
A Novel Transgenic Sf9 Cell Line for Quick and Easy Virus Quantification

Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Construction of the Transfer Vector
2.3. Virus-Inducible Transient Expression
2.4. Generation of the Transgenic Cell Line
2.5. Virus Titration
2.6. Fluorescence Intensity Measurement
3. Results and Discussion
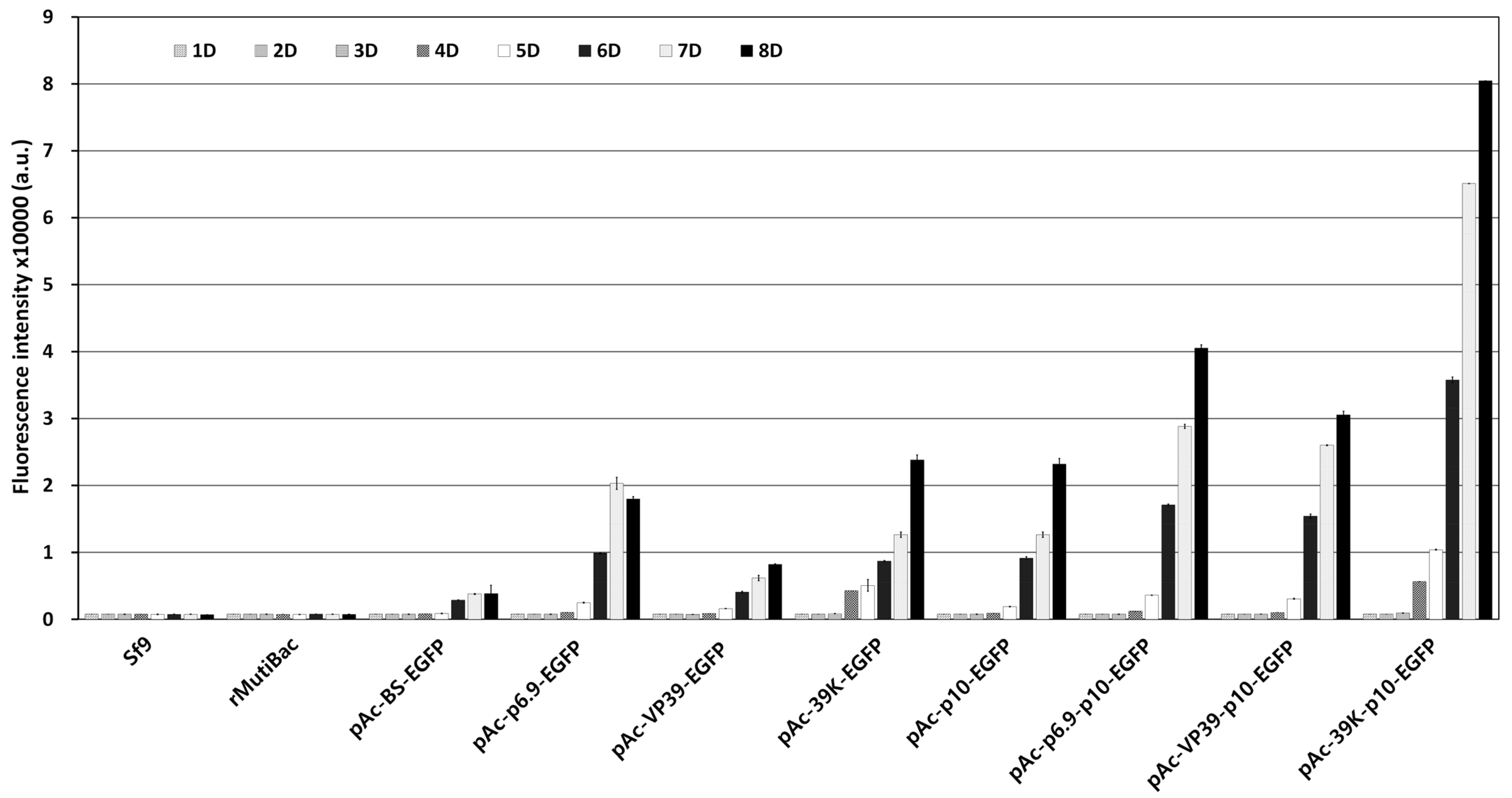
3.1. Optimal Expression Structure of the Marker Gene
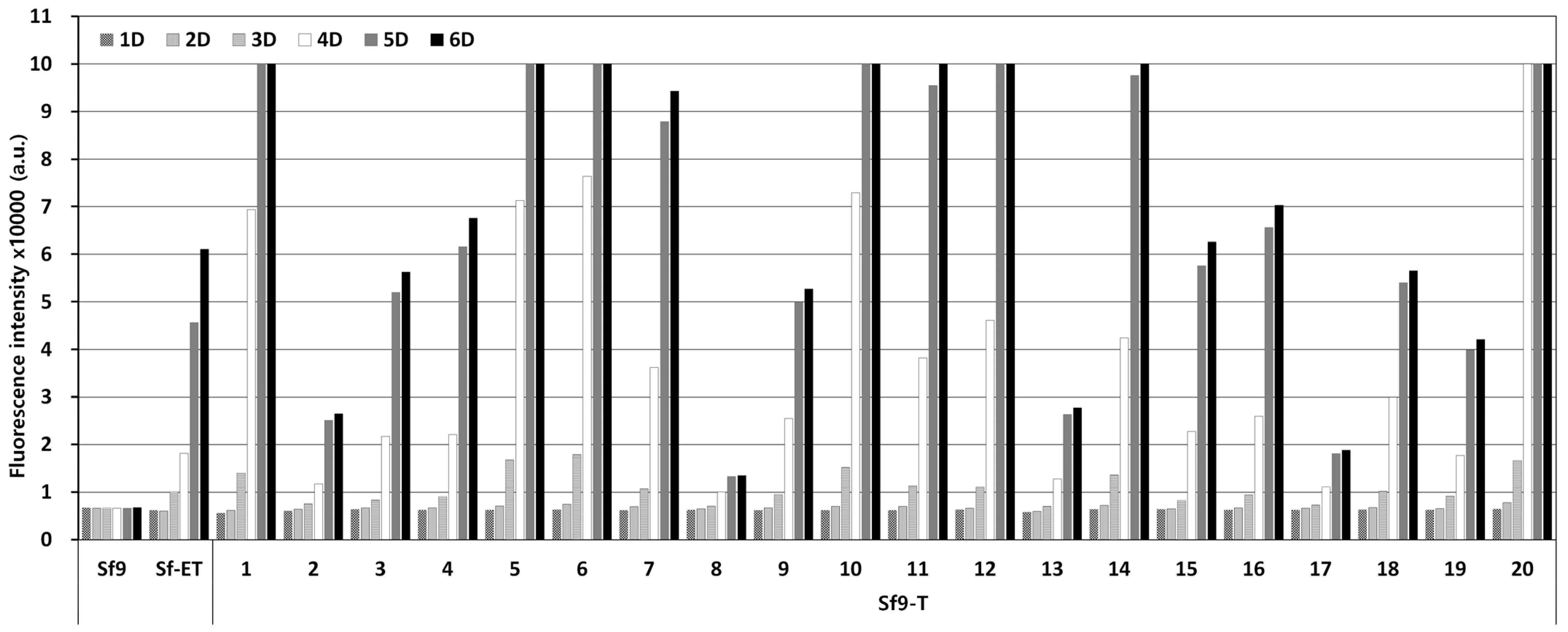
3.2. The Generation and Selection of the Transgenic Cell Lines
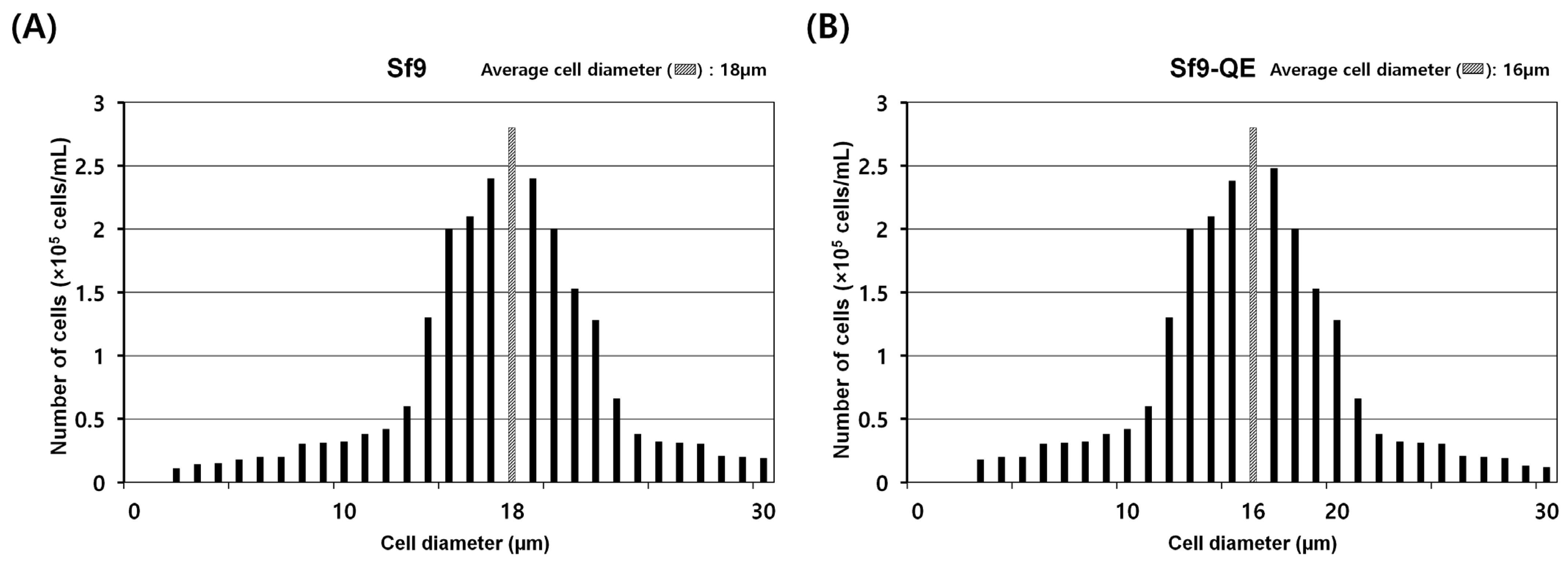
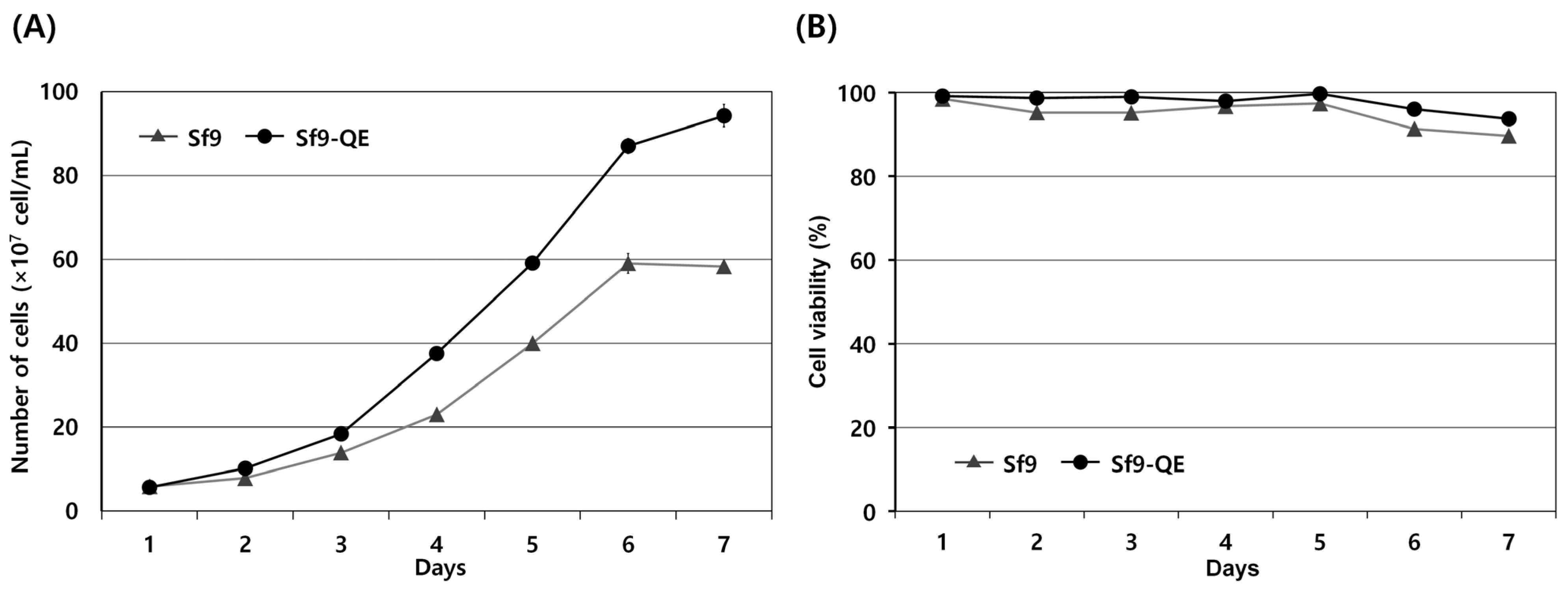
3.3. Characteristics of Sf9-QE
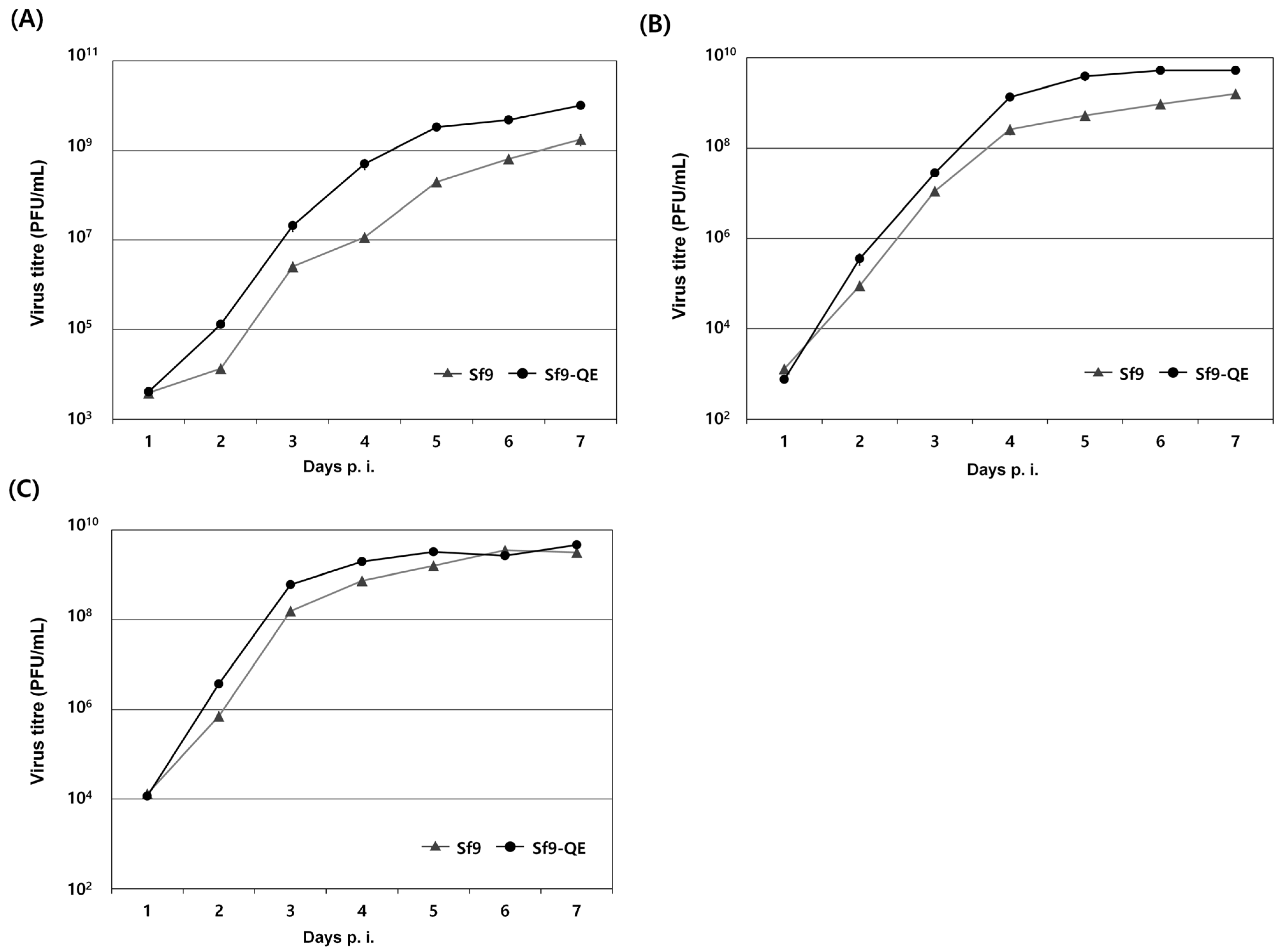
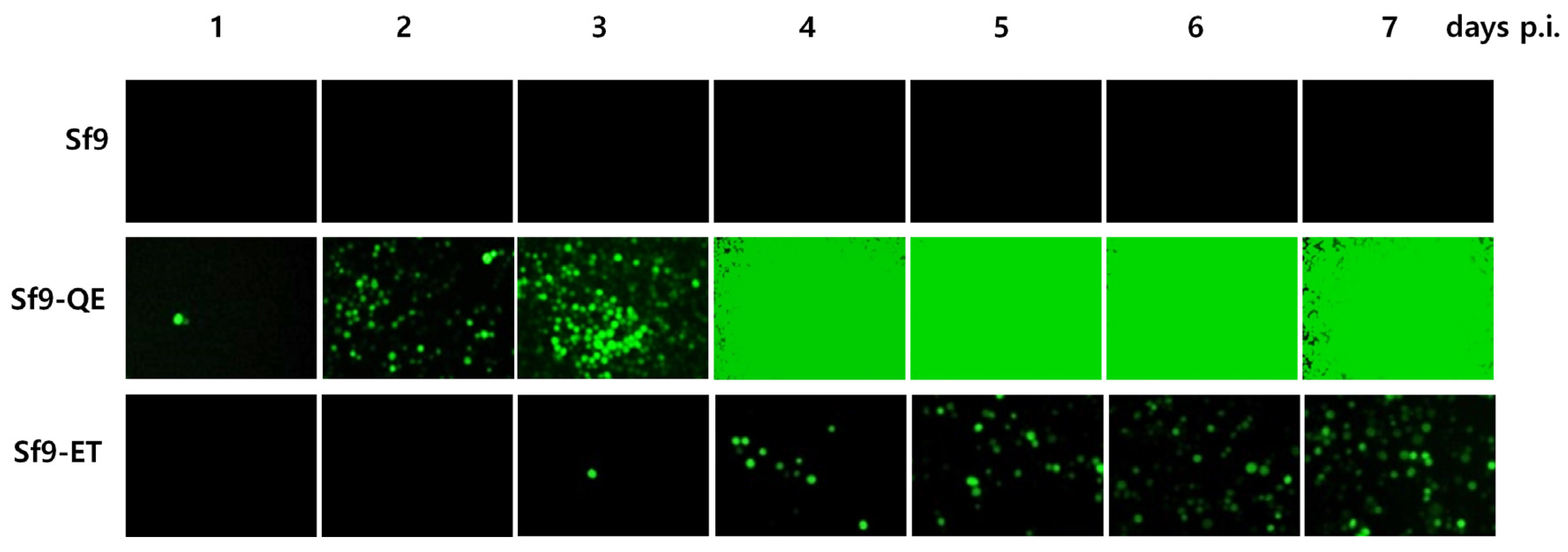
3.4. Suitability of the Sf9-QE Cells for Virus Quantification
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, N.; Kong, X.; Zhao, S.; Xiaofeng, W. Post-translational modification of baculovirus-encoded proteins. Virus Res. 2020, 279, 197865. [Google Scholar] [CrossRef] [PubMed]
- Hink, W.F.; Vail, P.V. A plaque assay for titration of alfalfa looper nuclear polyhedrosis virus in a cabbage looper (TN-368) cell line. J. Invertebr. Pathol. 1973, 22, 168–174. [Google Scholar] [CrossRef]
