Role of Fruit Epicuticular Waxes in Preventing Bactrocera oleae (Diptera: Tephritidae) Attachment in Different Cultivars of Olea europaea





Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Olive Fruits
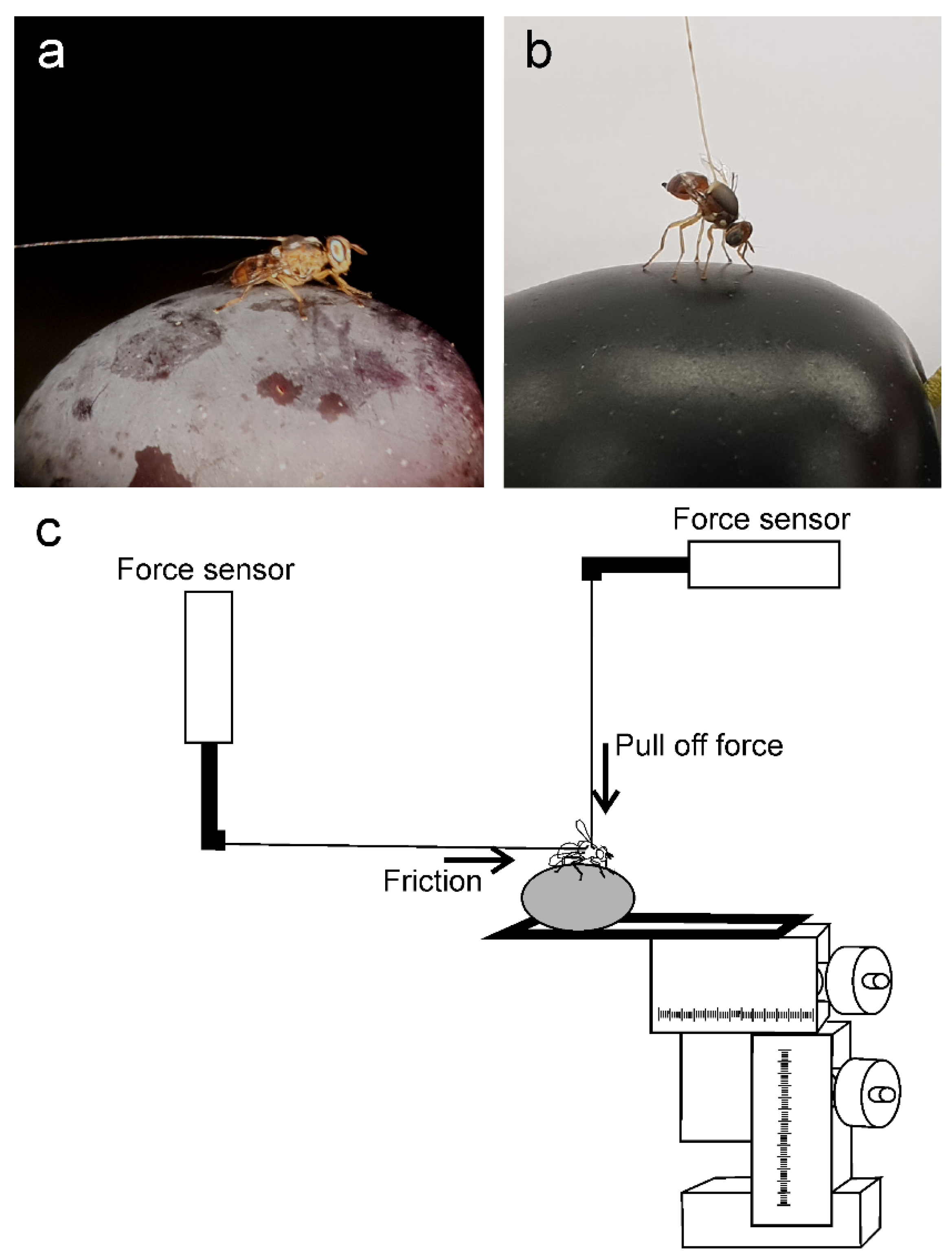
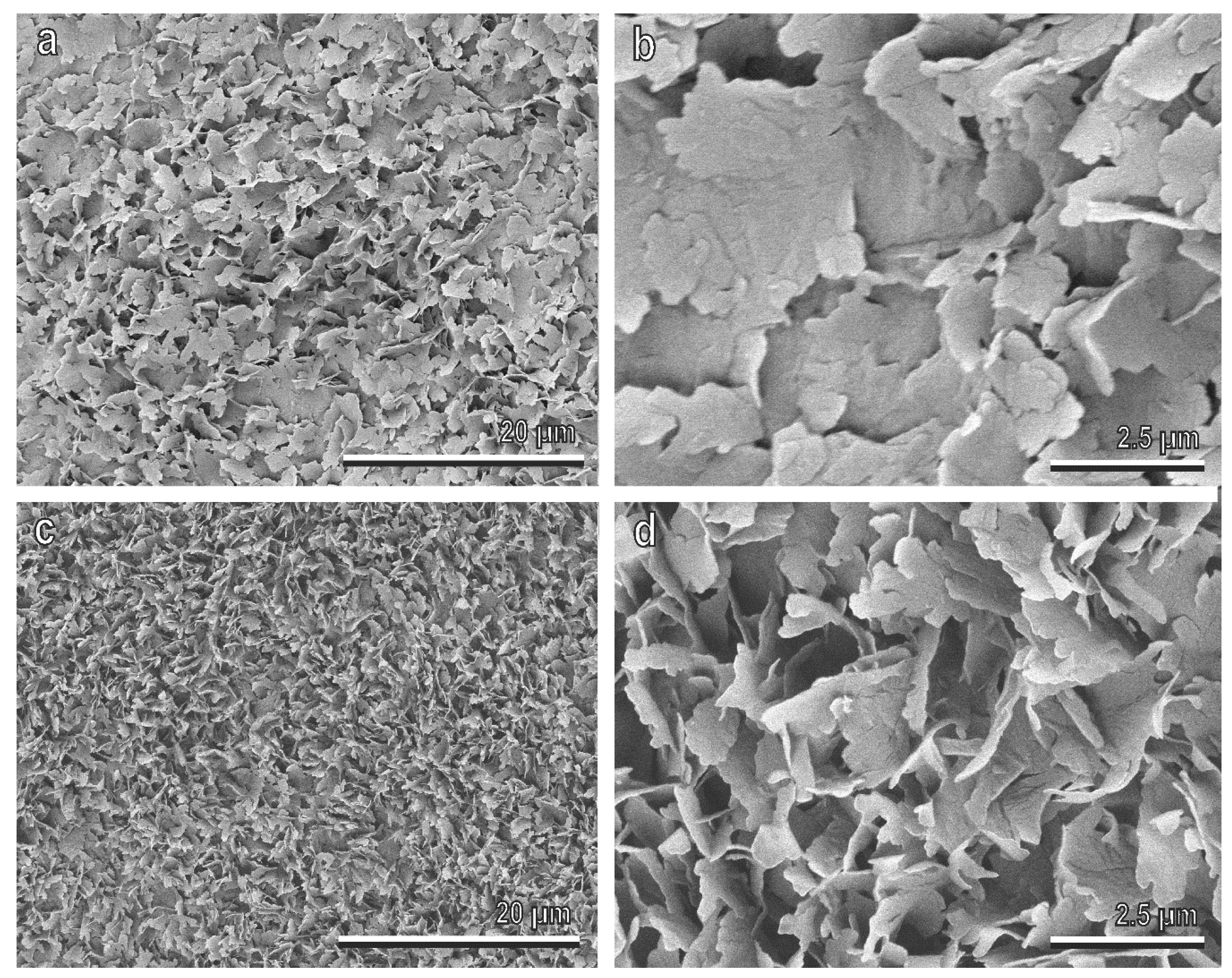
2.3. Cryo Scanning Electron Microscopy (Cryo-SEM)
2.4. Evaluation of Pulvilli Area
2.5. Characterization of Wettability of Natural (Olive Fruits) and Artificial (Hydrophilic and Hydrophobic Glass) Surfaces
2.6. Force Measurements
2.7. Statistical Analysis
3. Results
3.1. Morphology of Bactrocera oleae Attachment Organs
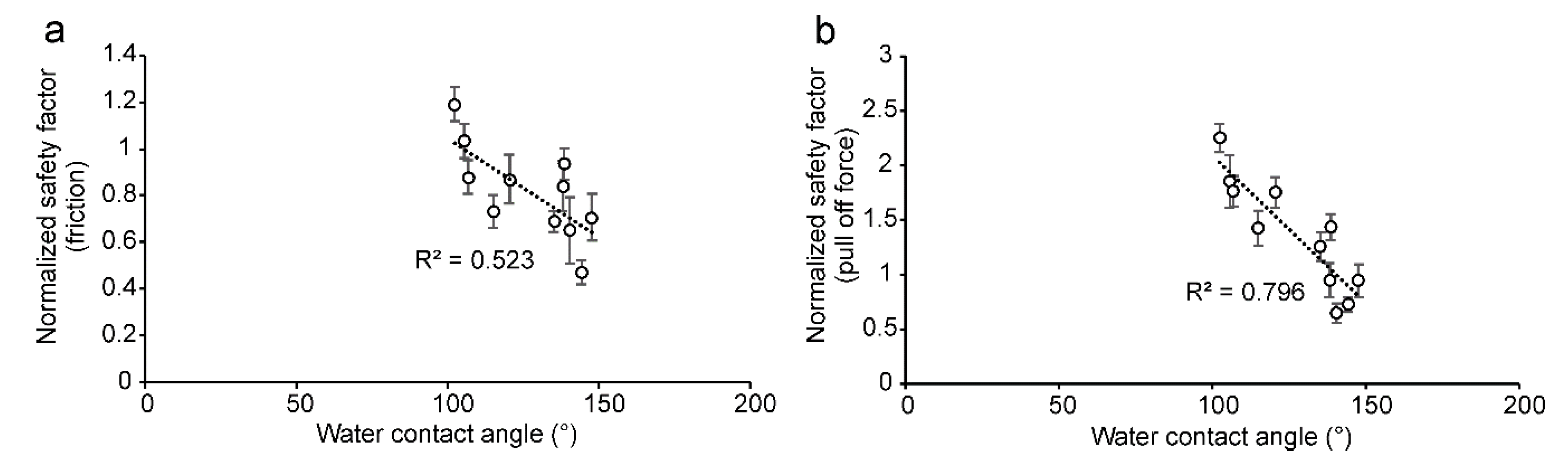
3.2. Surface Morphology and Wettability of Olive Fruits
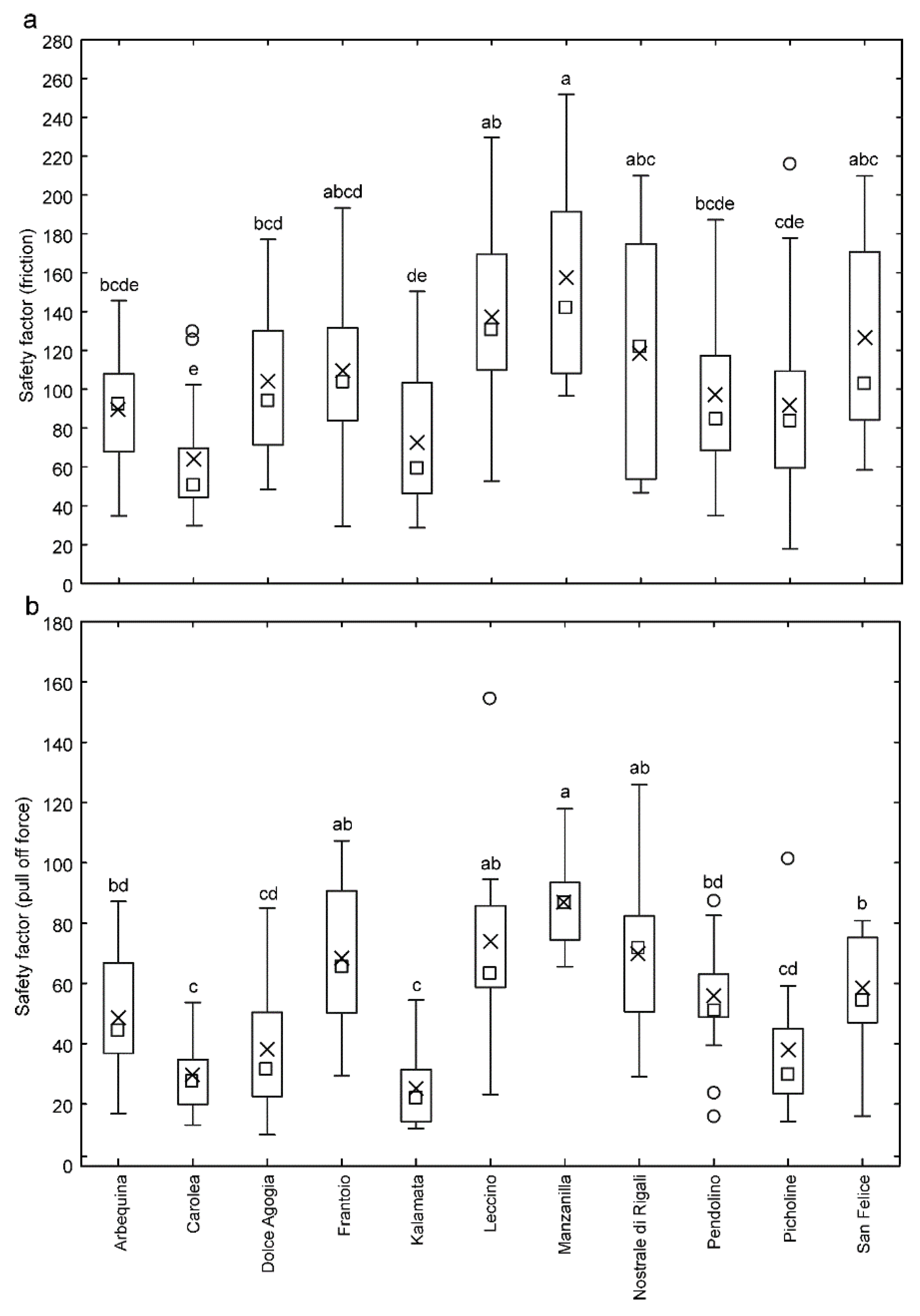
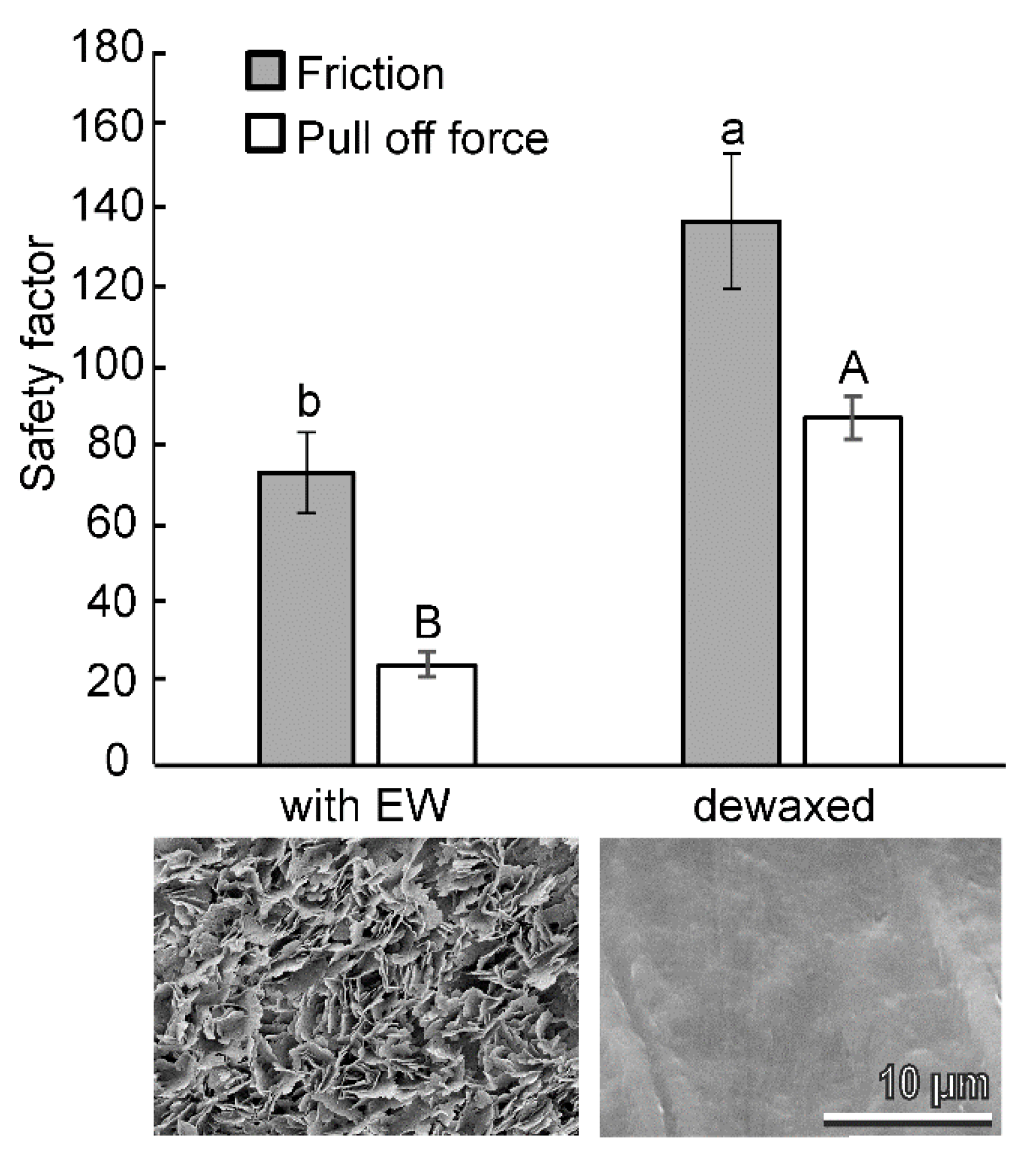
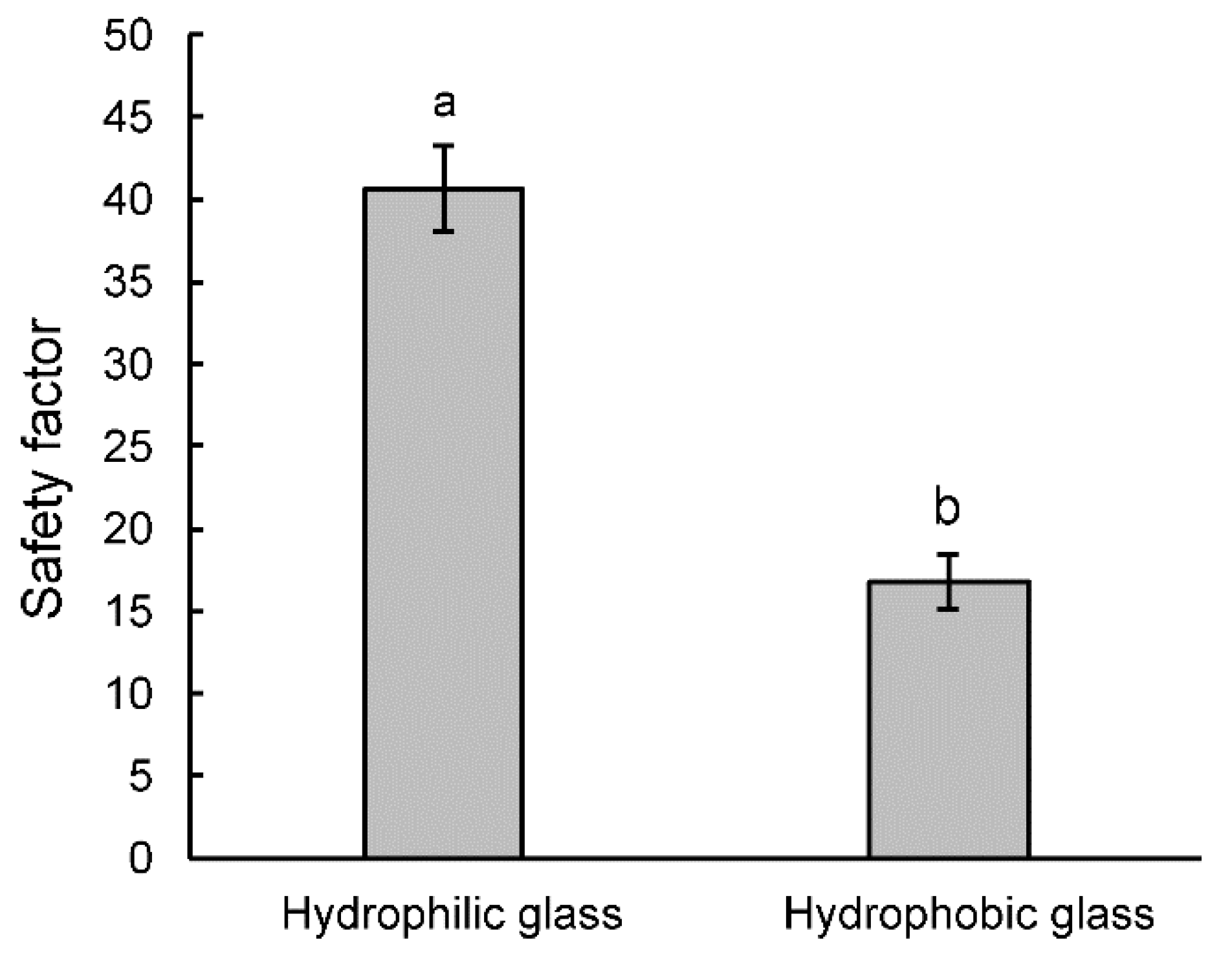
3.3. Attachment Ability of Bactrocera oleae Females to the Olive Fruit Surface of Different Cultivars and to Hydrophilic and Hydrophobic Glass
4. Discussion
4.1. EWs Effect on the Attachment Ability of Bactrocera oleae to Olive Fruit Surfaces
4.2. Different Attachment Ability of Bactrocera oleae to the Olive Fruits of Different Cultivars of Olea europaea
4.3. Attachment Ability of Bactrocera oleae and Cultivar Susceptibility to the Olive Fruit Fly
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barthlott, W.; Neinhuis, C.; Cutler, D.; Ditsch, F.; Meusel, I.; Theisen, I.; Wilhelmi, H. Classification and terminology of plant epicuticular waxes. Bot. J. Linn. Soc. 1998, 126, 237–260. [Google Scholar] [CrossRef]
- Bargel, H.; Koch, K.; Cerman, Z.; Neinhuis, C. Structure–function relationships of the plant cuticle and cuticular waxes—A smart material? Funct. Plant Biol. 2006, 33, 893–910. [Google Scholar] [CrossRef] [Green Version]
- Jetter, R.; Kunst, L.; Samuels, A.L. Composition of plant cuticular waxes. In Biology of the Plant Cuticle; Riederer, M., Müller, C., Eds.; Blackwell: Oxford, UK, 2006; pp. 145–181. [Google Scholar]
- Jeffree, C.E. The cuticle, epicuticular waxes and trichomes of plants, with reference to their structure, function and evolution. In Insects and the Plant Surface; Juniper, B., Southwood, T.R.E., Eds.; Edward Arnold Publishers: London, UK, 1986; pp. 23–63. [Google Scholar]
