
Transcriptomic Characterization of Odorant Binding Proteins in Cacia cretifera thibetana and Their Association with Different Host Emitted Volatiles

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. RNA Extraction and Quantification
2.3. cDNA Library Preparation and Transcriptome Sequencing
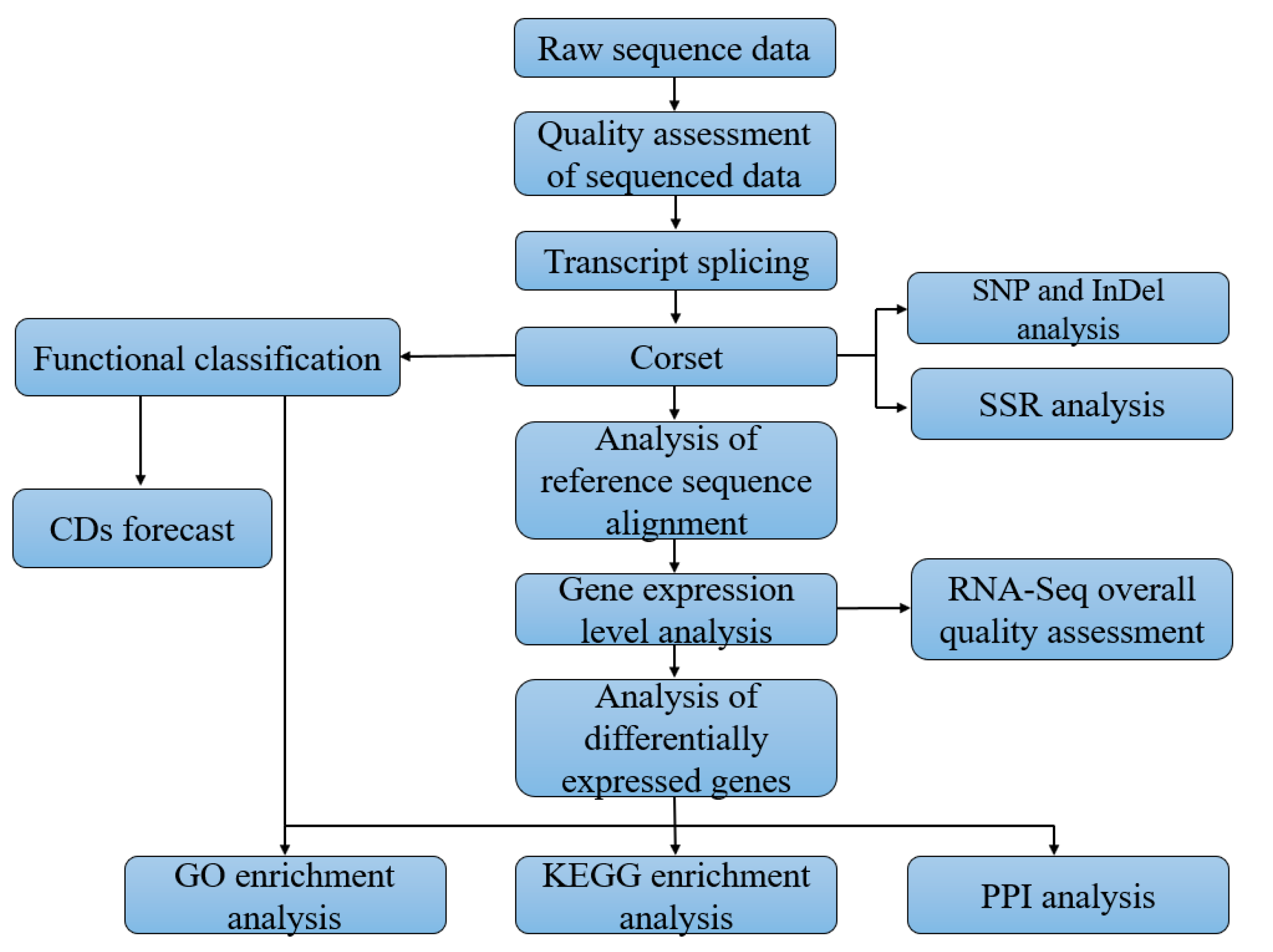
2.4. Quality Control, De Novo Assembly and Functional Annotation
2.5. Identification of Odor Binding Protein (OBP) Genes
2.6. Quantitative Real-Time PCR
2.7. Differential Expression Analysis
2.8. GO and KEGG Enrichment Analysis of Differentially Expressed Genes
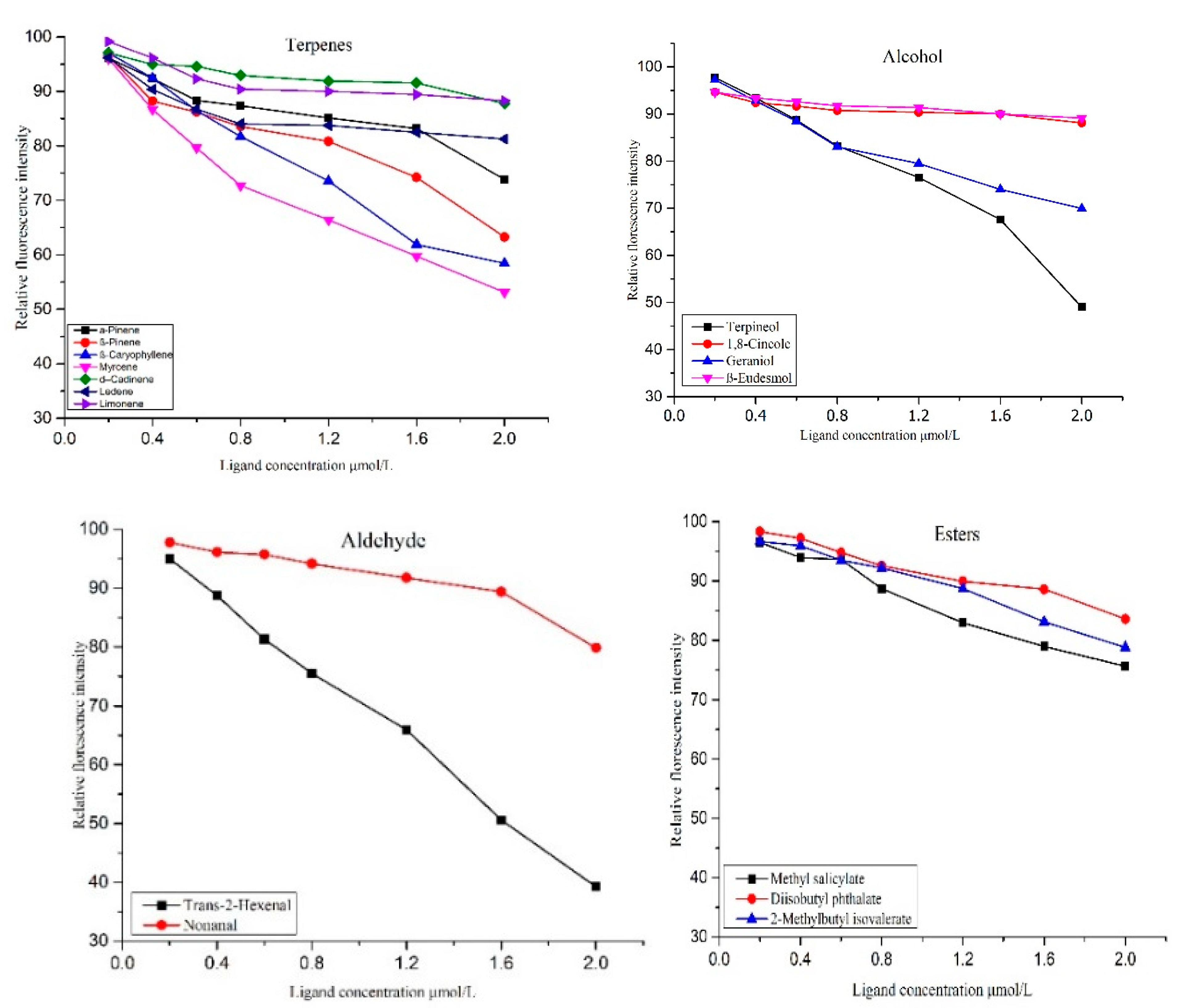
2.9. Evaluation of Binding Affinities of OBP Genes with Host Compounds
3. Results
3.1. The RNA-Sequencing Data
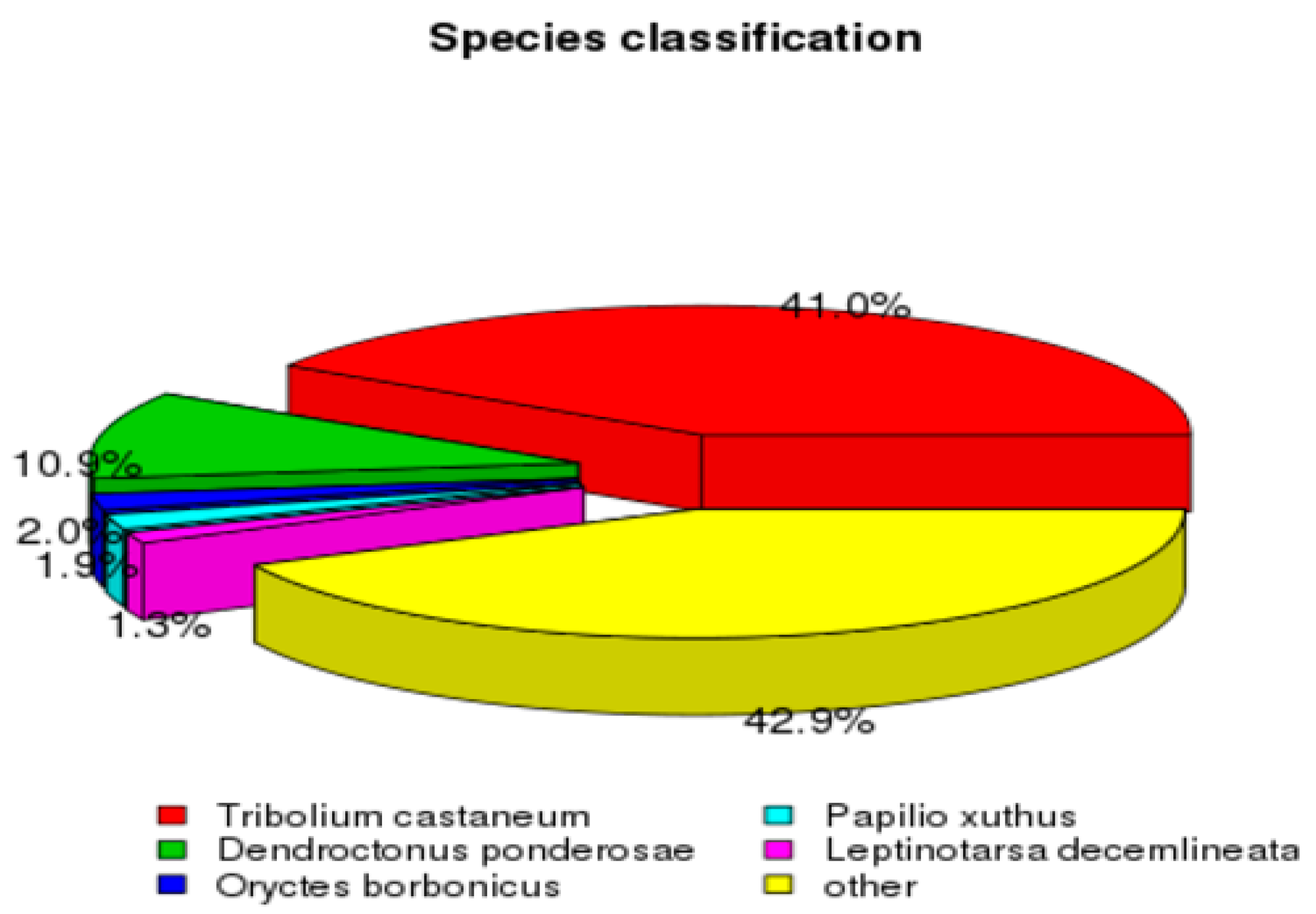
3.2. De Novo Assembly and Transcriptome Annotation of C. cretifera thibetana
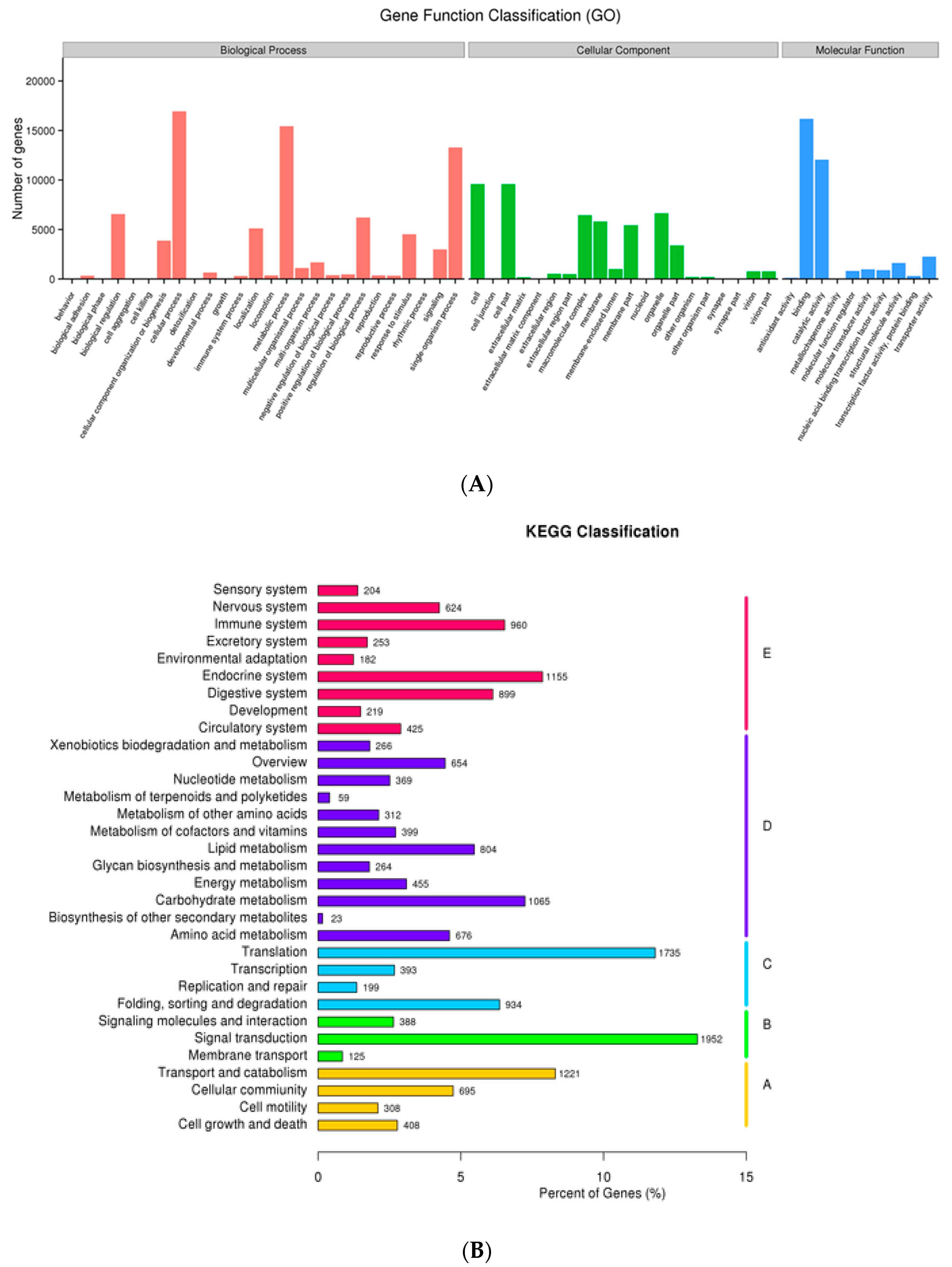
3.3. The Unigene Functional Annotation
3.4. Gene Expression Analyses
3.4.1. The RNA-Seq Data Mapping to Reference Sequence and Transcripts Distribution in Samples
3.4.2. Identification of Differentially Expressed Genes and Functional Enrichment Analyses
3.4.3. Quantitative Real-Time PCR Analysis
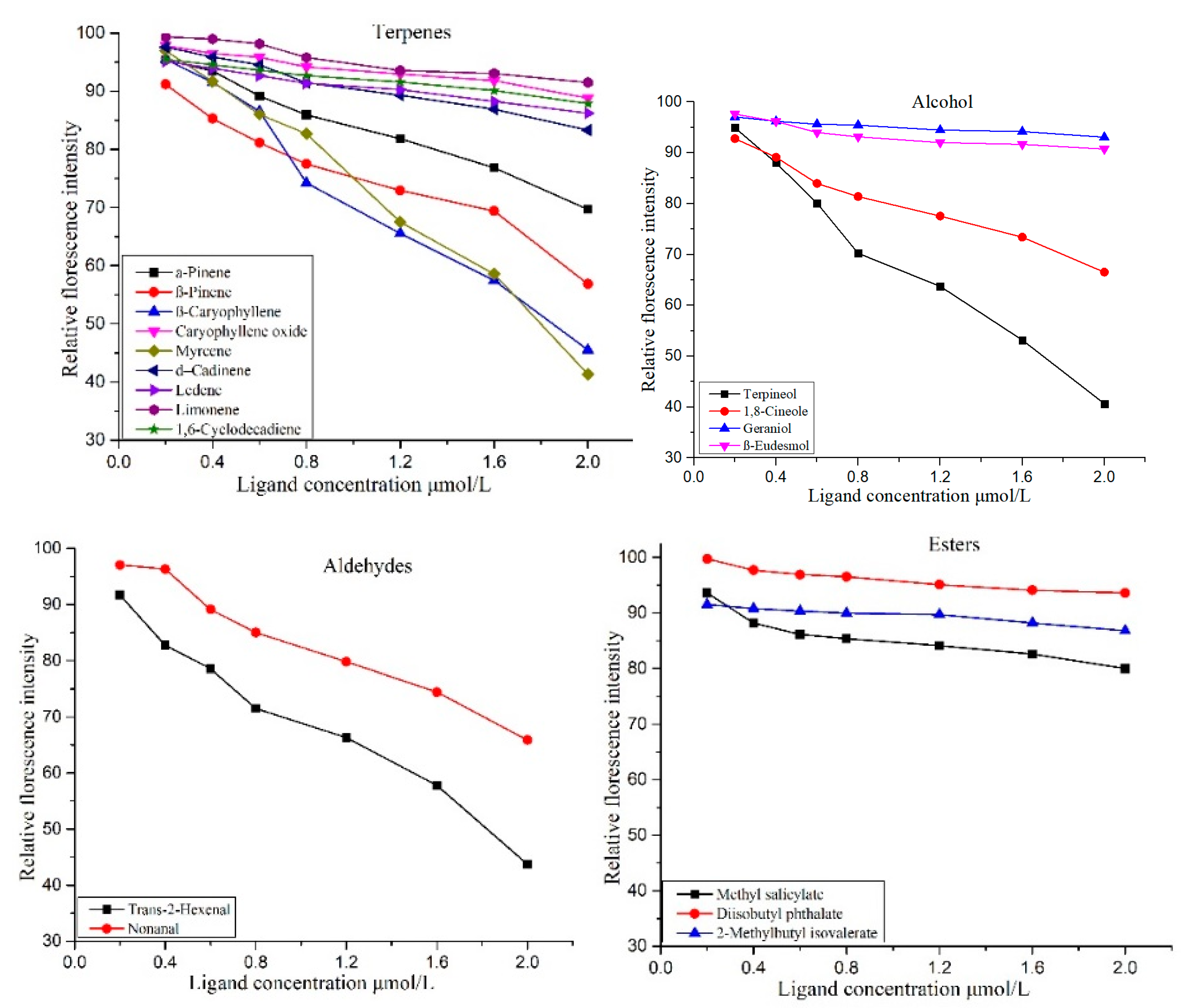
3.4.4. Functional Analysis of Odorant Binding Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Y.; Ding, X. Damage characteristics and comprehensive prevention and control technology of Anoplophora nobilis. Sci. Technol. Qinghai Agric. For. 2016, 1, 35–43. [Google Scholar]
