

The ZcVg3 Gene Regulates the Reproduction and Lifespan of Female Zeugodacus cucurbitae (Coquillett) Mediated by Short-Term High Temperatures

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Design and Preparation of siRNA
2.3. Injection of siRNA
2.4. qRT-PCR Analysis
2.5. Western Blot
2.6. Effect of Silencing ZcVg3 Gene on Reproduction and Lifespan of Z. cucurbitae (Coquillett)
2.7. Effect of Silencing ZcVg3 Gene on Ovarian Development of Z. cucurbitae (Coquillett)
2.8. Statistics
3. Results
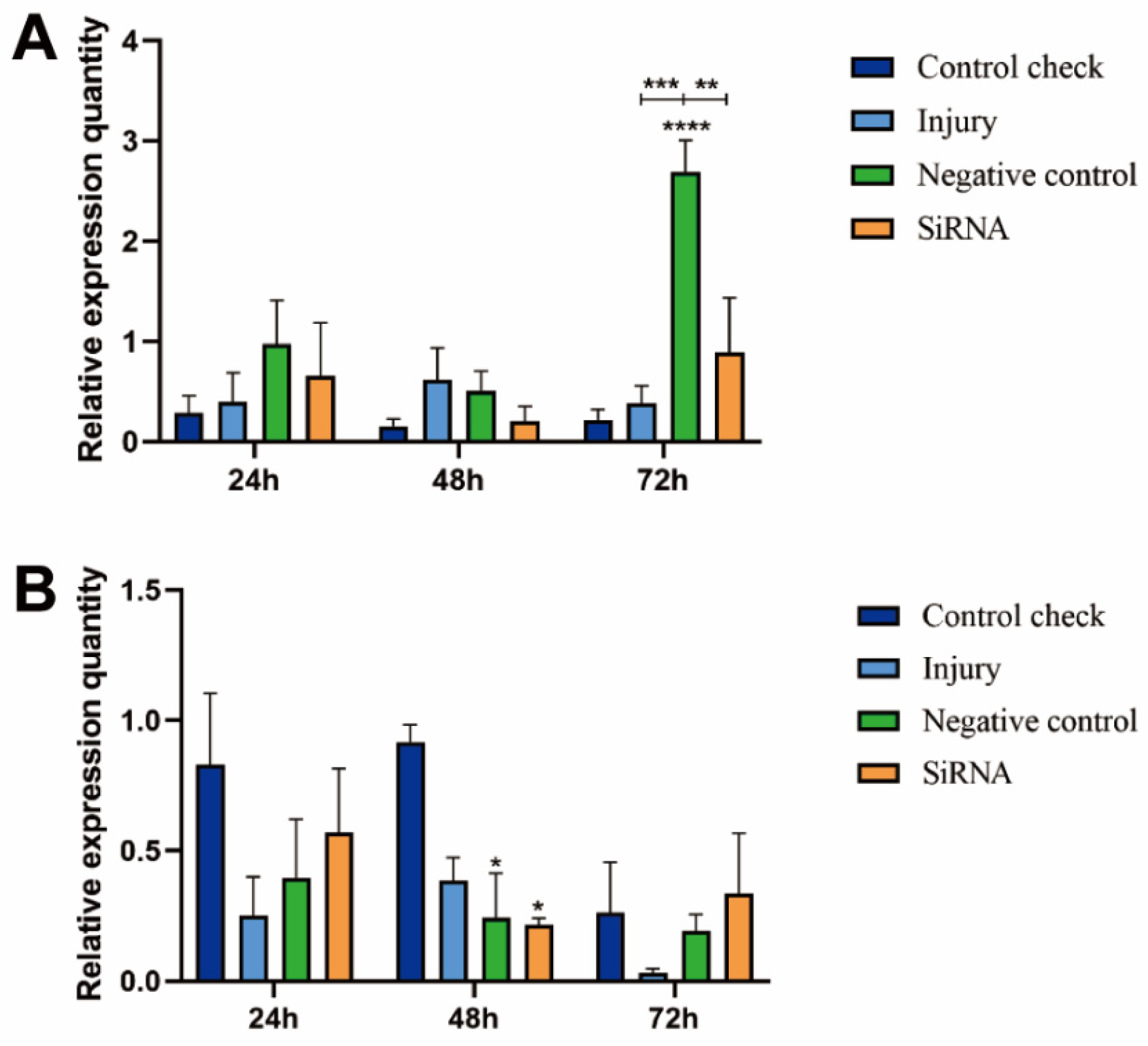
3.1. Silencing Efficiency of ZcVg3 Gene
3.2. Effect of Silencing ZcVgR Gene on Vitellogenin-3 Expression
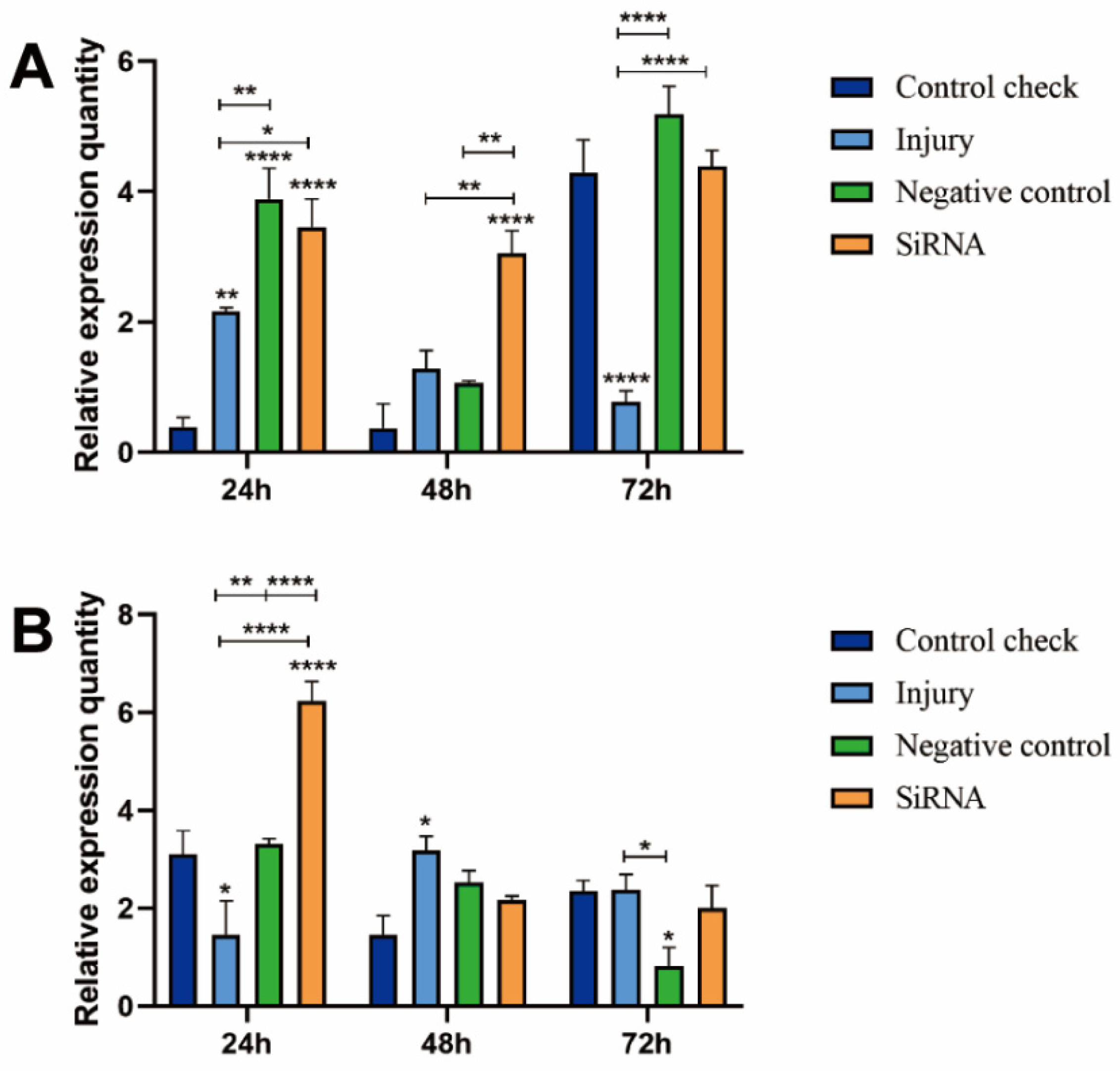
3.3. Effect of Silencing ZcVg3 Gene on ZcVgR Gene Expression
3.4. Effect of Silencing ZcVg3 Gene on ZcVg1 Gene Expression
3.5. Effect of Silencing ZcVg3 Gene on ZcVg2 Gene Expression
3.6. Effect of Silencing ZcVg3 Gene on Reproduction and Lifespan of Z. cucurbitae (Coquillett)
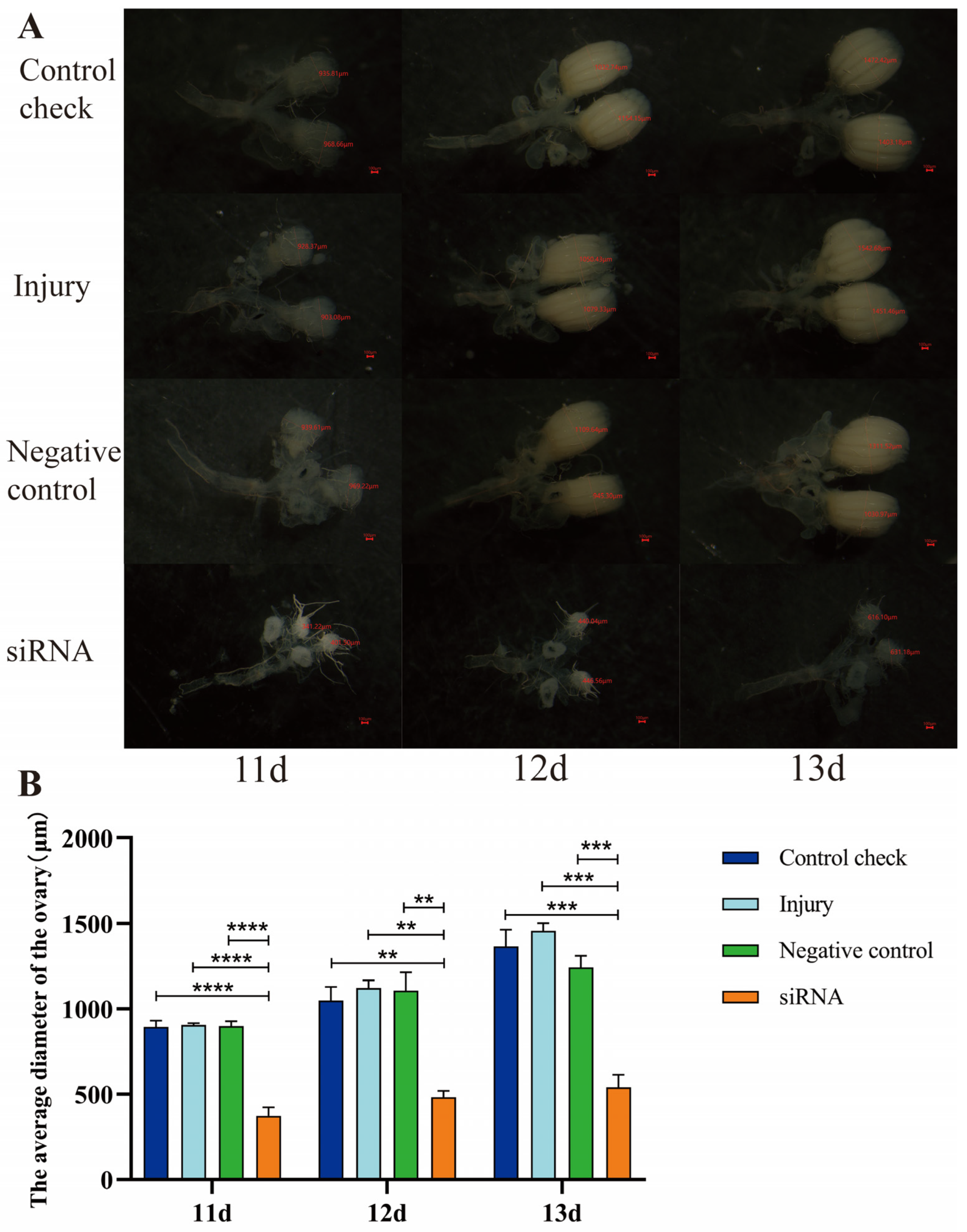
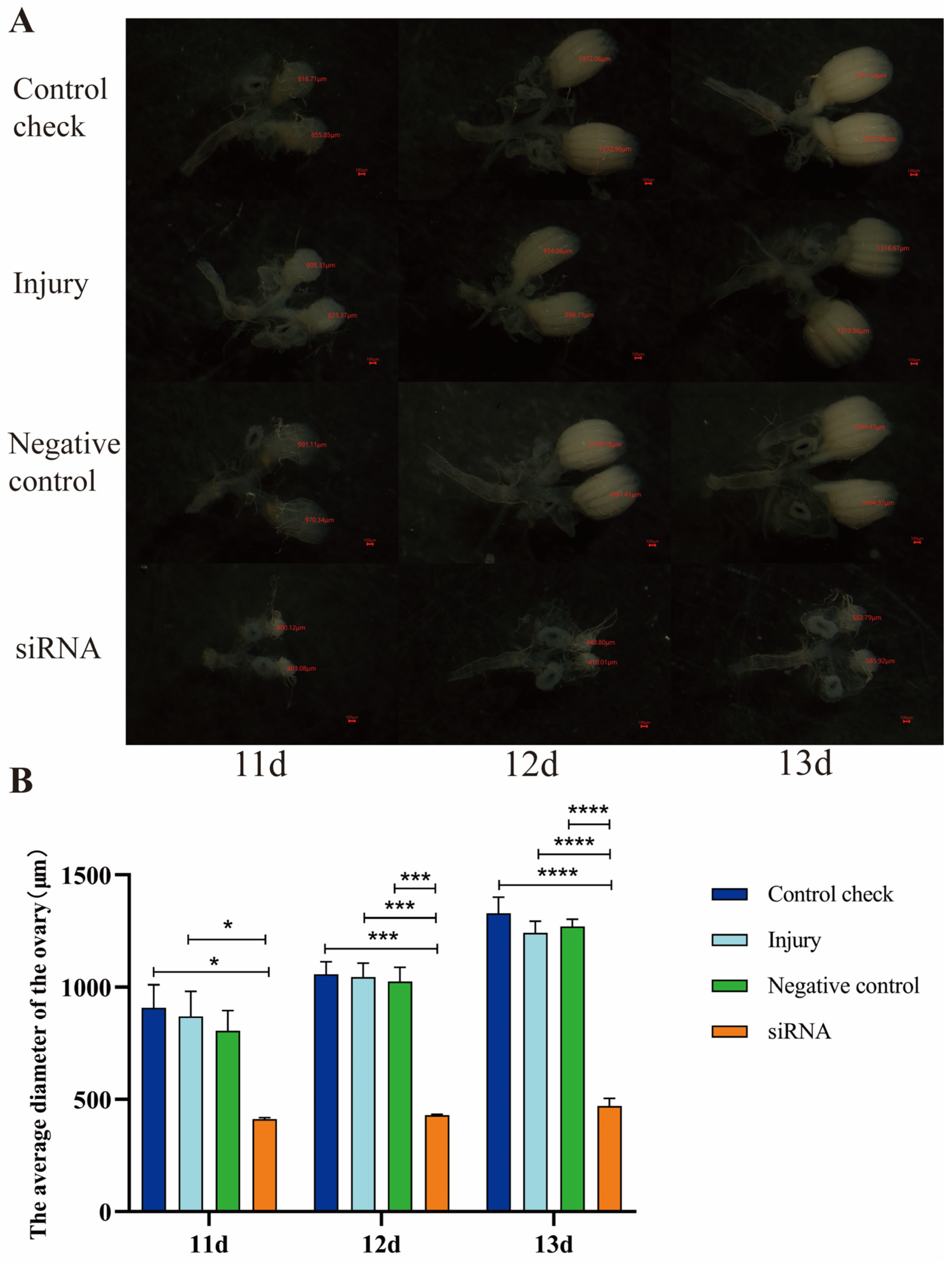
3.7. Effect of Silencing ZcVg3 Gene on Ovarian Development of Z. cucurbitae (Coquillett)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sajan, I.V.; Subramanian, S.; Sharma, K. Attractancy potential of bacterial volatiles from symbiotic bacteria of wild population of Zeugodacus cucurbitae Coquillett (Diptera: Tephritidae). Phytoparasitica 2022, 50, 411–421. [Google Scholar] [CrossRef]
- Shivaramu, S.; Parepally, S.K.; Chakravarthy, A.K.; Pagadala Damodaram, K.J.; Kempraj, V. Ridge gourd volatiles are attractive to gravid female melon fly, Zeugodacus cucurbitae (Coquillett) (Diptera: Tephritidae). J. Appl. Entomol. 2022, 146, 539–546. [Google Scholar] [CrossRef]
