


Higher Essential Amino Acid and Crude Protein Contents in Pollen Accelerate the Oviposition and Colony Foundation of Bombus breviceps (Hymenoptera: Apidae)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Queen Collection, Rearing and Colony Management
2.2. Pollen Selection and Treatments
2.3. Palynological Analysis
2.4. Determination of Pollen Nutritional Composition
2.5. Observation of Queenright Colony Foundation
2.6. Statistics and Analysis
3. Results
3.1. Palynological Analysis
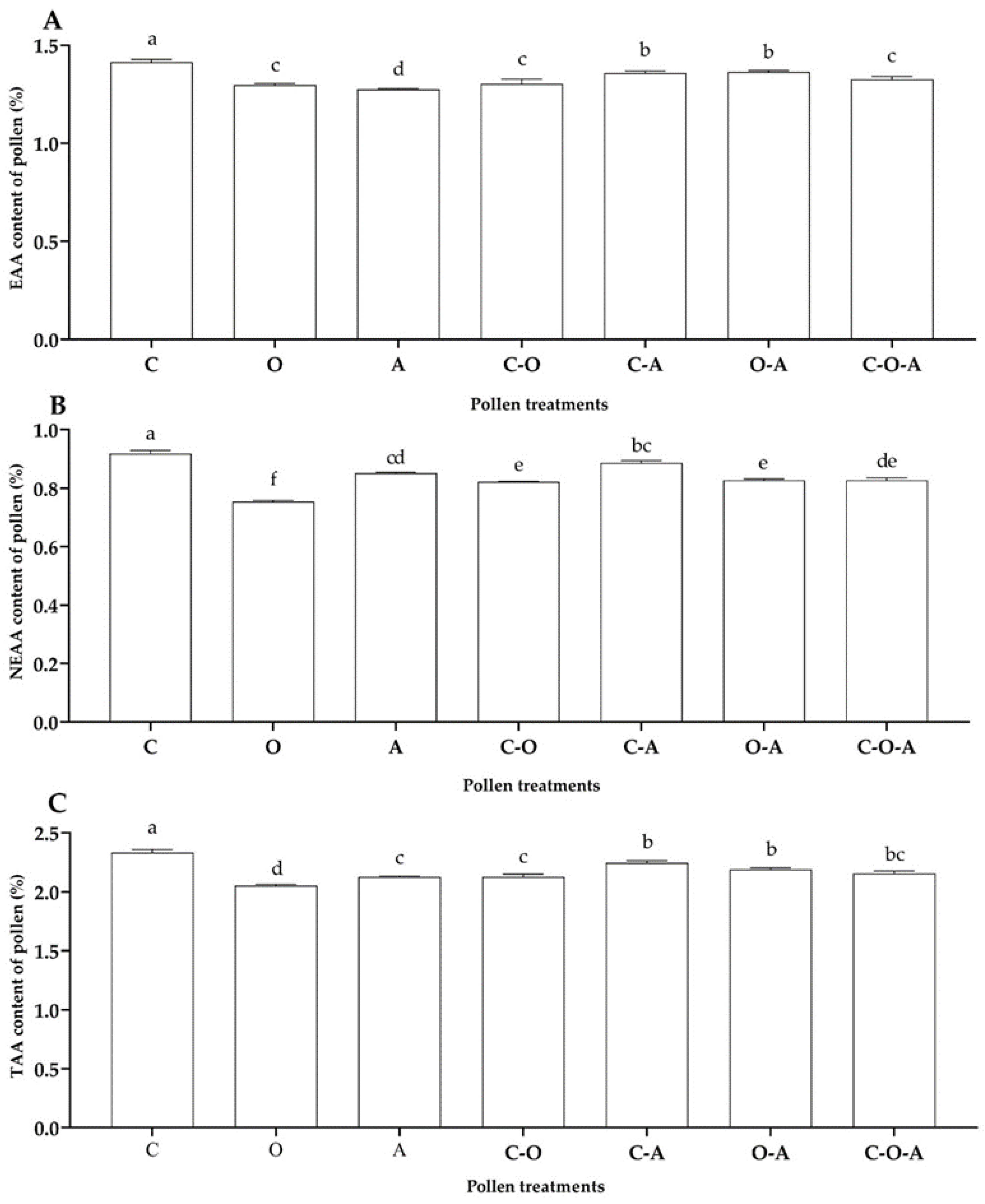
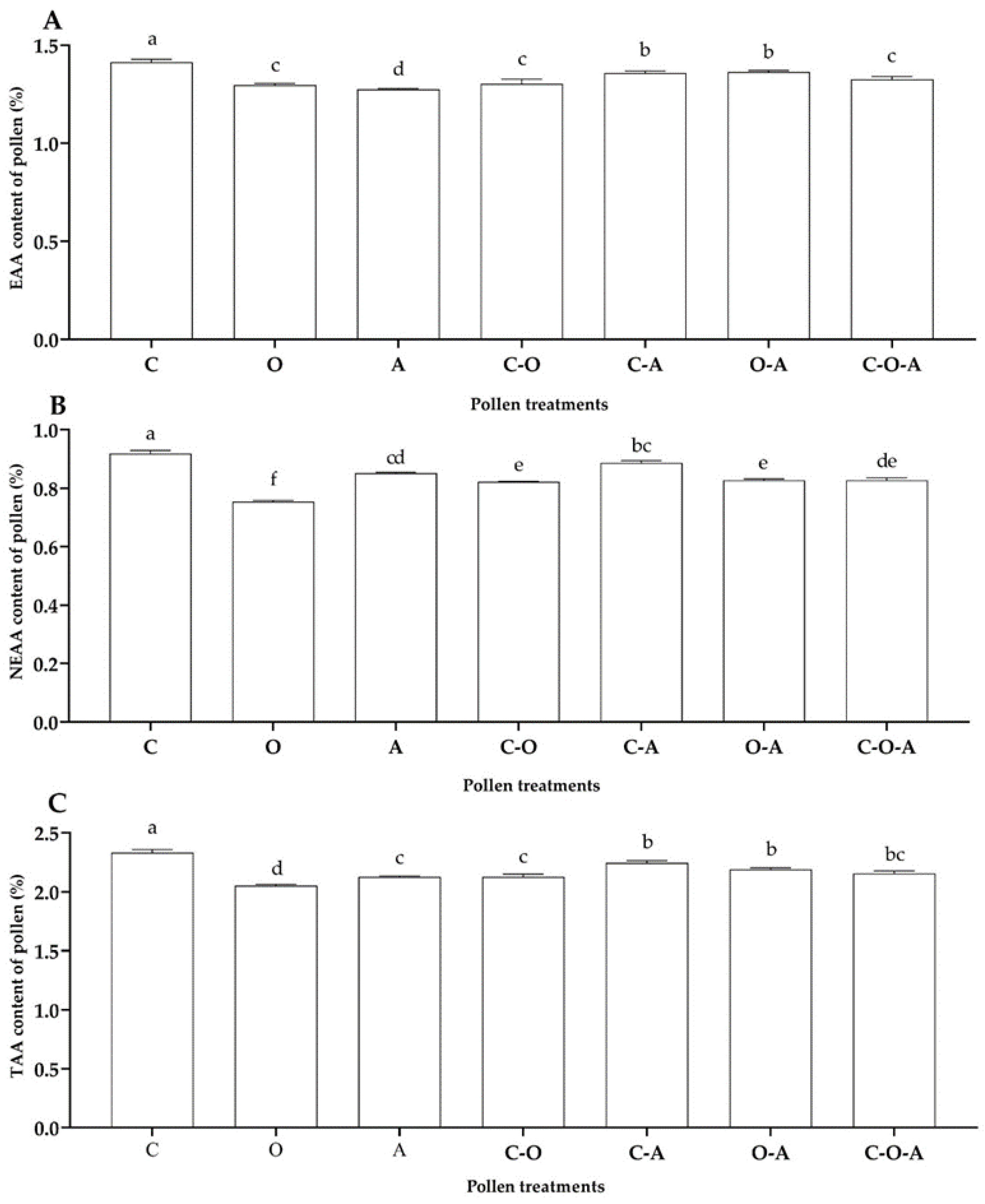
3.2. Determination of Pollen Nutritional Composition
3.3. Observation of Queenright Colony Development
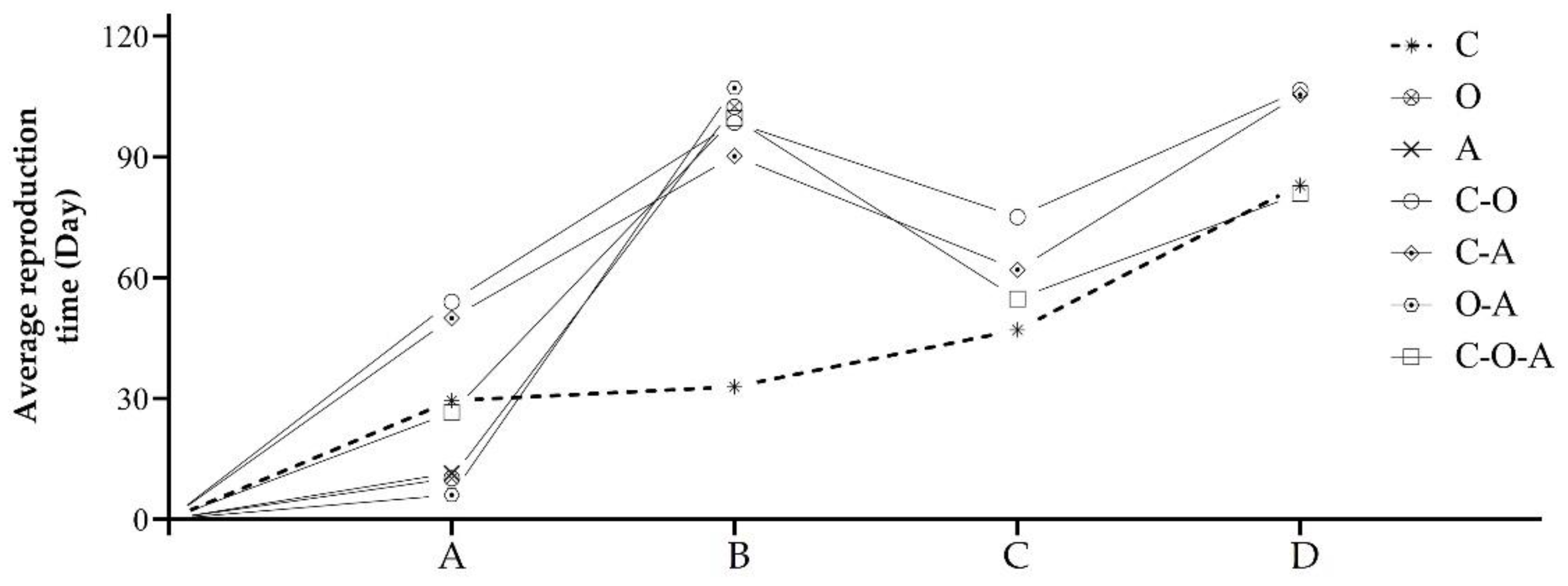
3.3.1. Performance and Reproductive Time
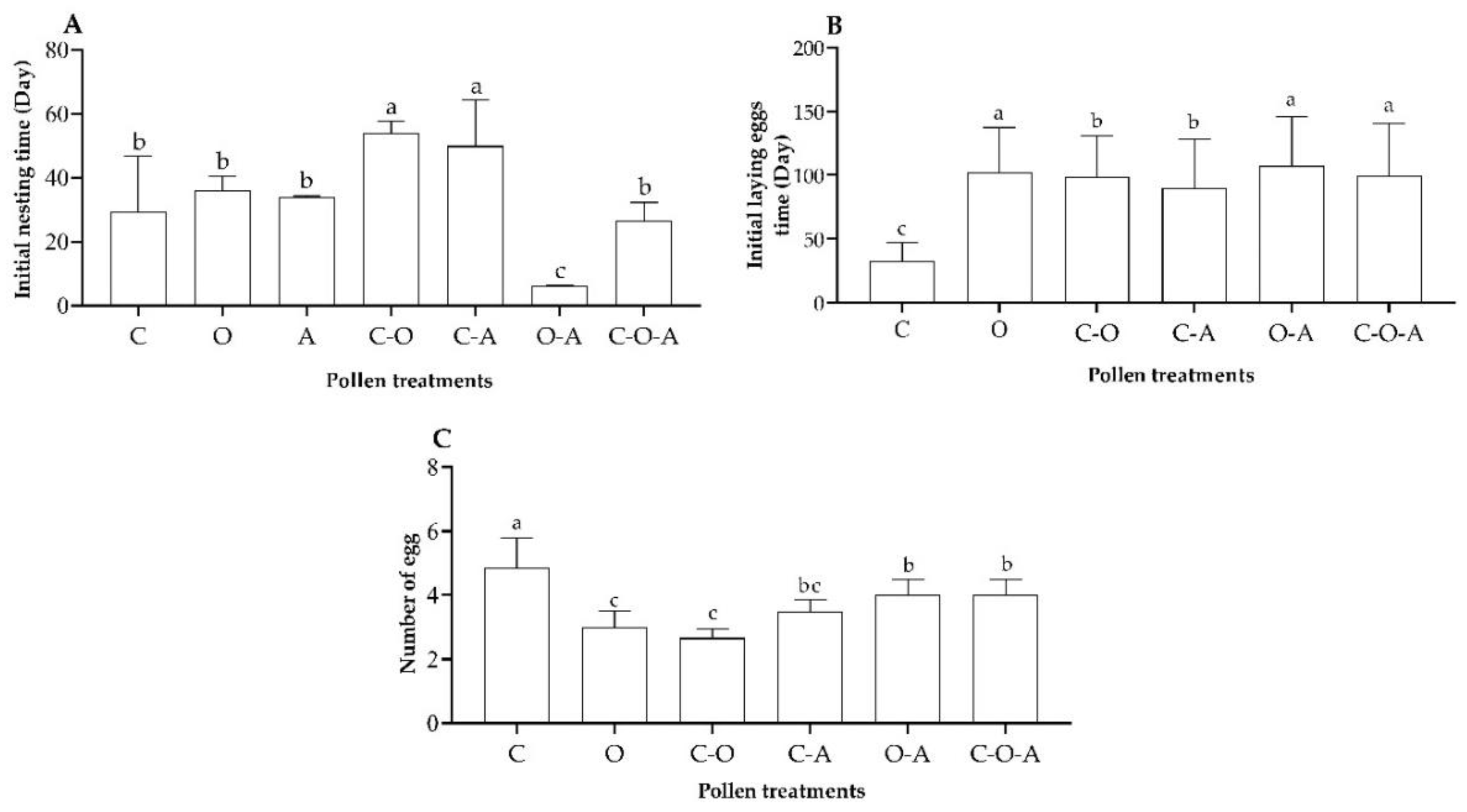
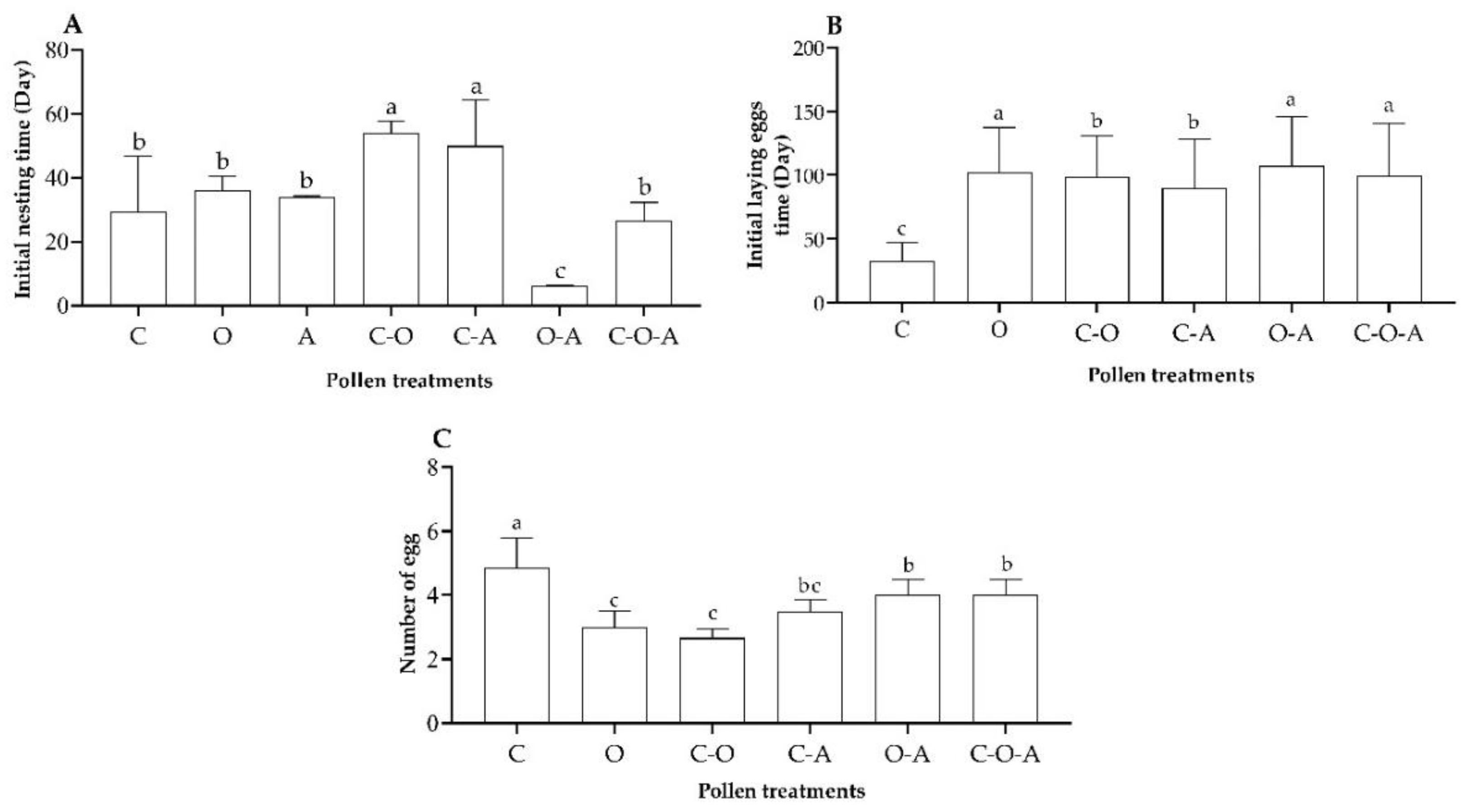
3.3.2. Nest Building and Egg Laying
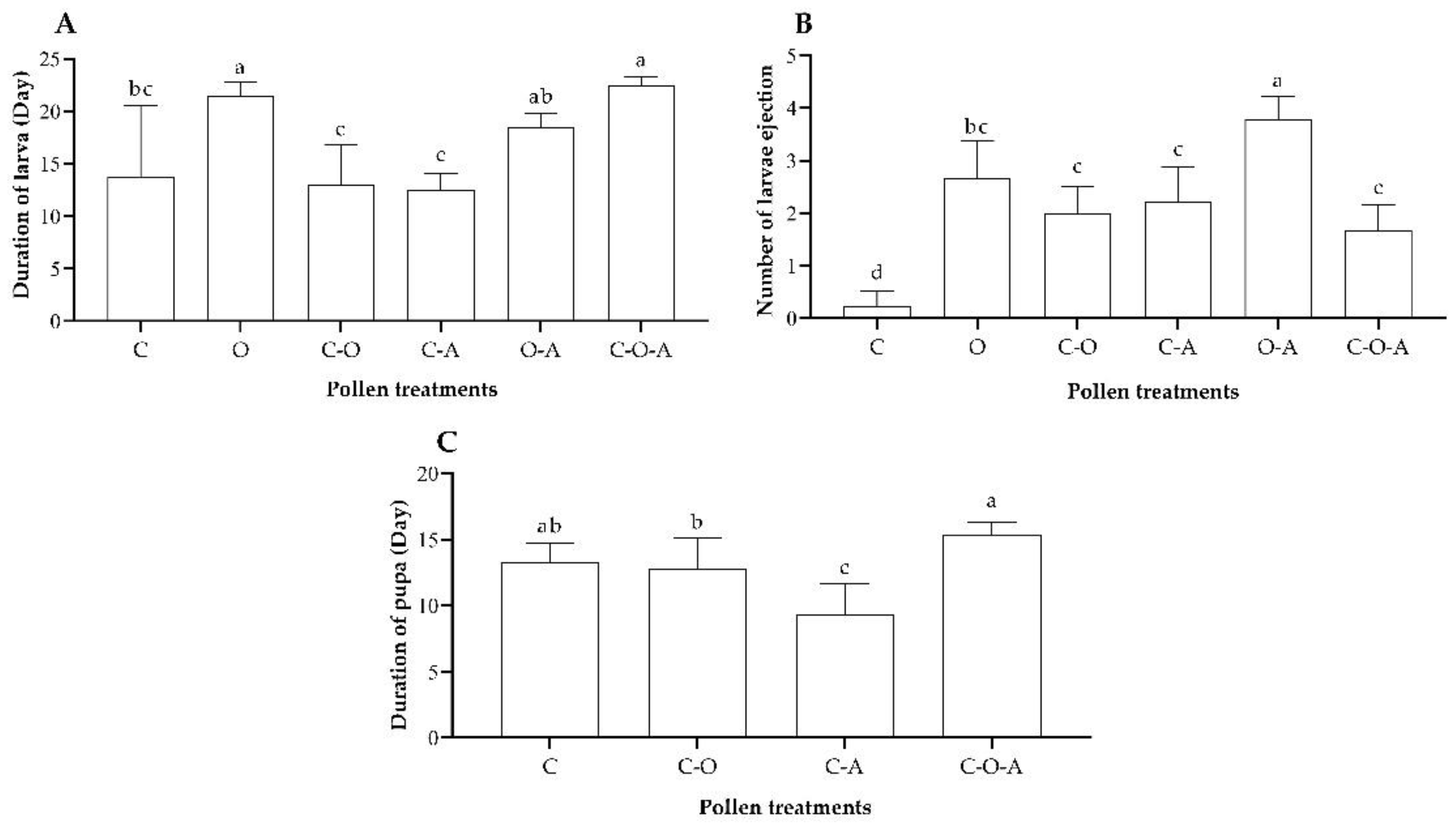
3.3.3. Development of Larvae and Pupae
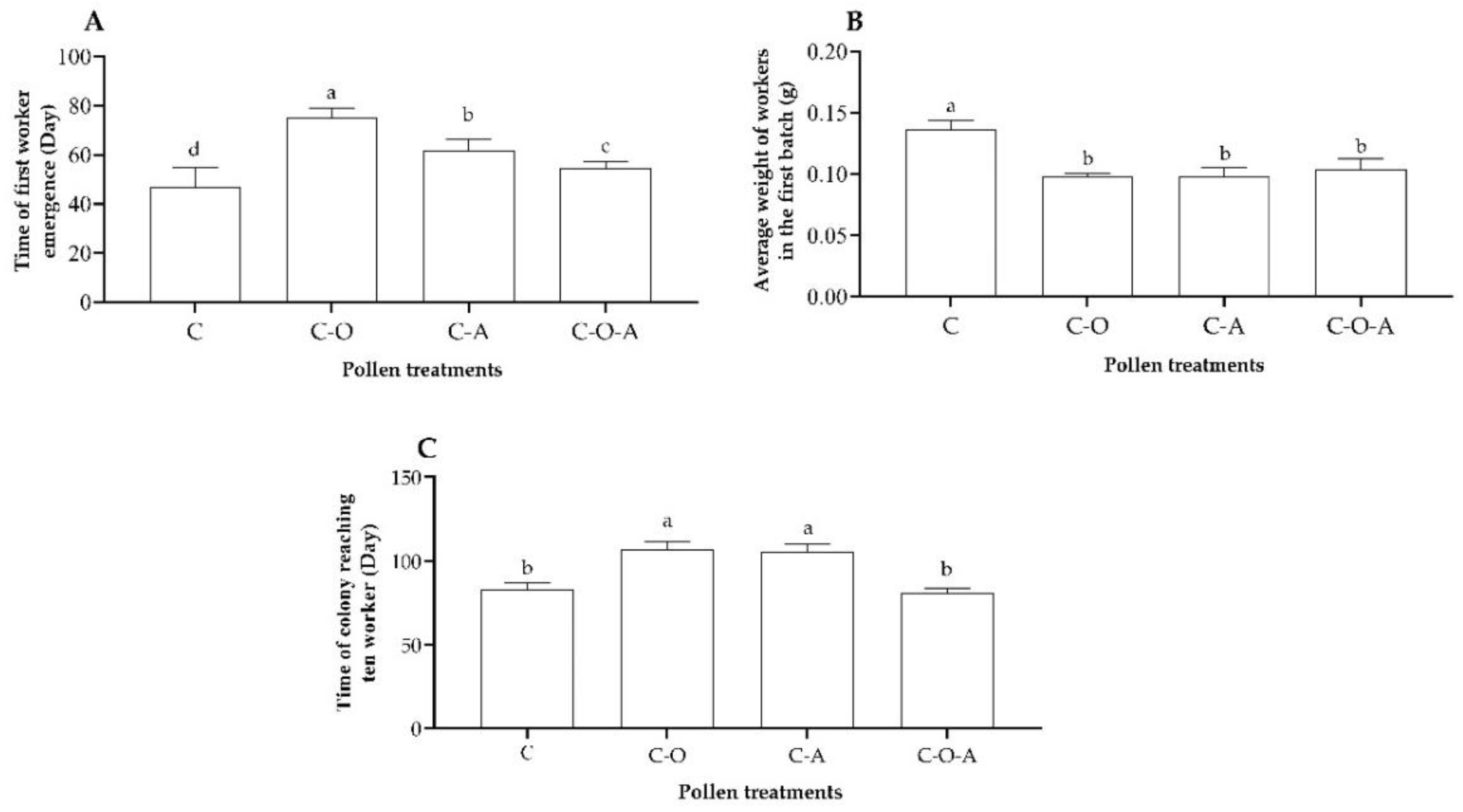
3.3.4. Development of Workers in Colony Foundation Stage
3.4. Analysis of the Relationship between Pollen Nutrition and Reproduction Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Velthuis, H.; Doorn, A. A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie 2006, 37, 421–451. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and conservation of bumble bees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M.M.; Tucker, E.M.; Mathiasson, M.E.; Rehan, S.M. Decline of bumble bees in northeastern North America, with special focus on Bombus terricola. Biol. Conserv. 2018, 217, 437–445. [Google Scholar] [CrossRef]
- Rasmont, P.; Coppée, A.; Michez, D.; De Meulemeester, T. An overview of the Bombus terrestris (L. 1758) subspecies (Hymenoptera: Apidae). Ann. Société Entomol. Fr. 2008, 44, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.; Brown, M. Parasites and genetic diversity in an invasive bumble bee. J. Anim. Ecol. 2014, 83, 1428–1440. [Google Scholar] [CrossRef] [Green Version]
- Schmid-Hempel, R.; Eckhardt, M.; Goulson, D.; Heinzmann, D.; Lange, C.; Plischuk, S.; Escudero, L.R.; Salathé, R.; Scriven, J.J.; Schmid-Hempel, P. The invasion of southern South America by imported bumblebees and associated parasites. J. Anim. Ecol. 2014, 83, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, C.; Yokoyama, J.; Washitani, I. Invasion status and potential ecological impacts of an invasive alien bumblebee, Bombus terrestris L. (Hymenoptera: Apidae) naturalized in southern Hokkaido, Japan. Glob. Environ. Res. 2003, 8, 51–66. [Google Scholar]