- Williams, D.W.; Lee, H.-P.; Kim, I.-K. Distribution and abundance of Anoplophora glabripennis (Coleoptera: Cerambycidae) in natural acer stands in South Korea. Environ. Entomol. 2004, 33, 540–545. [Google Scholar] [CrossRef]
- Xiaona, C. Recognition and identification of damage features of main longhorn beetles in China. For. Pest Dis. 2009, 6, 1–112. [Google Scholar]
- Hardie, J.; Minks, A.K. Pheromones of Non-Lepidopteran Insects Associated with Agricultural Plants; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Zhang, Q.-H.; Schlyter, F. Olfactory recognition and behavioural avoidance of angiosperm nonhost volatiles by conifer-inhabiting bark beetles. Agric. For. Entomol. 2004, 6, 1–20. [Google Scholar] [CrossRef]
- Erbilgin, N.; Krokene, P.; Kvamme, T.; Christiansen, E. A host monoterpene influences Ips typographus (Coleoptera: Curculionidae, Scolytinae) responses to its aggregation pheromone. Agric. For. Entomol. 2007, 9, 135–140. [Google Scholar] [CrossRef]
- Andersson, M.N.; Larsson, M.C.; Blazenec, M.; Jakuš, R.; Zhang, Q.-H.; Schlyter, F. Peripheral modulation of pheromone response by inhibitory host compound in a beetle. J. Exp. Biol. 2010, 213, 3332–3339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, P.; Wang, J.; Cui, M.; Tao, J.; Luo, Y. Antennal transcriptome analysis of the Asian longhorned beetle Anoplophora glabripennis. Sci. Rep. 2016, 6, 26652. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.N. Mechanisms of Odor Coding in Coniferous Bark Beetles: From Neuron to Behavior and Application. Psyche J. Entomol. 2012, 2012, 149572. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Gu, S.; Zhang, Y.; Guo, Y.; Wang, G. Candidate Olfaction Genes Identified within the Helicoverpa armigera Antennal Transcriptome. PLoS ONE 2012, 7, e48260. [Google Scholar] [CrossRef] [Green Version]
