1. Introduction
Recent studies place the origin of Achilidae Stål, 1866 in the Jurassic period [
1], and this may even extend to the end of the Triassic [
2,
3]. By the mid-Cretaceous period, approximately 100 million years ago, all major lineages of Achilidae had emerged and began diversifying [
2,
3]. It is therefore not surprising to find a significant presence of Achilidae in Burmese amber inclusions, where they are the second most diverse of extant planthopper families in the Cretaceous fossil record, following Cixiidae Spinola, 1839, and surpassing Derbidae Spinola, 1839 and Nogodinidae Melichar, 1898 [
4,
5,
6,
7,
8,
9].
The taxon Achilidae was initially separated by Stål as the subfamily ‘Achilida’ within the family ‘Fulgorida‘ in his work “Hemiptera Africana” in 1866 [
10]. The taxon was later formally recognized as a distinct family by Muir in 1923 [
11]. The evolutionary history and taxonomic divisions within Achilidae have been reviewed by Brysz and Szwedo [
12].
The Achilidae currently group 162 genera and 521 species, respectively only accounting for 6.4% and 3.7% of Fulgoromorpha diversity [
13]. Following Emeljanov’s classification system [
14,
15], they were initially subdivided into three supertribes, currently upgraded to subfamilies by Bartlett et al. [
16], and 12 tribes: Achilinae Stål, 1866 (Achilini Stål, 1866, Achillini Emeljanov, 1991), Apatesoninae Metcalf, 1938 (Apatesonini Metcalf, 1938, Ilvini Emeljanov, 1991, Seviini Emeljanov, 1991, Tropiphlepsiini Emeljanov, 1991), and Myconinae Fennah, 1950 (Amphignomini Emeljanov, 1991, Mycarini Emeljanov, 1991, Myconini Fennah, 1950, Plectoderini Fennah, 1950, Rhotalini Fennah, 1950, and the fossil tribe Waghildini Szwedo, 2006). The placement of another fossil tribe, Ptychoptilini Emeljanov, 1990, within the Achilidae or Derbidae, remains uncertain [
5,
17].
Achilidae are distributed worldwide, with a latitudinal profile particularly well represented in the northern hemisphere between 5° and 55° [
13]. As obligate phytophagous species, they are predominantly associated with Fagales, Pinales, Ericales, and Rosales, accounting for 23.5%, 14.1%, 11.8%, and 8.2% of the records, respectively [
13]. They are generally recognized as one of the planthopper groups best adapted to temperate climates [
18].
The documented fossil taxa of Achilidae in the Cretaceous period are rare, with only three genera and four species recognized. Hamilton [
19] described the first fossil genus
Acixiites Hamilton, 1990, with
A. immodesta Hamilton, 1990 (the type species), and
A. costalis Hamilton, 1990, from the Crato Formation in Brazil representing the oldest fossil records of Achilidae from the Cretaceous period. Cockerell [
20] initially described the first Myanmar amber planthopper taxon in the Delphacid genus
Liburnia Stål, 1866, transferred to Achilidae by Shcherbakov [
21], and redescribed by Szwedo [
4] in the new genus
Niryasaburnia Szwedo, 2004, with the type species
N. burmitina (Cockerell, 1917). According to Cruickshank and Ko [
22], the specimen belongs to the ‘old mines’ location in Hukawng Valley near Tanai. Brysz et al. [
8] described the second genus from the Hukawng Valley also,
Amphignokachinia Brysz & Szwedo, 2023, with the type species
A. subversa Brysz & Szwedo, 2023, as the first mid-Cretaceous representative of the tribe Amphignomini in the subfamily Myconinae.
In this paper, a new Burmese amber species Niryasaburnia nigrutomia sp. nov. is described of Achilidae from the Kachin state (Tanai) of northern Myanmar, easily distinguished by its forewing pattern coloration and metatibiotarsal formula from N. burmitina (Cockerell, 1917). Additionally, a new Burmese amber genus Sinuovenaxius gen. nov., along with S. kachinensis sp. nov., is also described from this location. This discovery marks the third amber genus and the sixth fossil species of the family from the Cretaceous period. Furthermore, a new tribe Niryasaburniini trib. nov. is established within Myconinae to include the genera Niryasaburnia and Sinuovenaxius gen. nov.
