An Acylsucrose-Producing Tomato Line Derived from the Wild Species Solanum pimpinellifolium Decreases Fitness of the Whitefly Trialeurodes vaporariorum


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tomato Plants and Whitefly Population
2.2. Trichome Observation and Acylsucrose Accumulation Quantification
2.3. T. vaporariorum Settling Preference
2.4. Oviposition of T. vaporariorum
2.5. Survival of Immature T. vaporariorum
3. Results
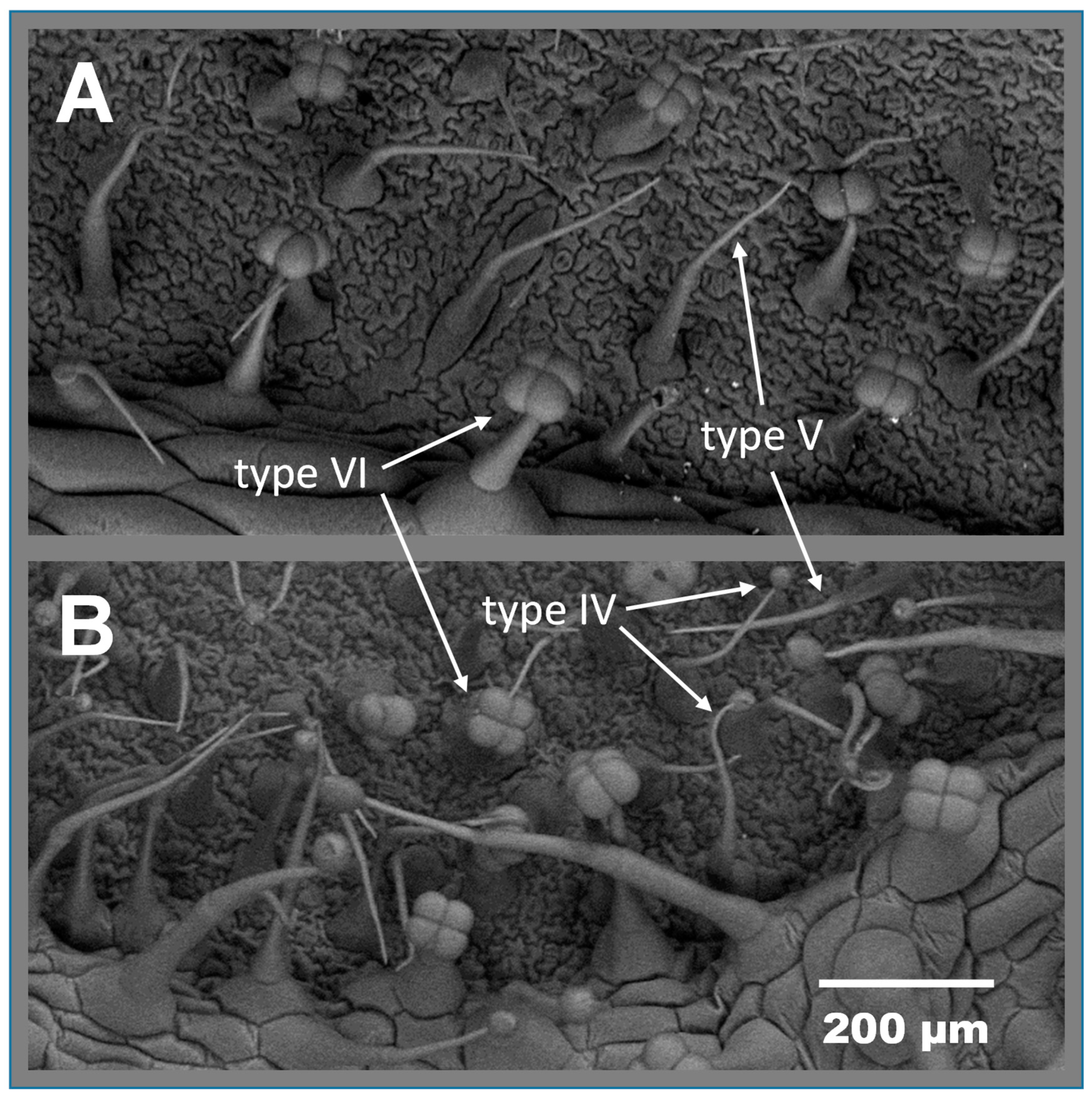
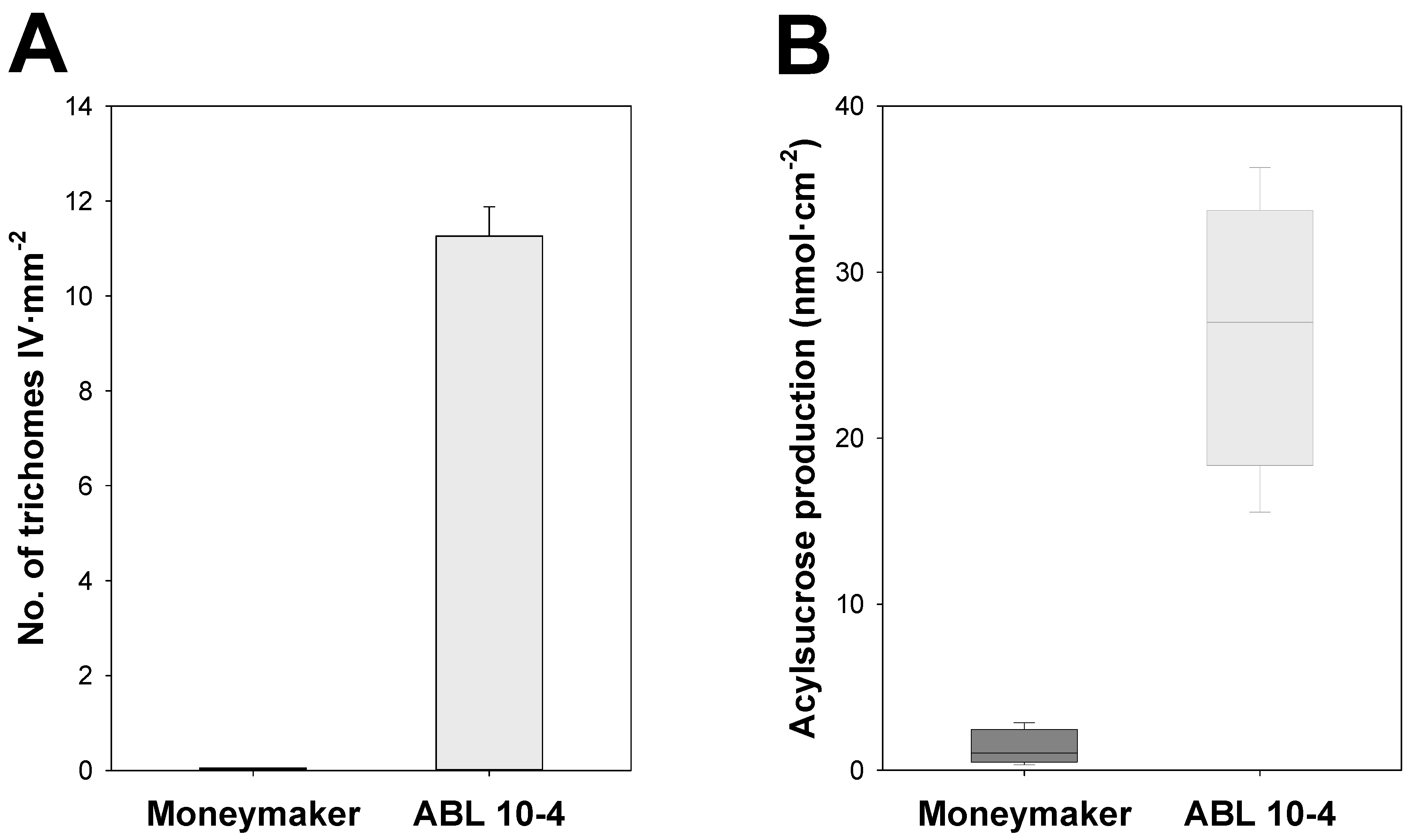
3.1. ABL 10-4 Accumulates Type IV Glandular Trichomes and Acylsucroses in Leaves
3.2. T. vaporariorum Repellence in the Genotype with Type IV Glandular Trichomes
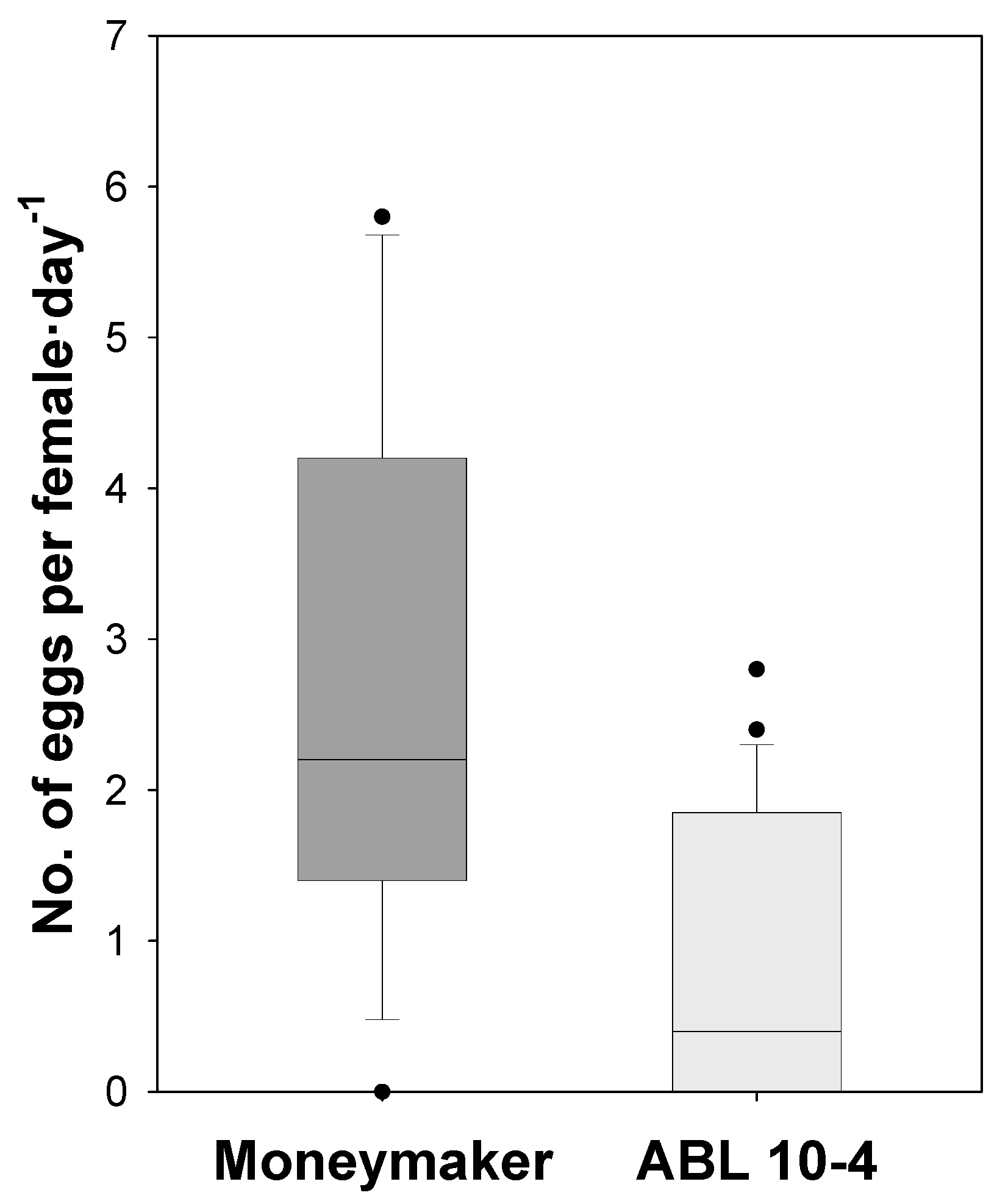
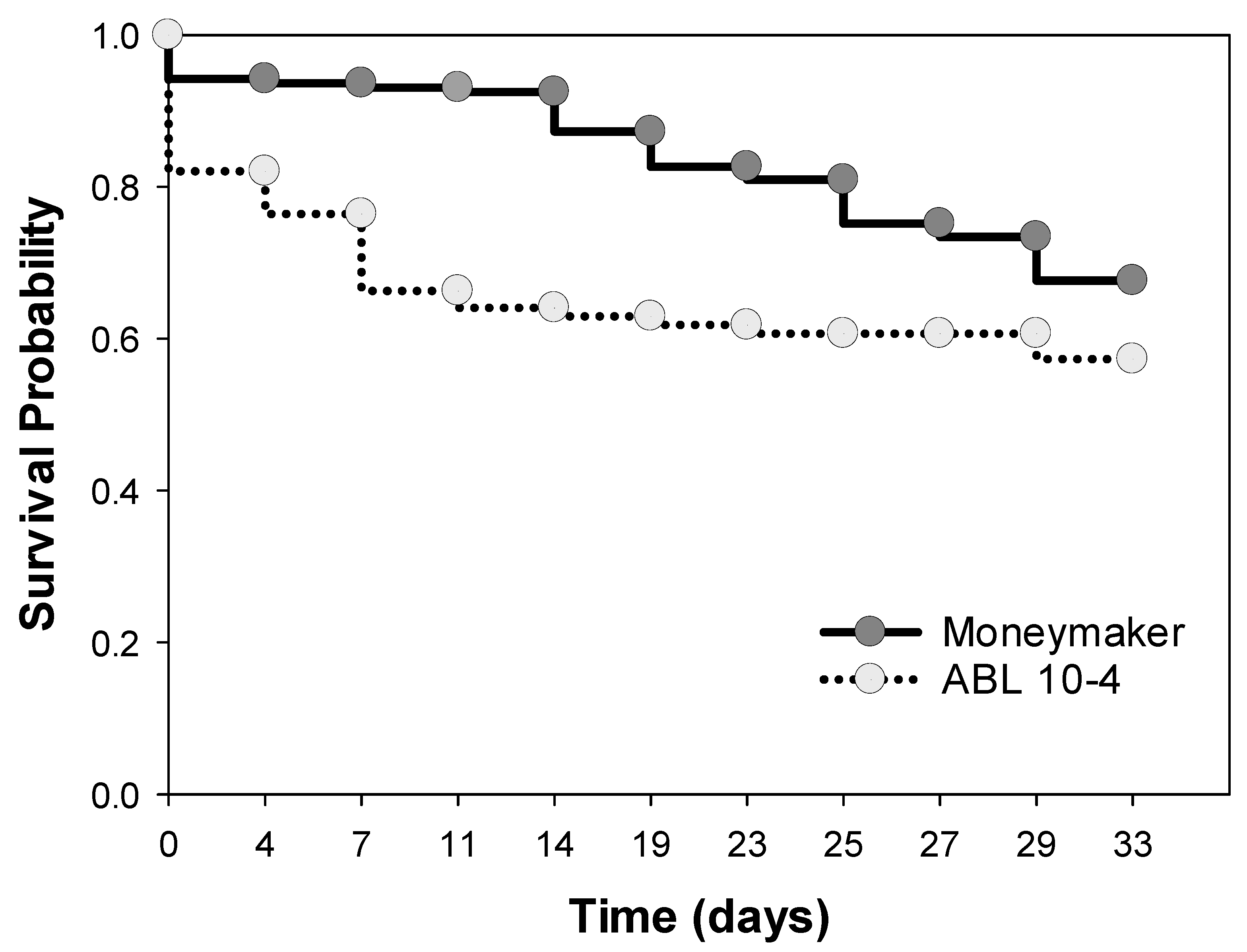
3.3. Negative Effect of ABL 10-4 on Oviposition and Survival of T. vaporariorum
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wainaina, J.M.; De Barro, P.; Kubatko, L.; Kehoe, M.A.; Harvey, J.; Karanja, D.; Boykin, L.M. Global phylogenetic relationships, population structure and gene flow estimation of Trialeurodes vaporariorum (Greenhouse whitefly). Bull. Entomol. Res. 2017, 108, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Van Lenteren, J.C. Biological control in protected crops: Where do we go? Pestic. Sci. 1992, 36, 321–327. [Google Scholar] [CrossRef]
- Gorman, K.; Hewitt, F.; Denholm, I.; Devine, G.J. New developments in insecticide resistance in the glasshouse whitefly (Trialeurodes vaporariorum) and the two-spotted spider mite (Tetranychus urticae) in the UK. Pest Manag. Sci. 2002, 58, 123–130. [Google Scholar] [CrossRef]
- Navas-Castillo, J.; Fiallo-Olive, E.; Sanchez-Campos, S. Emerging virus diseases transmitted by whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.K.; Czosnek, H. Whitefly transmission of plant viruses. Adv. Bot. Res. 2002, 36, 65–100. [Google Scholar]
- Navas-Castillo, J.; Lopez-Moya, J.J.; Aranda, M.A. Whitefly-transmitted RNA viruses that affect intensive vegetable production. Ann. Appl. Biol. 2014, 165, 155–171. [Google Scholar] [CrossRef]
- Wintermantel, W.M. Emergence of Greenhouse Whitefly (Trialeurodes vaporariorum) Transmitted Criniviruses as threats to Vegetable and Fruit Production in North America; APSnet Feature Story: St. Paul, MN, USA, 2004. [Google Scholar]