- Leal, W.S. Odorant Reception in Insects: Roles of Receptors, Binding Proteins, and Degrading Enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef]
- Nehme, M.E.; Keena, M.A.; Zhang, A.; Baker, T.C.; Xu, Z.; Hoover, K. Evaluating the Use of Male-Produced Pheromone Components and Plant Volatiles in Two Trap Designs to Monitor Anoplophora glabripennis. Environ. Entomol. 2010, 39, 169–176. [Google Scholar] [CrossRef]
- Tan, L.; Wang, W.; Li, C. Progress on integrated control of longhorn beetles in China. Hubei Agric. Sci. 2008, 47, 232–237. [Google Scholar]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef] [Green Version]
- Hull, J.J.; Perera, O.P.; Snodgrass, G.L. Cloning and expression profiling of odorant-binding proteins in the tarnished plant bug, Lygus lineolaris. Insect Mol. Biol. 2013, 23, 78–97. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, O.; Brillada, C.; Yazawa, S.; Maffei, M.E.; Arimura, G.I. Transcriptome pyrosequencing of the parasitoid wasp Cotesia vestalis: Genes involved in the antennal odorant-sensory system. PLoS ONE 2012, 7, e50664. [Google Scholar] [CrossRef] [Green Version]
- Davidson, N.M.; Oshlack, A. Corset: Enabling differential gene expression analysis for de novo assembled transcriptomes. Genome Biol. 2014, 15, 410. [Google Scholar]
- Yuan, H.B.; Ding, Y.X.; Gu, S.H.; Sun, L.; Zhu, X.Q.; Liu, H.W.; Dhiloo, K.H.; Zhang, Y.J.; Guo, Y.Y. Molecular characterization and expression profiling of odorant-binding proteins in Apolygus lucorum. PLoS ONE 2015, 10, e0140562. [Google Scholar]
