [Google Scholar] [PubMed]
- Kim, E.R.; Chang, D.K. Colorectal cancer in inflammatory bowel disease: The risk, pathogenesis, prevention and diagnosis. World J. Gastroenterol. 2014, 20, 9872–9881. [Google Scholar] [CrossRef]
- Schembre, S.M.; Cheng, I.; Wilkens, L.R.; Albright, C.L.; Le Marchand, L. Variations in bitter-taste receptor genes, dietary intake, and colorectal adenoma risk. Nutr. Cancer 2013, 65, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Xi, R.; Liu, J.; Peng, X.; Zhao, L.; Zhou, X.; Li, J.; Zheng, X.; Xu, X. TAS2R16 Activation Suppresses LPS-Induced Cytokine Expression in Human Gingival Fibroblasts. Front. Immunol. 2021, 12, 726546. [Google Scholar] [CrossRef]
- Buzzard, K.; Chan, W.H.; Kilpatrick, T.; Murray, S. Multiple Sclerosis: Basic and Clinical. Adv. Neurobiol. 2017, 15, 211–252. [Google Scholar]
- Giovannoni, G.; Cutter, G.R.; Lunemann, J.; Martin, R.; Münz, C.; Sriram, S.; Steiner, I.; Hammerschlag, M.R.; Gaydos, C.A. Infectious causes of multiple sclerosis. Lancet Neurol. 2006, 5, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Slots, J. Herpesviruses in periodontal diseases. Periodontol. 2000 2005, 38, 33–62. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.-J.; Lin, H.-C. Association between multiple sclerosis and chronic periodontitis: A population-based pilot study. Eur. J. Neurol. 2013, 20, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Dalesio, N.M.; Sebastian Pluznick, J.L.; Berkowitz, D.E. Olfactory, Taste, and Photo Sensory Receptors in Non-sensory Organs: It Just Makes Sense. Front. Physiol. 2018, 9, 424638. [Google Scholar] [CrossRef] [PubMed]
- Wölfle, U.; Haarhaus, B.; Kersten, A.; Fiebich, B.; Hug, M.J.; Schempp, C.M. Salicin from Willow Bark can Modulate Neurite Outgrowth in Human Neuroblastoma SH-SY5Y Cells. Phytother. Res. 2015, 29, 1494–1500. [Google Scholar] [CrossRef]
- Manchery, N.; Henry, J.D.; Nangle, M.R. A systematic review of oral health in people with multiple sclerosis. Community Dent. Oral Epidemiol. 2019, 48, 89–100. [Google Scholar] [CrossRef]


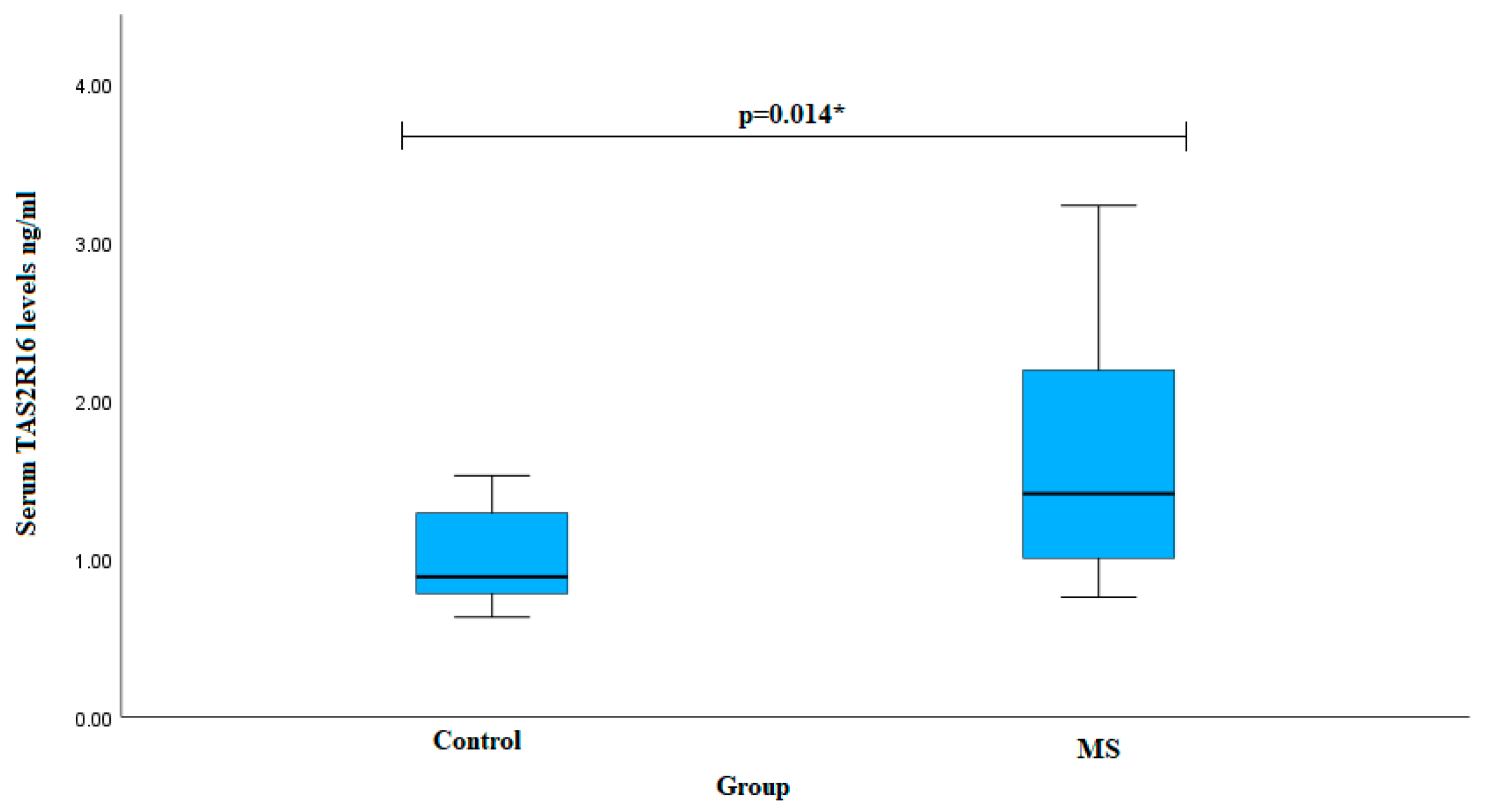{kind=link}
| Characteristic | Group | |||
|---|---|---|---|---|
| MS Group | Control Group | p-Value | ||
| Gender | Females, n (%) | 111 (50.9) | 169 (63.8) | 0.004 * |
| Males, n (%) | 107 (49.1) | 96 (36.2) | ||
| Age median (IQR) | 38 (15.25) | 36 (27.5) | 0.568 ** | |
| SNP | MS Group n (%) (n = 218) | Control Group n (%) (n = 265) | p-Value |