- Fiallo-Olive, E.; Pan, L.L.; Liu, S.S.; Navas-Castillo, J. Transmission of begomoviruses and other whitefly-borne viruses: Dependence on the vector species. Phytopathology 2020, 110, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Fiallo-Olive, E.; Navas-Castillo, J. Tomato chlorosis virus, an emergent plant virus still expanding its geographical and host ranges. Mol. Plant Pathol. 2019, 20, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.W.; Caprio, L.C.; Coughlin, J.A.; Tabashnik, B.E.; Rosenheim, J.A.; Welter, S.C. Effect of Trialeurodes vaporariorum (Homoptera: Aleyrodidae) on yield of fresh market tomatoes. J. Econ. Entomol. 1992, 85, 2370–2376. [Google Scholar] [CrossRef]
- Mansilla-Córdova, P.J.; Bampi, D.; Rondinel-Mendoza, N.V.; Melo, P.C.T.; Lourenção, A.L.; Rezende, J.A.M. Screening tomato genotypes for resistance and tolerance to Tomato chlorosis virus. Plant Pathol. 2018, 67, 1231–1237. [Google Scholar] [CrossRef]
- Omer, A.D.; Johnson, M.W.; Tabashnik, B.E.; Ullman, D.E. Association between insecticide use and greenhouse whitefly (Trialeurodes vaporariorum Westwood) resistance to insecticides in Hawaii. Pestic. Sci. 1993, 37, 253–259. [Google Scholar] [CrossRef]
- Dittrich, V.; Uk, S.; Ernst, G.H. Chemical control and insecticide resistance of whiteflies. In Whiteflies: Their Bionomics, Pest Status and Management; Intercept Press: Andover, Hants, UK, 1990; pp. 263–285. [Google Scholar]
- van Lenteren, J.C.; van Roermund, H.J.W.; Sütterlin, S. Biological control of greenhouse whitefly (Trialeurodes vaporariorum) with the parasitoid Encarsia formosa: How does It work? Biol. Control 1996, 6, 1–10. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; Maranhao, E.A.A.; Valverde-Garcia, P.; Santiago-Alvarez, C. Selection of Beauveria bassiana isolates for control of the whiteflies Bemisia tabaci and Trialeurodes vaporariorum on the basis of their virulence, thermal requirements, and toxicogenic activity. Biol. Control 2006, 36, 274–287. [Google Scholar] [CrossRef]
- Meekes, E.T.M.; Fransen, J.J.; van Lenteren, J.C. Pathogenicity of Aschersonia spp. against whiteflies Bemisia argentifolii and Trialeurodes vaporariorum. J. Invertebr. Pathol. 2002, 81, 1–11. [Google Scholar] [CrossRef]
- Byrne, D.N.; Bellows, T.S.J. Whitefly Biology. Annu. Rev. Entomol. 1991, 36, 431–457. [Google Scholar] [CrossRef]
- Coombe, P.E. Visual behaviour of the greenhouse whitefly, Trialeurodes vaporariorum. Physiol. Entomol. 1982, 7, 243–251. [Google Scholar] [CrossRef]
- Broekgaarden, C.; Snoeren, T.A.L.; Dicke, M.; Vosman, B. Exploiting natural variation to identify insect-resistance genes. Plant Biotechnol. J. 2011, 9, 819–825. [Google Scholar] [CrossRef]