- Xu, Y.-L.; He, P.; Zhang, L.; Fang, S.Q.; Dong, S.-L.; Zhang, Y.J.; Li, F. Large-scale identification of odorant-binding proteins and chemosensory proteins from expressed sequence tags in insects. BMC Genom. 2009, 10, 632. [Google Scholar] [CrossRef] [Green Version]
- Grosse-Wilde, E.; Kuebler, L.S.; Bucks, S.; Vogel, H.; Wicher, D.; Hansson, B. Antennal transcriptome of Manduca sexta. Proc. Natl. Acad. Sci. USA 2011, 108, 7449–7454. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Q.; Chen, D.B.; Liu, H.H.; Hu, H.L.; Bian, H.X.; Zhang, R.S.; Yang, R.S.; Jiang, X.F.; Shi, S.L. The complete mitochondrial genome of the longhorn beetle Dorysthenes paradoxus (Coleoptera: Cerambycidae: Prionini) and the implication for the phylogenetic relationships of the Cerambycidae species. J. Insect Sci. 2018, 18, 21. [Google Scholar] [CrossRef]
- Dodd, A. Beetle; Reaktion Books: London, UK, 2015. [Google Scholar]
- Aw, T.; Schlauch, K.; Keeling, C.I.; Young, S.; Bearfield, J.C.; Blomquist, G.J.; Tittiger, C. Functional genomics of mountain pine beetle (Dendroctonus ponderosae) midguts and fat bodies. BMC Genom. 2010, 11, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keeling, C.I.; Henderson, H.; Li, M.; Yuen, M.; Clark, E.L.; Fraser, J.D.; Huber, D.P.; Liao, N.Y.; Docking, T.R.; Birol, I. Transcriptome and full-length cDNA resources for the mountain pine beetle, Dendroctonus ponderosae Hopkins, a major insect pest of pine forests. Insect Biochem. Mol. Biol. 2012, 42, 525–536. [Google Scholar] [CrossRef]