- Hitchman, R.B.; Locanto, E.; Possee, R.D.; King, L.A. Optimizing the baculovirus expression vector system. Methods 2011, 55, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.R.; Chao, Y.C. Rapid titer determination of baculovirus by quantitative real-time polymerase chain reaction. Biotechnol. Prog. 2004, 20, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Li, Y. Establishment, verification and application of rapid detection of baculovirus infectious titer by flow cytometry. J. Virol. Methods 2022, 303, 114495. [Google Scholar] [CrossRef]
- Kitts, P.A.; Green, G. An immunological assay for determination of baculovirus titers in 48 hours. Anal. Biochem. 1999, 268, 173–178. [Google Scholar] [CrossRef]
- Cresta, D.; Warren, D.C.; Quirouette, C.; Smith, A.P.; Lane, L.C.; Smith, A.M.; Beauchemin, C.A.A. Time to revisit the endpoint dilution assay and to replace the TCID50 as a measure of a virus sample’s infection concentration. PLoS Comput. Biol. 2021, 17, e1009480. [Google Scholar] [CrossRef]
- Hopkins, R.F.; Esposito, D. A rapid method for titrating baculovirus stocks using the Sf-9 easy titer cell line. BioTechniques 2009, 47, 785–788. [Google Scholar] [CrossRef]
- Takahashi, T. Routine management of microalgae using autofluorescence from chlorophyll. Molecules 2019, 24, 4441. [Google Scholar] [CrossRef]
