Feeding Performance of Argentine Stem Weevil Is Reduced by Peramine from Perennial Ryegrass Infected with Endophyte Fungus

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Evaluation of Endophytes in Plants of L. perenne
2.3. Isolation and Characterization of Endophytic Fungus in Lolium perenne
2.4. Alkaloid Extraction Procedure and Analysis
2.5. Insects
2.6. No-Choice Bioassay
2.7. Alkaloidal Extract Bioassays
2.8. Artificial Diet Tests
2.9. Data Analysis
3. Results
3.1. Endophyte Infection and Quantification of Peramine
3.2. No-Choice Bioassay
3.3. Alkaloid Extract Bioassays
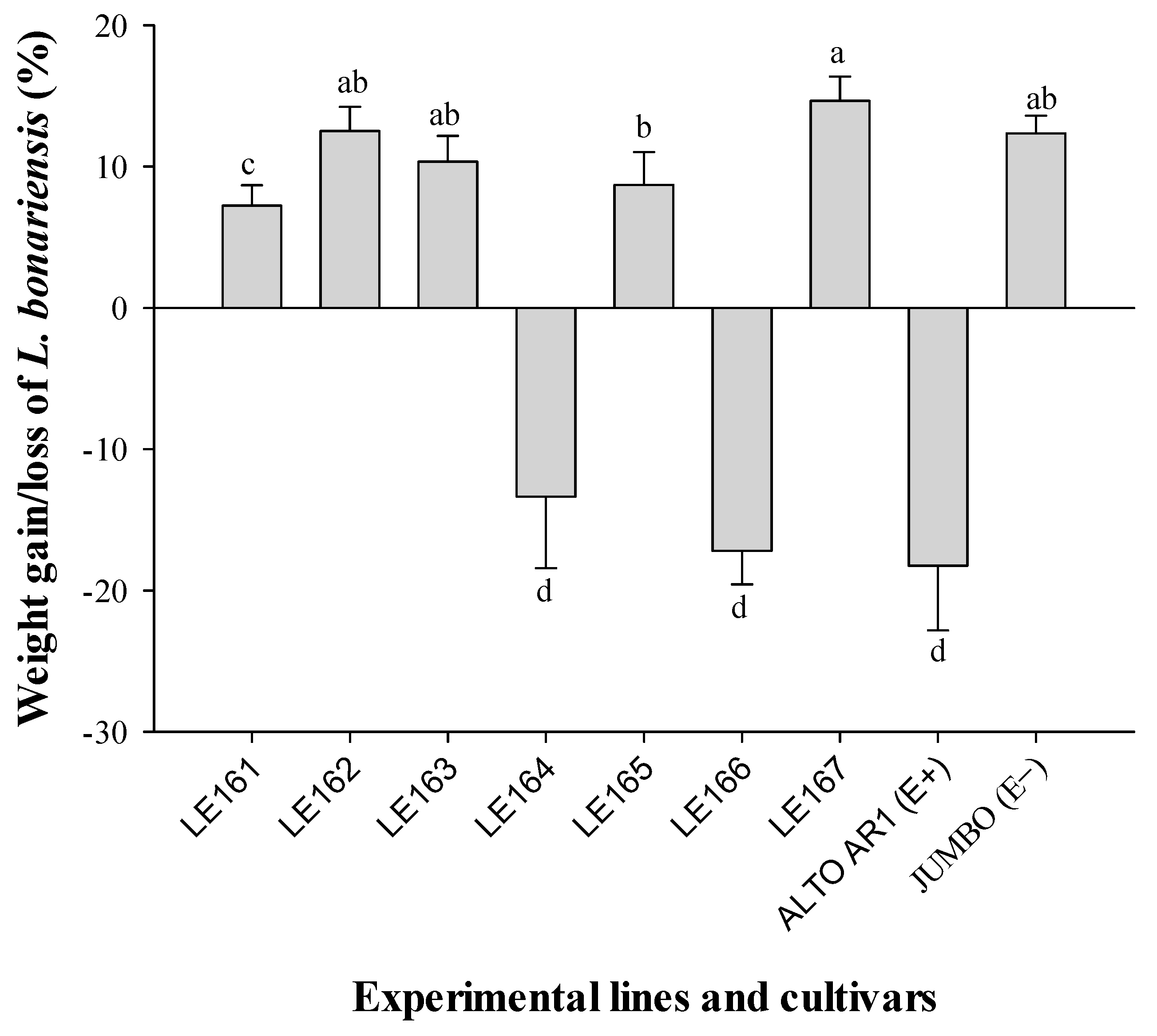
3.4. Artificial Diet Bioassays
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cardoso, A.; Longhini, V.Z.; BerÇa, A.S.; Ongaratto, F.; Siniscalchi, D.; Reis, R.A.; Ruggier, A.C. Pasture Management and Greenhouse Gases Emissions. Biosci. J. 2022, 38, e38099. [Google Scholar] [CrossRef]
- Majeed, Y.; Rasheed, A.; Awan, S.I.; Xu, Z.J.; Javed, S.O.; Gillani, S.F.A.; Wu, Z.M. Physicochemical response of Alfalfa (Medicago sativa L.) and rye grass (Lolium perenne L.) genotypes to induce drought stress at seedling stage. Appl. Ecol. Environ. Res. 2023, 21, 3335–3348. [Google Scholar] [CrossRef]
- Lasley, P.; Hogberg, M.; Helsel, Z.; Larson, A. People, grassland, and livestock in revitalized rural communities. In Grassland Quietness and Strength for a New American Agriculture; Wedin, W.F., Fales, S.L., Eds.; American Society of Agronomy, Crop Science Society of America and Soil Science Society of America: Madison, WI, USA, 2009; pp. 173–188. [Google Scholar]
- Kane, K. Effects of endophyte infection on drought stress tolerance of Lolium perenne accessions from the Mediterranean region. Environ. Exp. Bot. 2011, 71, 337–344. [Google Scholar] [CrossRef]
- Lamelas-López, L.; Borges, P.A.V.; Tarantino, E.; Juliano, M.M.; Fontes, J.C.; Moules, C.; Rodrigues, R.; Machado, J.; Mota, J.A.; Sousa, B.; et al. Monitorin ten insect pests in selected orchards in three Azorean Islands: The project CUARENTAGRI. Biodivers. Data J. 2023, 11, e100942. [Google Scholar] [CrossRef]
- Barratt, B.; Barton, D.; Philip, B.; Ferguson, C.; Goldson, S. Is the invasive species Listronotus bonariensis (Kuschel) (Coleoptera: Curculionidae) (Argentine Stem Weevil) a threat to New Zealand Natural grassland ecosystems? Front. Plant Sci. 2016, 7, 1091. [Google Scholar] [CrossRef]
- Hennessy, L.M.; Popay, A.J.; Glare, T.R.; Finch, S.C.; Cave, V.M.; Rostás, M. Olfactory responses of Argentine stem weevil to herbivory and endophyte-colonisation in perennial ryegrass. J. Pest Sci. 2022, 95, 263–277. [Google Scholar] [CrossRef]