- Walling, L.L. Adaptive defense responses to pathogens and insects. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2009; Volume 51, pp. 551–612. [Google Scholar]
- Fraser, R.S.S. Integrated pest and disease management in protected crops. Outlook Agric. 1992, 21, 169–175. [Google Scholar] [CrossRef]
- Fereres, A.; Moreno, A. Integrated Control Measures against Viruses and Their Vectors; Caister Academic Press: Wymondham, UK, 2011; pp. 237–261. [Google Scholar]
- Nombela, G.; Beitia, F.; Muniz, M. Variation in tomato host response to Bemisia tabaci (Hemiptera: Aleyrodidae) in relation to acyl sugar content and presence of the nematode and potato aphid resistance gene Mi. Bull. Entomol. Res. 2000, 90, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Bas, N.; Mollema, C.; Lindhout, P. Resistance in Lycopersicon hirsutum f. glabratum to the greenhouse whitefly (Trialeurodes vaporariorum) increases with plant age. Euphytica 1992, 64, 189–195. [Google Scholar] [CrossRef]
- Rodriguez-Lopez, M.J.; Garzo, E.; Bonani, J.P.; Fereres, A.; Fernandez-Muñoz, R.; Moriones, E. Whitefly resistance traits derived from the wild tomato Solanum pimpinellifolium affect the preference and feeding behavior of Bemisia tabaci and reduce the spread of Tomato yellow leaf curl virus. Phytopathology 2011, 101, 1191–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Q.; Peng, Z.; Tong, H.; Yang, F.; Xing, G.; Wang, L.; Zheng, J.; Zhang, Y.; Su, Q. Tomato plant flavonoids increase whitefly resistance and reduce spread of tomato yellow leaf curl virus. J. Econ. Entomol. 2019, 112, 2790–2796. [Google Scholar] [CrossRef]
- Walling, L.L. The myriad plant responses to herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [CrossRef] [PubMed]
- Bleeker, P.M.; Mirabella, R.; Diergaarde, P.J.; VanDoorn, A.; Tissier, A.; Kant, M.R.; Prins, M.; de Vos, M.; Haring, M.A.; Schuurink, R.C. Improved herbivore resistance in cultivated tomato with the sesquiterpene biosynthetic pathway from a wild relative. Proc. Natl. Acad. Sci. USA 2012, 109, 20124–20129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanDoorn, A.; de Vos, M. Resistance to sap-sucking insects in modern-day agriculture. Front. Plant Sci. 2013, 4, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glas, J.J.; Schimmel, B.C.J.; Alba, J.M.; Escobar-Bravo, R.; Schuurink, R.C.; Kant, M.R. Plant glandular trichomes as targets for breeding or engineering of resistance to herbivores. Int. J. Mol. Sci. 2012, 13, 17077–17103. [Google Scholar] [CrossRef] [Green Version]
