
Fossils Shed a New Light on the Diversity and Disparity of the Family Limoniidae (Diptera, Nematocera)

 , , , and
, , , and
Abstract
:Simple Summary
Abstract

1. Introduction
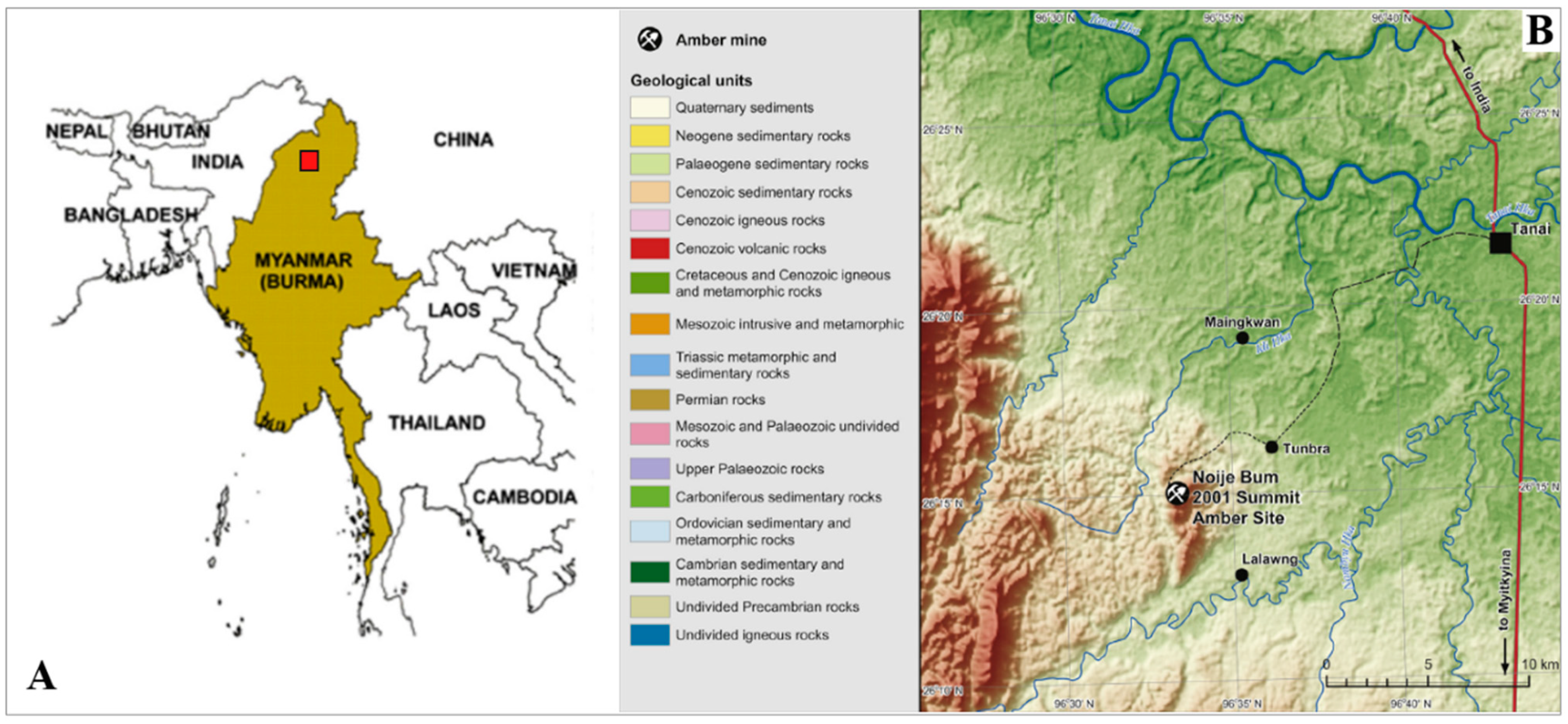
2. Materials and Methods
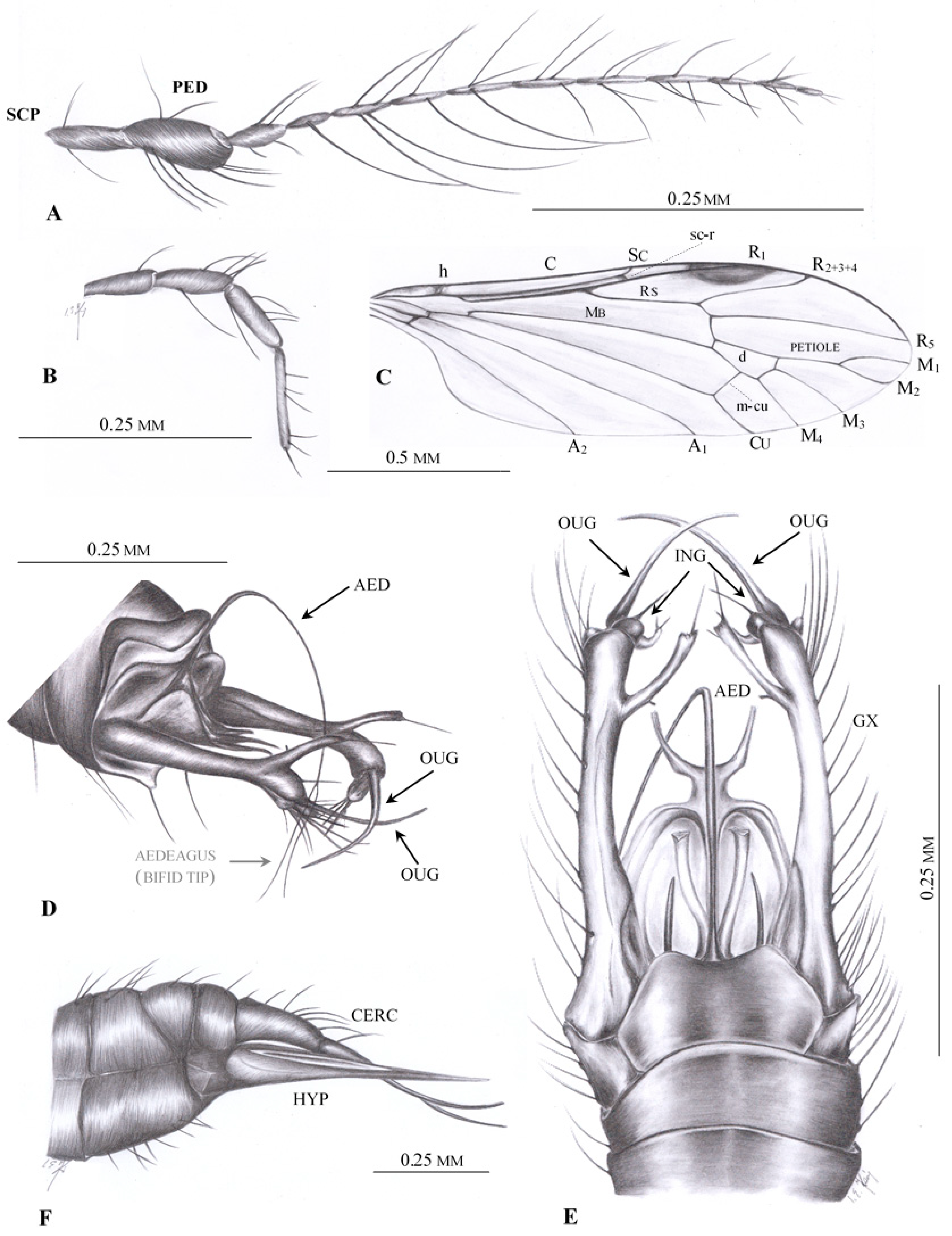
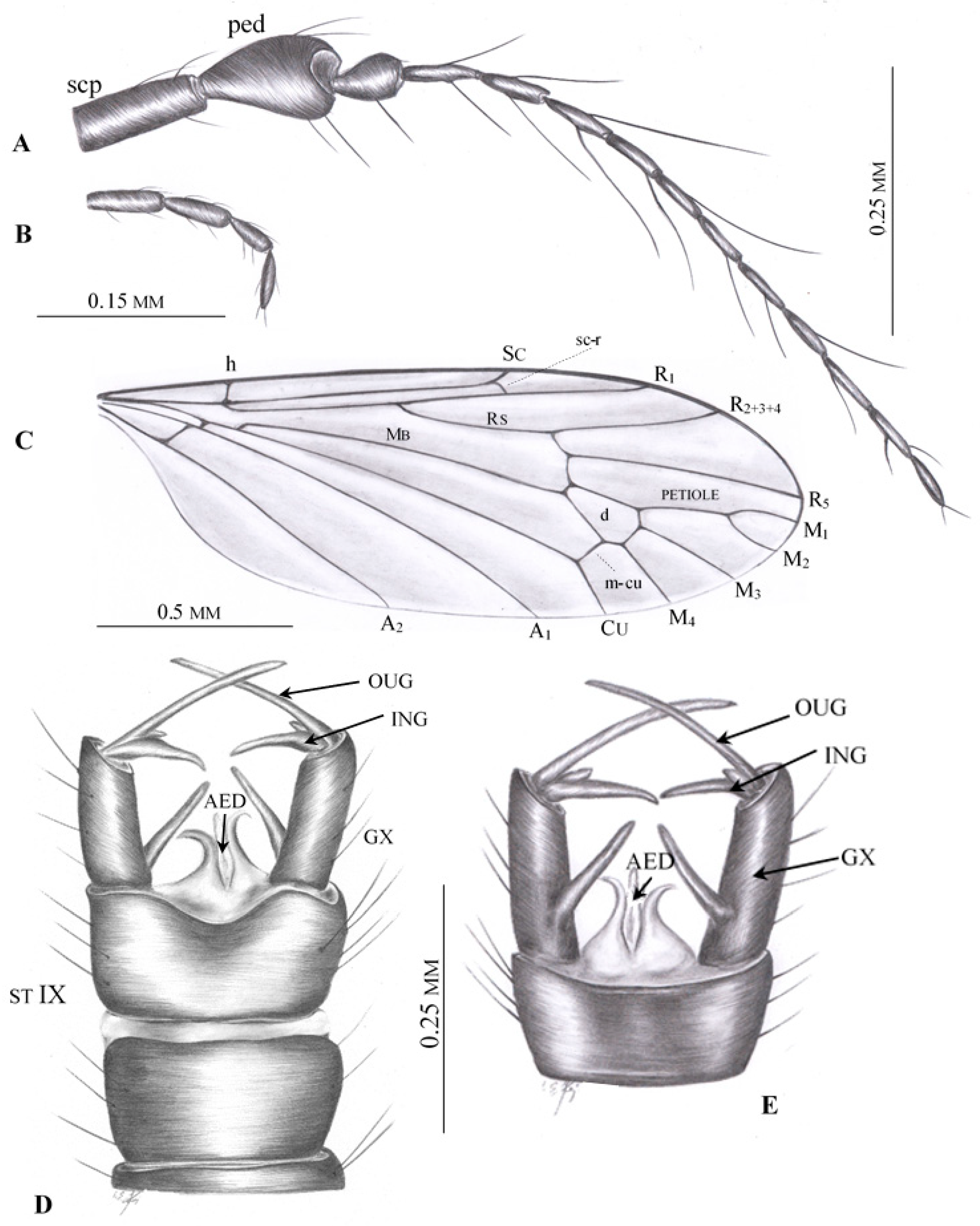
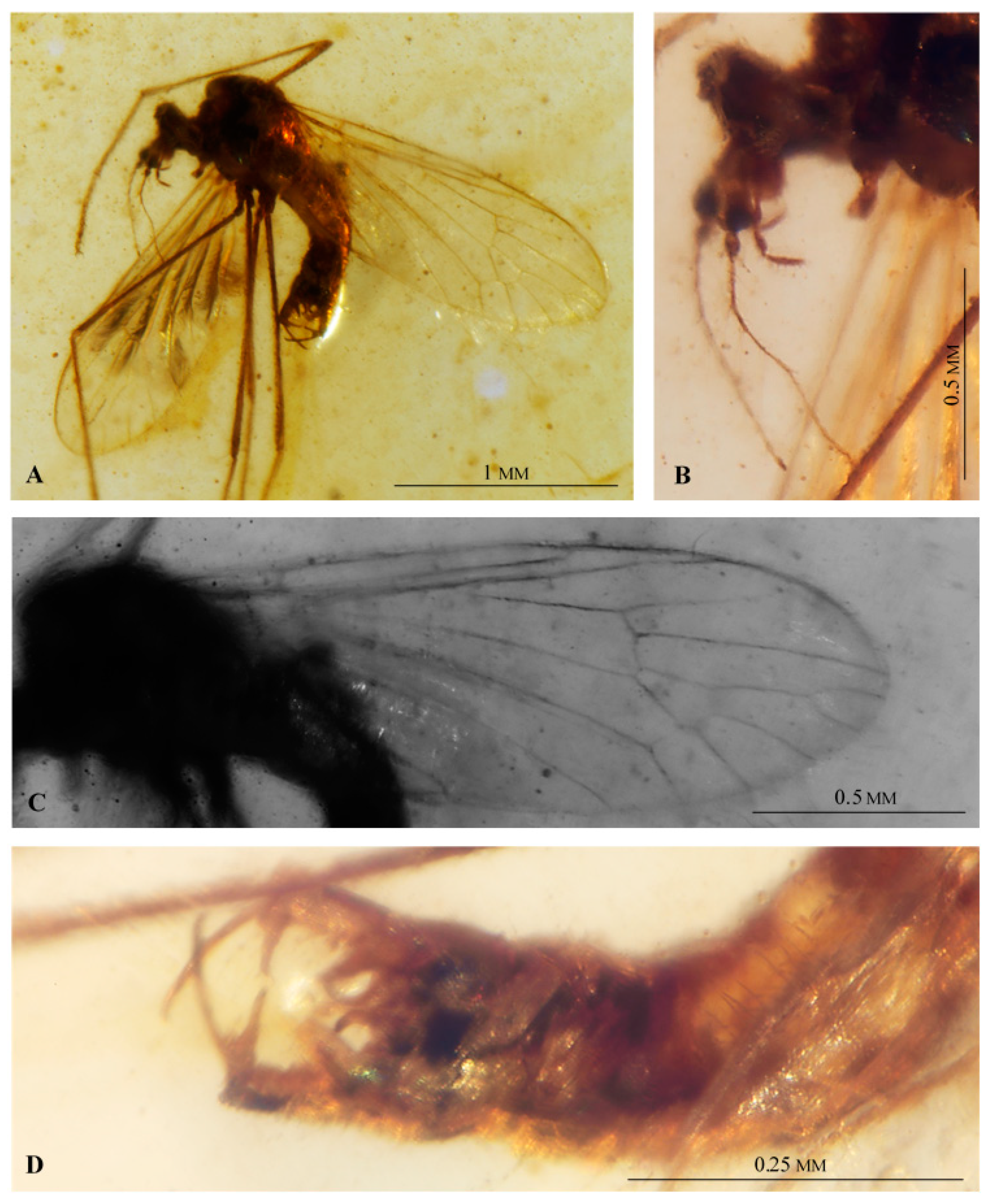
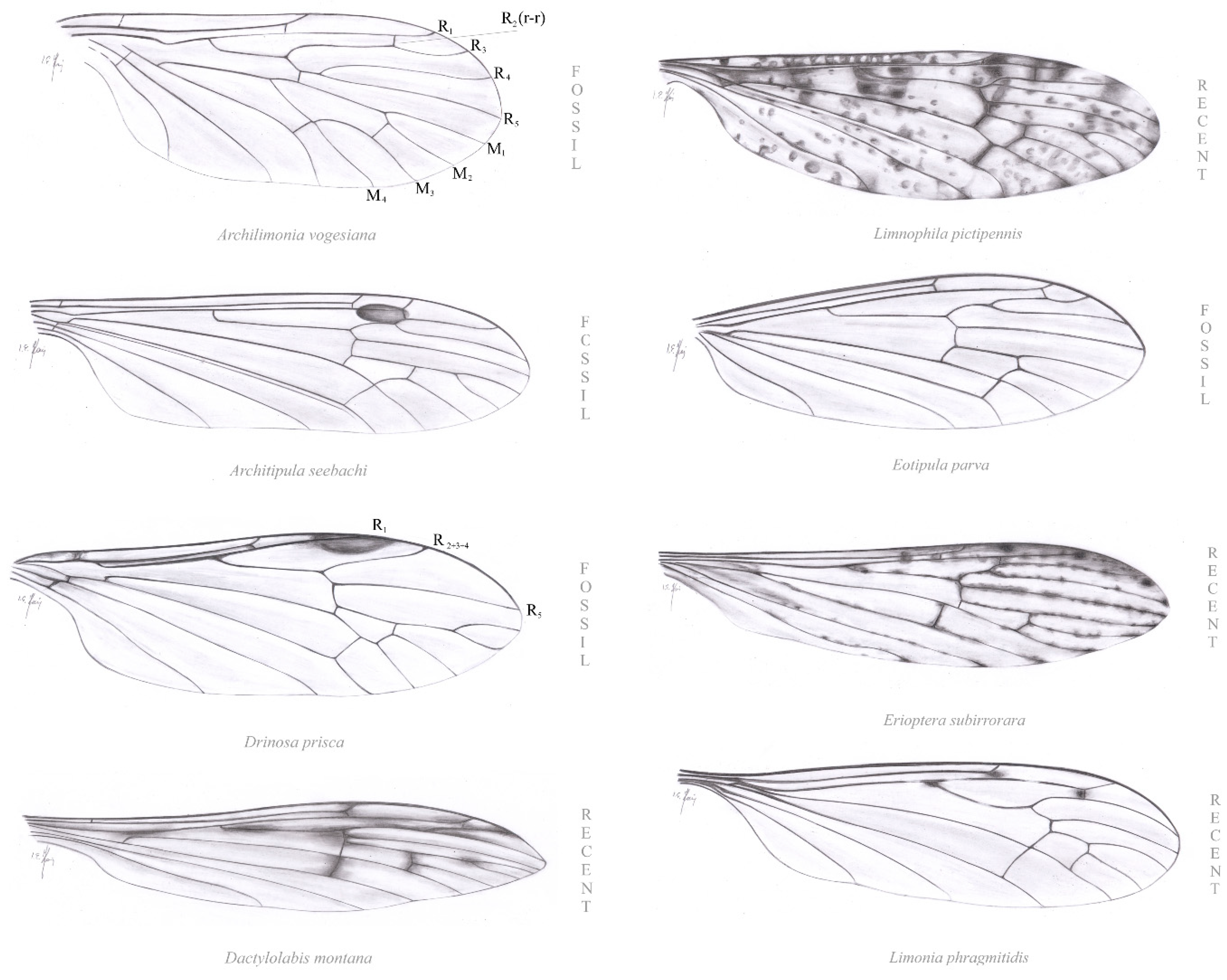
3. Results
Systematic Paleontology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krzemiński, W.; Evenhuis, N.L. Review of Diptera palaeontological. In 2000 Contributions to a Manual of Palaearctic Diptera; Papp, L., Davas, B., Eds.; Science Heraid: Budapest, Hungary, 2000; Volume 1, pp. 535–564. [Google Scholar]
- Krzemiński, W.; Krzemińska, E.; Papier, F. Grauvogelia arzvilleriana sp. n.—The oldest Diptera species (Lower/Middle Triassic of France). Acta Zool. Crac. 1994, 37, 95–99. [Google Scholar]
- Grimaldi, D.A.; Engel, M.S.; Nascimbene, P.C. Fossiliferous Cretaceous amber from Myanmar (Burma): Its rediscovery, biotic diversity, and paleontological significance. Am. Mus. Novit. 2002, 3361, 1–71. [Google Scholar] [CrossRef] [Green Version]
