Notes on the Nest Architecture and Colony Composition in Winter of the Yellow-Legged Asian Hornet, Vespa velutina Lepeletier 1836 (Hym.: Vespidae), in Its Introduced Habitat in Galicia (NW Spain)



Abstract
:1. Introduction
- The Asian chestnut wasp (Dryocosmus kuriphilus Yasumatsu 1951 (Hymenoptera: Cynipidae)) was detected in 2004. It has caused significant losses in chestnut production and weakened vitally important chestnut agroforestry cultivars.
- The potato flea (Epitrix similaris Gentner 1944 (Coleoptera: Chrysomelidae)) and the Guatemalan moth (Tecia solanivora Povolny 1973 (Lepidoptera: Gelechiidae)) were detected in 2009 and 2015, respectively. Their presence has important implications, and the growth of the potato is now forbidden in some areas—a plan implemented by an invasive species task force.
- The red palm weevil (Rhynchophorus ferrugineus Olivier, 1790 (Coleoptera: Curculionidae)) was detected in 2013. The species actively destroys the interior of the palm tree.
- The Asian boxy moth (Cydalima perspectalis Walker, 1859 (Lepidoptera: Crambidae)) was detected in 2014. Its larvae defoliate the boxy bush.
- The African citrus psyllid (Trioza erythreae Del Guercio, 1918 (Hemiptera: Triozidae)) was detected in 2014. It weakens trees, thereby deforming them, and is a vector for various diseases.
- The yellow-legged Asian hornet (Vespa velutina Lepeletier 1836 (Hymenoptera: Vespidae)) was detected in Galicia in 2012.
2. Materials and Methods
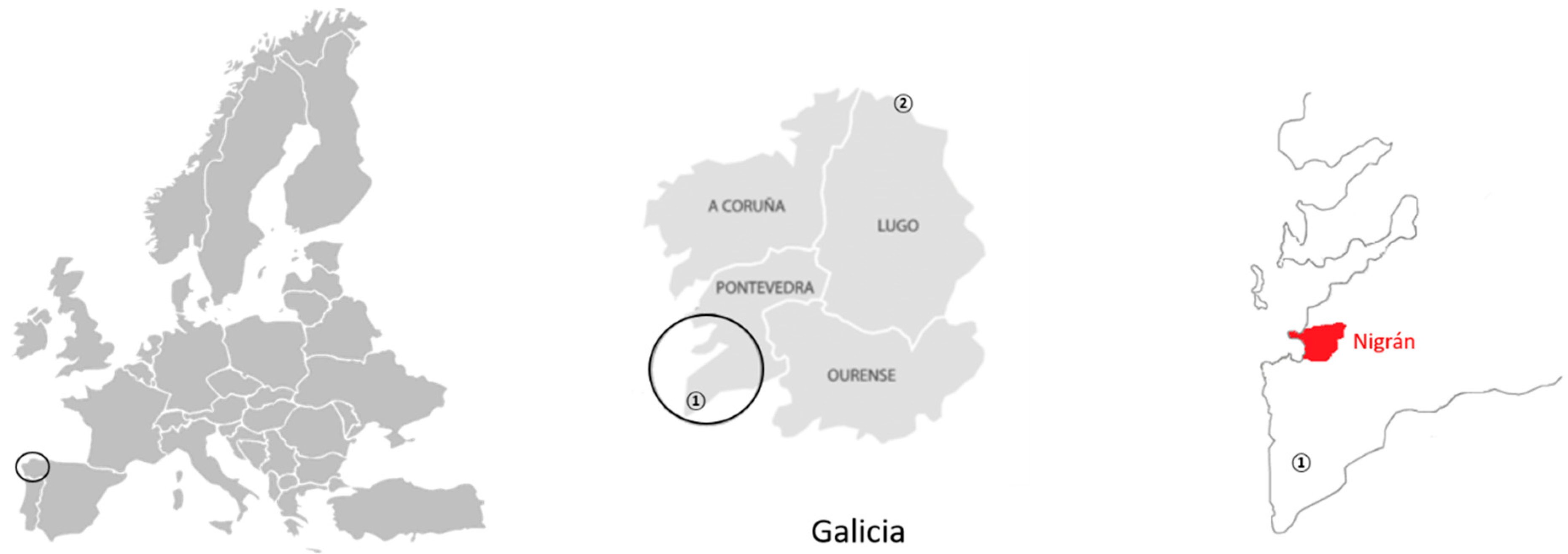
2.1. Sampling Area
2.2. Nest Collection
2.3. Nest Structural Features
2.3.1. External
2.3.2. Internal
2.4. Nest Colony Composition
3. Results
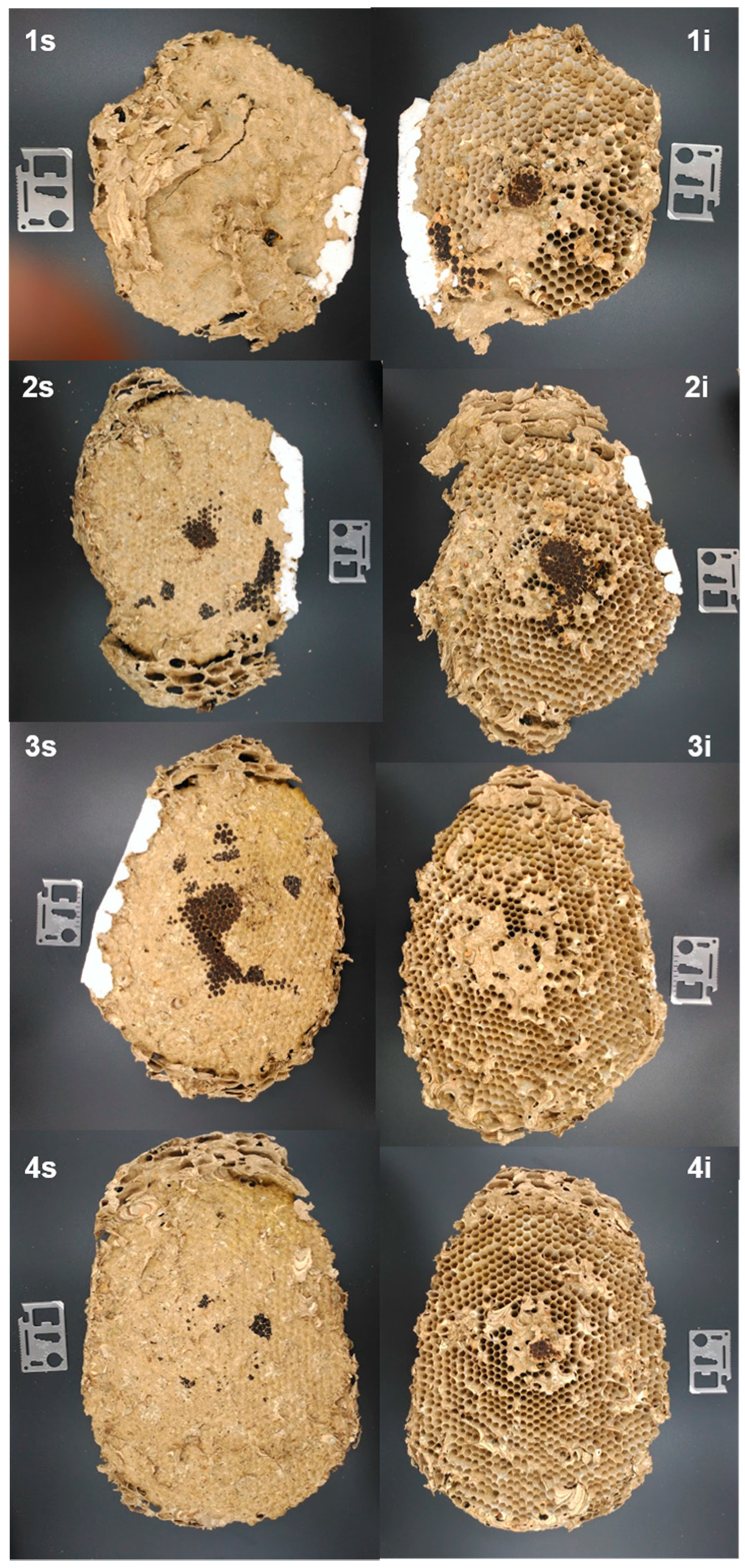
3.1. External Nest Structure
3.2. Internal Nest Structure
3.3. Nest Colony Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xunta de Galicia. Available online: https://cmatv.xunta.gal/c/document_library/get_file?file_path=/portal-web/Documentos_DXConservacion_da_Natureza/Biodiversidade/as_especies_exoticas_invasoras_en_galicia_diagnostico_e_%20proposta_de_actuacion.pdf (accessed on 12 June 2019).