- Sáez, A.; Morales, C.L.; Garibaldi, L.A.; Aizen, M.A. Invasive bumble bees reduce nectar availability for honey bees by robbing raspberry flower buds. Basic Appl. Ecol. 2017, 19, 26–35. [Google Scholar] [CrossRef]
- Dohzono, I.; Kunitake, Y.K.; Yokoyama, J.; Goka, K. Alien bumble bee affects native plant reproduction through interactions with native bumble bees. Ecology 2008, 89, 3082–3092. [Google Scholar] [CrossRef]
- Kanbe, Y.; Okada, I.; Yoneda, M.; Goka, K.; Tsuchida, K. Interspecific mating of the introduced bumblebee Bombus terrestris and the native Japanese bumblebee Bombus hypocrita sapporoensis results in inviable hybrids. Naturwissenschaften 2008, 95, 1003–1008. [Google Scholar] [CrossRef]
- Tsuchida, K.; Yamaguchi, A.; Kanbe, Y.; Goka, K. Reproductive Interference in an Introduced Bumblebee: Polyandry may Mitigate Negative Reproductive Impact. Insects 2019, 10, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kingston, A.; Marsden-Smedley, J.; Driscoll, D.; Corbett, S.; Fenton, J.; Anderson, R.; Plowman, C.; Mowling, F.; Jenkin, M.; Matsui, K.; et al. Extent of invasion of Tasmanian native vegetation by the exotic bumblebee Bombus terrestris (Apoidea: Apidae). Austral Ecol. 2002, 27, 162–172. [Google Scholar] [CrossRef]
- Inoue, M.N.; Yokoyama, J.; Washitani, I. Displacement of Japanese native bumblebees by the recently introduced Bombus terrestris (L.) (Hymenoptera: Apidae). J. Insect Conserv. 2008, 12, 135–146. [Google Scholar] [CrossRef]
- Tsuchida, K.; Kondo, N.; Inoue, M.; Goka, K. Reproductive disturbance risks to indigenous Japanese bumblebees from introduced Bombus terrestris. Appl. Entomol. Zool. 2010, 45, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Macfarlane, R.P.; Patten, K.D.; Royce, L.A.; Wyatt, B.; Mayer, D.F. Management potential of sixteen North American bumble bee species. Melanderia 1994, 50, 1–12. [Google Scholar]
- Mah, Y.I.; Lee, M.Y.; Bilinski, M. Some charateristics of Korean indigenous bumblebee species (Hymenoptera; Bombus spp.) under laboratory conditions. Acta Hortic. 2001, 561, 287–291. [Google Scholar] [CrossRef]
- Ono, M.; Mitsuhata, M.; Sasaki, M. Use of Introduced Bombus terrestris Worker Helpers for rapid development of Japanese native B. hypocrita colonies (Hymenoptera, Apidae). Appl. Entomol. Zool. 1994, 29, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Maeta, Y.; Hoshikawa, K. Colony development of two species of Japanese bumblebees Bombus (Bombus) ignitus and Bombus (Bombus) hypocrita reared under artificial condition (Hymenoptera, Apidae). Jpn. J. Entomol. 1997, 65, 343–354. [Google Scholar]
