
Revealing Changes in Ovarian and Hemolymphatic Metabolites Using Widely Targeted Metabolomics between Newly Emerged and Laying Queens of Honeybee (Apis mellifera)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Acquisition of Experimental Samples
2.2. Gathering the Ovary and Hemolymph of the Queen Bee
2.3. Widely Targeted Metabolomics Profiling Methods and Conditions
2.4. Data Analysis and Statistics
3. Results
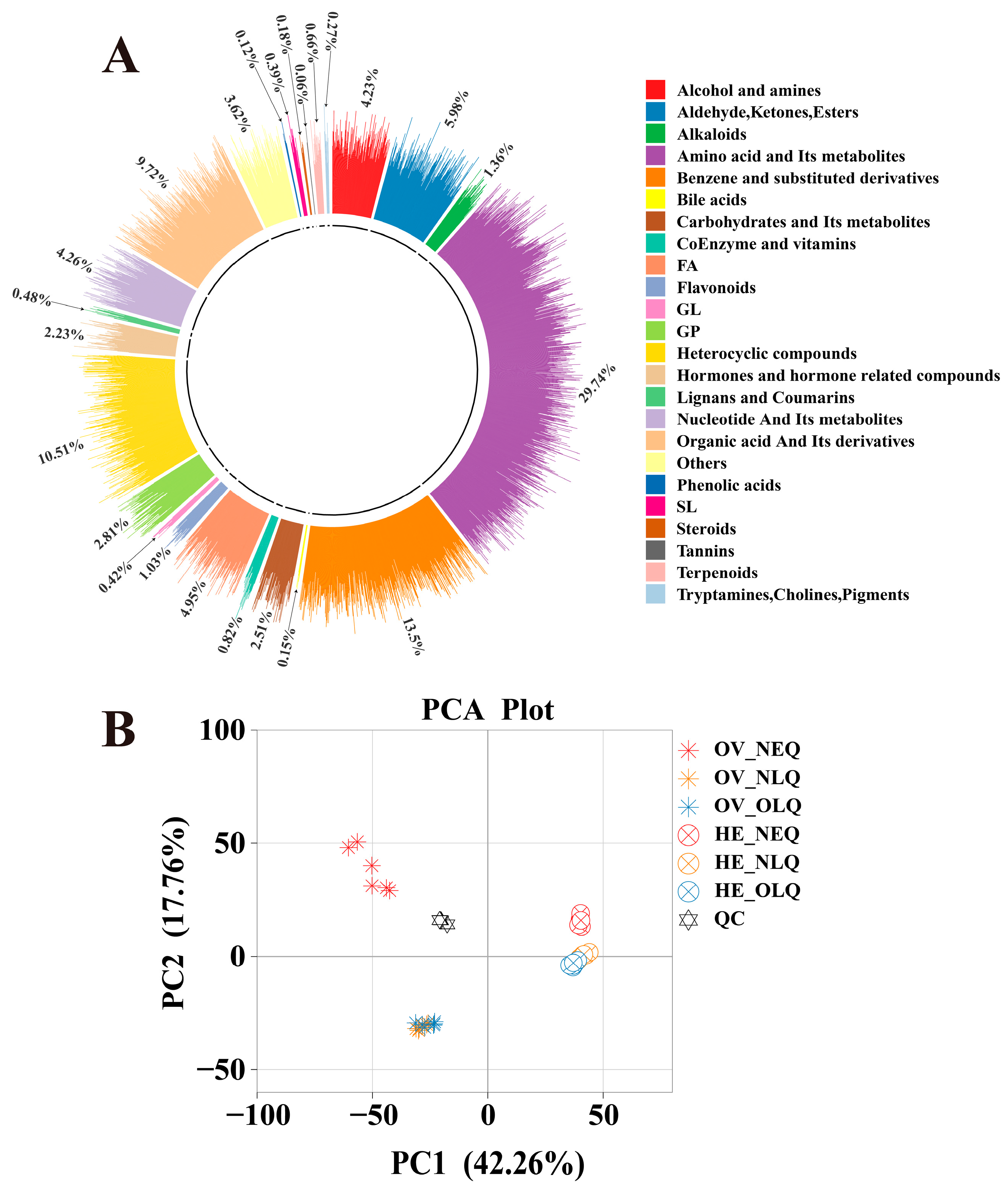
3.1. Identification of Metabolites and Widely Targeted Metabolomic Analysis
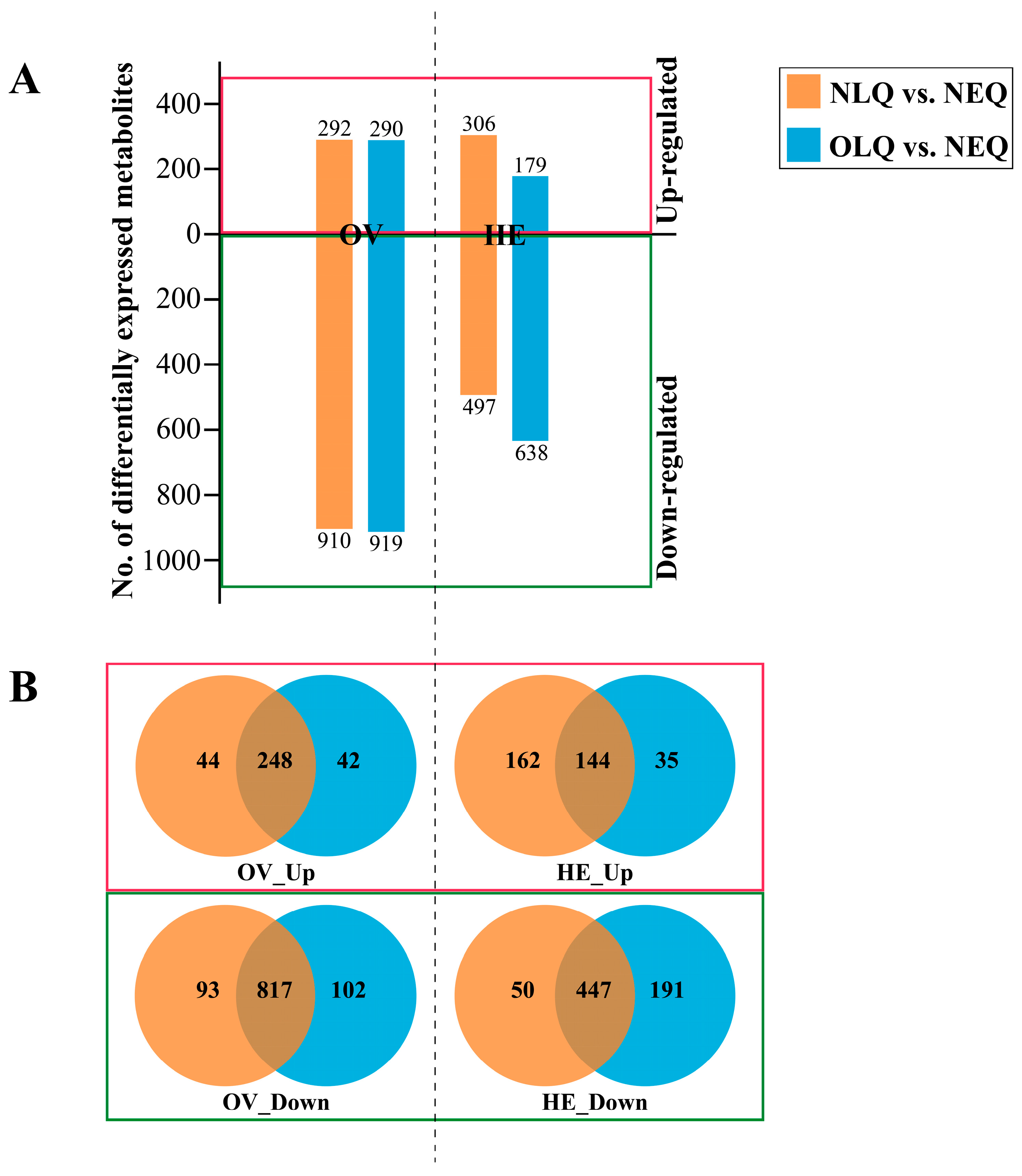
3.2. DEMs in Ovary and Hemolymph
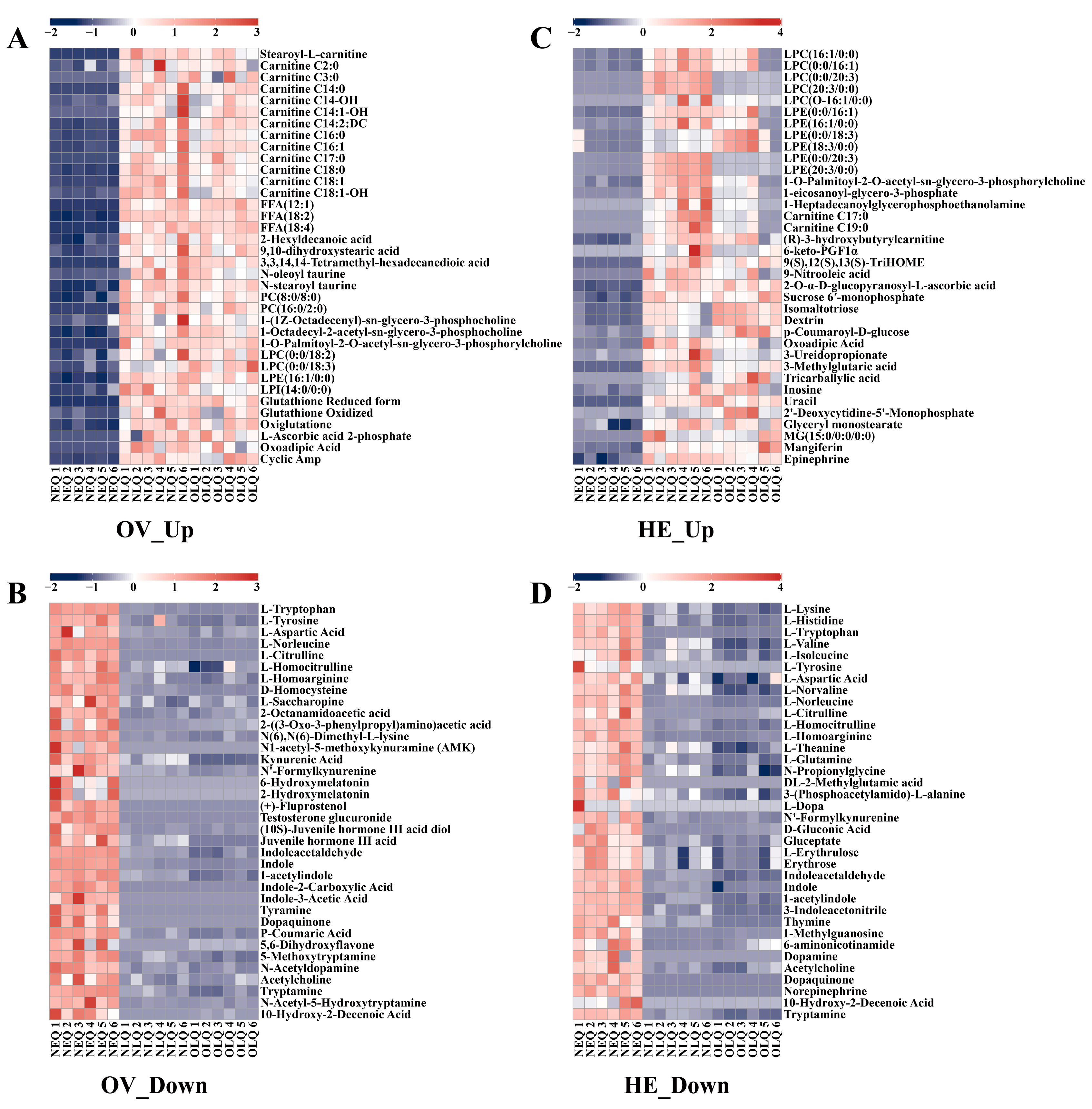
3.3. Analysis of Key Metabolites with Consistent Trends
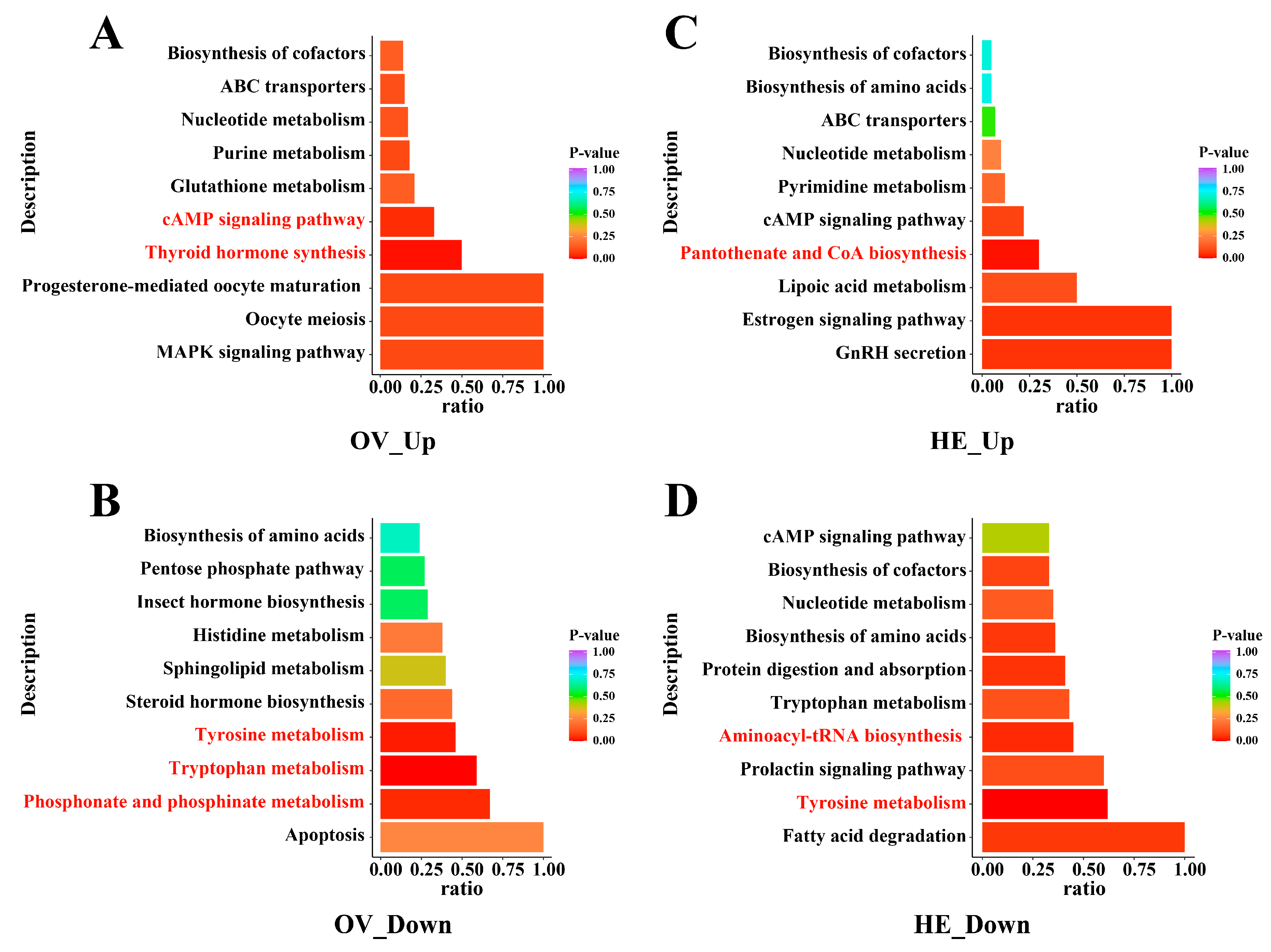
3.4. Important Metabolic Pathways Affected by Metabolite Variations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Snodgrass, R.E. Anatomy of the Honey Bee; Cornell University Press: Ithaca, NY, USA, 1956. [Google Scholar]
- Büchler, R.; Andonov, S.; Bienefeld, K.; Costa, C.; Hatjina, F.; Kezic, N.; Kryger, P.; Spivak, M.; Uzunov, A.; Wilde, J. Standard methods for rearing and selection of Apis mellifera queens. J. Apic. Res. 2013, 52, 1–30. [Google Scholar] [CrossRef]
- Kumar, N. Evaluation of larval grafted queen and natural reared queen of Italian honey bees (Apis mellifera L.). J. Pharmacogn. Phytochem. 2018, 7, 3181–3183. [Google Scholar]
- Woyke, J. Correlations between the age at which honeybee brood was grafted, characteristics of the resultant queens, and results of insemination. J. Apic. Res. 1971, 10, 45–55. [Google Scholar] [CrossRef]
- Yi, Y.; Liu, Y.-B.; Barron, A.B.; Zeng, Z.-J. Effects of commercial queen rearing methods on queen fecundity and genome methylation. Apidologie 2021, 52, 282–291. [Google Scholar] [CrossRef]
- Al-Ghzawi, A.A.M.; Zaitoun, S. Origin and rearing season of honeybee queens affect some of their physiological and reproductive characteristics. Entomol. Res. 2008, 38, 139–148. [Google Scholar] [CrossRef]
- Taha, E.-K.A.; Al-Kahtani, S.N. The relationship between comb age and performance of honey bee (Apis mellifera) colonies. Saudi J. Biol. Sci. 2020, 27, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Taha, E.-K.A.; Shawer, M.B.; Taha, R.; Elashmawy, A.; Gaber, S.; Mousa, K. Comb age significantly influences the emergency queen rearing, morphometric and reproductive characteristics of the queens. J. Apic. Res. 2024. [Google Scholar] [CrossRef]
- Tarpy, D.R. Genetic diversity within honeybee colonies prevents severe infections and promotes colony growth. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Mattila, H.R.; Seeley, T.D. Genetic diversity in honey bee colonies enhances productivity and fitness. Science 2007, 317, 362–364. [Google Scholar] [CrossRef] [PubMed]
- Simeunovic, P.; Stevanovic, J.; Cirkovic, D.; Radojicic, S.; Lakic, N.; Stanisic, L.; Stanimirovic, Z. Nosema ceranae and queen age influence the reproduction and productivity of the honey bee colony. J. Apic. Res. 2014, 53, 545–554. [Google Scholar] [CrossRef]
- Amiri, E.; Strand, M.K.; Rueppell, O.; Tarpy, D.R. Queen quality and the impact of honey bee diseases on queen health: Potential for interactions between two major threats to colony health. Insects 2017, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.T.; Tarpy, D.R.; Fahrbach, S.E. Histological estimates of ovariole number in honey bee queens, Apis mellifera, reveal lack of correlation with other queen quality measures. J. Insect Sci. 2011, 11, 82. [Google Scholar] [CrossRef] [PubMed]
