
Detection of Lotmaria passim, Crithidia mellificae and Replicative Forms of Deformed Wing Virus and Kashmir Bee Virus in the Small Hive Beetle (Aethina tumida)




Abstract
:1. Introduction
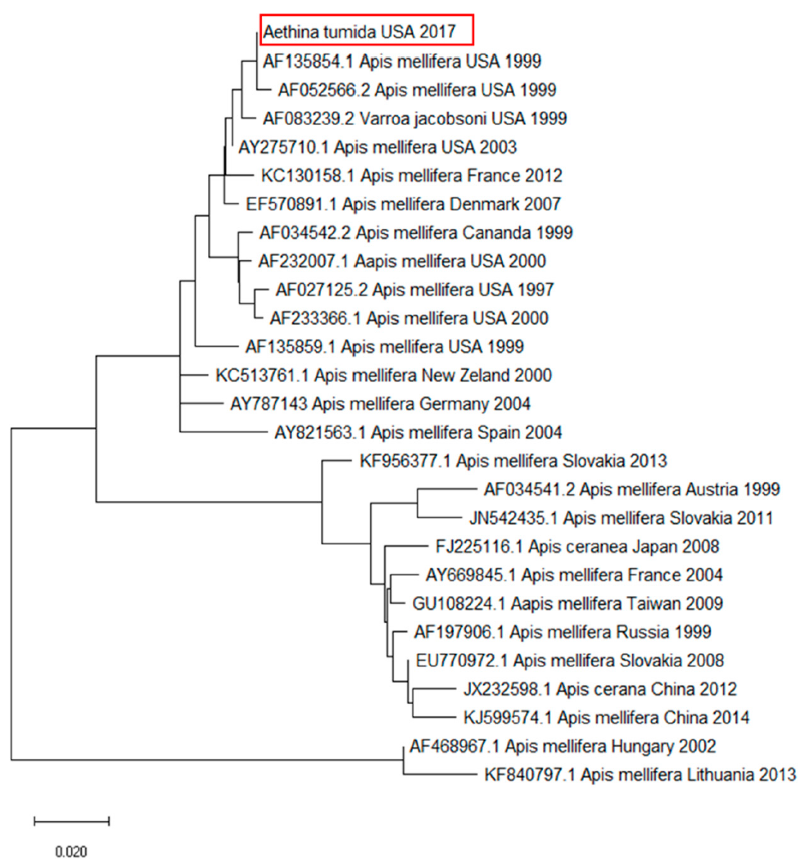
2. Results
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Extraction of Total Nucleic Acids
4.3. PCR Assays to Detect Bacteria and Protozoa DNA
4.4. PCR Assays to Detect Virus RNA
4.5. Strand-Specific RT-PCR
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Murray, A. List of Coleoptera received from Old Calabar. Ann. Mag. Nat. Hist. Lond. 1867, 19, 167–179. [Google Scholar] [CrossRef]
- Lundie, A.E. The Small Hive Beetle, Aethina túmida. Sci. Bull. 1940, 220, 40. [Google Scholar]
- Schäfer, M.O.; Cardaio, I.; Cilia, G.; Cornelissen, B.; Crailsheim, K.; Formato, G.; Lawrence, A.K.; Le Conte, Y.; Mutinelli, F.; Nanetti, A.; et al. How to slow the global spread of small hive beetles, Aethina tumida. Biol. Invasions 2019, 21, 1451–1459. [Google Scholar] [CrossRef]
- Ellis, J.D.; Hepburn, H.R. An ecological digest of the small hive beetle (Aethina tumida), a symbiont in honey bee colonies (Apis mellifera). Insectes Soc. 2006, 53, 8–19. [Google Scholar] [CrossRef]
- Al Toufailia, H.; Alves, D.A.; De Bená, D.C.; Bento, J.M.S.; Iwanicki, N.S.A.; Cline, A.R.; Ellis, J.D.; Ratnieks, F.L.W. First record of small hive beetle, Aethina tumida Murray, in South America. J. Apic. Res. 2017, 56, 76–80. [Google Scholar] [CrossRef]
- Lee, S.; Hong, K.-J.; Cho, Y.S.; Choi, Y.S.; Yoo, M.-S.; Lee, S. Review of the subgenus Aethina Erichson s. str. (Coleoptera: Nitidulidae: Nitidulinae) in Korea, reporting recent invasion of small hive beetle, Aethina tumida. J. Asia. Pac. Entomol. 2017, 20, 553–558. [Google Scholar] [CrossRef]
- Neumann, P.; Pettis, J.S.; Schäfer, M.O. Quo vadis Aethina tumida? Biology and control of small hive beetles. Apidologie 2016, 47, 427–466. [Google Scholar] [CrossRef] [Green Version]
- Boncristiani, H.; Ellis, J.D.; Bustamante, T.; Graham, J.; Jack, C.; Kimmel, C.B.; Mortensen, A.; Schmehl, D.R. World Honey Bee Health: The Global Distribution of Western Honey Bee (Apis mellifera L.) Pests and Pathogens. Bee World 2021, 98, 2–6. [Google Scholar] [CrossRef]
- Granato, A.; Zecchin, B.; Baratto, C.; Duquesne, V.; Negrisolo, E.; Chauzat, M.-P.; Ribière-Chabert, M.; Cattoli, G.; Mutinelli, F. Introduction of Aethina tumida (Coleoptera: Nitidulidae) in the regions of Calabria and Sicily (southern Italy). Apidologie 2017, 48, 194–203. [Google Scholar] [CrossRef] [Green Version]
- da Silva, M.J.V. The First Report Of Aethina tumida In The European Union, Portugal, 2004. Bee World 2014, 91, 90–91. [Google Scholar] [CrossRef]
- Palmeri, V.; Scirtò, G.; Malacrinò, A.; Laudani, F.; Campolo, O. A scientific note on a new pest for European honeybees: First report of small hive beetle Aethina tumida, (Coleoptera: Nitidulidae) in Italy. Apidologie 2015, 46, 527–529. [Google Scholar] [CrossRef] [Green Version]