3. Systematic Paleontology
Order: Hemiptera Linnaeus, 1758
Suborder: Fulgoromorpha Evans, 1946
Superfamily: Fulgoroidea Latreille, 1807
Family: Achilidae Stål, 1866
Subfamily: Myconinae Fennah, 1950
Key to tribes of Myconinae
1. Metatibia with more than 4 lateral spines..................................................................................2
-. Metatibia with no more than 3 lateral spines.............................................................................3
2. CuA 2-branched in prenodal region on forewing; apical teeth of metatarsomeres I and II both with subapical platellar sensilla....................................................Waghildini Szwedo, 2006
-. CuA 3-branched in prenodal region on forewing; apical teeth of metatarsomere I without subapical platellar sensilla, but metatarsomere II with this sensilla..........................................................................................................Rhotalini Fennah, 1950
3. Genae with subantennal carinae; mesonotum without lateral carinae...........................................................................................Amphignomini Emeljanov, 1991
-. Genae without subantennal carinae; mesonotum with lateral carinae....................................4
4. MP with at least 6 terminals, CuA with at least 4 terminals on forewing......................................................................................................Myconini Fennah, 1950
-. MP with no more than 5 terminals, CuA with no more than 3 terminals on forewing.............................................................................................................................................5
5. Head with compound eyes around 2/3 length of pronotum.................................................................................................Plectoderini Fennah, 1950
-. Head with compound eyes around half length of pronotum..................................................6
6. Forewing with ScP+R and CuA forking before level of fusion of Pcu and A1; hindwing with CuA 3-branched............................................................................Mycarini Emeljanov, 1991
-. Forewing with ScP+R and CuA forking after level of fusion of Pcu and A1; hindwing with CuA 2-branched......................................................................................Niryasaburniini trib. nov.
Niryasaburniini Wang & Bourgoin trib. nov.
LSIDurn:lsid:zoobank.org:act:28F1995F-0AF4-49B3-BBAF-238A49F7533C
Type genus. Niryasaburnia Szwedo, 2004.
Composition. Niryasaburnia Szwedo, 2004, Sinuovenaxius gen. nov.
Diagnosis: Head with compound eyes around half length of pronotum. Vertex with anterior margin almost straight. Frons with median carina elevated; lateral margins subparallel above level of compound eyes, then strongly diverging below lower margin of compound eyes and curved. On forewings, basal cell narrow, elongated; precostal and stigmal areas without veinlets connecting anterior margin, no pterostigma area individualized; rather simple venation, MP with two or three terminals; CuA with two terminals; ScP+R and CuA forking late, well after the level of fusion of Pcu and A1; MP forking late well after nodal line level. On hindwings, RP simple with one terminal; both MP and CuA with two terminals; A2 simple, reaching posterior margin. Hind tibia with two or three small lateral spines including subgenual one; metatibial apical teeth strong, long, in row widely diverging apically; apical teeth of metatarsomeres I and II both with subapical platellar sensilla.
Niryasaburnia Szwedo, 2004
Type species:
Niryasaburnia burmitina (Cockerell, 1917), by monotypy [
4].
Modified Diagnosis: Niryasaburnia can be distinguished by the late forking of MP well after nodal line level, ScP+R and CuA forking after level of the fusion of Pcu+A1, ScP+R forking before end of clavus and before forking of CuA, C4 shorter than C3, RP 2-branched, MP forked twice with three terminals on forewing; the ventral margin of frons strongly diverging ventrally (twice as wide than between compound eyes) well below the lower margin of the compound eyes, frons with distinct median carina from anterior margin to posterior margin; long anteclypeus reaching the base of prolegs; rostrum exceeding metatrochanter; hind tibia with two lateral spines including subgenual one; metatibial apical teeth strong, long in row widely diverging apically, the apical teeth of metatarsomeres I and II both with subapical platellar sensilla.
Niryasaburnia nigrutomia Deng & Bourgoin sp. nov.
LSIDurn:lsid:zoobank.org:act:F17983FC-AEBF-4821-AC96-147897DCCE82
Diagnosis: This new species differs from
Niryasaburnia burmitina (Cockerell, 1917) by the following characteristics: (1) tegmen with irregular brown patches as in
Figure 1A (without patches in
N. burmitina); (2) forewing, RA
2 with only one terminal (two in
N. burmitina); (3) transverse veins
r-m1 and
m-cu1 placed at same level of the forking of MP (distad in
N. burmitina); (4) lateral carinae of the frontoclypeus without distinct granulation (present in
N. burmitina); (5) metatibiotarsal formula: 2-8/8/7, versus 2-9/10/8 in
N. burmitina.
Etymology: The name refers to the black thorax, nigrum pronotum in Latin, arbitrarily concatenated into nigrutomia.
Type material: Holotype, MDHP130. Female adult, in Burmese amber, from Hukawng Valley (Tanai location), Kachin State, Northern Myanmar.