- Alqarni, A.S.; Balhareth, H.M.; Owayss, A.A. Queen morphometric and reproductive characters of Apis mellifera jemenitica, a native honey bee to Saudi Arabia. Bull. Insectol. 2013, 66, 239–244. [Google Scholar]
- Gilley, D.C.; Tarpy, D.R.; Land, B.B. Effect of queen quality on interactions between workers and dueling queens in honeybee (Apis mellifera L.) colonies. Behav. Ecol. Sociobiol. 2003, 55, 190–196. [Google Scholar] [CrossRef]
- Gregorc, A.; Škerl, M.I.S. Characteristics of honey bee (Apis mellifera carnica, Pollman 1879) queens reared in Slovenian commercial breeding stations. J. Apic. Sci. 2015, 59, 5–12. [Google Scholar] [CrossRef]
- Martins, J.R.; Nunes, F.M.; Cristino, A.S.; Simões, Z.L.; Bitondi, M.M. The four hexamerin genes in the honey bee: Structure, molecular evolution and function deduced from expression patterns in queens, workers and drones. BMC Mol. Biol. 2010, 11, 23. [Google Scholar] [CrossRef]
- Martins, J.R.; Bitondi, M.M.G. The HEX 110 hexamerin is a cytoplasmic and nucleolar protein in the ovaries of Apis mellifera. PLoS ONE 2016, 11, e0151035. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Qu, Y.; Dong, K.; He, S.; Jie, W.; Huang, J. Characterization and developmental expression patterns of four hexamerin genes in the bumble bee, Bombus terrestris (Hymenoptera: Apidae). J. Insect Sci. 2021, 21, 13. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.R.; Nunes, F.M.F.; Simões, Z.L.P.; Bitondi, M.M.G. A honeybee storage protein gene, hex 70a, expressed in developing gonads and nutritionally regulated in adult fat body. J. Insect Physiol. 2008, 54, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.R.; Anhezini, L.; Dallacqua, R.P.; Simoes, Z.L.; Bitondi, M.M. A honey bee hexamerin, HEX 70a, is likely to play an intranuclear role in developing and mature ovarioles and testioles. PLoS ONE 2011, 6, e29006. [Google Scholar] [CrossRef] [PubMed]
- Amdam, G.V.; Simões, Z.L.; Hagen, A.; Norberg, K.; Schrøder, K.; Mikkelsen, Ø.; Kirkwood, T.B.; Omholt, S.W. Hormonal control of the yolk precursor vitellogenin regulates immune function and longevity in honeybees. Exp. Gerontol. 2004, 39, 767–773. [Google Scholar] [CrossRef]
- Corona, M.; Velarde, R.A.; Remolina, S.; Moran-Lauter, A.; Wang, Y.; Hughes, K.A.; Robinson, G.E. Vitellogenin, juvenile hormone, insulin signaling, and queen honey bee longevity. Proc. Natl. Acad. Sci. USA 2007, 104, 7128–7133. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.C. The Circulatory System of Insects; Charles C. Thomas: Springfield, IL, USA, 1977. [Google Scholar]
- Hillyer, J.F.; Pass, G. The insect circulatory system: Structure, function, and evolution. Annu. Rev. Entomol. 2020, 65, 121–143. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.-I.; Moeller, F. Comparison of the free amino acid composition in the hemolymph of healthy and Nosema-infected female honey bees. J. Invertebr. Pathol. 1970, 15, 202–206. [Google Scholar] [CrossRef]
- Mayack, C.; Naug, D. Parasitic infection leads to decline in hemolymph sugar levels in honeybee foragers. J. Insect Physiol. 2010, 56, 1572–1575. [Google Scholar] [CrossRef] [PubMed]