- Kim, J.; Choi, M.; Moon, T. Occurrence of Vespa velutina Lepeletier from Korea, and a revised key for Korean Vespa species (Hymenoptera: Vespidae). Entomol. Res. 2006, 36, 112–115. [Google Scholar] [CrossRef]
- Haxaire, J.; Bouguet, J.-P.; Tamisier, J.-P. Vespa velutina Lepeletier, 1836, une redoutable nouveauté pour la faune de France (Hymenoptera, Vespidae). Bull. Soc. Entomol. France 2006, 111, 194. [Google Scholar]
- Minoshima, Y.; Yamane, S.K.; Ueno, T. An invasive alien hornet, Vespa velutina nigrithorax du Buysson (Hymenoptera, Vespidae), found in Kitakyushu, Kyushu Island: A first record of the species from mainland Japan. Jpn. J. Syst. Entomol. 2015, 21, 259–261. [Google Scholar]
- Takahashi, R.; Sakai, Y.; Yamamura, T.; Kiyoshi, T.; Takahashi, J. Analysis the nest of alian hornet, Vespa velutina, found for the first time in Tsushima Island, Japan. Trans. Nagasaki Biol. Soc. 2015, 76, 49–56. [Google Scholar]
- Castro, L.; Pagola-Carte, S. Vespa velutina Lepeletier, 1836 (Hymenoptera: Vespidae), recolectada en la Península Ibérica. Heteropterus Rev. Entomol. 2010, 10, 193–196. [Google Scholar]
- Grosso-Silva, J.M.; Maia, M. Vespa velutina Lepeletier, 1836 (Hymenoptera, Vespidae), new species for Portugal. Arquivos Entomolóxicos 2012, 6, 53–54. [Google Scholar]
- Rome, Q.; Perrard, A.; Muller, F.; Villemant, C. Monitoring and control modalities of a honeybee predator, the yellow-legged hornet Vespa velutina nigrithorax (Hymenoptera: Vespidae). Aliens Invasive Species Bull. 2011, 31, 7–15. [Google Scholar]
- Bertolino, S.; Lioy, S.; Laurino, D.; Manino, A.; Porporato, M. Spread of the invasive yellow-legged hornet Vespa velutina (Hymenoptera: Vespidae) in Italy. Appl. Entomol. Zool. 2016, 51, 589–597. [Google Scholar] [CrossRef]
- Witt, R. Esrtfund eines Nestes der Asiatischen Hornisse Vespa velutina Lepeletier, 1838 in Deutschland und Details zumm Nestbau (Hymenosptera, Vespinae). Ampulex 2015, 7, 42–53. [Google Scholar]
- Smit, J.; Noordijk, J.; Zeegers, T. Will the Asian hornet (Vespa velutina) settle in the Netherlands? Entomol. Ber. 2018, 78, 2–6. [Google Scholar]