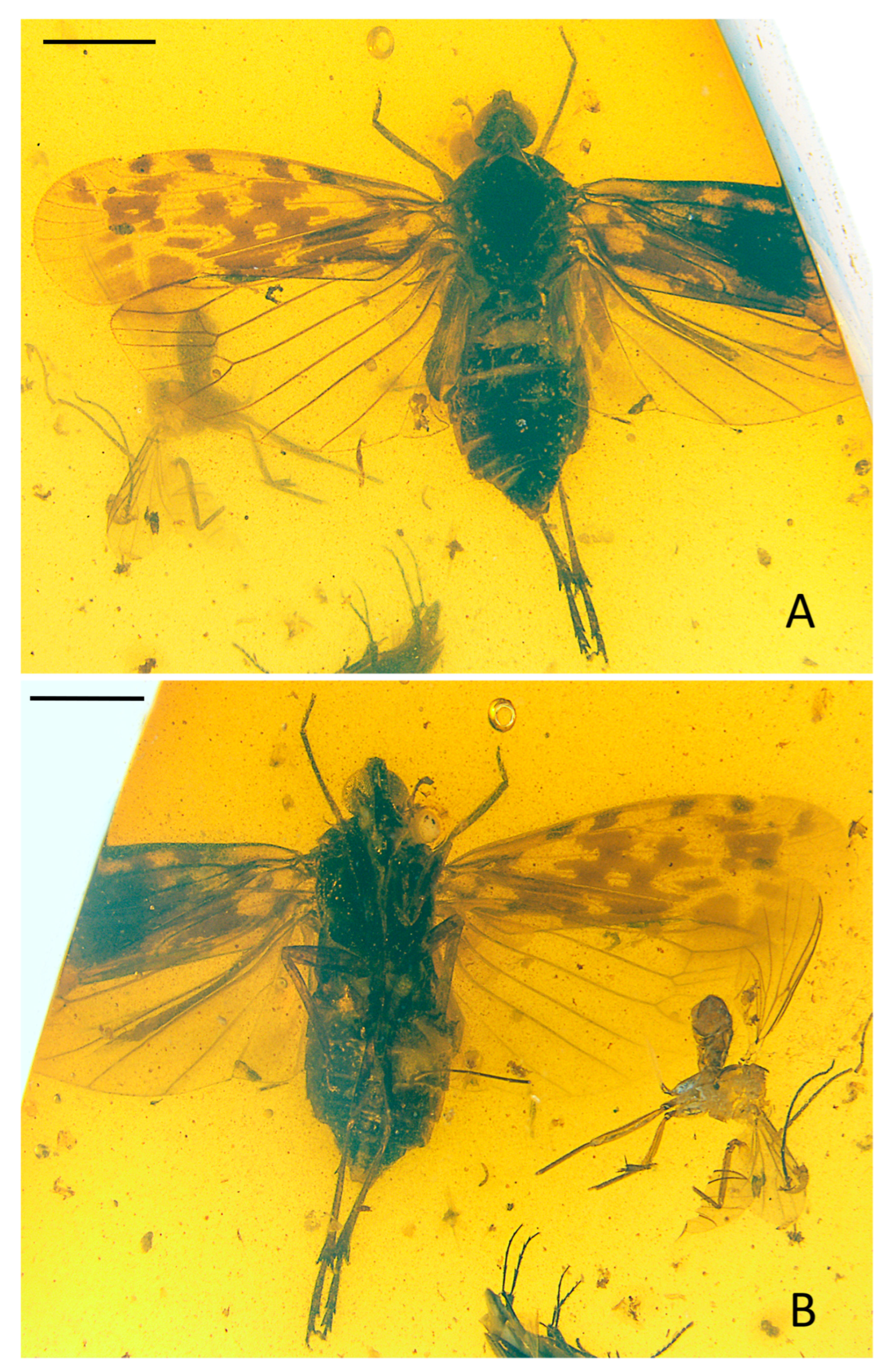
Description:
Small-size insect (
Figure 1,
Figure 2 and
Figure 3). Total length including tegmina 4.99 mm. The forewing and hindwing in right side not completed due to the fossil conditions. Anal lobe of forewing in left side folded but well visible in right side. Head slightly split from thorax in the specimen. Frons and clypeus slightly slanted in ventral view in the specimen. The apex of left middle leg missing.