- Aliferis, K.A.; Copley, T.; Jabaji, S. Gas chromatography–mass spectrometry metabolite profiling of worker honey bee (Apis mellifera L.) hemolymph for the study of Nosema ceranae infection. J. Insect Physiol. 2012, 58, 1349–1359. [Google Scholar] [CrossRef] [PubMed]
- Burritt, N.L.; Foss, N.J.; Neeno-Eckwall, E.C.; Church, J.O.; Hilger, A.M.; Hildebrand, J.A.; Warshauer, D.M.; Perna, N.T.; Burritt, J.B. Sepsis and hemocyte loss in honey bees (Apis mellifera) infected with Serratia marcescens strain sicaria. PLoS ONE 2016, 11, e0167752. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Li, J.; Zhang, Y.; Li, Z.; Evans, J.D.; Rose, R.; Gilligan, T.M.; LeBrun, A.; He, N.; Zheng, T. A novel method for the detection and diagnosis of virus infections in honey bees. J. Virol. Methods 2021, 293, 114163. [Google Scholar] [CrossRef] [PubMed]
- Ilijević, K.; Vujanović, D.; Orčić, S.; Purać, J.; Kojić, D.; Zarić, N.; Gržetić, I.; Blagojević, D.P.; Čelić, T.V. Anthropogenic influence on seasonal and spatial variation in bioelements and non-essential elements in honeybees and their hemolymph. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 239, 108852. [Google Scholar] [CrossRef]
- Horvatinec, J.; Svečnjak, L. Infrared (FTIR) spectral features of honey bee (Apis mellifera L.) hemolymph. J. Cent. Eur. Agric. 2020, 21, 37–41. [Google Scholar] [CrossRef]
- Sappington, T.W.; Raikhel, A.S. Molecular characteristics of insect vitellogenins and vitellogenin receptors. Insect Biochem. Mol. Biol. 1998, 28, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Guidugli, K.R.; Piulachs, M.D.; Bellés, X.; Lourenço, A.P.; Simões, Z.L. Vitellogenin expression in queen ovaries and in larvae of both sexes of Apis mellifera. Arch. Insect Biochem. Physiol. Publ. Collab. Entomol. Soc. Am. 2005, 59, 211–218. [Google Scholar] [CrossRef]
- Excels, W. Occurrence and significance of vitellogenins in female castes of social Hymenoptera. Am. Zool. 1974, 14, 1229–1237. [Google Scholar] [CrossRef]
- Fleig, R. Role of the follicle cells for yolk uptake in ovarian follicles of the honey bee Apis mellifera L. (Hymenoptera: Apidae). Int. J. Insect Morphol. Embryol. 1995, 24, 427–433. [Google Scholar] [CrossRef]
- Jung, J. Metabolomic Studies in Apis mellifera. J. Apic. 2023, 38, 151–162. [Google Scholar] [CrossRef]
- Shi, T.; Burton, S.; Wang, Y.; Xu, S.; Zhang, W.; Yu, L. Metabolomic analysis of honey bee, Apis mellifera L. response to thiacloprid. Pestic. Biochem. Physiol. 2018, 152, 17–23. [Google Scholar] [CrossRef]
- Gao, J.; Yang, Y.; Ma, S.; Liu, F.; Wang, Q.; Wang, X.; Wu, Y.; Zhang, L.; Liu, Y.; Diao, Q. Combined transcriptome and metabolite profiling analyses provide insights into the chronic toxicity of carbaryl and acetamiprid to Apis mellifera larvae. Sci. Rep. 2022, 12, 16898. [Google Scholar] [CrossRef]
- Wang, B.; Habermehl, C.; Jiang, L. Metabolomic analysis of honey bee (Apis mellifera L.) response to glyphosate exposure. Mol. Omics 2022, 18, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Shi, J.; Yu, L.; Wu, X. Metabolic profiling of Apis mellifera larvae treated with sublethal acetamiprid doses. Ecotoxicol. Environ. Saf. 2023, 254, 114716. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-L.; Zhou, C.-X.; Wu, P.-J.; Xu, J.; Guo, Y.-Q.; Xue, F.; Getachew, A.; Xu, S.-F. Brain metabolomic profiling of eastern honey bee (Apis cerana) infested with the mite Varroa destructor. PLoS ONE 2017, 12, e0175573. [Google Scholar] [CrossRef]