- Vosman, B.; van’t Westende, W.P.C.; Henken, B.; van Eekelen, H.D.L.M.; de Vos, R.C.H.; Voorrips, R.E. Broad spectrum insect resistance and metabolites in close relatives of the cultivated tomato. Euphytica 2018, 214, 46. [Google Scholar] [CrossRef] [Green Version]
- Muigai, S.G.; Bassett, M.J.; Schuster, D.J.; Scott, J.W. Greenhouse and field screening of wild Lycopersicon germplasm for resistance to the whitefly Bemisia argentifolii. Phytoparasitica 2003, 31, 27–38. [Google Scholar] [CrossRef]
- Lucatti, A.F.; Alvarez, A.E.; Machado, E.G.; Gilardón, E. Resistance of tomato genotypes to the greenhouse whitefly Trialeurodes vaporariorum (West.) (Hemiptera: Aleyrodidae). Neotrop. Entomol. 2010, 39, 792–798. [Google Scholar] [CrossRef]
- Romanow, L.R.; de Ponti, O.M.B.; Mollema, C. Resistance in tomato to the greenhouse whitefly: Analysis of population dynamics. Entomol. Exp. Appl. 1991, 60, 247–259. [Google Scholar] [CrossRef]
- De Ponti, O.M.B.; Mollema, C. Emerging breeding strategies for insect resistance. In Plant Breeding in the 1990s; Stalker, H.T., Murphy, J.P., Eds.; CAB International: Wallingford, UK, 1992; pp. 323–346. [Google Scholar]
- Maliepaard, C.; Bas, N.; van Heusden, S.; Kos, J.; Pet, G.; Verkerk, R.; Vrieunk, R.; Zabel, P.; Lindhout, P. Mapping of QTLs for glandular trichome densities and Trialeurodes vaporariorum (greenhouse whitefly) resistance in an F2 from Lycopersicon esculentum × Lycopersicon hirsutum F. glabratum. Heredity 1995, 75, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Wagner, G.J.; Wang, E.; Shepherd, R.W. New approaches for studying and exploiting an old protuberance, the plant trichome. Ann. Bot. 2004, 93, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dicke, M.; Baldwin, I.T. The evolutionary context for herbivore-induced plant volatiles: Beyond the ‘cry for help’. Trends Plant Sci. 2010, 15, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Bleeker, P.M.; Diergaarde, P.J.; Ament, K.; Guerra, J.; Weidner, M.; Schutz, S.; de Both, M.T.J.; Haring, M.A.; Schuurink, R.C. The role of specific tomato volatiles in tomato-whitefly interaction. Plant Physiol. 2009, 151, 925–935. [Google Scholar] [CrossRef] [Green Version]
- Alba, J.M.; Montserrat, M.; Fernandez-Munoz, R. Resistance to the two-spotted spider mite (Tetranychus urticae) by acylsucroses of wild tomato (Solanum pimpinellifolium) trichomes studied in a recombinant inbred line population. Exp. Appl. Acarol. 2009, 47, 35–47. [Google Scholar] [CrossRef]