- Dhillon, M.K.; Singh, R.; Naresh, J.S.; Sharma, H.C. The melon fruit fly, Bactrocera cucurbitae: A review of its biology and management. J. Insect Sci. 2005, 5, 40. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Kong, Q.; Yuan, S.Y.; Shen, D.R.; Xie, K.; Wang, C.M. Effects of temperature and relative humidity stress on the survival and fecundity of the melon fly, Bactrocera cucurbitae (Diptera: Tephritidae). N. Hortic. 2019, 15, 36–43. [Google Scholar]
- Subedi, K.; Regmi, R.; Thapa, R.B.; Tiwari, S. Evaluation of net house and mulching effect on Cucurbit fruit fly (Bactrocera cucurbitae Coquillett) on cucumber (Cucumis sativus L.). J. Agric. Food Chem. 2021, 3, 100–103. [Google Scholar] [CrossRef]
- Diksha, M.E.; Singh, S.; Sohal, S.K. Potential biological control agents of Zeugodacus cucurbitae (Coquillett): A review. J. Appl. Entomol. 2022, 146, 917–929. [Google Scholar] [CrossRef]
- Zeng, B.; Zhu, W.J.; Fu, Y.G.; Zhou, S.H. Influence of high-temperature exposure on the mating, oviposition and thermotaxis of Bactrocera cucurbitae (Coquillet) (Diptera:Tephritidae). PLoS ONE 2018, 13, e0204065. [Google Scholar] [CrossRef] [PubMed]
- Meehl, G.A.; Tebaldi, C. More intense, more frequent, and longer lasting heat waves in the 21st century. Science 2004, 305, 994–997. [Google Scholar] [CrossRef]
- Alexander, L.V.; Zhang, X.; Peterson, T.C.; Caesar, J.; Gleason, B.E.; Klein Tank, A.M.G.; Haylock, M.; Collins, D.; Trewin, B.; Rahimzadeh, F.; et al. Global observed changes in daily climate extremes of temperature and precipitation. J. Geophys. Res. 2006, 111, D05109. [Google Scholar] [CrossRef]