- Asada, S.; Ono, M. Difference in colony development of two Japanese bumblebees, Bombus hypocrita and B. ignitus (Hymenoptera: Apidae). Appl. Entomol. Zool. 2000, 35, 597–603. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhou, Z.Y.; Huang, J.X.; Yuan, X.L.; Ding, G.L.; An, J.D. Queen traits and colony size of four bumblebee species of China. Insectes Sociaux 2018, 65, 537–547. [Google Scholar] [CrossRef]
- An, J.D.; Huang, J.X.; Shao, Y.Q.; Zhang, S.W.; Wang, B.; Liu, X.Y.; Wu, J.; Williams, P.H. The bumblebees of North China (Apidae, Bombus Latreille). Zootaxa 2014, 3830, 1–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.X.; An, J.D. Species diversity, pollination application and strategy for conservation of the bumblebees of China. Biodivers. Sci. 2018, 26, 486–497. [Google Scholar] [CrossRef]
- Liang, C.; Zhang, X.W.; Huang, J.X.; Song, W.F.; Zhang, H.; Luo, W.T.; An, J.D. Biogeography and species diversity of bumblebees in Yunnan, Southwest China. Chin. J. Appl. Entomol. 2018, 55, 1045–1053. [Google Scholar]
- Williams, P.H.; Tang, Y.; Yao, J.W.; Cameron, S.A. The bumblebees of Sichuan (Hymenoptera: Apidae, Bombini). Syst. Biodivers. Conserv. 2009, 7, 101–189. [Google Scholar] [CrossRef] [Green Version]
- Deka, T.N.; Gudade, B.; Saju, K.A.; Vijayan, A.; Chhetri, P. Foraging behaviour of Bumble bee (Bombus breviceps Smith) in large cardamom (Amomum subulatum Roxb.) at Dzongu, North Sikkim. Ecol. Environ. Conserv. 2014, 20, 1593–1595. [Google Scholar]
- Deka, T.N.; Sudharshan, M.R.; Saju, K.A. New record of bumble bee, Bombus breviceps Smith as a pollinator of large cardamom. Curr. Sci. 2011, 100, 926–928. [Google Scholar]
- Liang, C.; Ding, G.L.; Huang, J.X.; Zhang, X.W.; Miao, C.H.; An, J.D. Characteristics of the two Asian bumblebee species Bombus friseanus and Bombus breviceps (Hymenoptera: Apidae). Insects 2020, 11, 163. [Google Scholar] [CrossRef] [Green Version]
- Roulston, T.H.; Cane, J.H. Pollen nutritional content and digestibility for animals. In Pollen and Pollination; Springer: Vienna, Austria, 2000; pp. 187–209. [Google Scholar] [CrossRef]