|---|---|---|---|
| Genotype | |||
| TT | 63 (28.9) 1 | 100 (37.7) 1 | <0.001 |
| CT | 138 (63.3) | 164 (61.9) | |
| CC | 17 (7.8) 2 | 1 (0.4) 2 | |
| Total | 218 (100) | 265 (100) | |
| Allele | |||
| T | 264 (60.6) | 364 (68.7) | 0.008 |
| C | 172 (39.4) | 166 (31.3) |
| SNP | MS Group n (%) (n = 218) | Control Group n (%) (n = 265) | p-Value |
|---|---|---|---|
| TAS2R16 rs978739 | |||
| Genotype | |||
| TT | 104 (47.7) | 142 (53.6) | 0.222 |
| CT | 99 (45.4) | 100 (37.7) | |
| CC | 15 (6.9) | 23 (8.7) | |
| Total | 218 (100) | 265 (100) | |
| Allele | |||
| T | 307 (70.4) | 384 (72.5) | 0.484 |
| C | 129 (29.6) | 146 (27.5) | |
| TAS2R16 rs1357949 | |||
| Genotype | |||
| AA | 102 (46.8) | 118 (44.5) | 0.884 |
| AG | 94 (43.1) | 119 (44.9) | |
| GG | 22 (10.1) | 28 (10.6) | |
| Total | 218 (100) | 265 (100) | |
| Allele | |||
| A | 298 (68.3) | 355 (67.0) | 0.651 |
| G | 138 (31.7) | 175 (33.0) |
| Model | Genotype/Allele | OR (95% CI) | p-Value | AIC |
|---|---|---|---|---|
| Codominant | CT vs. TT CC vs. TT | 1.336 (0.906–1.969) 26.984 (3.504–207.790) | 0.144 0.002 | 645.637 |
| Dominant | CC + CT vs. TT | 1.491 (1.016–2.189) | 0.041 | 662.795 |
| Over-dominant | CT vs. TT + CC | 1.062 (0.733–1.539) | 0.749 | 666.897 |
| Recessive | CC vs. CT + TT | 22.328 (2.947–169.181) | 0.003 | 645.789 |
| Additive | C | 1.792 (1.267–2.533) | <0.001 | 655.774 |
| Model | Genotype/Allele | OR (95% CI) | p-Value | AIC |
|---|---|---|---|---|
| TAS2R16 rs978739 | ||||
| Codominant | CT vs. TT CC vs. TT | 1.352 (0.928–1.968) 0.890 (0.443–1.789) | 0.116 0.745 | 666.985 |
| Dominant | CC + CT vs. TT | 1.265 (0.884–1.812) | 0.199 | 666.345 |
| Over-dominant | CT vs. TT + CC | 1.373 (0.953–1.976) | 0.088 | 664.092 |
| Recessive | CC vs. CT + TT | 0.777 (0.395–1.530) | 0.466 | 666.461 |
| Additive | C | 1.107 (0.834–1.467) | 0.482 | 666.505 |
| TAS2R16 rs1357949 | ||||
| Codominant | AG vs. AA GG vs. AA | 0.914 (0.626–1.334) 0.909 (0.490–1.686) | 0.641 0.762 | 668.753 |
| Dominant | GG + AG vs. AA | 0.913 (0.637–1.308) | 0.620 | 666.753 |
| Over-dominant | AG vs. AA + GG | 0.930 (0.648–1.335) | 0.694 | 666.845 |
| Recessive | GG vs. AG + AA | 0.950 (0.527–1.713) | 0.865 | 666.970 |
| Additive | G | 0.939 (0.715–1.233) | 0.650 | 666.794 |
| SNP | MS Group n (%) (n = 111) | Control Group n (%) (n = 169) | p-Value |
|---|---|---|---|
| TAS2R16 rs860170 | |||
| Genotype | |||
| TT | 32 (28.8) | 68 (40.2) | <0.001 |