- Mitsutake, H.; Kobayashi, J. piggyBac-mediated stable transformation of cultured Bombyx mori cells using in vitro synthesized transposase mRNA. J. Insect Biotechnol. Sericol. 2011, 80, 15–24. [Google Scholar] [CrossRef]
- Gwak, W.S.; Kim, H.S.; Bae, J.S.; Kim, T.H.; Bae, S.M.; Woo, S.D. Development of a novel enhanced baculovirus expression vector via promoter combination. J. Asia-Pac. Entomol. 2020, 23, 909–914. [Google Scholar] [CrossRef]
- Lin, C.H.; Jarvis, D.L. Utility of temporally distinct baculovirus promoters for constitutive and baculovirus-inducible transgene expression in transformed insect cells. J. Biotechnol. 2013, 165, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Luria, N.; Lu, L.; Chejanovsky, N. Conserved structural motifs at the C-terminus of baculovirus protein IE0 are important for its functions in transactivation and supporting hr5-mediated DNA replication. Viruses 2012, 4, 761–776. [Google Scholar] [CrossRef]
- Toth, A.M.; Kuo, C.W.; Khoo, K.H.; Jarvis, D.L. A new insect cell glycoengineering approach provides baculovirus-inducible glycogene expression and increases human-type glycosylation efficiency. J. Biotechnol. 2014, 182–183, 19–29. [Google Scholar] [CrossRef]
