
Phylogenetic Structure Revealed through Combining DNA Barcodes with Multi-Gene Data for Agrodiaetus Blue Butterflies (Lepidoptera, Lycaenidae)

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
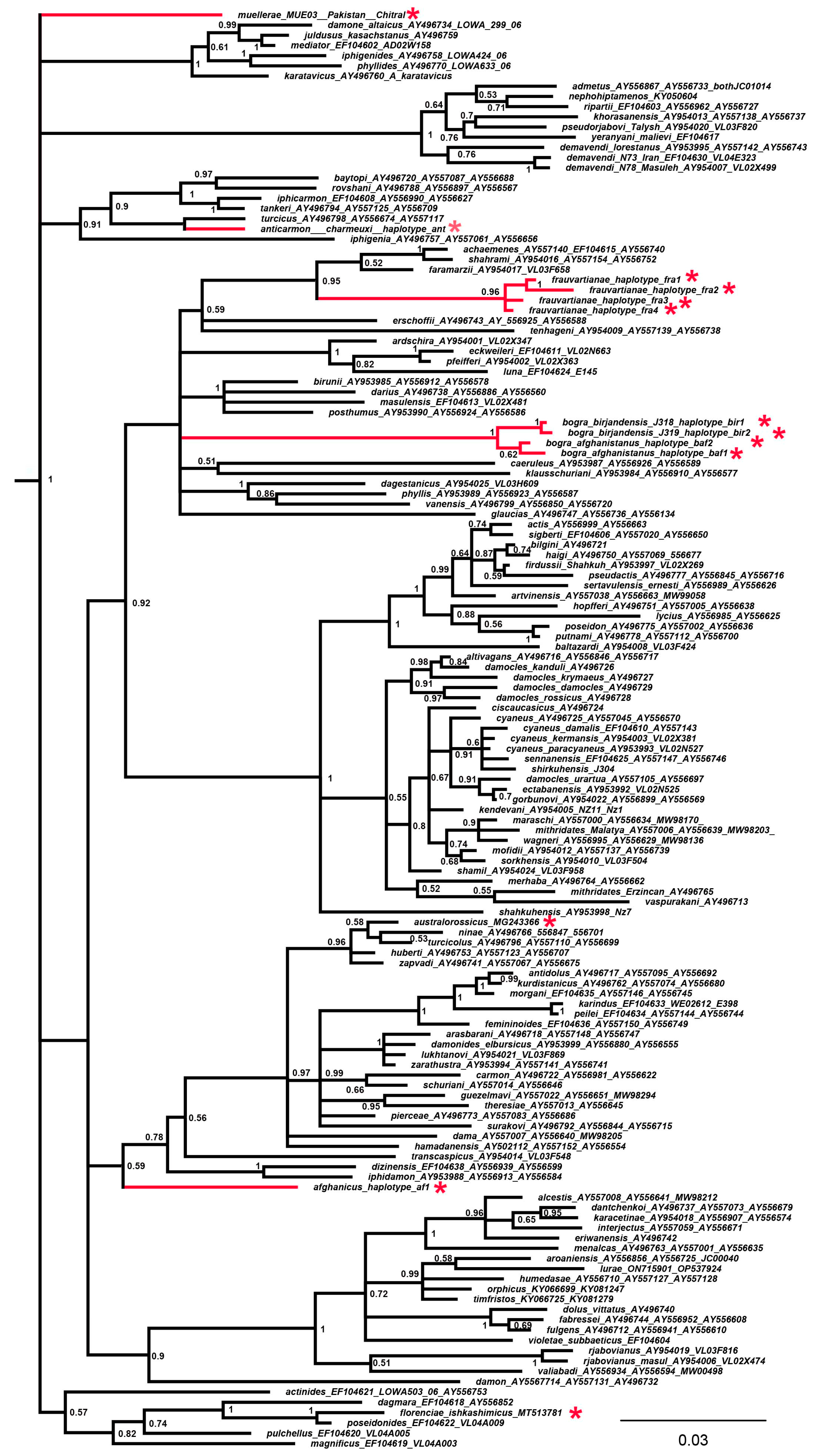
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- Checklist of the species groups and species of Agrodiaetus
References
- Rubinoff, D.; Holland, B.S. Between two extremes: Mitochondrial DNA is neither the panacea nor the nemesis of phylogenetic and taxonomic inference. Syst. Biol. 2005, 54, 952–961. [Google Scholar] [CrossRef] [PubMed]
- Will, K.W.; Mishler, B.D.; Wheeler, Q.D. The perils of DNA barcoding and the need for integrative taxonomy. Syst. Biol. 2005, 54, 844–851. [Google Scholar] [CrossRef]
- Knowles, L.L.; Carstens, B.C. Delimiting species without monophyletic gene trees. Syst. Biol. 2007, 56, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Talavera, G.; Lukhtanov, V.A.; Pierce, N.E.; Vila, R. Establishing criteria for higher-level classification using molecular data: The systematics of Polyommatus blue butterflies (Lepidoptera, Lycaenidae). Cladistics 2013, 29, 166–192. [Google Scholar] [CrossRef] [PubMed]
- Gapon, D.A.; Kuznetsova, V.G.; Maryańska-Nadachowska, A. A new species of the genus Rhaphidosoma Amyot et Serville, 1843 (Heteroptera, Reduviidae), with data on its chromosome complement. Comp. Cytogenet. 2021, 15, 467–505. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Nat. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef]
- Talavera, G.; Lukhtanov, V.A.; Pierce, N.; Vila, R. DNA barcodes combined with multilocus data of representative taxa can generate reliable higher-level phylogenies. Syst. Biol. 2022, 71, 382–395. [Google Scholar] [CrossRef]
- Hu, S.J.; Condamine, F.L.; Monastyrskii, A.L.; Cotton, A.M. A new species of the Graphium (Pazala) mandarinus group from Central Vietnam (Lepidoptera: Papilionidae). Zootaxa 2019, 4554, 286–300. [Google Scholar] [CrossRef]
- Xu, Z.-B.; Wang, Y.-Y.; Condamine, F.L.; Cotton, A.M.; Hu, S.-J. Are the yellow and red marked club-tail Losaria coon the same species? Insects 2020, 11, 392. [Google Scholar] [CrossRef]