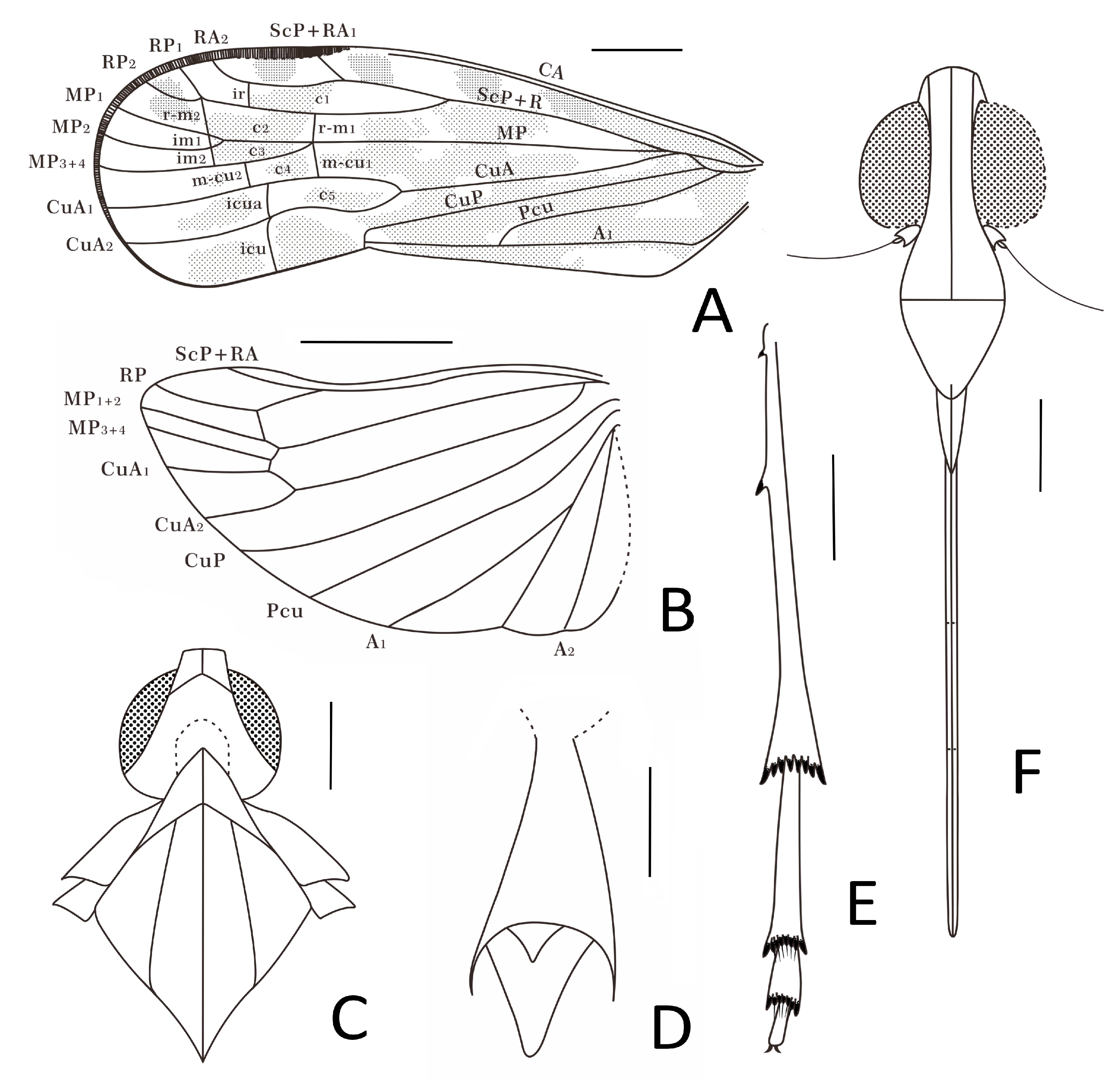
Head. Head width with compound eyes (
Figure 2A) 0.61 mm. Vertex (
Figure 2A) length 0.16 mm in midline, width in anterior margin 0.16 mm, width at middle 0.19 mm, width at base 0.26 mm. Compound eyes (
Figure 2A) length 0.35 mm, width 0.12 mm. Frons (
Figure 2B) length 0.65 mm, width in anterior margin 0.15 mm, width at middle 0.13 mm, width at base 0.31 mm. Clypeus (
Figure 2B) length 0.37 mm. Rostrum (
Figure 1B) 1.34 mm.
Thorax. Pronotum (
Figure 2A) length 0.22 mm, width 1.04 mm. Mesonotum (
Figure 2A) length 0.98 mm, widest width 0.90 mm.