- Riederer, M.; Schreiber, L. Protecting against water loss: Analysis of the barrier properties of plant cuticles. J. Exp. Bot. 2001, 52, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- Barnes, J.D.; Cardoso-Vilhena, J. Interactions between electromagnetic radiation and the plant cuticle. In Plant Cuticles, an Integrated Approach; Kerstiens, G., Ed.; Bios Scientific Publisher: Oxford, UK, 1996; pp. 157–170. [Google Scholar]
- Barthlott, W.; Neinhuis, C. Purity of the sacred lotus or escape from contamination in biological surfaces. Planta 1997, 202, 1–7. [Google Scholar] [CrossRef]
- Fürstner, R.; Barthlott, W.; Neinhuis, C.; Walzel, P. Wetting and self cleaning properties of artificial superhydrophobic surfaces. Langmuir 2005, 21, 956–961. [Google Scholar] [CrossRef]
- Garcia, S.; Garcia, C.; Heinzen, H.; Moyna, P. Chemical basis of the resistance of barley seeds to pathogenic fungi. Phytochemistry 1997, 44, 415–418. [Google Scholar] [CrossRef]
- Gorb, S.N.; Gorb, E. Anti-adhesive effects of plant wax coverage on insect attachment. J. Exp. Bot. 2017, 68, 5323–5337. [Google Scholar] [CrossRef] [Green Version]
- Eigenbrode, S.D. Plant surface waxes and insect behaviour. In Plant Cuticles—An Integrated Functional Approach; Kerstiens, G., Ed.; BIOS Scientific Publishers: Oxford, UK, 1996; pp. 201–222. [Google Scholar]
- Müller, C. Plant-insect interactions on cuticular surfaces. In Biology of the Plant Cuticle; Riederer, M., Müller, C., Eds.; Blackwell: Oxford, UK, 2006; pp. 398–422. [Google Scholar]
- Daane, K.M.; Johnson, M.W. Olive Fruit Fly: Managing an Ancient Pest in Modern Times. Annu. Rev. Entomol. 2010, 55, 151–169. [Google Scholar] [CrossRef]
- Malheiro, R.; Casal, S.; Baptista, P.; Pereira, J.A. A review of Bactrocera oleae (Rossi) impact in olive products: From the tree to the table. Trends Food Sci. Technol. 2015, 44, 226–242. [Google Scholar] [CrossRef]
- Neuenschwander, P.; Michelakis, S. Olive Fruit Drop Caused by Dacus oleae (Gmel) (Dipt. Tephritidae). Z. Angew. Entomol. 1981, 91, 193–205. [Google Scholar] [CrossRef]
- Barrios, G.; Mateu, J.; Ninot i Cort, A.; Romero, A.; Vichi, S. Sensibilidad varietal del olivo a Bactrocera oleae y su incidencia en la Gestión Integrada de Plagas. Phytoma España Rev. Prof. Sanid. Veg. 2015, 268, 21–28. [Google Scholar]
- Grasso, F.; Coppola, M.; Carbone, F.; Baldoni, L.; Alagna, F.; Perrotta, G.; PeÂrez-Pulido, A.J.; Garonna, A.; Facella, P.; Daddiego, L.; et al. The transcriptional response to the olive fruit fly (Bactrocera oleae) reveals extended differences between tolerant and susceptible olive (Olea europaea L.) varieties. PLoS ONE 2017, 12, e0183050. [Google Scholar] [CrossRef] [Green Version]