- Jousse, C.; Dalle, C.; Abila, A.; Traïkia, M.; Diogon, M.; Lyan, B.; El Alaoui, H.; Vidau, C.; Delbac, F. A combined LC-MS and NMR approach to reveal metabolic changes in the hemolymph of honeybees infected by the gut parasite Nosema ceranae. J. Invertebr. Pathol. 2020, 176, 107478. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hou, M.; Qiu, Y.; Zhao, B.; Nie, H.; Su, S. Changes in antioxidant enzymes activity and metabolomic profiles in the guts of honey bee (Apis mellifera) larvae infected with Ascosphaera apis. Insects 2020, 11, 419. [Google Scholar] [CrossRef] [PubMed]
- Ricigliano, V.A.; Cank, K.B.; Todd, D.A.; Knowles, S.L.; Oberlies, N.H. Metabolomics-guided comparison of pollen and microalgae-based artificial diets in honey bees. J. Agric. Food Chem. 2022, 70, 9790–9801. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Ding, G.; Jia, G.; Feng, M.; Huang, J. Hemolymph Metabolism Analysis of Honey Bee (Apis mellifera L.) Response to Different Bee Pollens. Insects 2023, 14, 37. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, F.; Li, G.; Chi, X.; Wang, Y.; Wang, H.; Ma, L.; Han, K.; Zhao, G.; Guo, X. Metabolite support of long-term storage of sperm in the spermatheca of honeybee (Apis mellifera) queens. Front. Physiol. 2020, 11, 574856. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-L.; Huang, Q.; Li, J.-L.; Wu, P.; Wei, B.; Li, X.-J.; Tang, Q.-H.; Dong, Z.-X.; Xiong, J.; Tang, H. Gut microbiota-driven regulation of queen bee ovarian metabolism. Microbiol. Spectr. 2023, 11, e02145-23. [Google Scholar] [CrossRef]
- Kostecki, R. Investigation on the Haemocytes and Haemolymph of Honeybees. J. Apic. Res. 1965, 4, 49–54. [Google Scholar] [CrossRef]
- Garrido, P.M.; Martin, M.L.; Negri, P.; Eguaras, M.J. A standardized method to extract and store haemolymph from Apis mellifera and the ectoparasite Varroa destructor for protein analysis. J. Apic. Res. 2013, 52, 67–68. [Google Scholar] [CrossRef]
- Borsuk, G.; Ptaszyńska, A.A.; Olszewski, K.; Domaciuk, M.; Krutmuang, P.; Paleolog, J. A new method for quick and easy hemolymph collection from apidae adults. PLoS ONE 2017, 12, e0170487. [Google Scholar] [CrossRef] [PubMed]
- Fritz, I.B. Carnitine and its role in fatty acid metabolism. Adv. Lipid Res. 1963, 1, 285–334. [Google Scholar] [PubMed]
- Bremer, J. Carnitine--metabolism and functions. Physiol. Rev. 1983, 63, 1420–1480. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.-Q.; Jia, B.-Y.; Li, J.-J.; Fu, X.-W.; Zhou, G.-B.; Hou, Y.-P.; Zhu, S.-E. L-carnitine enhances oocyte maturation and development of parthenogenetic embryos in pigs. Theriogenology 2011, 76, 785–793. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Lee, J.; Hyun, S.-H.; Lee, E. L-carnitine treatment during oocyte maturation improves in vitro development of cloned pig embryos by influencing intracellular glutathione synthesis and embryonic gene expression. Theriogenology 2012, 78, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Phongnimitr, T.; Liang, Y.; Srirattana, K.; Panyawai, K.; Sripunya, N.; Treetampinich, C.; Parnpai, R. Effect of L-carnitine on maturation, cryo-tolerance and embryo developmental competence of bovine oocytes. Anim. Sci. J. 2013, 84, 719–725. [Google Scholar] [CrossRef]