- Vega, F.E.; Infante, F.; Johnson, A.J. The genus Hypothenemus, with emphasis on H. hampei, the coffee berry borer. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press Elsevier: London, UK, 2015; pp. 427–494. [Google Scholar] [CrossRef]
- Gu, X.-C.; Zhang, Y.-N.; Kang, K.; Dong, S.-L.; Zhang, L.-W. Antennal transcriptome analysis of odorant reception genes in the red turpentine beetle (RTB), Dendroctonus valens. PLoS ONE 2015, 10, e0125159. [Google Scholar] [CrossRef] [Green Version]
- Pelosi, P.; Zhou, J.-J.; Ban, L.; Calvello, M. Soluble proteins in insect chemical communication. Cell. Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef]
- Liu, R.; He, X.; Lehane, S.; Lehane, M.; Hertz-Fowler, C.; Berriman, M.; Field, L.; Zhou, J.-J. Expression of chemosensory proteins in the tsetse fly Glossina morsitans is related to female host-seeking behaviour. Insect Mol. Biol. 2011, 21, 41–48. [Google Scholar] [CrossRef] [Green Version]
- McKenzie, S.K.; Oxley, P.R.; Kronauer, D.J. Comparative genomics and transcriptomics in ants provide new insights into the evolution and function of odorant binding and chemosensory proteins. BMC Genom. 2014, 15, 718. [Google Scholar] [CrossRef] [Green Version]
- Manoharan, M.; Ng Fuk Chong, M.; Vaïtinadapoulé, A.; Frumence, E.; Sowdhamini, R.; Offmann, B. Comparative genomics of odorant binding proteins in Anopheles gambiae, Aedes aegypti, and Culex quinquefasciatus. Genome Biol. Evol. 2013, 5, 163–180. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Gracia, A.; Vieira, F.; Rozas, J. Molecular evolution of the major chemosensory gene families in insects. Heredity 2009, 103, 208–216. [Google Scholar] [CrossRef]