Forewings. Tegmen (
Figure 1A,B,
Figure 2C and
Figure 3A) length 3.96 mm in longest part, width 1.33 mm in widest part; RA
2 with one terminal; transverse veins
r-m1 and
m-cu1 placed at same level of the forking of MP; tegmen hyaline, large area of irregular brown patches on tegmen, five of them almost equidistant located anteriorly on costal and radial area, wider markings posteriorly more or less confluent and irregular, as figured; clavus with darker brown markings; brown patches distally lighter or absent; costal membrane (
Figure 2D) clearly visible.
Hindwings. Hindwing (
Figure 1A and
Figure 2C) 3.30 mm long in longest part, 1.78 mm in widest part.
Legs. Fore femur, tibia, and tarsus (
Figure 1B and
Figure 2F) length 0.79 mm, 0.84 mm, 0.36 mm; middle femur, tibia and tarsus (
Figure 1B and
Figure 2G) length 0.88 mm, 1.18 mm, 0.22 mm; hind femur, tibia and tarsus (
Figure 1B and
Figure 2H) length 0.82 mm, 1.34 mm, 0.75 mm; basitarsomere (
Figure 2H) length 0.49 mm, metatarsomere II (
Figure 2H) length 0.16 mm, metatarsomere III (
Figure 2H) length 0.10 mm; metatibiotarsal formula: 2-8/8/7 (
Figure 2H and
Figure 3E).
Female terminalia. A female specimen but not clear enough and complete for description. In dorsal view, anal tube (
Figure 2E and
Figure 3D) sub-quadrangular with apical margin concave at middle, anal styles (
Figure 2E and
Figure 3D) developed.
Sinuovenaxius Wang & Bourgoin gen. nov.
LSIDurn:lsid:zoobank.org:act:7CAD226A-27DA-4C78-AFC8-CF6FEA73562C
Type species: Sinuovenaxius kachinensis sp. nov.
Diagnosis: From other Burmese amber Achilid fossils, the new genus approaches Niryasaburnia, but differs in the following characters on the forewing: (1) ScP+R forking late after end of clavus (before in Niryasaburnia), and after forking of CuA (before in Niryasaburnia); (2) all branches single after nodal line level (RP and MP1+2 forked once in Niryasaburnia); (3) CuA2 strongly sinuated at base (slightly sinuated in Niryasaburnia).
Sinuovenaxius gen. nov. differs from all currently known Achilids by its rather simple venation, by the combination of the following characters: (1) late forking of ScP+R after nodal level and after CuA forking; (2) as in several Plectoderini genera CuA2 on forewing distinctly sinuated but stronger in Sinuovenaxius with a late forking of CuA well after the level of fusion of Pcu +A1.