- Knitlova, D.; Hulinska, P.; Jeseta, M.; Hanzalova, K.; Kempisty, B.; Machatkova, M. Supplementation of l-carnitine during in vitro maturation improves embryo development from less competent bovine oocytes. Theriogenology 2017, 102, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Dunning, K.R.; Akison, L.K.; Russell, D.L.; Norman, R.J.; Robker, R.L. Increased beta-oxidation and improved oocyte developmental competence in response to l-carnitine during ovarian in vitro follicle development in mice. Biol. Reprod. 2011, 85, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Moawad, A.R.; Xu, B.; Tan, S.L.; Taketo, T. L-carnitine supplementation during vitrification of mouse germinal vesicle stage–oocytes and their subsequent in vitro maturation improves meiotic spindle configuration and mitochondrial distribution in metaphase II oocytes. Hum. Reprod. 2014, 29, 2256–2268. [Google Scholar] [CrossRef]
- Li, L.; Zhu, S.; Shu, W.; Guo, Y.; Guan, Y.; Zeng, J.; Wang, H.; Han, L.; Zhang, J.; Liu, X. Characterization of metabolic patterns in mouse oocytes during meiotic maturation. Mol. Cell 2020, 80, 525–540.e529. [Google Scholar] [CrossRef]
- Geer, B.; Dolph, W. A dietary choline requirement for egg production in Drosophila melanogaster. Reproduction 1970, 21, 9–15. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gülçin, I. Antioxidant and antiradical activities of L-carnitine. Life Sci. 2006, 78, 803–811. [Google Scholar] [CrossRef]
- Solarska, K.; Lewińska, A.; Karowicz-Bilińska, A.; Bartosz, G. The antioxidant properties of carnitine in vitro. Cell. Mol. Biol. Lett. 2010, 15, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Choi, A.; Hope, Y.Y.; Czerniak, S.M.; Holick, E.A.; Paolella, L.J.; Agarwal, A.; Combelles, C.M. Fluctuations in total antioxidant capacity, catalase activity and hydrogen peroxide levels of follicular fluid during bovine folliculogenesis. Reprod. Fertil. Dev. 2011, 23, 673–680. [Google Scholar] [CrossRef]
- Park, Y.S.; You, S.Y.; Cho, S.; Jeon, H.-J.; Lee, S.; Cho, D.-H.; Kim, J.-S.; Oh, J.S. Eccentric localization of catalase to protect chromosomes from oxidative damages during meiotic maturation in mouse oocytes. Histochem. Cell Biol. 2016, 146, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Corona de Lau, C. Correlation between follicle levels of superoxide dismutase and oocyte quality, fertilization rates and embryo development. Ginecol. Obstet. Mex. 2004, 72, 335–344. [Google Scholar]
- Tatemoto, H.; Muto, N.; Sunagawa, I.; Shinjo, A.; Nakada, T. Protection of porcine oocytes against cell damage caused by oxidative stress during in vitro maturation: Role of superoxide dismutase activity in porcine follicular fluid. Biol. Reprod. 2004, 71, 1150–1157. [Google Scholar] [CrossRef] [PubMed]
- Meijide, S.; Hernández, M.L.; Navarro, R.; Larreategui, Z.; Ferrando, M.; Ruiz-Sanz, J.I.; Ruiz-Larrea, M.B. Glutathione S-transferase activity in follicular fluid from women undergoing ovarian stimulation: Role in maturation. Free Radic. Biol. Med. 2014, 75, S41. [Google Scholar] [CrossRef] [PubMed]