- Leza, M.; Miranda, M.Á.; Colomar, V. First detection of Vespa velutina nigrithorax (Hymenoptera: Vespidae) in the Balearic Islands (Western Mediterranean): A challenging study case. Biol. Invasions 2018, 20, 1643–1649. [Google Scholar] [CrossRef]
- Budge, G.E.; Hodgetts, J.; Jones, E.P.; Ostojá-Starzewski, J.C.; Hall, J.; Tomkies, V.; Semmence, N.; Brown, M.; Wakefield, M.; Stainton, K. The invasion, provenance and diversity of Vespa velutina Lepeletier (Hymenoptera: Vespidae) in Great Britain. PLoS ONE 2017, 12, e0185172. [Google Scholar] [CrossRef]
- Takahashi, J.; Okuyama, H.; Kiyoshi, T.; Takeuchi, T.; Martin, S.J. Origins of Vespa velutina hornets that recently invaded Iki Island, Japan and Jersey Island, UK. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2019, 30, 434–439. [Google Scholar] [CrossRef]
- McGlynn, T.P. The Ecology of Nest Movement in Social Insects. Annu. Rev. Entomol. 2012, 57, 291–308. [Google Scholar] [CrossRef] [Green Version]
- Klingner, R.; Richter, K.; Schmolz, E.; Keller, B. The role of moisture in the nest thermoregulation of social wasps. Naturwissenschaften 2005, 92, 427–430. [Google Scholar] [CrossRef]
- Jones, J.C.; Oldroyd, B.P. Nest Thermoregulation in Social Insects. Adv. Insect Phys. 2006, 33, 153–191. [Google Scholar]
- Feás, X. Vespa velutina Spreading from 2 Nests to 10’642 in Just 4 Years in Northern Spain. 2017. Available online: https://www.vespavelutina.co.uk/vespavelutinanews/asian-hornet-spreading-from-2-nests-to-10642-in-just-4-years-in-northern-spain (accessed on 25 April 2019).
- Feás, X. A Apicultura, unha actividade necesaria para a vida na Terra. Presented at the Academia de Ciencias Veterinarias de Galicia-ACVG, Santiago de Compostela, Spain, 23 February 2012. [Google Scholar]
- Feás, X. A Vespa Velutina en Galicia: A Necesidade de Avances No Campo Científico. 2019. Available online: https://youtu.be/7ftQcbJwdHM (accessed on 5 April 2019).
- De Haro, L.; Labadie, M.; Chanseau, P.; Cabot, C.; Blanc-Brisset, I.; Penouil, F. Medical consequences of the Asian black hornet (Vespa velutina) invasion in Southwestern France. Toxicon 2010, 55, 650–652. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, S.; Zhou, Y.; Xie, C.; Zhu, B.; Zhu, H.; Liu, S.; Wang, W.; Chen, H.; Ji, Y. Deciphering the Venomic Transcriptome of Killer-Wasp Vespa velutina. Sci. Rep. 2015, 5, 9454. [Google Scholar] [CrossRef]