- Murilhas, A.M. Aethina tumida arrives in Portugal. Will it be eradicated? EurBee Newsl. 2004, 2, 7–9. [Google Scholar]
- Arbogast, R.T.; Torto, B.; Willms, S.; Teal, P.E.A. Trophic habits of Aethina tumida (Coleoptera: Nitidulidae): Their adaptive significance and relevance to dispersal. Environ. Entomol. 2009, 38, 561–568. [Google Scholar] [CrossRef] [Green Version]
- McMenamin, A.J.; Genersch, E. Honey bee colony losses and associated viruses. Curr. Opin. Insect Sci. 2015, 8, 121–129. [Google Scholar] [CrossRef]
- Brutscher, L.M.; McMenamin, A.J.; Flenniken, M.L. The Buzz about Honey Bee Viruses. PLoS Pathog. 2016, 12, e1005757. [Google Scholar] [CrossRef]
- Grupe, A.C.; Quandt, C.A. A growing pandemic: A review of Nosema parasites in globally distributed domesticated and native bees. PLoS Pathog. 2020, 16, e1008580. [Google Scholar] [CrossRef]
- Genersch, E.; Evans, J.D.; Fries, I. Honey bee disease overview. J. Invertebr. Pathol. 2010, 103, S2–S4. [Google Scholar] [CrossRef]
- Eyer, M.; Chen, Y.P.; Schäfer, M.O.; Pettis, J.; Neumann, P. Small hive beetle, Aethina tumida, as a potential biological vector of honeybee viruses. Apidologie 2009, 40, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, M.O.; Ritter, W.; Pettis, J.; Neumann, P. Small hive beetles, Aethina tumida, are vectors of Paenibacillus larvae. Apidologie 2010, 41, 14–20. [Google Scholar] [CrossRef]
- Cilia, G.; Cardaio, I.; dos Santos, P.E.J.; Ellis, J.D.; Nanetti, A. The first detection of Nosema ceranae (Microsporidia) in the small hive beetle, Aethina tumida Murray (Coleoptera: Nitidulidae). Apidologie 2018, 49, 619–624. [Google Scholar] [CrossRef] [Green Version]
- Eyer, M.; Chen, Y.P.; Schäfer, M.O.; Pettis, J.S.; Neumann, P. Honey bee sacbrood virus infects adult small hive beetles, Aethina tumida (Coleoptera: Nitidulidae). J. Apic. Res. 2009, 48, 296–297. [Google Scholar] [CrossRef]
- de Landa, G.F.; Porrini, M.P.; Revainera, P.; Porrini, D.P.; Farina, J.; Correa-Benítez, A.; Maggi, M.D.; Eguaras, M.J.; Quintana, S. Pathogens Detection in the Small Hive Beetle (Aethina tumida (Coleoptera: Nitidulidae)). Neotrop. Entomol. 2020, 1–5. [Google Scholar] [CrossRef]
- Huwiler, M.; Papach, A.; Cristina, E.; Yañez, O.; Williams, G.R.; Neumann, P. Deformed wings of small hive beetle independent of virus infections and mites. J. Invertebr. Pathol. 2020, 172, 107365. [Google Scholar] [CrossRef]
- Arismendi, N.; Caro, S.; Castro, M.P.; Vargas, M.; Riveros, G.; Venegas, T. Impact of Mixed Infections of Gut Parasites Lotmaria passim and Nosema ceranae on the Lifespan and Immune-related Biomarkers in Apis mellifera. Insects 2020, 11, 420. [Google Scholar] [CrossRef]