- Thaker, R.; Mishra, V.; Gor, M.; Agarwal, R.; Sheth, H.; Kapadia, P.; Kumar, S. The role of stimulation protocol, number of oocytes retrieved with respect to follicular fluid oxidative stress and IVF outcome. Hum. Fertil. 2020, 23, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Barros, F.D.d.A.; Adona, P.R.; Guemra, S.; Damião, B.C.M. Oxidative homeostasis in oocyte competence for in vitro embryo development. Anim. Sci. J. 2019, 90, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Hao, Y.; Liu, Z.; Li, S.; Wang, C.; Wang, B.; Liu, Y.; Liu, G.; Dai, Y. Effect of exogenous glutathione supplementation on the in vitro developmental competence of ovine oocytes. Theriogenology 2021, 173, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Kere, M.; Siriboon, C.; Lo, N.-W.; Nguyen, N.T.; Ju, J.-C. Ascorbic acid improves the developmental competence of porcine oocytes after parthenogenetic activation and somatic cell nuclear transplantation. J. Reprod. Dev. 2013, 59, 78–84. [Google Scholar] [CrossRef]
- Yu, X.-X.; Liu, Y.-H.; Liu, X.-M.; Wang, P.-C.; Liu, S.; Miao, J.-K.; Du, Z.-Q.; Yang, C.-X. Ascorbic acid induces global epigenetic reprogramming to promote meiotic maturation and developmental competence of porcine oocytes. Sci. Rep. 2018, 8, 6132. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Takasaki, A.; Miwa, I.; Taniguchi, K.; Maekawa, R.; Asada, H.; Taketani, T.; Matsuoka, A.; Yamagata, Y.; Shimamura, K. Oxidative stress impairs oocyte quality and melatonin protects oocytes from free radical damage and improves fertilization rate. J. Pineal Res. 2008, 44, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Kizuka, F.; Lee, L.; Tamura, I.; Maekawa, R.; Aasada, H.; Yamagata, Y. The role of melatonin as an antioxidant in the follicle. J. Ovarian Res. 2012, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, K.; Itoh, M.T.; Takahashi, N.; Tarumi, W.; Ishizuka, B. The rat oocyte synthesises melatonin. Reprod. Fertil. Dev. 2013, 25, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.; Williams, V.; Evans, J. Sperm storage and antioxidative enzyme expression in the honey bee, Apis mellifera. Insect Mol. Biol. 2004, 13, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Weirich, G.F.; Collins, A.M.; Williams, V.P. Antioxidant enzymes in the honey bee, Apis mellifera. Apidologie 2002, 33, 3–14. [Google Scholar] [CrossRef]
- Herbert, E., Jr.; Vanderslice, J.; Higgs, D. Effect of dietary vitamin C levels on the rate of brood production of free-flying and confined colonies of honey bees. Apidologie 1985, 16, 385–394. [Google Scholar] [CrossRef]
- Farjan, M.; Dmitryjuk, M.; Lipiński, Z.; Biernat-Łopieńska, E.; Żółtowska, K. Supplementation of the honey bee diet with vitamin C: The effect on the antioxidative system of Apis mellifera carnica brood at different stages. J. Apic. Res. 2012, 51, 263–270. [Google Scholar] [CrossRef]
- Farjan, M.; Łopieńska-Biernat, E.; Lipiński, Z.; Dmitryjuk, M.; Żółtowska, K. Supplementing with vitamin C the diet of honeybees (Apis mellifera carnica) parasitized with Varroa destructor: Effects on antioxidative status. Parasitology 2014, 141, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, M.; Arai, N.; Kohno, K.; Ushio, S.; Fukuda, S. Anti-oxidative and anti-aging activities of 2-O-α-glucopyranosyl-L-ascorbic acid on human dermal fibroblasts. Eur. J. Pharmacol. 2012, 674, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Hanada, Y.; Iomori, A.; Ishii, R.; Gohda, E.; Tai, A. Protection of free radical-induced cytotoxicity by 2-O-α-d-glucopyranosyl-l-ascorbic acid in human dermal fibroblasts. Biosci. Biotechnol. Biochem. 