- Pohl, H.W.; Wipfler, B.; Boudinot, B.; Beutel, R.G. On the value of Burmese amber for understanding insect evolution: Insights from Heterobathmilla—An exceptional stem group genus of Strepsiptera (Insecta). Cladisctics 2020, 1–19. [Google Scholar] [CrossRef]
- Peris, D.; Ruzzier, E.; Perrichot, W.; Delclòs, X. Evolutionary and paleobiological implications of Coleoptera (Insecta) from Tethyan-influenced Cretaceous ambers. Geosci. Front. 2016, 7, 695–706. [Google Scholar] [CrossRef] [Green Version]
- Peris, D.; Kundrata, R.; Delclòs, X.; Mähler, B.; Ivie, M.A.; Rust, J.; Labandeira, C.C. Unlocking the mystery of the mid-Cretaceous Mysteriomorphidae (Coleoptera: Elateroidea) and modalities in transiting from gymnosperms to angiosperms. Sci. Rep. 2020, 10, 16854. [Google Scholar] [CrossRef]
- Blagoderov, V.; Grimaldi, D.A. Fossil Sciaroidea (Diptera) in Cretaceous ambers, exclusive Cecidimyiidae, Sciaridae, and Keroplatidae. Am. Mus. Novit. 2009, 3433, 1–76. [Google Scholar] [CrossRef]