- Choi, M.B.; Kim, T.G.; Kwon, O. Recent Trends in Wasp Nest Removal and Hymenoptera Stings in South Korea. J. Med. Entomol. 2018, 56, 254–260. [Google Scholar] [CrossRef]
- Gaertner, M.; Wilson, J.; Cadotte, M.; MacIvor, J.S.; Zenni, R.; Richardson, D. Non-native species in urban environments: patterns, processes, impacts and challenges. Biol. Invasions 2017, 19, 3461–3469. [Google Scholar] [CrossRef] [Green Version]
- Turchi, L.; Derijard, B. Options for the biological and physical control of Vespa velutina nigrithorax (Hym.: Vespidae) in Europe: A review. J. Appl. Entomol. 2018, 142, 553–562. [Google Scholar] [CrossRef]
- Rome, Q.; Muller, F.J.; Touret-Alby, A.; Darrouzet, E.; Perrard, A.; Villemant, C. Caste differentiation and seasonal changes in Vespa velutina (Hym.: Vespidae) colonies in its introduced range. J. Appl. Entomol. 2015, 139, 771–782. [Google Scholar] [CrossRef]
- Ueno, T. Establishment of the invasive hornet Vespa velutina (Hymenoptera: Vespidae) in Japan. Int. J. Chem. Env. Biol. Sci. 2014, 2, 220–222. [Google Scholar]
- Pérez-de-Heredia, I.; Darrouzet, E.; Goldarazena, A.; Romón, P.; Iturrondobeitia, J.C. Differentiating between gynes and workers in the invasive hornet Vespa velutina (Hymenoptera, Vespidae) in Europe. J. Hymenopt. Res. 2017, 6, 119–133. [Google Scholar] [CrossRef]
- Feás, X. The Asian Hornet (Vespa velutina nigrithorax): An exotic predator in Europe. What does the future hold? Presented at the 42nd British Beekeepers Association (BBKA) Spring Convention, Newport Shropshire, UK, 4 April 2019. [Google Scholar]
- Barthélémy, C. A provisional identification guide to the social Vespids of Hong Kong (Hymenoptera: Vespidae). 2008. Available online: https://www.academia.edu/1884577/Barthelemy_2008_A_Provisional_Identification_Guide_To_The_Social_Vespids_Of_Hong_Kong_Hymenoptera_Vespidae (accessed on 25 April 2019).
- Matsuura, M.; Yamane, S. Biology of the Vespine Wasps; Springer: Berlin, Germany, 1990; 323p. [Google Scholar]