- Vosman, B.; Kashaninia, A.; van’t Westende, W.; Meijer-Dekens, F.; van Eekelen, H.; Visser, R.G.F.; de Vos, R.C.H.; Voorrips, R.E. QTL mapping of insect resistance components of Solanum galapagense. Theor. Appl. Genet. 2018, 132, 531–541. [Google Scholar] [CrossRef] [Green Version]
- Goffreda, J.C.; Mutschler, M.A.; Tingey, W.M. Feeding behavior of potato aphid affected by glandular trichomes of wild tomato. Entomol. Exp. Appl. 1988, 48, 101–107. [Google Scholar] [CrossRef]
- Oriani, M.A.d.G.; Vendramim, J.D. Influence of trichomes on attractiveness and ovipositional preference of Bemisia tabaci (Genn.) B biotype (Hemiptera: Aleyrodidae) on tomato genotypes. Neotrop. Entomol. 2010, 39, 1002–1007. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Munoz, R.; Salinas, M.; Alvarez, M.; Cuartero, J. Inheritance of resistance to two-spotted spider mite and glandular leaf trichomes in wild tomato Lycopersicon pimpinellifolium (Jusl.) Mill. J. Am. Soc. Hortic. Sci. 2003, 128, 188–195. [Google Scholar] [CrossRef] [Green Version]
- Gentile, A.G.; Stoner, A.K.; Webb, R.E. Resistance in Lycopersicon and Solanum to greenhouse whiteflies. J. Econ. Entomol. 1968, 61, 1355–1357. [Google Scholar] [CrossRef]
- Firdaus, S.; van Heusden, A.W.; Hidayati, N.; Supena, E.D.J.; Visser, R.G.F.; Vosman, B. Resistance to Bemisia tabaci in tomato wild relatives. Euphytica 2012, 187, 31–45. [Google Scholar] [CrossRef] [Green Version]
- McDaniel, T.; Tosh, C.R.; Gatehouse, A.M.R.; George, D.; Robson, M.; Brogan, B. Novel resistance mechanisms of a wild tomato against the glasshouse whitefly. Agron. Sustain. Dev. 2016, 36, 14. [Google Scholar] [CrossRef] [PubMed]
- Maluf, W.R.; Maciel, G.M.; Gomes, L.A.A.; Cardoso, M.d.G.; Goncalves, L.D.; da Silva, E.C.; Knapp, M. Broad-spectrum arthropod resistance in hybrids between high- and low-acylsugar tomato lines. Crop Sci. 2010, 50, 439–450. [Google Scholar] [CrossRef]
- Silva, K.F.A.S.; Michereff-Filho, M.; Fonseca, M.E.N.; Silva-Filho, J.G.; Texeira, A.C.A.; Moita, A.W.; Torres, J.B.; Fernandez-Muñoz, R.; Boiteux, L.S. Resistance to Bemisia tabaci biotype B of Solanum pimpinellifolium is associated with higher densities of type IV glandular trichomes and acylsugar accumulation. Entomol. Exp. Appl. 2014, 151, 218–230. [Google Scholar] [CrossRef] [Green Version]
- Rojas, M.R.; Macedo, M.A.; Maliano, M.R.; Soto-Aguilar, M.; Souza, J.O.; Briddon, R.W.; Kenyon, L.; Bustamante, R.F.R.; Zerbini, F.M.; Adkins, S.; et al. World management of geminiviruses. Annu. Rev. Phytopathol. 2018, 56, 637–677. [Google Scholar] [CrossRef]