- Grimaldi, D.A.; Arillo, A.; Cumming, J.M.; Hauser, M. Brachyceran Diptera (Insecta) in Cretaceous ambers, Part IV, Significant New Orthorrhaphous Taxa. ZooKeys 2011, 148, 293–332. [Google Scholar] [CrossRef]
- Shi, G.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.; Lei, W.; Li, Q.; Li, X. Age constraint on Burmese amber based on U-Pb dating of zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Ross, A.J. Burmese (Myanmar) amber checklist and bibliography 2018. Palaeoentomology 2020, 2, 22–84. [Google Scholar] [CrossRef]
- Krzemiński, W.; Krzemińska, E. Triassic Diptera: Review, revisions and descriptions. Acta Zool. Crac. 2003, 46 (Suppl. Fossil Insects), 153–184. [Google Scholar]
- Krzemiński, W. Fossil Limoniidae (Diptera, Tipulomorpha) from Lower Cretaceous Burmese amber of Myanmar. J. Syst. Palaeontol. 2004, 2, 123–125. [Google Scholar] [CrossRef]
- Podenas, S.; Poinar, G.O. New crane flies (Diptera: Limoniidae) from Burmese amber. Proc. Entomol. Soc. Wash. 2009, 111, 470–492. [Google Scholar] [CrossRef]