Etymology: The Latin name refers to the sinuate CuA2 vein on the forewing, sinuosus vena, arbitrarily concatenated with -xius, gender masculine.
Description:
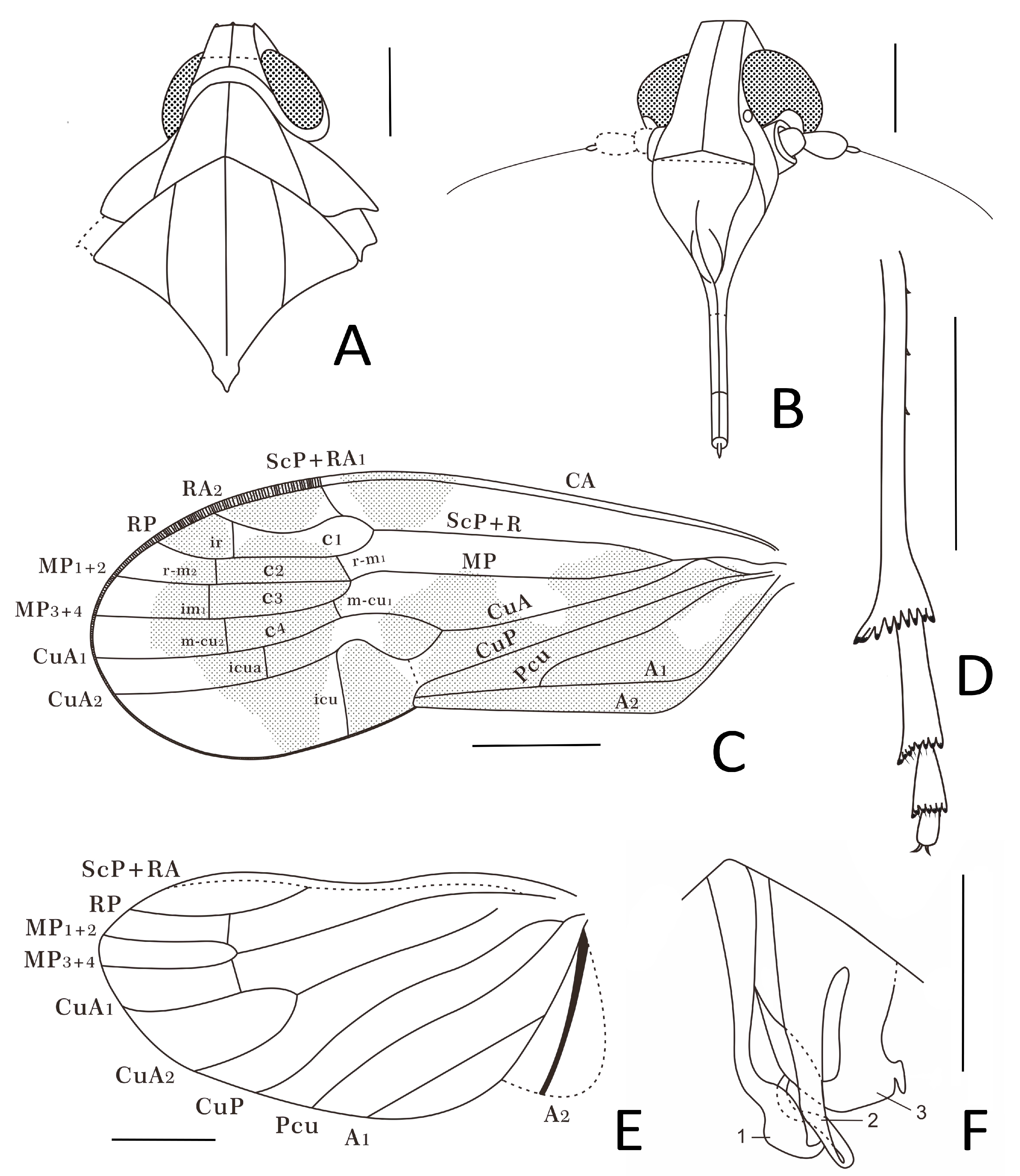
Head. Vertex (
Figure 4A,
Figure 5A and
Figure 6A) trapezoid, slightly wider than long, with a distinct median carina, reaching anterior and posterior margins of vertex; anterior margin almost straight, surpassing the upper margin of compound eyes; lateral margins inclined outwards to the base; posterior margin roundly concave. Frons (
Figure 4B,
Figure 5B and
Figure 6B) much longer than wide, the length in midline of frons about 2.3 times longer than wide, with distinct elevated median carina from the apical margin to the base of frons; apical margin straight; lateral margins subparallel above the lower margin of compound eyes, and then expanded outward. Median ocellus absent. Gena (
Figure 5B and
Figure 6B) with a pair of lateral ocelli touching compound eyes. Compound eyes (
Figure 5B and
Figure 6B) large, bulged. Antennae (
Figure 5B and
Figure 6B) with scape and elongated pedicel globulous; in frontal view, pedicel surpassing external margin of compound eyes; flagellum twice longer than pedicel, basal bulb of flagellum oval. Frontoclypeal suture (
Figure 5B and
Figure 6B) slightly angular. Clypeus (
Figure 5B and
Figure 6B) large, dorsally almost same as frons at length, triangular; with median carina well present in the basal 2/3; lateral margins converging to base. Rostrum (
Figure 4B,
Figure 5E and
Figure 6B) short, just reaching mesocoxae.
Thorax. Pronotum (
Figure 5A and
Figure 6A) strong, wide and large, length along midline longer than length of vertex, saddle-like; anterior margin roundly convex, reaching to the middle level of compound eyes; posterior margin angularly concave, forming an angle around 120° at middle; disc with three distinctly elevated carinae derived from anterior margin reaching to posterior margin. Mesonotum (
Figure 5A and
Figure 6A) diamond-shaped, slightly wider than length in midline, about 3.3 times longer in midline than the length of pronotum in midline; with three obvious longitudinal carinae from anterior margin to posterior margin, lateral carinae subparallel to median carina, the lateral areas slant ventrally. Mesoscutellum (
Figure 5A and
Figure 6A) with lateral margins subparallel, and then triangular.