- Bruder, M.R.; Aucoin, M.G. Utility of alternative promoters for foreign gene expression using the baculovirus expression vector system. Viruses 2022, 14, 2670. [Google Scholar] [CrossRef]
- Cao, M.; Kuang, X.; Li, H.; Lei, X.; Xiao, W.; Dong, Z.; Zhang, J.; Hu, N.; Chen, T.T.; Lu, C.; et al. Screening and optimization of an efficient Bombyx mori nucleopolyhedrovirus inducible promoter. J. Biotechnol. 2016, 231, 72–80. [Google Scholar] [CrossRef]
- Ishiyama, S.; Ikeda, M. High-level expression and improved folding of proteins by using the vp39 late promoter enhanced with homologous DNA regions. Biotechnol. Lett. 2010, 32, 1637–1647. [Google Scholar] [CrossRef]
- Rankl, N.B.; Rice, J.W.; Gurganus, T.M.; Barbee, J.L.; Burns, D.J. The production of an active protein kinase C-δ in insect cells is greatly enhanced by the use of the basic protein promoter. Protein Expr. Purif. 1994, 5, 346–356. [Google Scholar] [CrossRef]
- Cha, H.J.; Dalal, N.G.; Pham, M.Q.; Kramer, S.F.; Vakharia, V.N.; Bentley, W.E. Monitoring foreign protein expression under baculovirus p10 and polh promoters in insect larvae. Biotechniques 2002, 32, 986–992. [Google Scholar] [CrossRef]