- Kania, I.; Bo, W.; Szwedo, J. Dicranoptycha Osten Sacken, 1860 (Diptera, Limoniidae) from the earliest Cenomanian Burmese amber. Cretac. Res. 2015, 52, 522–530. [Google Scholar] [CrossRef]
- Men, Q.; Hu, Z.; Mu, D. New species of Lebania (Diptera: Limoniidae) from mid-Cretaceous amber of northern Myanmar. Cretac. Res. 2019, 104, 1–5. [Google Scholar] [CrossRef]
- Kopeć, K.; Krzemiński, W.; Skowron, K.; Coram, R. The genera Architipula Handlirsch, 1906 and Grimmenia Krzemiński and Zessin, 1990 (Diptera: Limoniidae) from the Lower Jurassic of England. Palaeontol. Electron. 2017, 20, 1–7. [Google Scholar] [CrossRef]
- Lepeletier, A.L.M.; Serville, J.G.A. Entomologie, ou histoire naturelle des crustacés, des arachnides et des insectes. Encycl. Method. Hist. Nat. 1828, 10, 345–833. [Google Scholar]
- Meigen, J.W. Systematische Beschreibung der Bekannten Europäischen Zweiflügeligen Insecten; Erster Theil, F.W., Ed.; Forstmann: Aachen, Germany, 1818; Volume XXXVI, p. 332. [Google Scholar]
- Osten-Sacken, C. New genera and species of North American Tipulidae with short palpi, with an attempt at a new classification of the tribe. Proc. Acad. Nat. Sci. Phila. 1859, 1860, 197–256. [Google Scholar]