Forewings. Tegmen (
Figure 4A,
Figure 4B,
Figure 5C and
Figure 6C) distinctly longer than wide, color markings interspersed on the surface, with a subapical line delimitating seven distal apical open cells. Costal margin slightly curved; apical forewing margin rounded. Clavus almost half of tegmen length, Pcu and A
1 fused slightly after middle of clavus, clavus closed, the stem of Pcu+A
1 reaching CuP at the apex of clavus. The common stalk ScP+R very long, subparallel to costal margin; ScP+R forking close to nodal line level after apex of clavus level, at about 2/3 of tegmen length; ScP+RA
1 single, reaching anterior margin; RA
2 and RP base curved just distal to fork, both with only one terminal. Common stem ScP+R+MP short; MP bifurcated in two branches MP
1+2 and MP
3+4 around apical 1/3 of tegmen, slightly after the forking of ScP+R, both MP
1+2 and MP
3+4 simple and nearly straight. Two
r-m, respectively, after the ScP+R fork and
ir;
im1 slightly after
r-m2; two
m-cu, respectively, after proximally stalked C3 and before
im1. C1 and C5 expanded at base; C2 and C3 subparallel; C4 quadrangular, shorter than C3. Stem CuA bifurcated in CuA
1 and CuA
2 slightly before apex of clavus; CuA
1 single, distinctly curved upward in the basal half; CuA
2 single, strongly S-shaped sinuated proximally;
icua before other subdistal veinlets, but C5 still the longest cell;
icu long, from the middle of C5 to forewing margin.
Hindwings. Hindwing (
Figure 4A,
Figure 4B,
Figure 5C and
Figure 6E) more acute at MP
1+2 level; venation pattern similar to
Niryasaburnia. ScP+RA and RP separating at 3/5 length of costal margin; RP simple, reaching apical margin; MP bifurcate to simple MP
1+2 and MP
3+4 distad to the forking of ScP+RA and RP; CuA with two terminals, fork of CuA nearly at the same level as the fork of ScP+RA and RP; veins CuP and Pcu simple, slightly sinuate; A
1 straight; A
2 thick, slightly arcuate anteriorly, reaching posterior margin; transverse veins
r-m and
m-cu slightly after the forking of MP.
Legs. Fore and middle legs (
Figure 4B and
Figure 5E) robust; hind femur (
Figure 4B) equal width, tibia (
Figure 5D and
Figure 6D) with three lateral spines in basal half including subgenual one, apical portion slightly wider with strong apical teeth placed in a row strongly widening; basitarsomere (
Figure 5D and
Figure 6D) longer than the combined length of metatarsomeres II and III (
Figure 5D and
Figure 6D), apical teeth on metatarsomeres I and II equal length, subapical platellar sensilla present.
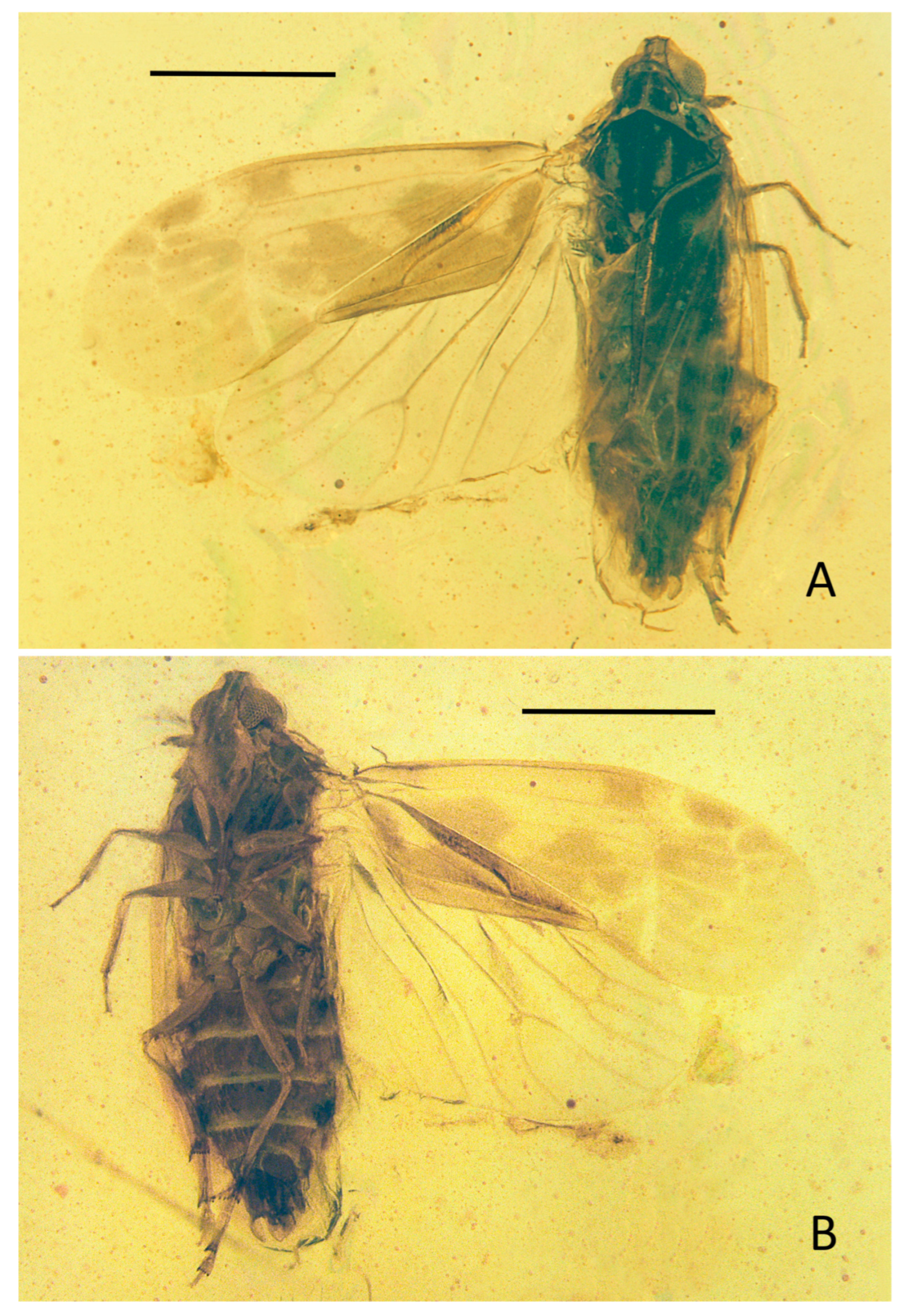