- DiFalco, M.R.; Bakopanos, E.; Patricelli, M.; Chan, G.; Congote, L.F. The influence of various insect cell lines, p10 and polyhedrin promoters in the production of secreted insulin-like growth factor-interleukin-3 chimeras in the baculovirus expression system. J. Biotechnol. 1997, 56, 49–56. [Google Scholar] [CrossRef]
- van Oers, M.M.; Malarme, D.; Jore, J.M.; Vlak, J.M. Expression of the Autographa californica nuclear polyhedrosis virus p10 gene: Effect of polyhedrin gene expression. Arch. Virol. 1992, 123, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bleckmann, M.; Schürig, M.; Chen, F.F.; Yen, Z.Z.; Lindemann, N.; Meyer, S.; Spehr, J.; van den Heuvel, J. Identification of essential genetic baculoviral elements for recombinant protein expression by transactivation in Sf21 insect cells. PLoS ONE 2016, 11, e0149424. [Google Scholar] [CrossRef] [PubMed]
- Cary, L.C.; Goebel, M.; Corsaro, B.G.; Wang, H.G.; Rosen, E.; Fraser, M.J. Transposon mutagenesis of baculoviruses: Analysis of Trichoplusia ni transposon IFP2 insertions within the FP-locus of nuclear polyhedrosis viruses. Virology 1989, 172, 156–169. [Google Scholar] [CrossRef]
- Yusa, K. piggyBac transposon. Microbiol. Spectr. 2015, 3, MDNA3-0028-2014. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Jiang, E.; Chen, S.; Gu, Y.; Shangguan, A.J.; Lv, T.; Luo, L.; Yu, Z. PiggyBac transposon vectors: The tools of the human gene encoding. Transl. Lung Cancer Res. 2016, 5, 120–125. [Google Scholar] [CrossRef]