- Bigot, J.M.F. Essai d’une classification génér insectsale et synoptique de l’ordre des insectes diptères (3è memoire). Tribu de Tipulidii (mihi). Ann. De La Société Entomol. De Fr. 1854, 3, 447–482. [Google Scholar]
- Rondani, C. Progetto di una Classificazione in Famiglie degli Insetti Ditteri Europei. Memoria Terza per Servir alla Ditterologia Italiana; Donati: Parma, Italy, 1841; pp. 1–28. [Google Scholar]
- Alexander, C.P. New or little-known Tipulidae (Diptera). III. Ethiopian species. Ann. Mag. Nat. Hist. 1920, 9, 465–472. [Google Scholar] [CrossRef]
- Speiser, P. 4 Orthoptera. Orthoptera Nematocera. Wissenschaftliche Ergebnisse der Schwedischen Zoologische Expededition nach Kilimandjaro. Meru 10 (Diptera) 1909, 31–65. [Google Scholar]
- Handlirsch, A. Die fossilen Insekten und Die Phylogenie der Rezenten Formen; Ein Handbuch für Paläontologen und Zoologen; Engelmann: Leipzig, Germany, 1906; pp. 481–640. [Google Scholar]
- Kalugina, N.S.; Kovalev, V.G. Dipterous Insects of Jurassic Siberia. Paleontological Institute; Akademia Nauk: Moscow, Russia, 1985; 198p. (In Russian) [Google Scholar]