Sinuovenaxius kachinensis Wang & Bourgoin sp. nov.
LSIDurn:lsid:zoobank.org:act:1C4A45C9-8067-4463-A8CB-93EB726978C8
Diagnosis: This species could be quickly recognized by the processes of the phallic complex and the metatibiotarsal formula 3-9/8/7.
Etymology: This name refers to the location of this species in Kachin state from Myanmar.
Type material: Holotype, MDHP78. Male adult, in Burmese amber, from Hukawng Valley (Tanai location), Kachin State, Northern Myanmar.
Description:
Head. Head width with compound eyes (
Figure 5A) 0.49 mm. Vertex (
Figure 5A) length 0.12 mm in midline, width in anterior margin 0.13 mm, width at middle 0.16 mm, width at base 0.20 mm. Compound eyes (
Figure 5A) length 0.27 mm, width 0.16 mm. Antennae (
Figure 5B) scape length 0.07 mm, pedicel length 0.18 mm, flagellum length 0.43 mm. Frons (
Figure 5B) length 0.42 mm, width in anterior margin 0.13 mm, width at middle 0.18 mm, width at base 0.28 mm. Clypeus (
Figure 5B) length 0.40 mm. Rostrum (
Figure 4B) 0.39 mm.
Thorax. Pronotum (
Figure 5A) length 0.20 mm, width 0.84 mm. Mesonotum (
Figure 5A) length 0.66 mm, widest width 0.74 mm.
Forewings. Tegmen length 2.72 mm in longest part, width 1.06 mm in widest part; tegmen (
Figure 4A,B and
Figure 5C) hyaline, the surface of tegmina with irregular brown patches on apical half in the cell areas, beyond ScP, on basal 1/4 and the middle area from MP to clavus margin; the markings close to clavus darker; veinlets (
Figure 5C) in apical half and CuP yellow, others brown.
Hindwings. Hindwing (
Figure 4A,B and
Figure 5C) hyaline, 2.24 mm long in longest part, 1.22 mm in widest part.
Legs. Fore femur, tibia, and tarsus (
Figure 4B and
Figure 5E) length 0.56 mm, 0.37 mm, 0.22 mm; middle femur, tibia, and tarsus (
Figure 4B and
Figure 5E) length 0.55 mm, 0.46 mm, 0.23 mm; hind femur, tibia, and tarsus (
Figure 4B) length 0.55 mm, 0.68 mm, 0.51 mm; basitarsomere (
Figure 5D) length 0.27 mm, metatarsomere II (
Figure 5D) length 0.12 mm, metatarsomere III (
Figure 5D) length 0.12 mm; metatibiotarsal formula: 3-9/8/7 (
Figure 5D and
Figure 6D).
Male terminalia. In ventral view, phallic complex (
Figure 5F and
Figure 6F) asymmetric; the left process of periandrium (1) long, with apex broader and sinuated, curved to middle; the middle process, aedeagus
s.s. (2), slender and twisty to left side in the apex; the right process of periandrium (3) shorter and broad, with a large sharp triangular process in the left side directed to basal and a small short spine-like process in the right side directed to caudal.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