- Jia, J.J.; Zhu, W.J.; Fu, Y.G.; Zhou, S.H. Impact of Warm-Temperature Treatments on Six Physiological Metabolism Indices of Bactrocera cucurbitae (Coquillet). Southwest Entomol. 2019, 44, 181–187. [Google Scholar] [CrossRef]
- Li, M.; Wei, X.M.; Li, J.; Wei, S.M.; Zhang, J.L.; Chen, G.H.; Zhang, X.M. Effect of short-term exposure to high temperatures on the reproductive behavior and physiological enzyme activities in the fruit fly Zeugodacus tau (Walker). Front. Physiol. 2023, 14, 1036397. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.J.; Choi, K.S.; Huang, Y.B. Thermal effects on the development of Zeugodacus cucurbitae (Coquillett) (Diptera: Tephritidae) and model validation. Phytoparasitica 2022, 50, 601–616. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Sgrò, C.M. Climate Change and Evolutionary Adaptation. Nature 2011, 470, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.S.; Ma, G.; Pincebourde, S. Survive a Warming Climate: Insect Responses to Extreme High Temperatures. Annu. Rev. Entomol. 2020, 66, 163–184. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Zhu, W.; Fu, Y.; Zhou, S.H. Response Mechanism of Oviposition and Relevant Protein Expression of Bactrocera cucurbitae (Coquillet) to Short-Term High-Temperature Conditions. Neotrop. Entomol. 2019, 48, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Lian, Y.Y.; Jia, J.J.; Liu, Y.; Wang, A.Q.; Yang, H.M.; Li, J.L.; Yang, S.Y.; Peng, S.H.; Zhou, S.H. Multigenerational Effects of Short-Term High Temperature on the Development and Reproduction of the Zeugodacus cucurbitae (Coquillett, 1899). Agriculture 2022, 12, 954. [Google Scholar] [CrossRef]
- Pan, M.L.; Bell, W.J.; Telfer, W.H. Vitellogenic blood protein synthesis by insect fat body. Science 1969, 165, 393–394. [Google Scholar] [CrossRef]