- Robert, L.J. Nest Building. In Encyclopedia of Insects, 2nd ed.; Resh, V.H., Cardé, R.T., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 688–691. [Google Scholar]
- Oldroyd, B.P.; Pratt, S.C. Comb Architecture of the Eusocial Bees Arises from Simple Rules Used During Cell Building. Adv. Insect Physiol. 2015, 49, 101–121. [Google Scholar]
- Monceau, K.; Thiéry, D. Vespa velutina nest distribution at a local scale: An eight-year survey of the invasive honeybee predator. Insect Sci. 2017, 24, 663–674. [Google Scholar] [CrossRef]
- Asociación Galega de Apicultura, AGA. Tártago Asiático de Patas Amarelas. Available online: https://www.apiculturagalega.es/biblioteca/correo-apicola (accessed on 25 April 2019).
- Xunta de Galicia. Available online: https://mediorural.xunta.gal/es/areas/ganaderia/apicultura/vespa_velutina/ (accessed on 25 April 2019).
- Monceau, K.; Bonnard, O.; Thiéry, D. Vespa velutina: A new invasive predator of honeybees in Europe. J. Pest Sci. 2004, 87, 1–16. [Google Scholar] [CrossRef]
- Roitberg, B.D. Why pest management needs behavioral ecology and vice versa. Entomol. Res. 2007, 37, 14–18. [Google Scholar] [CrossRef]
- Kipyatkov, V.E. Life Cycles in Social Insects: Behaviour, Ecology and Evolution; St. Petersburg University Press: St. Petersburg, Russia, 2006; 156p. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
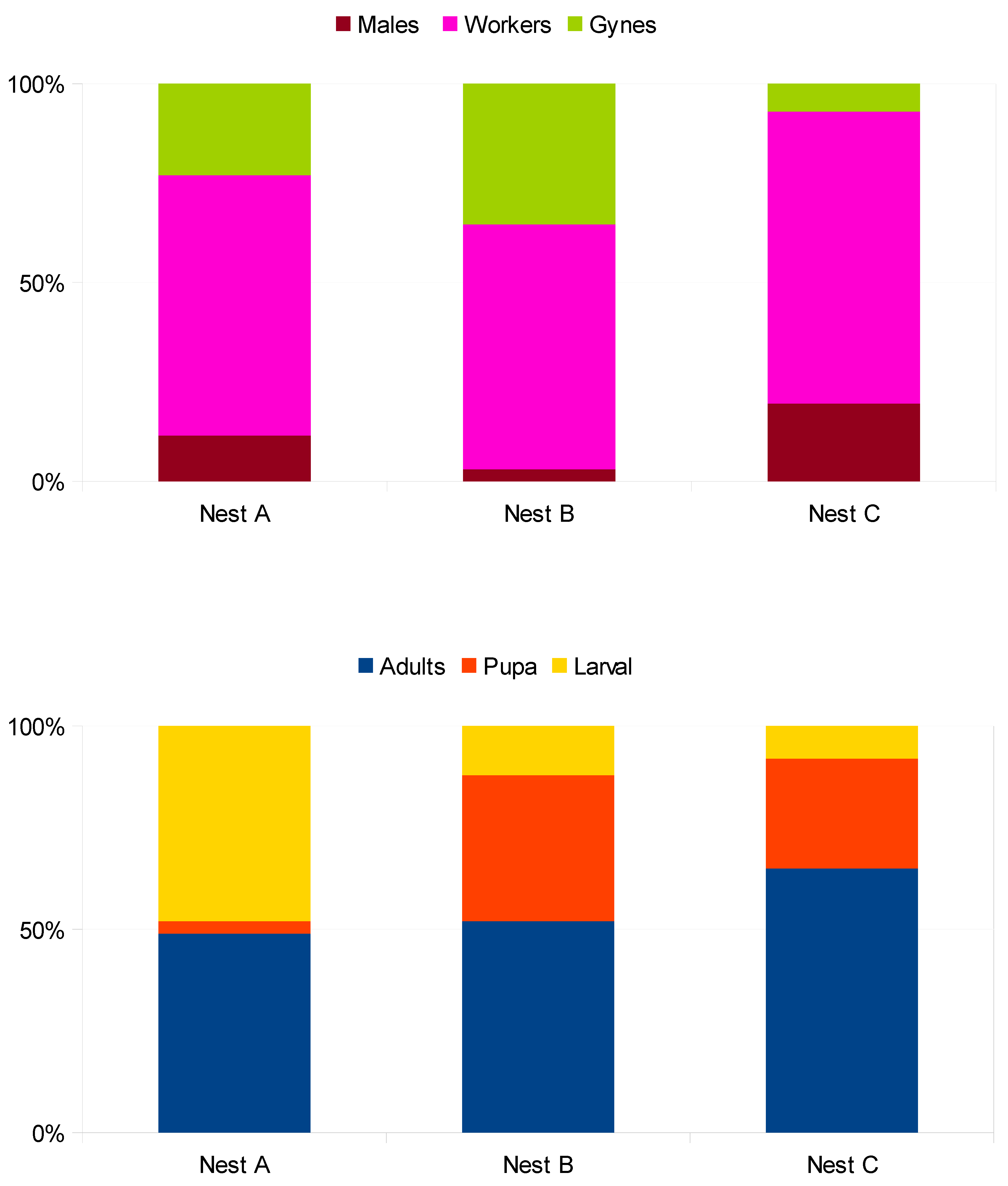{kind=link}
| Nest | LN | CN | WN | HN | VN | Tree | LMBC | DMB | NSB | DSB |
|---|---|---|---|---|---|---|---|---|---|---|
| A | 28.7 | 69.3 | 22 | 11 | 9.7 | Alnus glutinosa | 17 | 1.9 | - | - |
| B | 71.6 | 119.2 | 37.9 | 12 | 101.8 | Salix alba | 34 | 2.7 | 6 | 0.6; 0.9; 1; 1.4 and 1.5 |
| C | 54.4 | 115.6 | 36.8 | 10 | 57.0 | Salix babylonica | 2.5 | 3.7 | 4 | 1.4; 1.6; 2.5 and 2.6 |
| Nest A | Nest B | Nest C | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Comb num. | LC-max | LC-min | SC-max | NP | LC-max | LC-min | SC-max | NP | LC-max | LC-min | SC-max |
| 1 | 14.2 | 13.1 | 146.1 | 12 | 27.1 | 21.9 | 466.1 | 8 | 25.1 | 24.2 | 477.1 |
| 2 | 14.0 | 14.0 | 153.9 | 13 | 31.2 | 26.1 | 639.6 | 30 | 28.9 | 22.1 | 501.6 |
| 3 | 16.1 | 15.6 | 197.3 | 12 | 31.3 | 27.2 | 668.7 | 47 | 32.1 | 24.3 | 612.6 |