- Norambuena, H.; Gerding, M. Presencia de Listronotus bonariensis (Kuschel) en trigo y cebada en Chile. Simiente 1985, 55, 86–88. [Google Scholar]
- Cisternas, E. Plagas claves en la producción de praderas. In Seminario Praderas: Hacia un Nuevo Estilo Productivo; Opazo, L.A., Torres, E., Siebald, E., Eds.; Serie Actas N°9; INIA Remehue: Osorno, Chile, 2001; pp. 48–57. [Google Scholar]
- Parra, L.; Chacón, M.; Lizama, M.; Quiroz, A. Incidence of Listronotus bonariensis (Coleoptera: Curculionidae) in ryegrass pastures from southern Chile. New Background. J. Soil Sci. Plant Nutr. 2017, 17, 91–98. [Google Scholar] [CrossRef]
- Shields, M.W.; Wratten, S.D.; Koten, C.V.; Phillips, C.B.; Bennet, J.R.; Goldson, S.L. Different parasitoid species elicit varied Argentine stem weevil, Listronotus bonariensis avoidance responses. N. Z. J. Agric. Res. 2023, 1–10. [Google Scholar] [CrossRef]
- Cisternas, E.; Villagra, M. Gorgojo argentino de las ballicas Listronotus bonariensis (Kushel) en la Décima Región. In El Gorgojo y el Endófito de las Ballicas en la Producción Bovina de Leche y Carne en el sur de Chile; Lanuza, F., Torres, A., Cisternas, E., Eds.; Ministerio de Agricultura; INIA Remehue: Osorno, Chile, 2003; pp. 19–51. [Google Scholar]
- Urrutia, M. The Influence of the Diet of the Argentine Stem Weevil, Listronotus bonariensis (Kuschel) (Coleoptera: Curculionidae) on the Fitness of Its Parasitoid, Microctonus Hyperodae Loan (Hymenoptera: Braconidae): Prospects for “Indirect” Conservation Biocontrol. Master’s Thesis, Lincoln University, Lincoln, New Zeland, 2005; 89p. [Google Scholar]
- Bastias, D.A.; Ueno, A.C.; Gundel, P.E. Global change factors influence Plant-epichloe Associations. J. Fungi 2023, 9, 446. [Google Scholar] [CrossRef]
- Card, S.D.; Bastías, D.A.; Caradus, J.R. Antagonism to plant pathogens by epichloë fungal endophytes—A review. Plants 2021, 10, 1997. [Google Scholar] [CrossRef] [PubMed]
- Panaccione, D.G.; Beaulieu, W.T.; Cook, D. Bioactive alkaloids in vertically transmitted fungal endophytes. Funct. Ecol. 2014, 28, 299–314. [Google Scholar] [CrossRef]
- Aniszewski, T. Alkaloids–Secrets of Life Alkaloid Chemistry, Biological Significance, Applications and Ecological Role; Elsevier: Amsterdam, The Netherlands, 2007; 316p. [Google Scholar]
- Lin, W.; Gao, C.; Wang, J.; Xu, W.; Wang, M.; Li, M.; Ma, B.; Tian, P. Effects of drought stress on peramine and lolitrem B in Epichloe-endophyte-infected perennial ryegrass. Life 2022, 12, 1207. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.J.; De Bonth, A.C.M.; Briggs, L.; Caradus, J.R.; Finch, S.C.; Fleetwood, D.J.; Fletcher, L.R.; Hume, D.E.; Johnson, R.D.; Popay, A.; et al. The exploitation of epichloae endophytes for agricultural benefit. Fungal Divers. 2013, 60, 171–188. [Google Scholar] [CrossRef]