- Sappington, T.W.; Raikhel, A.S. Molecular characteristics of insect vitellogenins and vitellogenin receptors. Insect. Biochem. Mol. Biol. 1998, 28, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, D.; Zhang, M.; Chen, W.; Zhang, G. Food source affects the expression of vitellogenin and fecundity of a biological control agent, Neoseiulus cucumeris. Exp. Appl. Acarol. 2014, 63, 333–347. [Google Scholar] [CrossRef]
- Tufail, M.; Takeda, M. Insect vitellogenin/lipophorin receptors: Molecular structures, role in oogenesis, and regulatory mechanisms. J. Insect Physiol. 2009, 55, 88–104. [Google Scholar] [CrossRef]
- Corona, M.; Velarde, R.A.; Remolina, S.; Moran-Lauter, A.; Wang, Y.; Hughes, K.A.; Robinson, G.E. Vitellogenin, juvenile hormone, insulin signaling, and queen honey bee longevity. Proc. Natl. Acad. Sci. USA 2007, 104, 7128–7133. [Google Scholar] [CrossRef]
- Amdam, G.V.; Norberg, K.; Omholt, S.W.; Kryger, P.; Lourenço, A.P.; Bitondi, M.M.G.; Simões, Z.L.P. Higher vitellogenin concentrations in honey bee workers may be an adaptation to life in temperate climates. Insectes Soc. 2005, 52, 316–319. [Google Scholar] [CrossRef]
- Page, R.E., Jr.; Amdam, G.V. The making of a social insect: Developmental architectures of social design. Bioessays 2007, 29, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.M.; Ihle, K.E.; Fondrk, M.K.; Page, R.E., Jr.; Amdam, G.V. The gene vitellogenin has multiple coordinating effects on social organization. PLoS Biol. 2007, 5, e62. [Google Scholar] [CrossRef]