| 4 | 13.3 | 12.2 | 127.4 | 15 | 32.9 | 30.9 | 803.6 | 43 | 34.5 | 24.2 | 655.7 |
| 5 | - | - | - | 35.2 | 30.3 | 837.7 | 46 | 36.1 | 26.2 | 742.8 | |
| 6 | - | - | - | 33.1 | 29.2 | 759.1 | 43 | 33.9 | 25.8 | 686.9 | |
| 7 | - | - | - | 31.0 | 25.2 | 613.6 | 42 | 24.9 | 22.9 | 447.8 | |
| 8 | - | - | - | 29.1 | 21.2 | 484.5 | 22 | 7.9 | 6.1 | 37.8 | |
| 9 | - | - | - | 24.2 | 19.3 | 366.8 | 14 | - | - | - | |
| Vmin | 13.3 | 12.2 | 127.4 | 12 | 24.2 | 19.3 | 366.8 | 8 | 7.9 | 6.1 | 37.8 |
| Vmax | 16.1 | 15.6 | 197.3 | 15 | 35.2 | 30.9 | 837.7 | 47 | 36.1 | 26.2 | 742.8 |
| NC | 4 | 9 | 8 | ||||||||
| TSN | 624.7 | 5639.6 | 4162.5 | ||||||||
| Nest | IAPN | WCPN | Comb number | Eggs | Larval | Pupa |
|---|---|---|---|---|---|---|
| A | 87 | 176 | 1 | - | - | - |
| 2 | - | - | - | |||
| 3 | + | 22 | 2 | |||
| 4 | - | 62 | 3 | |||
| B | 1021 | 1979 | 1 | - | - | - |
| 2 | - | - | - | |||
| 3 | - | - | - | |||
| 4 | - | - | - | |||
| 5 | - | 52 | 2 | |||
| 6 | - | 48 | 42 | |||
| 7 | + | 68 | 143 | |||
| 8 | + | 48 | 285 | |||
| 9 | + | 30 | 240 | |||
| C | 430 | 662 | 1 | - | - | - |
| 2 | - | - | - | |||
| 3 | - | - | - | |||
| 4 | - | - | - | |||
| 5 | - | 12 | 76 | |||
| 6 | + | 11 | 34 | |||
| 7 | + | 27 | 72 | |||
| 8 | - | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feás Sánchez, X.; Charles, R.J. Notes on the Nest Architecture and Colony Composition in Winter of the Yellow-Legged Asian Hornet, Vespa velutina Lepeletier 1836 (Hym.: Vespidae), in Its Introduced Habitat in Galicia (NW Spain). Insects 2019, 10, 237. https://doi.org/10.3390/insects10080237
Feás Sánchez X, Charles RJ. Notes on the Nest Architecture and Colony Composition in Winter of the Yellow-Legged Asian Hornet, Vespa velutina Lepeletier 1836 (Hym.: Vespidae), in Its Introduced Habitat in Galicia (NW Spain). Insects. 2019; 10(8):237. https://doi.org/10.3390/insects10080237
Chicago/Turabian StyleFeás Sánchez, Xesús, and Rebecca Jane Charles. 2019. "Notes on the Nest Architecture and Colony Composition in Winter of the Yellow-Legged Asian Hornet, Vespa velutina Lepeletier 1836 (Hym.: Vespidae), in Its Introduced Habitat in Galicia (NW Spain)" Insects 10, no. 8: 237. https://doi.org/10.3390/insects10080237