- Hu, P.; Lu, L.; Hu, S.; Da, W.; Huang, C.-L.; Zhang, H.; Wang, D.; Zhang, Y.; Xu, Y.; Wang, R. Differentiation of the chestnut tiger butterfly Parantica sita (Lepidoptera: Nymphalidae: Danainae) in China. Front. Ecol. Evol. 2022, 10, 846499. [Google Scholar] [CrossRef]
- Cotton, A.M.; Doleck, T.; Zhang, X.; Inayoshi, Y.; Lohman, D.J.; Hu, S.J. Graphium septentrionicolus Page & Treadaway, 2013 (Lepidoptera: Papilionidae) is a distinct species. Zootaxa 2022, 5154, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.X.; Jiang, Z.H.; Wang, J.Q.; Song, K.; Zhang, C.; Hu, S.J. A revision of the Pieris napi-complex (Lepidoptera: Pieridae) and similar species with distribution in China. Arthropod Syst. Phylogeny 2023, 81, 257–287. [Google Scholar] [CrossRef]
- Hou, Y.; Cao, C.; Chiba, H.; Chang, Z.; Huang, S.; Zhu, L.; Kunte, K.; Huang, Z.; Wang, M.; Fan, X. Molecular phylogeny, historical biogeography, and classification of Pseudocoladenia butterflies (Lepidoptera: Hesperiidae). Mol. Phylogenet. Evol. 2023, 186, 107865. [Google Scholar] [CrossRef] [PubMed]
- Cong, Q.; Shen, J.; Zhang, J.; Li, W.; Kinch, L.N.; Calhoun, J.V.; Warren, A.D.; Grishin, N.V. Genomics reveals the origins of historical specimens. Mol. Biol. Evol. 2021, 38, 2166–2176. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, A.R.; Brown, J.M.; Stanger-Hall, K.; Lemmon, E.M. The effect of ambiguous data on phylogenetic estimates obtained by maximum likelihood and Bayesian inference. Syst. Biol. 2009, 58, 130–145. [Google Scholar] [CrossRef]
- Kandul, N.P.; Lukhtanov, V.A.; Dantchenko, A.V.; Coleman, J.W.S.; Sekercioglu, C.H.; Haig, D.; Pierce, N.E. Phylogeny of Agrodiaetus Hübner 1822 (Lepidoptera: Lycaenidae) inferred from mtDNA sequences of COI and COII and nuclear sequences of EF1-α: Karyotype diversification and species radiation. Syst. Biol. 2004, 53, 278–298. [Google Scholar] [CrossRef]
- Kandul, N.P.; Lukhtanov, V.A.; Pierce, N.E. Karyotypic diversity and speciation in Agrodiaetus butterflies. Evolution 2007, 61, 546–559. [Google Scholar] [CrossRef]