- Krzemiński, W.; Kovalev, V.G. The taxonomic status of Architipula fragmentosa (Bode) and the family Eoasilidae (Diptera) from the Lower Jurassic. Syst. Entomol. 1988, 13, 55–56. [Google Scholar] [CrossRef]
- Krzemiński, W. Triassic and Lower Jurassic stage of Diptera evolution. Mitt. Schweiz. Entomol. Ges. 1992, 65, 39–59. [Google Scholar]
- Zhang, J. Nematoceran Dipteras from the Jurassic of China (Insecta, Diptera: Limoniidae, Tanyderidae). Paleontol. Zhurnal 2004, 5, 53–57. (In Russian) [Google Scholar]
- Zhang, J. Jurassic limoniid Dipterans from China (Diptera: Limoniidae). Orient. Insects 2006, 40, 115–126. [Google Scholar] [CrossRef]
- Blagoderov, V.A.; Grimaldi, D.A.; Fraser, N.C. How time flies for flies: Diverse Diptera from the Triassic of Virginia and early radiation of the order. Am. Mus. Novit. 2007, 3572, 1–39. [Google Scholar] [CrossRef]
- Evenhuis, N.L. Catalogue of The Fossil Flies of the World (Insecta: Diptera). 2015. Available online: http://hbs.bishopmuseum.org/fossilcat/ (accessed on 29 May 2015).