- Rihani, K.; Ferveur, J.-F.; Briand, L. The 40-year mystery of insect odorant-binding proteins. Biomolecules 2021, 11, 509. [Google Scholar] [CrossRef]
- Liu, Z.; Liang, X.-F.; Xu, L.; Keesey, I.W.; Lei, Z.-R.; Smagghe, G.; Wang, J.-J. An antennae-specific odorant-binding protein is involved in Bactrocera dorsalis olfaction. Front. Ecol. Evol. 2020, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Brito, N.F.; Moreira, M.F.; Melo, A.C. A look inside odorant-binding proteins in insect chemoreception. J. Insect Physiol. 2016, 95, 51–65. [Google Scholar] [CrossRef]
- Sun, L.; Gu, S.-H.; Xiao, H.-J.; Zhou, J.-J.; Guo, Y.-Y.; Liu, Z.-W.; Zhang, Y.-J. The preferential binding of a sensory organ specific odorant binding protein of the alfalfa plant bug Adelphocoris lineolatus AlinOBP10 to biologically active host plant volatiles. J. Chem. Ecol. 2013, 39, 1221–1231. [Google Scholar] [CrossRef]
- Kim, M.-S.; Repp, A.; Smith, D.P. LUSH odorant-binding protein mediates chemosensory responses to alcohols in Drosophila melanogaster. Genetics 1998, 150, 711–721. [Google Scholar] [CrossRef]
- Mao, X.-Z.; Yang, B. EAG and behavioral responses of Cacia cretifera thibetana to seven walnut tree volatiles. Chin. J. Appl. Entomol. 2020, 57, 938. [Google Scholar]
- Campbell, C.; Gries, R.; Gries, G. Forty-two compounds in eleven essential oils elicit antennal responses from Aedes aegypti. Entomol. Exp. Et Appl. 2011, 138, 21–32. [Google Scholar] [CrossRef]
- Wicher, D. Olfactory Signaling in Insects. Molecular Basis of Olfaction; Academic Press Elsevier: New York, NY, USA, 2015; Volume 130, pp. 37–54. [Google Scholar] [CrossRef]