- Vassiliadis, S.; Reddy, P.; Hemsworth, J.; Spangenberg, G.C.; Guthridge, K.M.; Rochfort, S.J. Quantitation and distribution of epichloë-derived alkaloids in perennial ryegrass tissues. Metabolites 2023, 13, 205. [Google Scholar] [CrossRef] [PubMed]
- Duringer, J.M.; Blythe, L.L.; Estill, C.T.; Moon, A.; Murty, L.; Livesay, S.; Moon, A.; Murty, L.; Livesay, S.; Galen, A.; et al. Determination of a sub-chronic threshold for lolitrem B and perennial ryegrass toxicosis in Angus cattle consuming endophyte-infected perennial ryegrass (Lolium perenne) straw over 64 days. Livest. Sci. 2021, 250, 104570. [Google Scholar] [CrossRef]
- Hennessy, L.M.; Popay, A.J.; Finch, S.C.; Clearwater, M.J.; Cave, V.M. Temperature and plant genotype alter alkaloid concentration in ryegrass infected with an Epichloë endophyte and this affects an insect herbivore. Front. Plant Sci. 2016, 7, 1097. [Google Scholar] [CrossRef]
- Dombrowski, J.; Baldwin, J.; Azevedo, M.; Banowetz, G. A sensitive PCR-based assay to detect Neotyphodium fungi in seed and plant tissue of tall fescue and ryegrass species. Crop Sci. 2006, 46, 1064–1070. [Google Scholar] [CrossRef]
- Roylance, J.T.; Hill, N.S.; Agee, C.S. Ergovaline and peramine production in endophyte-infected tall fescue: Independent regulation and effects of plant and endophyte genotype. J. Chem. Ecol. 1994, 20, 2171–2183. [Google Scholar] [CrossRef]
- Parra, L.; Mutis, A.; Chacón, M.; Lizama, M.; Rojas, C.; Catrileo, A.; Rubilar, O.; Tortella, G.; Birkett, M.A.; Quiroz, A. Horn fly larval survival in cattle dung is reduced by endophyte infection of tall fescue pasture. Pest Manag. Sci. 2016, 72, 1328–1334. [Google Scholar] [CrossRef]
- Toledo, D.; Parra, L.; Mutis, A.; Ortega, F.; Hormazábal, E.; Quiroz, A. Influence of long-chain fatty acids on feeding behavior of Hylastinus obscurus Marsham (Coleoptera: Curculionidae). Cienc. E Investig. Agrar. 2014, 41, 357–364. [Google Scholar]
- Faccoli, M.; Schlyter, F. Conifer phenolic resistance markers are bark beetle antifeedant semiochemicals. Agric. For. Entomol. 2007, 9, 237–245. [Google Scholar] [CrossRef]
- Baldauf, M.; Mace, W.; Richmond, D. Endophyte-mediated resistance to black cutworm as a function of plant cultivar and endophyte strain in tall fescue. Environ. Entomol. 2011, 40, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Yue, Q.; Johnson-Cicalese, J.; Gianfagna, T.J.; Meyer, W.A. Alkaloid production and chinch bug resistance in endophyte-inoculated chewings and strong creeping red fescues. J. Chem. Ecol. 2000, 26, 279–292. [Google Scholar] [CrossRef]