2014, 78, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Hata, R.I.; Senoo, H. L-ascorbic acid 2-phosphate stimulates collagen accumulation, cell proliferation, and formation of a three-dimensional tissuelike substance by skin fibroblasts. J. Cell. Physiol. 1989, 138, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Saika, S.; Kanagawa, R.; Uenoyama, K.; Hiroi, K.; Hiraoka, J.-I. L-ascorbic acid 2-phosphate, a phosphate derivative of L-ascorbic acid, enhances the growth of cultured rabbit keratocytes. Graefe’s Arch. Clin. Exp. Ophthalmol. 1991, 229, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Shima, N.; Kimoto, M.; Yamaguchi, M.; Yamagami, S. Increased proliferation and replicative lifespan of isolated human corneal endothelial cells with L-ascorbic acid 2-phosphate. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8711–8717. [Google Scholar] [CrossRef]
- Johnson, J.N.; Hardgrave, E.; Gill, C.; Moore, D. Absence of consistent diel rhythmicity in mated honey bee queen behavior. J. Insect Physiol. 2010, 56, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Li, G.; Zhang, X.; Wang, Y.; Wang, C.; Xu, B.; Guo, X.; Li, H. The role of melatonin and Tryptophan-5-hydroxylase-1 in different abiotic stressors in Apis cerana cerana. J. Insect Physiol. 2021, 128, 104180. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Duan, J.; Chen, L.; Wang, Y.; Qin, Q.; Dang, X.; Zhou, Z. Melatonin enhances the antioxidant capacity to rescue the honey bee Apis mellifera from the ecotoxicological effects caused by environmental imidacloprid. Ecotoxicol. Environ. Saf. 2022, 239, 113622. [Google Scholar] [CrossRef] [PubMed]
- Finocchiaro, L.; Callebert, J.; Launay, J.; Jallon, J. Melatonin biosynthesis in Drosophila: Its nature and its effects. J. Neurochem. 1988, 50, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Estienne, M.; Barb, C. The control of adenohypophysial hormone secretion by amino acids and peptides in swine. Domest. Anim. Endocrinol. 2005, 29, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yin, Y.-L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Grohmann, U.; Bronte, V. Control of immune response by amino acid metabolism. Immunol. Rev. 2010, 236, 243–264. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Dai, Z.; Li, D.; Wang, J.; Wu, Z. Amino acid nutrition in animals: Protein synthesis and beyond. Annu. Rev. Anim. Biosci. 2014, 2, 387–417. [Google Scholar] [CrossRef] [PubMed]
- De Groot, A. Amino acid requirements for growth of the honeybee (Apis mellifica L.). Experientia 1952, 8, 192–194. [Google Scholar] [CrossRef]
- De Groot, A.P. Protein and amino acid requirements of the honeybee (Apis mellifica L.). Physiol. Comp. Oecol. 1953, 3, 197–285. [Google Scholar]
- Hrassnigg, N.; Leonhard, B.; Crailsheim, K. Free amino acids in the haemolymph of honey bee queens (Apis mellifera L.). Amino Acids 2003, 24, 205–212. [Google Scholar] [CrossRef]
- Sang, J.H.; King, R.C. Nutritional requirements of axenically cultured Drosophila melanogaster adults. J. Exp. Biol. 1961, 38, 793–809. [Google Scholar] [CrossRef]
- Alves, A.N.; Sgrò, C.M.; Piper, M.D.; Mirth, C.K. Target of Rapamycin Drives Unequal Responses to Essential Amino Acid Depletion for Egg Laying in Drosophila Melanogaster. Front. Cell Dev. Biol. 2022, 10, 822685. [Google Scholar] [CrossRef] [PubMed]
- Boselli, E.; Caboni, M.F.; Sabatini, A.G.; Marcazzan, G.L.; Lercker, G. Determination and changes of free amino acids in royal jelly during storage. Apidologie 2003, 34, 129–137. [Google Scholar] [CrossRef]