- Tanaka, C.S.; Ikemoto, M.; Nikkeshi, A.; Kanbe, Y.; Mitsuhata, M.; Yokoi, T. Ovarian development related to pollen feeding in workers of the bumblebee Bombus ignitus (Hymenoptera: Apidae). Appl. Entomol. Zool. 2018, 54, 85–89. [Google Scholar] [CrossRef]
- Free, J.B. Bumblebee economics. Nature 1979, 280, 850. [Google Scholar] [CrossRef]
- Moerman, R.; Vanderplanck, M.; Roger, N.; Decleves, S.; Wathelet, B.; Rasmont, P.; Fournier, D.; Michez, D. Growth rate of bumblebee larvae is related to pollen amino acids. J. Econ. Entomol. 2016, 109, 25–30. [Google Scholar] [CrossRef]
- Ryder, J.T.; Cherrill, A.; Thompson, H.M.; Walters, K.F.A. Lower pollen nutritional quality delays nest building and egg laying in Bombus terrestris audax micro-colonies leading to reduced biomass gain. Apidologie 2021, 52, 1033–1047. [Google Scholar] [CrossRef] [PubMed]
- Baloglu, G.H.; Gurel, F. The effects of pollen protein content on colony development of the bumblebee, Bombus terrestris L. J. Apic. Sci. 2015, 59, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Regali, A.; Rasmont, P. Nouvelles méthodes de test pour l’évaluation du régime alimentaire chez des colonies orphelines de Bombus terrestris (L) (Hymenoptera, Apidae). Apidologie 1995, 26, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Wynants, E.; Wackers, F.; Van Oystaeyen, A. Re-evaluation of a method used to study nutritional effects on bumble bees. Ecol. Entomol. 2022, 47, 959–966. [Google Scholar] [CrossRef]
- Vaudo, A.D.; Patch, H.M.; Mortensen, D.A.; Tooker, J.F.; Grozinger, C.M. Macronutrient ratios in pollen shape bumble bee (Bombus impatiens) foraging strategies and floral preferences. Proc. Natl. Acad. Sci. USA 2016, 113, 4035–4042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roger, N.; Michez, D.; Wattiez, R.; Sheridan, C.; Vanderplanck, M. Diet effects on bumblebee health. J. Insect Physiol. 2017, 96, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.M.; Sui, R.; Song, W.F.; Ma, Y.J.; Liang, C.; Dong, K. Effects of sugar syrups on worker colony development of the bumblebee. Apic. China 2020, 71, 4. [Google Scholar]
- Qin, J.M. Colony reproductive characteristics of Bombus breviceps in Yunnan of China. J. Environ. Entomol. 2022, 44, 440–445. [Google Scholar] [CrossRef]
- Tasei, J.N.; Aupinel, P. Nutritive value of 15 single pollens and pollen mixes tested on larvae produced by bumblebee workers (Bombus terrestris, Hymenoptera: Apidae). Apidologie 2008, 39, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Tasei, J.N.; Aupinel, P. Validation of a method using queenless Bombus terrestris micro-colonies for testing the nutritive value of commercial pollen mixes by comparison with queenright colonies. J. Econ. Entomol. 2008, 101, 1737–1742. [Google Scholar] [CrossRef]