- Venthur, H.; Zhou, J.-J. Odorant receptors and odorant-binding proteins as insect pest control targets: A comparative analysis. Front. Physiol. 2018, 9, 1163. [Google Scholar] [CrossRef] [PubMed]
- Hieu, T.T.; Jung, J.; Kim, S.I.; Ahn, Y.J.; Kwon, H.W. Behavioural and electroantennogram responses of the stable fly (Stomoxys calcitrans L.) to plant essential oils and their mixtures with attractants. Pest Manag. Sci. 2014, 70, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Grilo, F.S.; Wang, S.C. Walnut (Juglans regia L.) volatile compounds indicate kernel and oil oxidation. Foods 2021, 10, 329. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No | Female Samples | Male Samples | ||
|---|---|---|---|---|
| Organ/Tissue | Abbreviations | Organ/Tissue | Abbreviations | |
| 1 | Female wings | FCTc | Male wings | MCTc |
| 2 | Female abdomen | FCTf | Male Abdomen | MCTf |
| 3 | Female legs | FCTl | Male legs | MCT1 |
| 4 | Female head | FCTt | Male head | MCTt |
| 5 | Female antennae | FCTte | Male antennae | MCTte |
| 6 | Female thorax | FCTx | Male thorax | MCTx |
| GO Accession | Description | Term Type | Over Represented p-Value | Corrected p-Value | DEG Item | DEG List |
|---|---|---|---|---|---|---|
| GO:0055114 | oxidation reduction process | Biological Process | 1.2268 × 10−15 | 8.0529 × 10−12 | 730 | 6448 |
| GO:0016491 | Oxidoreductase activity | Molecular Function | 1.1892 × 10−14 | 3.903 × 10−11 | 702 | 6448 |
| GO:0016705 | oxidoreductase activity, acting on paired donors, oxidation or reduction of molecular oxygen | Molecular Function | 1.7425 × 10−13 | 3.8127 × 10−10 | 145 | 6448 |