- Xue, X.; Wu, L.; Wang, K. Chemical Composition of Royal Jelly. In Bee Products—Chemical and Biological Properties; Springer: Cham, Switzerland, 2017; pp. 181–190. [Google Scholar]
- Harlow, P.M. A study of ovarial development and its relation to adult nutrition in the blowfly Protophormia terrae-novae (RD). J. Exp. Biol. 1956, 33, 777–797. [Google Scholar] [CrossRef]
- Kobayashi, K.; Maezawa, T.; Tanaka, H.; Onuki, H.; Horiguchi, Y.; Hirota, H.; Ishida, T.; Horiike, K.; Agata, Y.; Aoki, M. The identification of d-tryptophan as a bioactive substance for postembryonic ovarian development in the planarian Dugesia ryukyuensis. Sci. Rep. 2017, 7, 45175. [Google Scholar] [CrossRef] [PubMed]
- Maezawa, T.; Ishikawa, M.; Sekii, K.; Nagamatsu, G.; Furukawa, R.; Kobayashi, K. D-Tryptophan enhances the reproductive organ-specific expression of the amino acid transporter homolog Dr-SLC38A9 involved in the sexual induction of planarian Dugesia ryukyuensis. Zool. Lett. 2021, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Christensen, B.M. Involvement of l-tyrosine and phenol oxidase in the tanning of Aedes aegypti eggs. Insect Biochem. Mol. Biol. 1993, 23, 739–748. [Google Scholar] [CrossRef]
- Zografou, E.; Tsiropoulos, G.; Margaritis, L. Effect of phenylalanine and tyrosine analogues on Bactrocera oleae Gmelin (Dipt., Tephritidae) reproduction. J. Appl. Entomol. 2001, 125, 365–369. [Google Scholar] [CrossRef]
- Fuchs, S.; Behrends, V.; Bundy, J.G.; Crisanti, A.; Nolan, T. Phenylalanine metabolism regulates reproduction and parasite melanization in the malaria mosquito. PLoS ONE 2014, 9, e84865. [Google Scholar] [CrossRef] [PubMed]
- Sterkel, M.; Oliveira, P.L. Developmental roles of tyrosine metabolism enzymes in the blood-sucking insect Rhodnius prolixus. Proc. R. Soc. B Biol. Sci. 2017, 284, 20162607. [Google Scholar] [CrossRef]
- Matsuyama, S.; Nagao, T.; Sasaki, K. Consumption of tyrosine in royal jelly increases brain levels of dopamine and tyramine and promotes transition from normal to reproductive workers in queenless honey bee colonies. Gen. Comp. Endocrinol. 2015, 211, 1–8. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, S.; Pan, L.; Wang, Z.; Zeng, Z. Revealing Changes in Ovarian and Hemolymphatic Metabolites Using Widely Targeted Metabolomics between Newly Emerged and Laying Queens of Honeybee (Apis mellifera). Insects 2024, 15, 263. https://doi.org/10.3390/insects15040263
Zhong S, Pan L, Wang Z, Zeng Z. Revealing Changes in Ovarian and Hemolymphatic Metabolites Using Widely Targeted Metabolomics between Newly Emerged and Laying Queens of Honeybee (Apis mellifera). Insects. 2024; 15(4):263. https://doi.org/10.3390/insects15040263
Chicago/Turabian StyleZhong, Shiqing, Luxia Pan, Zilong Wang, and Zhijiang Zeng. 2024. "Revealing Changes in Ovarian and Hemolymphatic Metabolites Using Widely Targeted Metabolomics between Newly Emerged and Laying Queens of Honeybee (Apis mellifera)" Insects 15, no. 4: 263. https://doi.org/10.3390/insects15040263
APA StyleZhong, S., Pan, L., Wang, Z., & Zeng, Z. (2024). Revealing Changes in Ovarian and Hemolymphatic Metabolites Using Widely Targeted Metabolomics between Newly Emerged and Laying Queens of Honeybee (Apis mellifera). Insects, 15(4), 263. https://doi.org/10.3390/insects15040263