| CT | 69 (62.2) | 100 (59.2) | |
| CC | 10 (9) 1 | 1 (0.6) 1 | |
| Total | 111 (100) | 169 (100) | |
| Allele | |||
| T | 133 (59.9) | 236 (69.8) | 0.010 |
| C | 89 (40.1)) | 102 (30.2) |
| SNP | MS Group n (%) (n = 111) | Control Group n (%) (n = 169) | p-Value |
|---|---|---|---|
| TAS2R16 rs978739 | |||
| Genotype | |||
| TT | 48 (43.2) | 87 (51.5) | 0.341 |
| CT | 53 (47.7) | 66 (39.1) | |
| CC | 10 (9) | 16 (9.5) | |
| Total | 111 (100) | 169 (100) | |
| Allele | |||
| T | 149 (67.1) | 240 (71) | 0.328 |
| C | 73 (32.9) | 98 (29) | |
| TAS2R16 rs1357949 | |||
| Genotype | |||
| AA | 50 (45) | 76 (45) | 0.979 |
| AG | 50 (45) | 75 (44.4) | |
| GG | 11 (9.9) | 18 (10.7) | |
| Total | 111 (100) | 169 (100) | |
| Allele | |||
| A | 150 (67.6) | 227 (67.2) | 0.920 |
| G | 72 (32.4) | 111 (32.8) |
| Model | Genotype/Allele | OR (95% CI) | p-Value | AIC |
|---|---|---|---|---|
| Codominant | CT vs. TT CC vs. TT | 1.466 (0.872–2.467) 21.250 (2.607–173.207) | 0.149 0.004 | 364.641 |
| Dominant | CC + CT vs. TT | 1.662 (0.995–2.776) | 0.052 | 374.211 |
| Over-dominant | CT vs. TT + CC | 1.134 (0.694–1.852) | 0.617 | 377.810 |
| Recessive | CC vs. CT + TT | 16.634 (2.098–131.870) | 0.008 | 364.750 |
| Additive | C | 1.995 (1.259–3.160) | 0.003 | 369.019 |
| Model | Genotype/Allele | OR (95% CI) | p-Value | AIC |
|---|---|---|---|---|
| TAS2R16 rs978739 | ||||
| Codominant | CT vs. TT CC vs. TT | 1.455 (0.879–2.411) 1.133 (0.477–2.691) | 0.145 0.778 | 377.913 |
| Dominant | CC + CT vs. TT | 1.393 (0.860–2.254) | 0.178 | 376.237 |
| Over-dominant | CT vs. TT + CC | 1.426 (0.879–2.314) | 0.151 | 375.993 |
| Recessive | CC vs. CT + TT | 0.947 (0.413–2.169) | 0.897 | 378.044 |
| Additive | C | 1.201 (0.832–1.733) | 0.329 | 377.107 |
| TAS2R16 rs1357949 | ||||
| Codominant | AG vs. AA GG vs. AA | 1.013 (0.611–1.680) 0.929 (0.405–2.132) | 0.959 0.862 | 380.018 |
| Dominant | GG + AG vs. AA | 0.997 (0.616–1.613) | 0.990 | 378.061 |
| Over-dominant | AG vs. AA + GG | 1.027 (0.635–1.663) | 0.913 | 378.049 |
| Recessive | GG vs. AG + AA | 0.923 (0.418–2.036) | 0.842 | 378.021 |
| Additive | G | 0.981 (0.682–1.412) | 0.919 | 378.050 |
| SNP Genotypes/Alleles | MS Group n (%) (n = 107) | Control Group n (%) (n = 96) | p-Value |
|---|---|---|---|
| Genotype | |||
| TT | |||
| CT | 31 (29) | 32 (33.3) | 0.036 |
| CC | 69 (64.5) | 64 (66.7) | |
| Total | 7 (6.5) | 0 (0) | |
| Allele | 107 (100) | 96 (100) | |