- Neuenschwander, P.; Michelakis, S.; Holloway, P.; Berchtol, W. Factors affecting the susceptibility of fruits of different olive varieties to attack by Dacus oleae (Gmel.) (Dipt, Tephritidae). Z. Angew. Entomol. 1985, 100, 174–188. [Google Scholar] [CrossRef]
- Kombargi, W.S.; Michelakis, S.E.; Petrakis, C.A. Effect of olive surface waxes on oviposition by Bactrocera oleae (Diptera: Tephritidae). J. Econ. Entomol. 1998, 91, 993–998. [Google Scholar] [CrossRef]
- Rizzo, R.; Caleca, V.; Lombardo, A. Relation of fruit color, elongation, hardness, and volume to the infestation of olive cultivars by the olive fruit fly, Bactrocera oleae. Entomol. Exp. Appl. 2012, 145, 15–22. [Google Scholar] [CrossRef]
- Vichi, S.; Cortés-Francisco, N.; Caixach, J.; Barrios, G.; Mateu, J.; Ninot, A. Epicuticular wax in developing olives (Olea europaea) is highly dependent upon cultivar and fruit ripeness. J. Agric. Food Chem. 2016, 64, 5985–5994. [Google Scholar] [CrossRef]
- Gorb, E.V.; Gorb, S.N. Functional surfaces in the pitcher of the carnivorous plant Nepenthes alata: A cryo-SEM approach. In Functional Surfaces in Biology—Adhesion Related Phenomena; Gorb, S.N., Ed.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2009; Volume 2, pp. 205–238. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Gorb, E.V.; Hosoda, N.; Miksch, C.; Gorb, S.N. Slippery pores: Anti-adhesive effect of nanoporous substrates on the beetle attachment system. J. R. Soc. Interface 2010, 7, 1571–1579. [Google Scholar] [CrossRef] [Green Version]
- StatSoft Inc. Statistica (Data Analysis Software System); Version 6; StatSoft Italia S.R.L: Vigonza (PD), Italy, 2001. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research; W.H. Freeman and Company: New York, NY, USA, 1998; p. 887. [Google Scholar]
- Anstey, T.H.; Moore, J.F. Inheritance of glossy foliage and cream petals in Green Sprouting Broccoli. J. Hered. 1954, 45, 39–41. [Google Scholar] [CrossRef]
- Edwards, P.B. Do waxes of juvenile Eucalyptus leaves provide protection from grazing insects? Aust. J. Ecol. 1982, 7, 347–352. [Google Scholar] [CrossRef]
- Bodnaryk, R.P. Leaf epicuticular wax, an antixenotic factor in Brassicaceae that affects the rate and pattern of feeding in flea beetles, Phyllotreta cruciferae (Goeze). Can. J. Plant Sci. 1992, 72, 1295–1303. [Google Scholar] [CrossRef]