- Tufail, M.; Bembenek, J.; Elgendy, A.M.; Takeda, M. Evidence for two vitellogenin-related genes in Leucophaea maderae: The protein primary structure and its processing. Arch. Insect Biochem. Physiol. 2007, 66, 190–203. [Google Scholar] [CrossRef] [PubMed]
- Tufail, M.; Takeda, M. Molecular characteristics of insect vitellogenins. J. Insect Physiol. 2008, 54, 1447–1458. [Google Scholar] [CrossRef]
- Tarone, A.M.; McIntyre, L.M.; Harshman, L.G.; Nuzhdin, S.V. Genetic variation in the Yolk protein expression network of Drosophila melanogaster: Sex-biased negative correlations with longevity. Heredity 2012, 109, 226–234. [Google Scholar] [CrossRef]
- Wei, D.; Jia, H.T.; Zhang, M.Y.; Li, R.; Smagghe, G.; Wang, J.J. Comparative analysis of differential gene expression profiling of sex-bias fat body of Bactrocera dorsalis (Diptera: Tephritidae) identifying a new vitellogenin gene. Ann. Entomol. Soc. Am. 2018, 111, 43–54. [Google Scholar] [CrossRef]
- Wurm, Y.; Wang, J.; Riba-Grognuz, O.; Corona, M.; Nygaard, S.; Hunt, B.G.; Ingram, K.K.; Keller, L. The genome of the fire ant Solenopsis invicta. Proc. Natl. Acad. Sci. USA 2011, 108, 5679–5684. [Google Scholar] [CrossRef] [PubMed]
- Tokar, D.R.; Veleta, K.A.; Canzano, J.; Hahn, D.A.; Hatle, J.D. Vitellogenin RNAi Halts Ovarian Growth and Diverts Reproductive Proteins and Lipids in Young Grasshoppers. Integr. Comp. Biol. 2014, 54, 931–941. [Google Scholar] [CrossRef]
- Coelho, R.R.; Souza Júnior, J.D.A.; Firmino, A.A.P.; Macedo, L.L.P.; Fonseca, F.C.A.; Terra, W.R.; Engler, G.; Almeida Engler, J.; Silva, M.C.M.; Grossi-de-Sa, M.F. Vitellogenin knockdown strongly affects cotton boll weevil egg viability but not the number of eggs laid by females. Meta Gene 2016, 9, 173–180. [Google Scholar] [CrossRef]