- Ball, O.J.-P.; Gwinn, K.D.; Pless, C.D.; Popay, A.J. Endophyte isolate and host grass effects on Chaetocnema pulicaria (Coleoptera: Chrysomelidae) feeding. J. Econ. Entomol. 2011, 104, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Karpyn, M.; Yen, A.; Rochfort, S.; Guthridge, K.; Powell, K.; Edwards, J.; Spangenberg, G. A review of perennial ryegrass endophytes and their potential use in the management of African black beetle in perennial grazing systems in Australia. Front. Plant Sci. 2017, 8, 3. [Google Scholar]
- Rutgers, J.A.; Clay, K. Endophyte symbiosis with tall fescue: How strong are the impacts on communities and ecosystem? Fungal Biol. Rev. 2007, 21, 107–124. [Google Scholar] [CrossRef]
- Raman, A.; Wheatly, W.; Popay, A. Endophytic fungus-vascular plant-insect interactions. Environ. Entomol. 2012, 41, 433–447. [Google Scholar] [CrossRef]
- Krauss, J.; Härri, S.A.; Bush, L.; Husi, R.; Bigler, L.; Power, S.A.; Müller, C.B. Effects of fertilizer, fungal endophytes and plant cultivar on the performance of insect herbivores and their natural enemies. Funct. Ecol. 2007, 21, 107–116. [Google Scholar] [CrossRef]
- Dougherty, C.T.; Knapp, F.W.; Bush, L.P. Mortality of horn fly larvae (Diptera: Muscidae) in bovine dung supplemented with ergotamine and N-formyl loline. J. Med. Entomol. 1999, 36, 73–77. [Google Scholar] [CrossRef]
- Ball, O.J.-P.; Prestidge, R.A.; Sprosen, J.M. Interrelationships between Acremonium lolii, peramine, and lolitrem B in perennial ryegrass. Appl. Environ. Microbiol. 1995, 61, 1527–1533. [Google Scholar] [CrossRef] [PubMed]
- Koulman, A.; Lane, G.; Christensen, M.; Fraser, K.; Tapper, B. Peramine and other fungal alkaloids are exuded in the guttation fluid of endophyte-infected grasses. Phytochemistry 2007, 68, 355–360. [Google Scholar] [CrossRef]
- Fuchs, B.; Krischke, M.; Mueller, M.; Krauss, J. Peramine and lolitrem B from endophyte-grass associations cascade up the food chain. J. Chem. Ecol. 2013, 39, 1385–1389. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.M.; Wang, R.; Li, X.D.; Jiang, Y.M.; Peng, F.; Xia, B. Alkaloid accumulation in different parts and ages of Lycoris chinensis. Z. Naturforsch. C. 2010, 65, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Krischke, M.; Mueller, M.; Krauss, J. Plant age and seasonal timing determine endophyte growth and alkaloid biosynthesis. Fungal Ecol. 2017, 29, 52–58. [Google Scholar] [CrossRef]
- Rowan, D.; Gaynor, D. Isolation of feeding deterrents against argentine stem weevil from ryegrass infected with the endophyte Acremonium loliae. J. Chem. Ecol. 1986, 12, 647–658. [Google Scholar] [CrossRef]