| GO:0020037 | heme binding | Molecular Function | 3.234 × 10−13 | 5.307 × 10−10 | 156 | 6448 |
| Term | ID | Sample Number | Background Number | p-Value |
|---|---|---|---|---|
| Cutin, suberine and wax biosynthesis | ko00073 | 26 | 40 | 0.0046 |
| Glycerophospholipid metabolism | ko00564 | 70 | 152 | 0.0086 |
| Choline metabolism in cancer | ko05231 | 48 | 98 | 0.0119 |
| Chemokine signaling pathway | ko04062 | 62 | 134 | 0.0119 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, N.; Mao, X.; Liu, N.; Liu, L.; Zhang, Z.; Ze, S.; Yang, B. Transcriptomic Characterization of Odorant Binding Proteins in Cacia cretifera thibetana and Their Association with Different Host Emitted Volatiles. Insects 2021, 12, 787. https://doi.org/10.3390/insects12090787
Zhao N, Mao X, Liu N, Liu L, Zhang Z, Ze S, Yang B. Transcriptomic Characterization of Odorant Binding Proteins in Cacia cretifera thibetana and Their Association with Different Host Emitted Volatiles. Insects. 2021; 12(9):787. https://doi.org/10.3390/insects12090787
Chicago/Turabian StyleZhao, Ning, Xiangzhong Mao, Naiyong Liu, Ling Liu, Zhixiao Zhang, Sangzi Ze, and Bin Yang. 2021. "Transcriptomic Characterization of Odorant Binding Proteins in Cacia cretifera thibetana and Their Association with Different Host Emitted Volatiles" Insects 12, no. 9: 787. https://doi.org/10.3390/insects12090787
APA StyleZhao, N., Mao, X., Liu, N., Liu, L., Zhang, Z., Ze, S., & Yang, B. (2021). Transcriptomic Characterization of Odorant Binding Proteins in Cacia cretifera thibetana and Their Association with Different Host Emitted Volatiles. Insects, 12(9), 787. https://doi.org/10.3390/insects12090787