- Moriyama, M.; Hosokawa, T.; Tanahashi, M.; Nikoh, N.; Fukatsu, T. Suppression of Bedbug’s Reproduction by RNA Interference of Vitellogenin. PLoS ONE 2016, 11, e0153984. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Han, H.L.; Li, W.J.; Wang, J.J.; Wei, D. Expression and Role of Vitellogenin Genes in Ovarian Development of Zeugodacus cucurbitae. Insects 2022, 13, 452. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.H.; Li, L.; Zeng, B.; Fu, Y.G. Effects of short-term high-temperature conditions on oviposition and differential gene expression of Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae). Int. J. Pest Manag. 2020, 66, 332–340. [Google Scholar] [CrossRef]
- Lian, Y.Y.; Peng, S.H.; Jia, J.J.; Li, L.; Wang, A.Q.; Yang, S.Y.; Zheng, R.J.; Yang, X.F.; Zhou, S.H. Function of Vitellogenin receptor gene in reproductive regulation of Zeugodacus cucurbitae (Coquillett) after short-term high-temperature treatment. Front. Physiol. 2022, 13, 995004. [Google Scholar] [CrossRef] [PubMed]
- Lian, Y.Y.; Peng, S.H.; Yang, X.F.; Jia, J.J.; Li, J.L.; Wang, A.Q.; Yang, S.Y.; Zheng, R.J.; Zhou, S.H. The vitellogenin receptor gene is involved in lifespan regulation of Zeugodacus cucurbitae (Coquillett) after short-term high-temperature treatment. Front. Physiol. 2022, 13, 1090348. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Yang, L.; He, Q.; Zhou, S. Regulatory mechanisms of vitellogenesis in insects. Front. Cell Dev. Biol. 2021, 8, 1863. [Google Scholar] [CrossRef]
- Ali, M.W.; Zhang, Z.Y.; Xia, S.; Zhang, H.Y. Biofunctional analysis of Vitellogenin and Vitellogenin receptor in citrus red mites, Panonychus citri by RNA interference. Sci. Rep. 2017, 7, 16123. [Google Scholar] [CrossRef]