- Rowan, D.D.; Dymock, J.J.; Bimble, M.A. Effect of fungal metabolite peramine and analogs on feeding and development of Argentine stem weevil (Listronotus bonariensis). J. Chem. Ecol. 1990, 16, 1683–1695. [Google Scholar] [CrossRef]
- Keogh, R.G.; Tapper, B.A.; Fletcher, R.H. Distribution of the fungal endophyte Acremonium lolii, and of the alkaloids lolitrem B and peramine, within perennial ryegrass. N. Z. J. Agric. Res. 1996, 39, 121–127. [Google Scholar] [CrossRef]
- Spiering, M.J.; Davies, E.; Tapper, B.A.; Schmid, J.; Lane, G.A. Simplified extraction of ergovaline and peramine for analysis of tissue distribution in endophyte-infected grass tillers. J. Agric. Food Chem. 2002, 50, 5856–5862. [Google Scholar] [CrossRef]
- Tanaka, A.; Tapper, B.A.; Popay, A.; Parker, E.J.; Scott, B. A symbiosis expressed non-ribosomal peptide synthetase from a mutualistic fungal endophyte of perennial ryegrass confers protection to the symbiotum from insect herbivory. Mol. Microbiol. 2005, 57, 1036–1050. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line Experimental | N° of Plants Tested | (E+) | (E−) | Endophyte (%) * |
|---|---|---|---|---|
| LE166 | 120 | 120 | 0 | 100 ± 0.0 a |
| LE164 | 120 | 102 | 18 | 85 ± 0.8 b |
| LE167 | 120 | 96 | 24 | 80 ± 1.1 b |
| LE162 | 120 | 96 | 24 | 80 ± 1.1 b |
| Alto AR1 (E+) | 120 | 96 | 24 | 80 ± 1.1 b |
| LE163 | 120 | 89 | 31 | 74 ± 2.5 c |
| LE165 | 120 | 0 | 120 | 0 ± 0.0 d |
| LE161 | 120 | 0 | 120 | 0 ± 0.0 d |
| Jumbo (E−) | 120 | 0 | 120 | 0 ± 0.0 d |
| Line Experimental | Peramine (ng/g DM) * |
|---|---|
| Alto AR1 (E+) | 184.2 ± 36.1 a |
| LE164 | 60.8 ± 0.4 b |
| LE166 | 46.5 ± 1.2 c |
| LE162 | nd |
| LE161 | nd |
| LE163 | nd |
| LE165 | nd |
| LE167 | nd |
| Jumbo (E−) | nd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chacón-Fuentes, M.; Martínez-Cisterna, D.; Vera, W.; Ortega-Klose, F.; Reyes, C.; Matamala, I.; Quiroz, A.; Bardehle, L. Feeding Performance of Argentine Stem Weevil Is Reduced by Peramine from Perennial Ryegrass Infected with Endophyte Fungus. Insects 2024, 15, 410. https://doi.org/10.3390/insects15060410
Chacón-Fuentes M, Martínez-Cisterna D, Vera W, Ortega-Klose F, Reyes C, Matamala I, Quiroz A, Bardehle L. Feeding Performance of Argentine Stem Weevil Is Reduced by Peramine from Perennial Ryegrass Infected with Endophyte Fungus. Insects. 2024; 15(6):410. https://doi.org/10.3390/insects15060410
Chicago/Turabian StyleChacón-Fuentes, Manuel, Daniel Martínez-Cisterna, Waleska Vera, Fernando Ortega-Klose, Claudio Reyes, Ignacio Matamala, Andrés Quiroz, and Leonardo Bardehle. 2024. "Feeding Performance of Argentine Stem Weevil Is Reduced by Peramine from Perennial Ryegrass Infected with Endophyte Fungus" Insects 15, no. 6: 410. https://doi.org/10.3390/insects15060410