- Ding, S.; Wu, X.; Li, G.; Han, M.; Zhuang, Y.; Xu, T. Efficient transposition of the piggyBac (PB) transposon in mammalian cells and mice. Cell 2005, 122, 473–483. [Google Scholar] [CrossRef]
- Li, X.; Ewis, H.E.; Hice, R.H.; Malani, N.; Parker, N.; Zhou, L.; Feschotte, C.; Bushman, F.D.; Atkinson, P.W.; Craig, N.L. A resurrected mammalian hAT transposable element and a closely related insect element are highly active in human cell culture. Proc. Natl. Acad. Sci. USA 2013, 110, 2347–2352. [Google Scholar] [CrossRef]
- Li, X.; Burnight, E.R.; Cooney, A.L.; Malani, N.; Brady, T.; Sander, J.D.; Staber, J.; Wheelan, S.J.; Joung, J.K.; McCray, P.B., Jr.; et al. piggyBac transposase tools for genome engineering. Proc. Natl. Acad. Sci. USA 2013, 110, E2279–E2287. [Google Scholar] [CrossRef]
- Wang, W.; Lin, C.; Lu, D.; Ning, Z.; Cox, T.; Melvin, D.; Wang, X.; Bradley, A.; Liu, P. Chromosomal transposition of PiggyBac in mouse embryonic stem cells. Proc. Natl. Acad. Sci. USA 2008, 105, 9290–9295. [Google Scholar] [CrossRef]