- Lukashevich, E.D.; Ribeiro, G.C. Mesozoic fossil and the phylogeny of Tipulomorpha (Insecta: Diptera). J. Syst. Paleontol. 2019, 17, 635–652. [Google Scholar] [CrossRef]
- Handlirsch, A. Neue Untersuchungenüber die fossilen Insekten. II. Ann. Nat. Mus. Wien 1939, 49, 1–240. [Google Scholar]
- Krzemiński, W.; Zessin, W. The lower Jurassic Limoniidae from Grimmen (Diptera, Nematocera). Dtsch. Entomol. Z. 1990, 37, 39–43. [Google Scholar] [CrossRef]
- Ansorge, J. Insekten aus dem Oberen Lias von Grimmen (Vorpommern, Norddeutschland). Neue Palaontol. Abh. 1996, 2, 1–132. [Google Scholar]
- Kania, I.; Krzemiński, W.; Krzemińska, E. The oldest representative of the genus Gonomyia (Diptera, Limoniidae) from Lebanese amber (Early Cretaceous): New subgenus and species. Cretac. Res. 2015, 52, 516–521. [Google Scholar] [CrossRef]
- Krzemiński, W.; Kania, I.; Krzemińska, E. A new species of Dactylolabis (Eobothrophorus) from Baltic amber (Diptera: Limoniidae). Acta Geol. Sin. 2010, 84, 768–771. [Google Scholar] [CrossRef]
- Krzemiński, W.; Kania, I.; Azar, D. The Early Cretaceous evidence of rapid evolution of the genus Helius Lepeletier and Serville, 1828 (Limoniidae, Diptera). Cretac. Res. 2014, 48, 96–101. [Google Scholar] [CrossRef]
- Krzemińska, E.; Krzemiński, W.; Dahl, C. Monograph of Fossil Trichoceridae (Diptera) over 180 Million Years of Evolution; Institute of Systematics and Evolution of Animals of the Polish Academy of Sciences: Krakow, Poland, 2009; 171p. [Google Scholar]
- McAlpine, J.F.; Peterson, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R.; Wood, D.M. Manual of Nearctic Diptera; Research Branch Agriculture; Canada Communication Group Pub: Ottawa, ON, Canada, 1981; Volume 1, pp. 27–674. [Google Scholar]
- Cruikshank, R.D.; Ko, K. Geology of an amber locality in the Hukawng Valley, Northern Myanmar. J. Asian Earth Sci. 2003, 21, 441–455. [Google Scholar] [CrossRef]
- Wandrey, C.J. Eocene to Miocene composite total petroleum system, Irrawaddy-Andaman and North Burma geologic provinces, Myanmar. In Petroleum Systems and Related Geologic Studies in Region 8, South Asia; Wandrey, C.J., Ed.; US Geological Survey US 2208-E; Department of the Interior: Washington, DC, USA, 2006; 26p. [Google Scholar]
- Kania, I.; Wojtoń, M.; Lukashevich, E.; Stanek-Tarkowska, J.; Wang, B.; Krzemiński, W. Anisopodidae (Insecta: Diptera) from Upper Cretaceous Amber of Northern Myanmar. Cretac. Res. 2019, 94, 190–206. [Google Scholar] [CrossRef]
- Linnaeus, C. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis; T. 1; Impensis Direct. Laurentii Salvii: Holmiae, Sweden, 1758; 800p. [Google Scholar]
- Rohdendorf, B.B. The oldest infraorders of Diptera from the Triassic of Middle Asia. Palaeontol. Zhurnal 1961, 2, 90–100. (In Russian) [Google Scholar]
- Loew, H. Ueber den Bernstein und die Bernsteinfauna. Program. der Königlichen Realschule zu Meseritz Mitler 1850, 1–44. [Google Scholar]
- Kopeć, K. A new species of Mesotipula (Limoniidae, Diptera) from the Lower Jurassic of England. Ann. Zool. 2018, 68, 93–96. [Google Scholar] [CrossRef]
- Kopeć, K.; Ansorge, J.; Soszyńska-Maj, A.; Krzemiński, W. Revision of the genus Mesotipula Handlirsch, 1920 (Diptera, Limoniidae, Architipulinae) from the Lower Jurassic of Northeast Germany. Hist. Biol. 2018, 32, 500–507. [Google Scholar] [CrossRef]
- Kopeć, K.; Soszyńska-Maj, A.; Gehler, A.; Ansorge, J.; Krzemiński, W. Mecoptera and Diptera from the early Toarcian (Early Jurassic) deposits of Wolfsburg e GroßeKley (Lower Saxony. Germany). Earth Environ. Sci. Trans. R. Soc. Edinb. 2018, 107, 163–171. [Google Scholar]
- Azar, D.; Maksoud, S. A new species of Protopsychoda Azar et al., 1999 from the Lower Cretaceous. Lebanese amber (Diptera: Psychodidae). Palaeoentomology 2020, 3, 352–356. [Google Scholar] [CrossRef]
- Maksoud, S.; Azar, D. Lebanese amber: Latest updates. Palaeoentomology 2020, 3, 125–155. [Google Scholar] [CrossRef]