- Lee, D.-H.; Nyrop, J.P.; Sanderson, J.P. Effect of host experience of the greenhouse whitefly, Trialeurodes vaporariorum, on trap cropping effectiveness. Entomol. Exp. Appl. 2010, 137, 193–203. [Google Scholar] [CrossRef]
- Darshanee, H.L.C.; Ren, H.; Ahmed, N.; Zhang, Z.F.; Liu, Y.H.; Liu, T.X. Volatile-mediated attraction of greenhouse whitefly Trialeurodes vaporariorum to tomato and eggplant. Front. Plant Sci. 2017, 8, 1285. [Google Scholar] [CrossRef] [Green Version]
- Escobar-Bravo, R.; Alba, J.M.; Pons, C.; Granell, A.; Kant, M.R.; Moriones, E.; Fernandez-Muñoz, R. A jasmonate-inducible defense trait transferred from wild into cultivated tomato establishes increased whitefly resistance and reduced viral disease Incidence. Front. Plant Sci. 2016, 7, 1732. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Lopez, M.J.; Garzo, E.; Bonani, J.P.; Fernandez-Muñoz, R.; Moriones, E.; Fereres, A. Acylsucrose-producing tomato plants forces Bemisia tabaci to shift its preferred settling and feeding site. PLoS ONE 2012, 7, e33064. [Google Scholar] [CrossRef] [Green Version]
- Goffreda, J.C.; Steffens, J.C.; Mutschler, M.A. Association of epicuticular sugars with aphid resistance in hybrids with wild tomato. J. Am. Soc. Hortic. Sci. 1990, 115, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Perring, T.M.; Stansly, P.A.; Liu, T.X.; Smith, H.A.; Andreason, S.A. Whiteflies: Biology, ecology, and management. In Sustainable Management of Arthropod Pests of Tomato; Wakil, W., Brust, G.E., Perring, T.M., Eds.; Academic Press: San Diego, CA, USA, 2018. [Google Scholar]
- Kaplan, E.L.; Meier, P. Nonparametric estimation from incomplete observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Hosmer, D.W.; Lemeshow, S. Introduction to the logistic regression model. In Applied Logistic Regression; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2000; pp. 1–30. [Google Scholar]
- Snyder, J.C.; Carter, C.D. Leaf trichomes and resistance of Lycopersicum hirsutum and Lycopersicum esculentum to spider mites. J. Am. Soc. Hortic. Sci. 1984, 109, 837–843. [Google Scholar]
- Goffreda, J.C.; Szymkowiak, E.J.; Sussex, I.M.; Mutschler, M.A. Chimeric tomato plants show that aphid resistance and triacylglucose production are epidermal autonomous characters. Plant Cell 1990, 2, 643–649. [Google Scholar] [PubMed] [Green Version]
- Fery, R.L.; Kennedy, G.G. Genetic analysis of 2-tridecanone concentration, leaf trichome characteristics, and tobacco hornwork resistance in tomato. J. Am. Soc. Hortic. Sci. 1987, 112, 886–891. [Google Scholar]