- Kost, T.A.; Condreay, J.P.; Jarvis, D.L. Baculovirus as versatile vectors for protein expression in insect and mammalian cells. Nat. Biotechnol. 2005, 23, 567–575. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | rAc-LacZ | rAc-PPV-VP2 | rMultiBac | ||||
|---|---|---|---|---|---|---|---|
| Cell Line | Titre (PFU/mL) | C. Days 1 | Titre (PFU/mL) | C. Days | Titre (PFU/mL) | C. Days | |
| Sf9 | 4.3 × 107 ± 2.3% | 12.0 ± 0.0 | 6.9 × 108 ± 3.9% | 12.0 ± 0.0 | 3.7 × 108 ± 5.8% | 11.3 ± 0.3 | |
| Sf9-QE | 3.2 × 107 ± 7.3% | 5.7 ± 0.3 | 7.3 × 108 ± 2.4% | 6.0 ± 0.0 | 8.0 × 108 ± 1.3% | 5.3 ± 0.3 | |
| Sf9-ET | 1.2 × 107 ± 8.1% | 10.7 ± 0.9 | 7.1 × 108 ± 4.8% | 10.0 ± 0.0 | 2.7 × 108 ± 9.8% | 10.0 ± 0.6 | |
| Virus | rAc-LacZ | rAc-PPV-VP2 | rMultiBac | ||||
|---|---|---|---|---|---|---|---|
| Cell Line | Titre (PFU/mL) | C. Days 1 | Titre (PFU/mL) | C. Days | Titre (PFU/mL) | C. Days | |
| Sf9-QE | Microscope | 3.2 × 107 ± 6.3% | 5.7 ± 0.3 | 7.3 × 108 ± 3.4% | 6.0 ± 0.0 | 8.0 × 108 ± 2.3% | 5.3 ± 0.3 |
| Photometry | 3.1 × 107 ± 7.3% | 6.3 ± 0.3 | 7.2 × 108 ± 2.4% | 6.7 ± 0.3 | 7.9 × 108 ± 1.3% | 6.0 ± 0.3 | |
| Sf9-ET | Microscope | 1.2 × 107 ± 8.1% | 10.7 ± 0.9 | 7.1 × 108 ± 4.9% | 10.0 ± 0.0 | 2.7 × 108 ± 9.8% | 10.0 ± 0.6 |
| Photometry | 1.2 × 107 ± 13.3% | 12 ± 0.0 | 6.4 × 108 ± 4.8% | 12 ± 0.0 | 2.6 × 108 ± 14.9% | 12 ± 0.0 | |