- Vaudo, A.D.; Stabler, D.; Patch, H.M.; Tooker, J.F.; Grozinger, C.M.; Wright, G.A. Bumble bees regulate their intake of essential protein and lipid pollen macronutrients. J. Exp. Biol. 2016, 219, 3962–3970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treanore, E.D.; Vaudo, A.D.; Grozinger, C.M.; Fleischer, S.J. Examining the nutritional value and effects of different floral resources in pumpkin agroecosystems on Bombus impatiens worker physiology. Apidologie 2019, 50, 542–552. [Google Scholar] [CrossRef]
- Cabrera, A.R.; Almanza, M.T.; Cutler, G.C.; Fischer, D.L.; Hinarejos, S.; Lewis, G.; Nigro, D.; Olmstead, A.; Overmyer, J.; Potter, D.A.; et al. Initial recommendations for higher-tier risk assessment protocols for bumble bees, Bombus spp. (Hymenoptera: Apidae). Integr. Environ. Assess. Manag. 2016, 12, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Klinger, E.G.; Camp, A.A.; Strange, J.P.; Cox-Foster, D.; Lehmann, D.M. Bombus (Hymenoptera: Apidae) microcolonies as a tool for biological understanding and pesticide risk assessment. Environ. Entomol. 2019, 48, 1249–1259. [Google Scholar] [CrossRef] [PubMed]
- Mommaerts, V.; Reynders, S.; Boulet, J.; Besard, L.; Sterk, G.; Smagghe, G. Risk assessment for side-effects of neonicotinoids against bumblebees with and without impairing foraging behavior. Ecotoxicology 2010, 19, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Van Oystaeyen, A.; Klatt, B.K.; Petit, C.; Lenaerts, N.; Wackers, F. Short-term lab assessments and microcolonies are insufficient for the risk assessment of insecticides for bees. Chemosphere 2021, 273, 128518. [Google Scholar] [CrossRef]
- GÜRel, F.; GalİÇ, A.; GÖSterİT, A. The effect of queen removal on sexual production in the bumble bee, Bombus terrestris (Hymenoptera: Apidae). Turk. J. Zool. 2009, 33, 4. [Google Scholar] [CrossRef]
- Smyth, A.P.; Seales, B.; Bradley, P.M. A pollen profile by scanning electron microscopy bracketing the mid-Holocene Tsuga canadensis decline at Poutwater Pond Bog, Holden, Massachusetts. Grana 2015, 54, 263–270. [Google Scholar] [CrossRef]
- Zavada, M.; Wei, Z.X. A Contribution to the Pollen Morphology of Camellia (Theaceae). Grana 1993, 32, 233–242. [Google Scholar] [CrossRef] [Green Version]