- Eckweiler, W.; Bozano, G.C. Guide to the Butterflies of the Palearctic Region. Lycaenidae Part IV; Bozano, G.C., Ed.; Omnes Artes: Milano, Italy, 2016; 132p, ISBN 9788887989182. [Google Scholar]
- de Lesse, H. Les nombres de chromosomes dans la classification du groupe d’Agrodiaetus ripartii Freyer (Lepidoptera, Lycaenidae). Rev. Fran. Entomol. 1960, 27, 240–264. [Google Scholar]
- de Lesse, H. Spéciation et variation chromosomique chez les Lépidoptères Rhopaloceres. Ann. Sci. Nat. Zool. Biol. Anim. 1960, 2, 1–223. [Google Scholar]
- Eckweiler, W.; Häuser, C.L. An illustrated checklist of Agrodiaetus Hübner, 1822, a subgenus of Polyommatus Latreille, 1804 (Lepidoptera: Lycaenidae). Nachr. Entomol. Ver. Apollo 1997, 16, 113–168. [Google Scholar]
- Olivier, A.; Puplesiene, J.; van der Pooten, D.; De Prins, W.; Wiemers, M. Revision of some taxa of Polyommatus (Agrodiaetus) transcaspicus group with description of a new species from Central Anatolia (Lepidoptera, Lycaenidae). Phegea 1999, 27, 281–296. [Google Scholar]
- Carbonell, F. Contribution a la conaissance du genre Agrodiaetus Hübner (1822), A. barmifiruze n. sp. et A. musa esfahensis n. ssp., en Iran méridional. Linneana Belg. 2000, 17, 211–217. [Google Scholar]
- Carbonell, F. Contribution a la connaissance du genre Agrodiaetus Hübner (1822), A. ahmadi et A. khorasanensis nouvelles espèces dans le Nord de l’Iran (Lepidoptera, Lycaenidae). Linneana Belg. 2001, 18, 105–110. [Google Scholar]
- Skala, P. New taxa of the subgenus Agrodiaetus Hübner, 1822 from Iran: Polyommatus (Agrodiaetus) faramarzii sp. n., P. (A.) shahrami sp. n. and P. (A.) pfeifferi astyages ssp. n. (Lepidoptera, Lycaenidae). Nachr. Entomol. Ver. Apollo 2001, 22, 101–108. [Google Scholar]
- Wiemers, M. Chromosome Differentiation and the Radiation of the Butterfly Subgenus Agrodiaetus (Lepidoptera: Lycaenidae: Polyommatus)—A Molecular Phylogenetic Approach. Ph.D. Dissertation, University of Bonn, Bonn, Germany, 2003; 203p. Available online: http://nbn-resolving.de/urn:nbn:de:hbz:5n-02787 (accessed on 15 August 2023).