- Schwarz, R.S.; Bauchan, G.R.; Murphy, C.A.; Ravoet, J.; de Graaf, D.C.; Evans, J.D. Characterization of Two Species of Trypanosomatidae from the Honey Bee Apis mellifera: Crithidia mellificae Langridge and McGhee, and Lotmaria passim n. gen., n. sp. J. Eukaryot. Microbiol. 2015, 62, 567–583. [Google Scholar] [CrossRef]
- Ruiz- Gonzalez, M.X.; Brown, M.J.F. Honey bee and bumblebee trypanosomatids: Specificity and potential for transmission. Ecol. Entomol. 2006, 31, 616–622. [Google Scholar] [CrossRef]
- Runckel, C.; DeRisi, J.; Flenniken, M.L. A Draft Genome of the Honey Bee Trypanosomatid Parasite Crithidia mellificae. PLoS ONE 2014, 9, e95057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Runckel, C.; Flenniken, M.L.; Engel, J.C.; Ruby, J.G.; Ganem, D.; Andino, R.; DeRisi, J.L. Temporal analysis of the honey bee microbiome reveals four novel viruses and seasonal prevalence of known viruses, Nosema, and Crithidia. PLoS ONE 2011, 6, e020656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravoet, J.; Maharramov, J.; Meeus, I.; De Smet, L.; Wenseleers, T.; Smagghe, G.; de Graaf, D.C. Comprehensive Bee Pathogen Screening in Belgium Reveals Crithidia mellificae as a New Contributory Factor to Winter Mortality. PLoS ONE 2013, 8, e72443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, R.S.; Evans, J.D. Single and mixed-species trypanosome and microsporidia infections elicit distinct, ephemeral cellular and humoral immune responses in honey bees. Dev. Comp. Immunol. 2013, 40, 300–310. [Google Scholar] [CrossRef]
- Strobl, V.; Yañez, O.; Straub, L.; Albrecht, M.; Neumann, P. Trypanosomatid parasites infecting managed honeybees and wild solitary bees. Int. J. Parasitol. 2019, 49, 605–613. [Google Scholar] [CrossRef]
- Buendía, M.; Martín-Hernández, R.; Ornosa, C.; Barrios, L.; Bartolomé, C.; Higes, M. Epidemiological study of honeybee pathogens in Europe: The results of Castilla-La Mancha (Spain). Spanish J. Agric. Res. 2018, 16, e0502. [Google Scholar] [CrossRef] [Green Version]
- Arismendi, N.; Bruna, A.; Zapata, N.; Vargas, M. PCR-specific detection of recently described Lotmaria passim (Trypanosomatidae) in Chilean apiaries. J. Invertebr. Pathol. 2016, 134, 1–5. [Google Scholar] [CrossRef]
- Vargas, M.; Arismendi, N.; Riveros, G.; Zapata, N.; Bruna, A.; Vidal, M.; Rodríguez, M.; Gerding, M. Viral and intestinal diseases detected in Apis mellifera in central and southern Chile. Chil. J. Agric. Res. 2017, 77, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Castelli, L.; Branchiccela, B.; Invernizzi, C.; Tomasco, I.; Basualdo, M.; Rodriguez, M.; Zunino, P.; Antúnez, K. Detection of Lotmaria passim in Africanized and European honey bees from Uruguay, Argentina and Chile. J. Invertebr. Pathol. 2019, 160, 95–97. [Google Scholar] [CrossRef]
- de Miranda, J.R.; Genersch, E. Deformed wing virus. J. Invertebr. Pathol. 2010, 103, S48–S61. [Google Scholar] [CrossRef] [PubMed]