| T | |||
| C | 131 (61.2) | 128 (66.7) | 0.254 |
| 83 (38.8) | 64 (33.3) |
| SNP | MS Group n (%) (n = 107) | Control Group n (%) (n = 96) | p-Value |
|---|---|---|---|
| TAS2R16 rs978739 | |||
| Genotype | |||
| TT | 56 (52.3) | 55 (57.3) | 0.461 |
| CT | 46 (43) | 34 (35.4) | |
| CC | 5 (4.7) | 7 (7.3) | |
| Total | 107 (100) | 96 (100) | |
| Allele | |||
| T | 158 (73.8) | 144 (75) | 0.788 |
| C | 56 (26.2) | 48 (25) | |
| TAS2R16 rs1357949 | |||
| Genotype | |||
| AA | 52 (48.6) | 42 (43.8) | 0.772 |
| AG | 44 (41.1) | 44 (45.8) | |
| GG | 11 (10.3) | 10 (10.4) | |
| Total | 107 (100) | 96 (100) | |
| Allele | |||
| A | 148 (69.2) | 128 (66.7) | 0.591 |
| G | 66 (30.8) | 64 (33.3) |
| Model | Genotype/Allele | OR (95% CI) | p-Value | AIC |
|---|---|---|---|---|
| TAS2R16 rs860170 | ||||
| Codominant | CT vs. TT CC vs. TT | 1.113 (0.611–2.027) - | 0.727 NA | 275.510 |
| Dominant | CC + CT vs. TT | 1.226 (0.676–2.223) | 0.503 | 282.372 |
| Over-dominant | CT vs. TT + CC | 0.908 (0.508–1.622) | 0.744 | 282.715 |
| Recessive | CC vs. CT + TT | NA | NA | 273.632 |
| Additive | C | 1.505 (0.879–2.578) | 0.136 | 280.575 |
| TAS2R16 rs978739 | ||||
| Codominant | CT vs. TT CC vs. TT | 1.329 (0.745–2.370) 0.720 (0.210–2.344) | 0.336 0.565 | 283.267 |
| Dominant | CC + CT vs. TT | 1.222 (0.702–2.127) | 0.479 | 282.320 |
| Over-dominant | CT vs. TT + CC | 1.375 (0.780–2.242) | 0.271 | 281.603 |
| Recessive | CC vs. CT + TT | 0.623 (0.191–2.033) | 0.433 | 282.197 |
| Additive | C | 1.066 (0.676–1.679) | 0.784 | 282.746 |
| TAS2R16 rs1357949 | ||||
| Codominant | AG vs. AA GG vs. AA | 0.808 (0.451–1.447) 0.888 (0.344–2.293) | 0.473 0.807 | 284.304 |
| Dominant | GG + AG vs. AA | 0.823 (0.473–1.431) | 0.489 | 282.343 |
| Over-dominant | AG vs. AA + GG | 0.825 (0.473–1.440) | 0.499 | 282.364 |
| Recessive | GG vs. AG + AA | 0.985 (0.399–2.435) | 0.975 | 282.820 |
| Additive | G | 0.892 (0.588–1.354) | 0.592 | 282.534 |
| Haplotype | TAS2R16 rs860170 | TAS2R16 rs978739 | TAS2R16 rs1357949 | Frequency (%) | OR (95% CI) | p-Value | |
|---|---|---|---|---|---|---|---|
| Control | MS | ||||||
| 1 | C | T | A | 30.78 | 27.2 | 1.00 | – |
| 2 | T | T | G | 31.28 | 19.15 | 0.76 (0.49–1.20) | 0.240 |
| 3 | T | C | A | 25.45 | 23.34 | 1.04 (0.67–1.63) | 0.860 |
| 4 | T | T | A | 10.39 | 14.84 | 1.52 (0.84–2.73) | 0.170 |
| 5 | T | C | G | 1.55 | 3.22 | 1.33 (0.39–4.58) | 0.650 |