- Pawłowski, J.; Kmieciak, D.; Szadziewski, R.; Burkiewicz, A. Próba izolacji DNA owadów z bursztynu bałtyckiego. Attempted isolation of DNA from insects embedded in the Baltic amber. Pr. Muz. Ziemi 1996, 44, 45–46. (In Polish) [Google Scholar]
- Austin, J.; Ross, A.; Smith, A.B.; Fortey, R.A.; Thomas, R.H. Problem of reproducibility does geologically ancient DNA survive in amber-preserved insects? Proc. R. Soc. Lond. 1997, 264, 467–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyfullah, L.; Beimforde, C.; Corso, J.D.; Perrichot, V.; Rikkinen, J.; Schmidt, A. Production and preservation of resins—Past and present. Biol. Rev. 2018, 93, 1684–1714. [Google Scholar] [CrossRef]
- Peris, D.; Janssen, K.; Barthel, J.H.; Bierbaum, G.; Delclòs, X.; Peñalver, E.; Solórzano-Kraemer, M.; Jordal, B.H.; Rust, J. DNA from resin-embedded organisms: Past, present and feature. PLoS ONE 2020, 15, e0239521. [Google Scholar] [CrossRef]
- Lukashevich, E.D. Limoniidae (Diptera) in the Upper Jurassic of Shar Teg, Mongolia. Zoosymposia 2009, 3, 131–154. [Google Scholar] [CrossRef]
- Gao, J.; Shih, C.; Zhao, Y.; Ren, D. New species of Cretolimonia and Mesotipula (Diptera: Limoniidae) from the middle Jurassic of Northeastern China. Acta Geol. Sin. 2015, 89, 1789–1796. [Google Scholar]
- Kopeć, K.; Krzemiński, W. The oldest representatives of the Pilaria (Diptera, Limoniidae) genus from Burmese amber. Cretac. Res. 2021, in press. [Google Scholar] [CrossRef]
- Geinitz, F.E. Über die Fauna des Dobbertiner Lias. Z. Dtsch. Geol. Ges. 1884, 36, 566–583. [Google Scholar]
- Schrank, F.; von Paula, V.E. Enumeratio Insectorum Austriae Indigenorum; Klett and Franck: Augsburg, Germany, 1781; Volume I–XXIV, pp. 1–548. [Google Scholar]
- Papp, L.; Schumann, H. Key to families. Adults. In 2000 Contributions to a Manual of Palaearctic Diptera; Papp, L., Darvas, B., Eds.; Science Heraid: Budapest, Hungary, 2000; Volume 1, pp. 163–200. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Subfamily | Time Scale | The Oledest Known Representative/Species | Age Range (MA) |
|---|---|---|---|---|
| 1. | Chioneinae Rondani, 1841 [21] | Cretaceous—Recent | Gonomyia (Azaria) libanensis Kania, Krzemiński, Krzemińska, 2015 [36] | 140 |
| 2. | Dactylolabinae Alexander, 1920 [22] | Eocene—Recent | Dactylolabis (Eobothrophorus) hoffeinsorum Krzemiński, Kania, Krzemińska, 2010 [37] | 45 |
| 3. | Limnophilinae Bigot, 1854 [20] | Cretaceous—Recent | Austrolimnophila joana Podenas, Poinar, 2009 [13] | 98.79 ± 0.62 |
| 4. | Limoniinae Speiser, 1909 [23] | Cretaceous—Recent | Helius ewa Krzemiński, Kania, Azar, 2014 [38] | 140 |
| 5. | Eotipulinae Kalugina, 1985 [25] | Jurassic | Eotipula parva Krzemiński, 1992 [27] | 221.5–205.6 |
| 6. | Architipulinae Handlirsch, 1906 [33] | Triassic | Architipula youngi Handlirsh, 1906 [33] | 183.0–182.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krzemiński, W.; Kania-Kłosok, I.; Krzemińska, E.; Ševčík, J.; Soszyńska-Maj, A. Fossils Shed a New Light on the Diversity and Disparity of the Family Limoniidae (Diptera, Nematocera). Insects 2021, 12, 206. https://doi.org/10.3390/insects12030206
Krzemiński W, Kania-Kłosok I, Krzemińska E, Ševčík J, Soszyńska-Maj A. Fossils Shed a New Light on the Diversity and Disparity of the Family Limoniidae (Diptera, Nematocera). Insects. 2021; 12(3):206. https://doi.org/10.3390/insects12030206
Chicago/Turabian StyleKrzemiński, Wiesław, Iwona Kania-Kłosok, Ewa Krzemińska, Jan Ševčík, and Agnieszka Soszyńska-Maj. 2021. "Fossils Shed a New Light on the Diversity and Disparity of the Family Limoniidae (Diptera, Nematocera)" Insects 12, no. 3: 206. https://doi.org/10.3390/insects12030206
APA StyleKrzemiński, W., Kania-Kłosok, I., Krzemińska, E., Ševčík, J., & Soszyńska-Maj, A. (2021). Fossils Shed a New Light on the Diversity and Disparity of the Family Limoniidae (Diptera, Nematocera). Insects, 12(3), 206. https://doi.org/10.3390/insects12030206