- Li, W.W.; Wang, Y.N.; Liu, L.Q.; Niu, Y.Y.; Zhao, S.R.; Zhang, S.K.; Wang, Y.T.; Liao, K. Pollen morphology of selected apricot (Prunus) taxa. Palynology 2021, 45, 95–102. [Google Scholar] [CrossRef]
- Ma, H.B.; Zhang, P.Q.; Wang, J.X.; Zhang, H.; Zhang, Z.H.; Zhang, Y.C.; Ning, Y.W. Preparation of a novel rape pollen shell microencapsulation and its use for protein adsorption and pH-controlled release. J. Microencapsul. 2014, 31, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Van Handel, E.; Day, J.F. Assay of lipids, glycogen and sugars in individual mosquitoes: Correlations with wing length in field-collected Aedes vexans. J. Am. Mosq. Control Assoc. 1988, 4, 549–550. [Google Scholar] [PubMed]
- de Groot, A.P. Protein and amino acid requirements of the honeybee (Apis mellifica L.). Physiol. Comp. Oecol. 1953, 3, 1–83. [Google Scholar]
- Day, S.; Beyer, R.; Mercer, A.; Ogden, S. The Nutrient Composition of Honeybee-Collected Pollen in Otago, New Zealand. J. Apic. Res. 1990, 29, 138–146. [Google Scholar] [CrossRef]
- Herbert, E.W. The Hive and the Honey Bee. In Honey Bee Nutrition; Graham, J.M., Ed.; Dadant & Sons: Hamilton, NY, USA, 1992; pp. 197–233. [Google Scholar]
- Vaudo, A.D.; Tooker, J.F.; Patch, H.M.; Biddinger, D.J.; Coccia, M.; Crone, M.K.; Fiely, M.; Francis, J.S.; Hines, H.M.; Hodges, M.; et al. Pollen Protein: Lipid Macronutrient Ratios May Guide Broad Patterns of Bee Species Floral Preferences. Insects 2020, 11, 132. [Google Scholar] [CrossRef] [Green Version]
- Mccaughey, W.F.; Gilliam, M.; Standifer, L.N. Amino acids and protein adequacy for honey bees of pollens from desert plants and other floral sources. Apidologie 1980, 11, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, J.O.; Thoenes, S.C.; Levin, M.D. Survival of Honey Bees, Apis mellifera (Hymenoptera: Apidae), Fed Various Pollen Sources. Ann. Entomol. Soc. Am. 1987, 80, 176–183. [Google Scholar] [CrossRef]
- Loper, G.M.; Cohen, A.C. Amino acid content of dandelion pollen, a honey bee (Hymenoptera: Apidae) nutritional evaluation. J. Econ. Entomol. 1987, 80, 14–17. [Google Scholar] [CrossRef]
- Moerman, R.; Vanderplanck, M.; Fournier, D.; Jacquemart, A.L.; Michez, D. Pollen nutrients better explain bumblebee colony development than pollen diversity. Insect Conserv. Divers. 2017, 10, 171–179. [Google Scholar] [CrossRef]
- Cook, S.M.; Awmack, C.S.; Murray, D.A.; Williams, I.H. Are honey bees’ foraging preferences affected by pollen amino acid composition? Ecol. Entomol. 2003, 28, 622–627. [Google Scholar] [CrossRef]