| 6 | C | C | A | 0.36 | 2.97 | 12.51 (1.59–104.12) | 0.020 |
| Rare | * | * | * | NA | NA | 6.85 (0.47–98.99) | 0.160 |
| Haplotype | TAS2R16 rs860170 | TAS2R16 rs978739 | TAS2R16 rs1357949 | Frequency (%) | OR (95% CI) | p-Value | |
|---|---|---|---|---|---|---|---|
| Control | MS | ||||||
| 1 | C | T | A | 29.83 | 24.93 | 1.00 | – |
| 2 | T | T | G | 31.26 | 16.33 | 0.73 (0.39–1.38) | 0.330 |
| 3 | T | C | A | 27.41 | 22.87 | 1.03 (0.56–1.88) | 0.930 |
| 4 | T | T | A | 9.92 | 15.23 | 1.87 (0.80–4.34) | 0.150 |
| 5 | T | C | G | 1.24 | 5.48 | 3.94 (0.85–18.28) | 0.081 |
| rare | * | * | * | - | NA | 7.76 (0.64–94.25) | 0.110 |
| Haplotype | TAS2R16 rs860170 | TAS2R16 rs978739 | TAS2R16 rs1357949 | Frequency (%) | OR (95% CI) | p-Value | |
|---|---|---|---|---|---|---|---|
| Control | MS | ||||||
| 1 | C | T | A | 32.57 | 29.33 | 1.00 | – |
| 2 | T | T | G | 31.15 | 22.41 | 0.87 (0.44–1.72) | 0.690 |
| 3 | T | C | A | 22.05 | 24.55 | 1.27 (0.62–2.58) | 0.520 |
| 4 | T | T | A | 11.28 | 14.25 | 1.43 (0.60–3.37) | 0.420 |
| 5 | T | C | G | 2.19 | 0 | 0.20 (0.00–91.98) | 0.610 |
| Rare | * | * | * | - | - | 1.79 (0.10–32.48) | 0.690 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gedvilaite, G.; Pileckaite, E.; Ramanauskas, I.; Kriauciuniene, L.; Balnyte, R.; Liutkeviciene, R. Investigating the Potential Influence of TAS2R16 Genetic Variants and Protein Levels on Multiple Sclerosis Development. J. Pers. Med. 2024, 14, 402. https://doi.org/10.3390/jpm14040402
Gedvilaite G, Pileckaite E, Ramanauskas I, Kriauciuniene L, Balnyte R, Liutkeviciene R. Investigating the Potential Influence of TAS2R16 Genetic Variants and Protein Levels on Multiple Sclerosis Development. Journal of Personalized Medicine. 2024; 14(4):402. https://doi.org/10.3390/jpm14040402
Chicago/Turabian StyleGedvilaite, Greta, Enrika Pileckaite, Ignas Ramanauskas, Loresa Kriauciuniene, Renata Balnyte, and Rasa Liutkeviciene. 2024. "Investigating the Potential Influence of TAS2R16 Genetic Variants and Protein Levels on Multiple Sclerosis Development" Journal of Personalized Medicine 14, no. 4: 402. https://doi.org/10.3390/jpm14040402
APA StyleGedvilaite, G., Pileckaite, E., Ramanauskas, I., Kriauciuniene, L., Balnyte, R., & Liutkeviciene, R. (2024). Investigating the Potential Influence of TAS2R16 Genetic Variants and Protein Levels on Multiple Sclerosis Development. Journal of Personalized Medicine, 14(4), 402. https://doi.org/10.3390/jpm14040402