- Calabrese, E.J.; Edwards, L.J. Light and gravity in leaf-side selection by green peach aphid, Myzus persicae. Ann. Entomol. Soc. Am. 1976, 69, 1145–1146. [Google Scholar] [CrossRef]
- Simmons, A.M. Settling of crawlers of Bemisia tabaci (Homoptera: Aleyrodidae) on five vegetable hosts. Ann. Entomol. Soc. Am. 2002, 95, 464–468. [Google Scholar] [CrossRef] [Green Version]
- Guershon, M.; Gerling, D. Effects of plant and prey characteristics on the predatory behavior of Delphastus catalinae. Entomol. Exp. Appl. 2006, 121, 15–21. [Google Scholar] [CrossRef]
- Liedl, B.E.; Lawson, D.M.; White, K.K.; Shapiro, J.A.; Cohen, D.E.; Carson, W.G.; Trumble, J.T.; Mutschler, M.A. Acylsugars of wild tomato Lycopersicon pennellii alters settling and reduces oviposition of Bemisia argentifolii (Homoptera:Aleyrodidae). J. Econ. Entomol. 1995, 88, 742–748. [Google Scholar] [CrossRef]
- Alba, J.M. Herencia de los Mecanismos de Resistencia a Arena Roja en Tomate. Ph.D. Thesis, Facultad de Ciencias, Universidad de Málaga, Málaga, Spain, 2006. [Google Scholar]
- Smeda, J.R.; Schilmiller, A.L.; Anderson, T.; Ben-Mahmoud, S.; Ullman, D.E.; Chappell, T.M.; Kessler, A.; Mutschler, M.A. Combination of acylglucose QTL reveals additive and epistatic genetic interactions and impacts insect oviposition and virus infection. Mol. Breed. 2017, 38, 3. [Google Scholar] [CrossRef]
- Leckie, B.M.; D’Ambrosio, D.A.; Chappell, T.M.; Halitschke, R.; De Jong, D.M.; Kessler, A.; Kennedy, G.G.; Mutschler, M.A. Differential and synergistic functionality of acylsugars in suppressing oviposition by insect herbivores. PLoS ONE 2016, 11, e0153345. [Google Scholar] [CrossRef] [Green Version]
- Buta, J.G.; Lusby, W.R.; Neal, J.W.; Waters, R.M.; Pittarelli, G.W. Sucrose esters from Nicotiana gossei active against the greenhouse whitefly Trialeuroides vaporariorum. Phytochemistry 1993, 32, 859–864. [Google Scholar] [CrossRef]
- Woets, J.; van Lenteren, J.C. The parasite-host relationship between Encarsia formosa (Hymenoptera: Aphelinidae) and Trialeurodes vaporariorum (Homoptera: Aleyrodidae). 6. The influence of the host plant of the greenhouse white fly and its parasite Encarsia formosa. Bull. Srop 1976, 1974, 151–164. [Google Scholar]
- van de Merendonk, S.; van Lenteren, J.V. Determination of mortality of greenhouse whitefly Trialeurodes vaporariorum (Westwood) (Homoptera: Aleyrodidae) eggs, larvae and pupae on four host-plant species: Eggplant (Solanum melongena L.), cucumber (Cucumis sativus L.), tomato (Lycopersicum esculentum L.) and paprika (Capsicum annuum L.). Med. Van De Fac. Landbouwwet. Rijksuniv. Gent 1978, 43, 421–429. [Google Scholar]