- Brennan, E.B.; Weinbaum, S.A. Effect of epicuticular wax on adhesion of psyllids to glaucous juvenile and glossy adult leaves of Eucalyptus globulus Labillardière. Aust. J. Entomol. 2001, 40, 270–277. [Google Scholar] [CrossRef]
- Chang, G.C.; Neufeld, J.; Eigenbrode, S.D. Leaf surface wax and plant morphology of peas influence insect density. Entomol. Exp. Appl. 2006, 119, 197–205. [Google Scholar] [CrossRef]
- Salerno, G.; Rebora, M.; Piersanti, S.; Gorb, E.V.; Gorb, S.N. Mechanical ecology of fruit-insect interaction in the adult Mediterranean fruit fly Ceratitis capitata (Diptera: Tephritidae). Zoology 2020, 139, 1257482. [Google Scholar] [CrossRef] [PubMed]
- Liquido, N.J.; Shinoda, L.A.; Cunningham, R.T. Host plants of the Mediterranean fruit fly (Diptera: Tephritidae): An annotated world review. Ann. Entomol. Soc. Am. 1991, 77, 1–52. [Google Scholar]
- White, I.M.; Elson-Harris, M.M. Fruit Flies of Economic Significance: Their Identification and Bionomics; CAB International: Wallingford, CT, USA; Oxon, UK, 1992. [Google Scholar]
- Bauchhenß, E.; Renner, M. Pulvillus of Calliphora erythrocephala Meig (Diptera; Calliphoridae). Int. J. Insect Morphol. Embryol. 1977, 6, 225–227. [Google Scholar] [CrossRef]
- Bauchhenß, E. Die Pulvillen von Calliphora erythrocephala Meig. (Diptera, Brachycera) als Adhäsionsorgane. Zoomorphologie 1979, 93, 99–123. [Google Scholar] [CrossRef]
- Walker, G.; Yule, A.B.; Ratcliffe, J. The adhesive organ of the blowfly, Calliphora vomitoria: A functional approach (Diptera: Calliphoridae). J. Zool. 1985, 205, 297–307. [Google Scholar] [CrossRef]
- Gorb, S.N. The design of the fly adhesive pad: Distal tenent setae are adapted to the delivery of an adhesive secretion. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1998, 265, 747–752. [Google Scholar] [CrossRef] [Green Version]
- Niederegger, S.; Gorb, S.N.; Jiao, Y. Contact behaviour of tenent setae in attachment pads of the blowfly Calliphora vicina (Diptera, Calliphoridae). J. Comp. Physiol. A 2002, 187, 961–970. [Google Scholar] [CrossRef]
- Gorb, S.N.; Schuppert, J.; Walther, P.; Schwarz, H. Contact behaviour of setal tips in the hairy attachment system of the fly Calliphora vicina (Diptera, Calliphoridae): A cryo-SEM approach. Zoology 2012, 115, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Gorb, S.N.; Gorb, E.V.; Kastner, V. Scale effects on the attachment pads and friction forces in syrphid flies (Diptera, Syrphidae). J. Exp. Biol. 2001, 204, 1421–1431. [Google Scholar] [PubMed]