- Rasool, K.G.; Mehmood, K.; Tufail, M.; Husain, M.; Alwaneen, W.S.; Aldawood, A.S. Silencing of vitellogenin gene contributes to the promise of controlling red palm weevil, Rhynchophorus ferrugineus (Olivier). Sci. Rep. 2021, 11, 21695. [Google Scholar] [CrossRef]
- Lu, K.; Shu, Y.; Zhou, J.; Zhang, X.; Zhang, X.; Chen, M.; Yao, Q.; Zhou, Q.; Zhang, W. Molecular characterization and RNA interference analysis of vitellogenin receptor from Nilaparvata lugens (Stål). J. Insect Physiol. 2015, 73, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Boldbaatar, D.; Umemiya-Shirafuji, R.; Liao, M.; Tanaka, T.; Xuan, X.; Fujisaki, K. Multiple vitellogenins from the Haemaphysalis longicornis tick are crucial for ovarian development. J. Insect Physiol. 2010, 56, 1587–1598. [Google Scholar] [CrossRef]
- Shen, Y.; Chen, Y.Z.; Lou, Y.H.; Zhang, C.X. Vitellogenin and Vitellogenin-Like Genes in the Brown Planthopper. Front. Physiol. 2019, 10, 1181. [Google Scholar] [CrossRef]
- Martin, D.; Piulachs, M.D.; Belles, X. Production and extraovarian processing of vitellogenin in ovariectomized Blattella germanica (L.) (Dictyoptera, Blattellidae). J. Insect Physiol. 1996, 42, 101–105. [Google Scholar] [CrossRef]
- Nagaba, Y.; Tufail, M.; Inui, H.; Takeda, M. Hormonal regulation and effects of four environmental pollutants on vitellogenin gene transcription in the giant water bug, Lethocerus deyrollei (Hemiptera: Belostomatidae). J. Insect Conserv. 2011, 15, 421–431. [Google Scholar] [CrossRef]
- Wyatt, G.R.; Davey, K.G. Cellular and molecular actions of juvenile hormone. II. roles of juvenile hormone in adult insects. Adv. Insect Physiol. 1996, 26, 1–155. [Google Scholar]