| Virus | rAc-LacZ | rAc-PPV-VP2 | rMultiBac | ||||
|---|---|---|---|---|---|---|---|
| Passages | Titre (PFU/mL) | C. Days 1 | Titre (PFU/mL) | C. Days | Titre (PFU/mL) | C. Days | |
| P.20 | 7.9 × 108 ± 3.7% | 5.7 ± 0.3 | 6.9 × 108 ± 6.1% | 6.7 ± 0.3 | 8.3 × 108 ± 5.3% | 6.0 ± 0.0 | |
| P.100 | 7.3 × 108 ± 3.1% | 5.3 ± 0.3 | 7.1 × 108 ± 6.9% | 6.3 ± 0.3 | 8.4 × 108 ± 5.1% | 5.7 ± 0.3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.-S.; Bae, J.-S.; Moon, H.-J.; Kim, D.-Y.; Woo, S.-D. A Novel Transgenic Sf9 Cell Line for Quick and Easy Virus Quantification. Insects 2024, 15, 686. https://doi.org/10.3390/insects15090686
Kim K-S, Bae J-S, Moon H-J, Kim D-Y, Woo S-D. A Novel Transgenic Sf9 Cell Line for Quick and Easy Virus Quantification. Insects. 2024; 15(9):686. https://doi.org/10.3390/insects15090686
Chicago/Turabian StyleKim, Kyu-Seek, Jun-Su Bae, Hyuk-Jin Moon, Do-Young Kim, and Soo-Dong Woo. 2024. "A Novel Transgenic Sf9 Cell Line for Quick and Easy Virus Quantification" Insects 15, no. 9: 686. https://doi.org/10.3390/insects15090686
APA StyleKim, K.-S., Bae, J.-S., Moon, H.-J., Kim, D.-Y., & Woo, S.-D. (2024). A Novel Transgenic Sf9 Cell Line for Quick and Easy Virus Quantification. Insects, 15(9), 686. https://doi.org/10.3390/insects15090686