- Gorb, E.V.; Gorb, S.N. Attachment ability of the beetle Chrysolina fastuosa on various plant surfaces. Entomol. Exp. Appl. 2002, 105, 13–28. [Google Scholar] [CrossRef]
- Friedemann, K.; Kunert, G.; Gorb, E.; Gorb, S.N.; Beutel, R.G. Attachment forces of pea aphids (Acyrthosiphon pisum) on different legume species. Ecol. Entomol. 2015, 40, 732–740. [Google Scholar] [CrossRef]
- Salerno, G.; Rebora, M.; Gorb, E.; Gorb, S. Attachment ability of the polyphagous bug Nezara viridula (Heteroptera: Pentatomidae) to different host plant surfaces. Sci. Rep. 2018, 8, 10975. [Google Scholar] [CrossRef] [Green Version]
- Gorb, E.; Haas, K.; Henrich, A.; Enders, S.; Barbakadze, N.; Gorb, S. Composite structure of the crystalline epicuticular wax layer of the slippery zone in the pitchers of the carnivorous plant Nepenthes alata and its effect on insect attachment. J. Exp. Biol. 2005, 208, 4651–4662. [Google Scholar] [CrossRef] [Green Version]
- Gorb, E.V.; Gorb, S.N. Do plant waxes make insect attachment structures dirty? Experimental evidence for the contamination hypothesis. In Ecology and Biomechanics: A Mechanical Approach to the Ecology of Animals and Plants; Herrel, A., Speck, T., Rowe, N.P., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 147–162. [Google Scholar]
- Scholz, I.; Bückins, L.; Dolge, L.; Erlinghagen, T.; Weth, A.; Hischen, F.; Mayer, J.; Hoffmann, S.; Riederer, M.; Riedel, M.; et al. Slippery surfaces of pitcher plants: Nepenthes wax crystals minimize insect attachment via microscopic surface roughness. J. Exp. Biol. 2010, 213, 1115–1125. [Google Scholar] [CrossRef] [Green Version]
- Gorb, S.N. Attachment Devices of Insect Cuticle; Kluwer Academic: Dordrecht, The Netherlands, 2001. [Google Scholar]
- Peressadko, A.; Gorb, S.N. Surface profile and friction force generated by insects. In First International Industrial Conference Bionik 2004; Boblan, I., Bannasch, R., Eds.; VDI Verlag: Düsseldorf, Germany, 2004; pp. 257–263. [Google Scholar]
- Voigt, D.; Schuppert, J.M.; Dattinger, S.; Gorb, S.N. Sexual dimorphism in the attachment ability of the Colorado potato beetle Leptinotarsa decemlineata (Coleoptera: Chrysomelidae) to rough substrates. J. Insect. Physiol. 2008, 54, 765–776. [Google Scholar] [CrossRef]