- Yamamoto, R.; Bai, H.; Dolezal, A.G.; Amdam, G.; Tatar, M. Juvenile hormone regulation of Drosophila aging. BMC Biol. 2013, 11, 85. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′ to 3′) | Use of Primers |
|---|---|---|
| Succinate dehydrogenase flavoprotein-F | TTGATTTCAAAATAGGCGCAGTG | Reference gene amplification in qPCR |
| Succinate dehydrogenase flavoprotein-R | CGATGGTACACGCATAAGGC | |
| Vitellogenin receptor-F | TGCTTTCCCGGTTATCGCTT | Target gene amplification in qPCR |
| Vitellogenin receptor-R | AACGTAATCGGTTGCTCCGT | |
| Vitellogenin-1-F | TGCCACGTGACTTAATCGGT | |
| Vitellogenin-1-R | ATGCTGTGGCTAGAGGCCAT | |
| Vitellogenin-2-F | GCTTGTAATGAGTCGGTGGCG | |
| Vitellogenin-2-R | AGTGGCGCAAAGAAATGCCT | |
| Vitellogenin-3-F | TTTGCTCCTCAGCACTCTCA | |
| Vitellogenin-3-R | GCGCCATATTTGATCGGCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Peng, S.; Wang, A.; Jia, J.; Wu, Q.; Yang, X.; Zhou, S. The ZcVg3 Gene Regulates the Reproduction and Lifespan of Female Zeugodacus cucurbitae (Coquillett) Mediated by Short-Term High Temperatures. Insects 2024, 15, 499. https://doi.org/10.3390/insects15070499
Yang S, Peng S, Wang A, Jia J, Wu Q, Yang X, Zhou S. The ZcVg3 Gene Regulates the Reproduction and Lifespan of Female Zeugodacus cucurbitae (Coquillett) Mediated by Short-Term High Temperatures. Insects. 2024; 15(7):499. https://doi.org/10.3390/insects15070499
Chicago/Turabian StyleYang, Shuyan, Sihua Peng, Aqiang Wang, Jingjing Jia, Qianxing Wu, Xiaofeng Yang, and Shihao Zhou. 2024. "The ZcVg3 Gene Regulates the Reproduction and Lifespan of Female Zeugodacus cucurbitae (Coquillett) Mediated by Short-Term High Temperatures" Insects 15, no. 7: 499. https://doi.org/10.3390/insects15070499