- Schurian, K.G.; ten Hagen, W. Polyommatus (Agrodiaetus) urmiaensis sp. n. aus Nordwestiran (Lepidoptera: Lycaenidae). Nachr. Entomol. Ver. Apollo 2003, 24, 1–5. [Google Scholar]
- Lukhtanov, V.A.; Kandul, N.P.; Plotkin, J.B.; Dantchenko, A.V.; Haig, D.; Pierce, N.E. Reinforcement of pre-zygotic isolation and karyotype evolution in Agrodiaetus butterflies. Nature 2005, 436, 385–389. [Google Scholar] [CrossRef]
- Wiemers, M.; Keller, A.; Wolf, M. ITS2 secondary structure improves phylogeny estimation in a radiation of blue butterflies of the subgenus Agrodiaetus (Lepidoptera: Lycaenidae: Polyommatus). BMC Evol. Biol. 2009, 9, 300. [Google Scholar] [CrossRef]
- Vodolazhsky, D.I.; Yakovlev, R.; Stradomsky, B. Study of taxonomic status of some specimens of subgenus Agrodiaetus (Lepidoptera: Lycaenidae: Polyommatus) from Western Mongolia based on mtDNA markers. Caucas. Entomol. Bull. 2011, 7, 81–82. [Google Scholar] [CrossRef]
- Lukhtanov, V.A.; Shapoval, N.A.; Dantchenko, A.V. Taxonomic position of several enigmatic Polyommatus (Agrodiaetus) species (Lepidoptera, Lycaenidae) from Central and Eastern Iran: Insights from molecular and chromosomal data. Comp. Cytogenet. 2014, 8, 313–322. [Google Scholar] [CrossRef]
- Rougerie, R.; Smith, M.A.; Fernandez-Triana, J.; Lopez-Vaamonde, C.; Ratnasingham, S.; Hebert, P.D. Molecular analysis of parasitoid linkages (MAPL): Gut contents of adult parasitoid wasps reveal larval host. Mol. Ecol. 2011, 20, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial Cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotech. 1994, 3, 294–299. [Google Scholar]
- deWaard, J.R.; Ivanova, N.V.; Hajibabaei, M.; Hebert, P.D.N. Assembling DNA barcodes: Analytical protocols. In Environmental Genomics, Methods in Molecular Biology; Martin, C.C., Ed.; Humana Press: Totowa, NJ, USA, 2008; Volume 410, pp. 275–283. ISBN 13: 978-1588297778. [Google Scholar]
- Hall, T. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Balint, Z. Reformation of the Polyommatus section with a taxonomic and biogeographic overview (Lepidoptera, Lycaenidae, Polyommatini). Neue Entomol. Nachricht. 1997, 40, 1–67. [Google Scholar]
- Coutsis, J.G. Notes concerning the taxonomic status of Agrodiaetus tankeri de Lesse (Lepidoptera: Lycaenidae). Nota Lepidopterol. 1985, 8, 8–14. [Google Scholar]
- Coutsis, J.G. The blue butterflies of the genus Agrodiaetus Hübner (Lep., Lycaenidae): Symptoms of taxonomic confusion. Nota Lepidopterol. 1986, 9, 159–169. [Google Scholar]
- Munguira, M.L.; Martin, L.; Perez-Valiente, M. Karyology and distribution as tools in the taxonomy of Iberian Agrodiaetus butterflies. Nota Lepidopterol. 1995, 17, 125–140. [Google Scholar]
- Kandul, N.P. The karyology and the taxonomy of the blue butterflies of the genus Agrodiaetus Hubner, (1822) from the Crimea. Atalanta 1997, 28, 111–119. [Google Scholar]
- Wiemers, M.; De Prins, J. Polyommatus (Agrodiaetus) paulae sp. nov. from Northwest Iran, discovered by means of molecular, karyological and morphological methods. Entomol. Z. 2004, 114, 155–162. [Google Scholar]
- Kolev, Z. Polyommatus dantchenkoi (Lukhtanov & Wiemers, 2003) tentatively identified as new to Europe, with a description of a new taxon from the Balkan Peninsula. Nota Lepidopterol. 2005, 28, 25–34. [Google Scholar]
- Hesselbarth, G.; Van Oorschot, H.; Wagener, S. Die Tagfalter der Türkei; Selbstverlag Sigbert Wagener: Bocholt, Germany, 1995; Volumes 1–3, 754p. [Google Scholar]
- Koçak, A.O.; Kemal, M. A study on the biodiversity, zoogeography and taxonomy of the section Agrodiaetus Hbn. in the genus Polyommatus Latr. (Lycaenidae, Lepidoptera). Centre Entomol. Stud. Misc. Pap. 2001, 78/79, 1–11. [Google Scholar]