- Wolff, J.O.; Gorb, S.N. Surface roughness effects on attachment ability of the spider Philodromus dispar (Araneae, Philodromidae). J. Exp. Biol. 2012, 215, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Robinson, A.; Steiner, U.; Federle, W. Insect adhesion on rough surfaces: Analysis of adhesive contact of smooth and hairy pads on transparent microstructured substrates. J. R. Soc. Interface 2014, 11, 20140499. [Google Scholar] [CrossRef] [Green Version]
- Zurek, D.B.; Gorb, S.N.; Voigt, D. Changes in tarsal morphology and attachment ability to rough surfaces during ontogenesis in the beetle Gastrophysa viridula (Coleoptera, Chrysomelidae). Arthropod Struct. Dev. 2017, 46, 130–137. [Google Scholar] [CrossRef]
- Kovalev, A.; Filippov, A.E.; Gorb, S.N. Critical roughness in animal hairy adhesive pads: A numerical modeling approach. Bioinspiration Biomim. 2018, 13, 66004. [Google Scholar] [CrossRef] [PubMed]
- Salerno, G.; Rebora, M.; Gorb, E.V.; Kovalev, A.; Gorb, S.N. Attachment ability of the southern green stink bug Nezara viridula (Heteroptera: Pentatomidae). J. Comp. Physiol. A 2017, 203, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lüken, D.; Voigt, D.; Gorb, S.N.; Zebitz, C.P.W. Die Tarsenmorphologie und die Haftfähigkeit des Schwarzen Batatenkäfers Cylas puncticollis (Boheman) auf glatten Oberflächen mit unterschiedlichen physiko-chemischen Eigenschaften. Mitt Dtsch. Ges Allg. Angew. Entomol. 2009, 17, 109–113. [Google Scholar]
- Gorb, E.V.; Gorb, S.N. Attachment ability of females and males of the ladybird beetle Cryptolaemus montrouzieri to different artificial surfaces. J. Insect Physiol. 2020, 121, 104011. [Google Scholar] [CrossRef]
- Hosoda, N.; Gorb, S.N. Underwater locomotion in a terrestrial beetle: Combination of surface de-wetting and capillary forces. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2012, 279, 4236–4242. [Google Scholar] [CrossRef] [Green Version]
- England, M.W.; Sato, T.; Yagihashi, M.; Hozumi, A.; Gorb, S.N.; Gorb, E.V. Surface roughness rather than surface chemistry essentially affects insect adhesion. Beilstein J. Nanotechnol. 2016, 7, 1471–1479. [Google Scholar] [CrossRef] [Green Version]
- Grohmann, C.; Blankenstein, A.; Koops, S.; Gorb, S.N. Attachment of Galerucella nymphaeae (Coleoptera, Chrysomelidae) to surfaces with different surface energy. J. Exp. Biol. 2014, 217, 4213–4220. [Google Scholar] [CrossRef] [Green Version]
- Lanza, B.; Di Serio, M.G. SEM characterization of olive (Olea europaea L.) fruit epicuticular waxes and epicarp. Sci. Hortic. 2015, 191, 49–56. [Google Scholar] [CrossRef]