- Mutschler, M.A.; Steffens, J.C.; Tingey, W. Broad based insect resistance from L. pennellii: Its nature and potential for pest control in tomato. In Molecular Biology of Tomato: Fundamental Advances and Crop Improvement; Yoder, J.I., Ed.; Technomic publishing Co., Inc.: Lancaster, PA, USA, 1993; pp. 285–290. [Google Scholar]
- Berlinger, M.J.; Dahan, R. Breeding for resistance to virus transmission by whiteflies in tomatoes. Int. J. Trop. Insect Sci. 1987, 8, 783–784. [Google Scholar] [CrossRef]
- Resende, J.T.d.; Maluf, W.R.; Cardoso, M.d.G.; Gonçalves, L.D.; Faria, M.V.; Nascimento, I.R.d. Resistance of tomato genotypes to the silverleaf whitefly mediated by acylsugars. Hortic. Bras. 2009, 27, 345–348. [Google Scholar] [CrossRef] [Green Version]
- De Ponti, O.M.B.; Romanow, L.R.; Berlinger, M.J. Whitefly plant relationships—Plant resistance. In Whiteflies: Their Bionomics, Pest Status and Management; Gerling, D., Ed.; Gerling Dan Intercept Press: Andover, UK, 1990; pp. 91–106. [Google Scholar]
- Van Lenteren, J.C. A greenhouse without pesticides: Fact or fantasy? Crop Prot. 2000, 19, 375–384. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-López, M.J.; Moriones, E.; Fernández-Muñoz, R. An Acylsucrose-Producing Tomato Line Derived from the Wild Species Solanum pimpinellifolium Decreases Fitness of the Whitefly Trialeurodes vaporariorum. Insects 2020, 11, 616. https://doi.org/10.3390/insects11090616
Rodríguez-López MJ, Moriones E, Fernández-Muñoz R. An Acylsucrose-Producing Tomato Line Derived from the Wild Species Solanum pimpinellifolium Decreases Fitness of the Whitefly Trialeurodes vaporariorum. Insects. 2020; 11(9):616. https://doi.org/10.3390/insects11090616
Chicago/Turabian StyleRodríguez-López, María J., Enrique Moriones, and Rafael Fernández-Muñoz. 2020. "An Acylsucrose-Producing Tomato Line Derived from the Wild Species Solanum pimpinellifolium Decreases Fitness of the Whitefly Trialeurodes vaporariorum" Insects 11, no. 9: 616. https://doi.org/10.3390/insects11090616
APA StyleRodríguez-López, M. J., Moriones, E., & Fernández-Muñoz, R. (2020). An Acylsucrose-Producing Tomato Line Derived from the Wild Species Solanum pimpinellifolium Decreases Fitness of the Whitefly Trialeurodes vaporariorum. Insects, 11(9), 616. https://doi.org/10.3390/insects11090616