- International Commission on Zoological Nomenclature. International Code of Zoological Nomenclature; The International Trust for Zoological Nomenclature: London, UK, 1999; Available online: https://www.iczn.org/the-code/the-code-online/ (accessed on 15 August 2023).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Laboratory ID | GeneBank | Haplotype | Country | Locality |
|---|---|---|---|---|---|
| P. (A.) afghanicus | AF05 | OR413713 | af1 | Afghanistan | Koh-i-Baba, Band-i-Amir, 34.8294° N, 67.1805° E, 2900–3000 m, 2 July 2009, Yu. Skrylnik leg. |
| BPAL2125 | OR413714 | af1 | Afghanistan | near Kabul, July 2010, I. Pljushch leg. | |
| BPAL2126 | OR413715 | af1 | Afghanistan | the same locality | |
| BPAL2127 | OR413716 | af1 | Afghanistan | the same locality | |
| BPAL2128 | OR413717 | af1 | Afghanistan | the same locality | |
| P. (A.) anticarmon (= charmeuxi) | CHAR01 | OR424389 | ant | Turkey | Hakkari Prov., vic. Üzümcū 1300 m, 28 June – 4 July 1976 |
| P. (A.) bogra afghanistanus | XX21 | OR424390 | baf1 | Afghanistan | Bamyan prov., 8 km S Bamyan, 2700 m, 31 May 2010, O. Pak leg. |
| AAF02 | OR424391 | baf2 | Afghanistan | the same locality | |
| AAF10 | OR424392 | baf1 | Afghanistan | Bamiyan prov., 34.2155° N, 66.8994° E, 2545 m, 23 June 2016, O. Pak leg. | |
| AAF11 | OR424393 | baf1 | Afghanistan | the same locality | |
| P. (A.) bogra birjandensis | J318 | OR413718 | bir1 | Iran | South Khorasan Prov., 26 km N of Birjand, 1900–2000 m, 14 July 2007, V. Lukhtanov leg. |
| J319 | OR413719 | bir2 | Iran | the same locality | |
| P. (A.) frauvartianae | AAF01 | OR424394 | fra1 | Afghanistan | Bamiyan prov., Yakawlang District, Bandi-Amir env., 3300 m, 3 August 2011, O. Pak leg. |
| AAF03 | OR424395 | fra2 | Afghanistan | the same locality | |
| AAF04 | OR424396 | fra1 | Afghanistan | the same locality | |
| AAF05 | OR424397 | fra1 | Afghanistan | the same locality | |
| AAF07 | OR424398 | fra3 | Afghanistan | the same locality | |
| AAF08 | OR424399 | fra4 | Afghanistan | Bamiyan prov., Panjab District, Kohi-Baba Mts., Rashak Mts., Shatu Pass, 3490 m, 6 August 2011, O. Pak leg. | |
| AAF09 | OR424400 | fra1 | Afghanistan | the same locality | |
| P. (A.) muellerae | MUE03 | OR413720 | mu1 | Pakistan | Chitral, Birmogh Lasht, 35.8981° N, 71.7712° E, 2600–3000 m, 1 July 2001, leg. V. Tuzov |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lukhtanov, V.A.; Shapoval, N.A.; Dantchenko, A.V.; Eckweiler, W. Phylogenetic Structure Revealed through Combining DNA Barcodes with Multi-Gene Data for Agrodiaetus Blue Butterflies (Lepidoptera, Lycaenidae). Insects 2023, 14, 769. https://doi.org/10.3390/insects14090769
Lukhtanov VA, Shapoval NA, Dantchenko AV, Eckweiler W. Phylogenetic Structure Revealed through Combining DNA Barcodes with Multi-Gene Data for Agrodiaetus Blue Butterflies (Lepidoptera, Lycaenidae). Insects. 2023; 14(9):769. https://doi.org/10.3390/insects14090769
Chicago/Turabian StyleLukhtanov, Vladimir A., Nazar A. Shapoval, Alexander V. Dantchenko, and Wolfgang Eckweiler. 2023. "Phylogenetic Structure Revealed through Combining DNA Barcodes with Multi-Gene Data for Agrodiaetus Blue Butterflies (Lepidoptera, Lycaenidae)" Insects 14, no. 9: 769. https://doi.org/10.3390/insects14090769
APA StyleLukhtanov, V. A., Shapoval, N. A., Dantchenko, A. V., & Eckweiler, W. (2023). Phylogenetic Structure Revealed through Combining DNA Barcodes with Multi-Gene Data for Agrodiaetus Blue Butterflies (Lepidoptera, Lycaenidae). Insects, 14(9), 769. https://doi.org/10.3390/insects14090769