- Bianchi, G.; Murelli, C.; Vlahov, G. Surface waxes from olive fruits. Phytochemistry 1992, 31, 3503–3506. [Google Scholar] [CrossRef]
- Bianchi, G.; Pozzi, N.; Vlahov, G. Pentacyclic triterpene acids in olives. Phytochemistry 1994, 37, 205–207. [Google Scholar] [CrossRef]
- Guinda, A.; Rada, M.; Delgado, T.; Gutiérrez-Adánez, P.; Castellano, J.M. Pentacylic triterpenoids from olive fruit and leaf. J. Agric. Food Chem. 2010, 58, 9685–9691. [Google Scholar] [CrossRef] [PubMed]
- Vlahov, G.; Rinaldi, G.; Del Re, P.; Giuliani, A.A. 13C nuclear magnetic resonance spectroscopy for determining the different components of epicuticular waxes of olive fruit (Olea europea) Dritta cultivar. Anal. Chim. Acta 2008, 624, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Vichi, S.; Cortés-Francisco, N.; Romero, A.; Caixach, J. Direct chemical profiling of olive (Olea europaea) fruit epicuticular waxes by direct electrospray-Ultrahigh resolution mass spectrometry. J. Mass Spectrom. 2015, 50, 558–566. [Google Scholar] [CrossRef]
- Diarte, C.; Lai, P.-H.; Huang, H.; Romero, A.; Casero, T.; Gatius, F.; Graell, J.; Medina, V.; East, A.; Riederer, M.; et al. Insights Into Olive Fruit Surface Functions: A Comparison of Cuticular Composition, Water Permeability, and Surface Topography in Nine Cultivars During Maturation. Front. Plant Sci. 2019, 10, 1484. [Google Scholar] [CrossRef]
- Gorb, E.; Voigt, D.; Eigenbrode, S.D.; Gorb, S. Attachment force of the beetle Cryptolaemus montrouzieri (Coleoptera, Coccinellidae) on leaflet surfaces of mutants of the pea Pisum sativum (Fabaceae) with regular and reduced wax coverage. Arthropod–Plant Interact. 2008, 2, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Gorb, E.; Böhm, S.; Jacky, N.; Maier, L.P.; Dening, K.; Pechook, S.; Pokroy, B.; Gorb, S. Insect attachment on crystalline bioinspired wax surfaces formed by alkanes of varying chain lengths. Beilstein J. Nanotechnol. 2014, 5, 1031–1041. [Google Scholar] [CrossRef] [Green Version]
- Corrado, G.; Garonna, A.; Cabanàs, C.G.-L.; Gregoriou, M.; Martelli, G.P.; Mathiopoulos, K.D.; Saponari, J.M.-B.M.; Tsoumani, K.T.; Rao, R. Host Response to Biotic Stresses. In The Olive Tree Genome; Rugini, E., Baldoni, L., Muleo, R., Sebastiani, L., Eds.; Springer International Publishing: Cham, Germany, 2016; pp. 75–98. [Google Scholar]