- Leonhardt, S.D.; Blüthgen, N. The same, but different: Pollen foraging in honeybee and bumblebee colonies. Apidologie 2012, 43, 449–464. [Google Scholar] [CrossRef] [Green Version]
- Nation, J.L. Insect Physiology and Biochemistry, 1st ed.; CRC Press: Boca Raton, FL, USA, 2002; Volume 22, p. 159. [Google Scholar] [CrossRef]
- Behmer, S.T.; Nes, W.D. Insect sterol nutrition and physiology: A global overview. Adv. Insect Physiol. 2003, 31, 1–72. [Google Scholar] [CrossRef]
- Feldlaufer, M.F.; Svoboda, J.A.; Herbert, E.W. Makisterone A and 24-methylenecholesterol from the ovaries of the honey bee, Apis mellifera L. Experientia 1986, 42, 200–201. [Google Scholar] [CrossRef]
- Vanderplanck, M.; Leroy, B.; Wathelet, B.; Wattiez, R.; Michez, D. Standardized protocol to evaluate pollen polypeptides as bee food source. Apidologie 2014, 45, 192–204. [Google Scholar] [CrossRef] [Green Version]
- Ruedenauer, F.A.; Raubenheimer, D.; Kessner-Beierlein, D.; Grund-Mueller, N.; Noack, L.; Spaethe, J.; Leonhardt, S.D. Best be(e) on low fat: Linking nutrient perception, regulation and fitness. Ecol. Lett. 2020, 23, 545–554. [Google Scholar] [CrossRef] [Green Version]
- McAulay, M.K.; Forrest, J.R.K. How do sunflower pollen mixtures affect survival of queenless microcolonies of bumblebees (Bombus impatiens)? Arthropod-Plant Interact. 2019, 13, 517–529. [Google Scholar] [CrossRef]
- Ribeiro, M.F.; Duchateau, M.J.; Velthuis, H.H.W. Comparison of the effects of two kinds of commercially available pollen on colony development and queen production in the bumble bee Bombus terrestris L. (Hymenoptera, Apidae). Apidologie 1996, 27, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Arien, Y.; Dag, A.; Zarchin, S.; Masci, T.; Shafir, S. Omega-3 deficiency impairs honey bee learning. Proc. Natl. Acad. Sci. USA 2015, 112, 15761–15766. [Google Scholar] [CrossRef] [Green Version]
- Muth, F.; Francis, J.S.; Leonard, A.S. Bees use the taste of pollen to determine which flowers to visit. Biol. Lett. 2016, 12, 20160356. [Google Scholar] [CrossRef] [Green Version]
- Human, H.; Nicolson, S.W.; Strauss, K.; Pirk, C.W.; Dietemann, V. Influence of pollen quality on ovarian development in honeybee workers (Apis mellifera scutellata). J. Insect Physiol. 2007, 53, 649–655. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pollen Treatments | Brassica sp. (%) | Camellia sp. (%) | Armeniaca sp. (%) | Seriphidium sp. (%) | Cirsium sp. (%) | Lactuca sp. (%) | Acer sp. (%) | Salix sp. (%) | Others (%) |