- Bartolini, G. Olive Germplasm (Olea europaea L.). Istituto per la Valorizzazione del Legno e delle Specie Arboree (IVALSA), Trees and Timber Institute. Available online: www.oleadb.it/olivodb.html (accessed on 20 January 2020). [CrossRef]
- Yeats, T.H.; Rose, J.K.C. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, P.; Nguyen, N.; Hykkerud, A.L.; Häggman, H.; Martinussen, I.; Jaakola, L.; Karppinen, K. Developmental and environmental regulation of cuticular wax biosynthesis in fleshy fruits. Front. Plant Sci. 2019, 10, 431. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Legs | Pulvillus Area (µm2) | n |
|---|---|---|
| ♀ * | ||
| foreleg | 10440 ± 234.0 b | 10 |
| midleg | 12644 ± 353.4 a | 10 |
| hindleg | 14118 ± 340.5 a | 10 |
| ♂ | ||
| foreleg | 7437 ± 236.9 A | 10 |
| midleg | 8879 ± 292.8 B | 10 |
| hindleg | 9512 ± 284.5 B | 10 |
| Cultivars | Contact Angle (°) | n |
|---|---|---|
| Arbequina | 135.18 ± 5.79 abc | 10 |
| Carolea | 144.18 ± 3.07 a | 11 |
| Dolce Agogia | 138.16 ± 5.59 ab | 9 |
| Frantoio | 120.40 ± 4.41 bd | 10 |
| Kalamata | 140.48 ± 3.50 ab | 10 |
| Leccino | 105.62 ± 5.95 d | 11 |
| Manzanilla | 102.30 ± 3.72 d | 10 |
| Nostrale | 106.88 ± 3.67 d | 8 |
| Pendolino | 115.02 ± 5.05 cd | 8 |
| Picholine | 147.62 ± 4.72 a | 10 |
| San Felice | 138.59 ± 3.92 ab | 9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rebora, M.; Salerno, G.; Piersanti, S.; Gorb, E.; Gorb, S. Role of Fruit Epicuticular Waxes in Preventing Bactrocera oleae (Diptera: Tephritidae) Attachment in Different Cultivars of Olea europaea. Insects 2020, 11, 189. https://doi.org/10.3390/insects11030189
Rebora M, Salerno G, Piersanti S, Gorb E, Gorb S. Role of Fruit Epicuticular Waxes in Preventing Bactrocera oleae (Diptera: Tephritidae) Attachment in Different Cultivars of Olea europaea. Insects. 2020; 11(3):189. https://doi.org/10.3390/insects11030189
Chicago/Turabian StyleRebora, Manuela, Gianandrea Salerno, Silvana Piersanti, Elena Gorb, and Stanislav Gorb. 2020. "Role of Fruit Epicuticular Waxes in Preventing Bactrocera oleae (Diptera: Tephritidae) Attachment in Different Cultivars of Olea europaea" Insects 11, no. 3: 189. https://doi.org/10.3390/insects11030189
APA StyleRebora, M., Salerno, G., Piersanti, S., Gorb, E., & Gorb, S. (2020). Role of Fruit Epicuticular Waxes in Preventing Bactrocera oleae (Diptera: Tephritidae) Attachment in Different Cultivars of Olea europaea. Insects, 11(3), 189. https://doi.org/10.3390/insects11030189