|---|---|---|---|---|---|---|---|---|---|
| C | 3.32 | 91.16 | 1.32 | - | - | 1.05 | 1.05 | - | 2.11 |
| O | 75.16 | - | 10.16 | 4.15 | 0.71 | - | - | 9.10 | 0.71 |
| A | 31.48 | - | 66.49 | 1.35 | - | - | - | - | 0.67 |
| C–O | 47.13 | 40.20 | 7.44 | 3.21 | - | 1.01- | - | - | 1.01 |
| C–A | 19.29 | 44.36 | 34.25 | - | - | - | - | - | 2.10 |
| O–A | 52.47 | - | 41.39 | 4.91 | - | - | - | - | 1.23 |
| C–O–A | 38.54 | 24.27 | 22.10 | 6.06 | - | 1.01 | - | 7.01 | 1.01 |
| Pollen Treatments | Crude Protein (%) | Lipid (%) | Carbohydrate (%) | P: L | C: L | C: P |
|---|---|---|---|---|---|---|
| C | 25.00 ± 0.06 abc | 2.49 ± 0.19 e | 25.99 ± 4.29 d | 10.01 | 10.40 | 1.04 |
| O | 26.18 ± 2.03 a | 5.58 ± 1.61 a | 46.44 ± 11.97 a | 4.69 | 8.32 | 1.77 |
| A | 24.67 ± 1.07 bc | 4.48 ± 0.05 c | 46.24 ± 6.23 a | 5.51 | 10.33 | 1.87 |
| C–O | 24.76 ± 2.45 bc | 2.23 ± 0.02 e | 36.56 ± 4.77 b | 11.13 | 16.43 | 1.48 |
| C–A | 23.93 ± 3.71 c | 3.55 ± 0.24 d | 31.62 ± 5.95 c | 6.74 | 8.91 | 1.32 |
| O–A | 24.97 ± 1.09 bc | 2.20 ± 0.47 e | 47.22 ± 3.24 a | 11.35 | 21.46 | 1.89 |
| C–O–A | 25.70 ± 1.18 ab | 5.03 ± 0.05 b | 45.12 ± 11.25 a | 5.11 | 8.97 | 1.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, C.-S.; Chang, Z.-M.; Han, L.; Chen, X.-S.; Long, J.-K. Higher Essential Amino Acid and Crude Protein Contents in Pollen Accelerate the Oviposition and Colony Foundation of Bombus breviceps (Hymenoptera: Apidae). Insects 2023, 14, 203. https://doi.org/10.3390/insects14020203
Ren C-S, Chang Z-M, Han L, Chen X-S, Long J-K. Higher Essential Amino Acid and Crude Protein Contents in Pollen Accelerate the Oviposition and Colony Foundation of Bombus breviceps (Hymenoptera: Apidae). Insects. 2023; 14(2):203. https://doi.org/10.3390/insects14020203
Chicago/Turabian StyleRen, Chang-Shi, Zhi-Min Chang, Lei Han, Xiang-Sheng Chen, and Jian-Kun Long. 2023. "Higher Essential Amino Acid and Crude Protein Contents in Pollen Accelerate the Oviposition and Colony Foundation of Bombus breviceps (Hymenoptera: Apidae)" Insects 14, no. 2: 203. https://doi.org/10.3390/insects14020203
APA StyleRen, C. -S., Chang, Z. -M., Han, L., Chen, X. -S., & Long, J. -K. (2023). Higher Essential Amino Acid and Crude Protein Contents in Pollen Accelerate the Oviposition and Colony Foundation of Bombus breviceps (Hymenoptera: Apidae). Insects, 14(2), 203. https://doi.org/10.3390/insects14020203