- Genersch, E.; Aubert, M. Emerging and re-emerging viruses of the honey bee (Apis mellifera L.). Vet. Res. 2010, 41, 54. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.J.; Highfield, A.C.; Brettell, L.; Villalobos, E.M.; Budge, G.E.; Powell, M.; Nikaido, S.; Schroeder, D.C. Global honey bee viral landscape altered by a parasitic mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef]
- Mordecai, G.J.; Wilfert, L.; Martin, S.J.; Jones, I.M.; Schroeder, D.C. Diversity in a honey bee pathogen: First report of a third master variant of the Deformed Wing Virus quasispecies. ISME J. 2016, 10, 1264–1273. [Google Scholar] [CrossRef] [Green Version]
- McMahon, D.P.; Natsopoulou, M.E.; Doublet, V.; Fürst, M.; Weging, S.; Brown, M.J.F.; Gogol-Döring, A.; Paxton, R.J. Elevated virulence of an emerging viral genotype as a driver of honeybee loss. Proc. Biol. Sci. 2016, 283, 20160811. [Google Scholar] [CrossRef]
- Yue, C.; Schroder, M.; Gisder, S.; Genersch, E. Vertical-transmission routes for deformed wing virus of honeybees (Apis mellifera). J. Gen. Virol. 2007, 88, 2329–2336. [Google Scholar] [CrossRef] [PubMed]
- Ball, B.V.; Allen, M.F. The prevalence of pathogens in honey bee (Apis mellifera) colonies infested with the parasitic mite Varroa jacobsoni. Ann. Appl. Biol. 1988, 113, 237–244. [Google Scholar] [CrossRef]
- Nordström, S. Distribution of deformed wing virus within honey bee (Apis mellifera) brood cells infested with the ectoparasitic mite Varroa destructor. Exp. Appl. Acarol. 2003, 29, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Cui, L.; Ostiguy, N.; Cox-Foster, D. Intricate transmission routes and interactions between picorna-like viruses (Kashmir bee virus and sacbrood virus) with the honeybee host and the parasitic varroa mite. J. Gen. Virol. 2005, 86, 2281–2289. [Google Scholar] [CrossRef]
- Lanzi, G.; de Miranda, J.R.; Boniotti, M.B.; Cameron, C.E.; Lavazza, A.; Capucci, L.; Camazine, S.M.; Rossi, C. Molecular and biological characterization of deformed wing virus of honeybees (Apis mellifera L.). J. Virol. 2006, 80, 4998–5009. [Google Scholar] [CrossRef] [Green Version]
- Gisder, S.; Aumeier, P.; Genersch, E. Deformed wing virus: Replication and viral load in mites (Varroa destructor). J. Gen. Virol. 2009, 90, 463–467. [Google Scholar] [CrossRef]
- Mazzei, M.; Carrozza, M.L.; Luisi, E.; Forzan, M.; Giusti, M.; Sagona, S.; Tolari, F.; Felicioli, A. Infectivity of DWV Associated to Flower Pollen: Experimental Evidence of a Horizontal Transmission Route. PLoS ONE 2014, 9, e113448. [Google Scholar] [CrossRef] [Green Version]
- Mockel, N.; Gisder, S.; Genersch, E. Horizontal transmission of deformed wing virus: Pathological consequences in adult bees (Apis mellifera) depend on the transmission route. J. Gen. Virol. 2011, 92, 370–377. [Google Scholar] [CrossRef]
- Chen, Y.; Evans, J.; Feldlaufer, M. Horizontal and vertical transmission of viruses in the honey bee, Apis mellifera. J. Invertebr. Pathol. 2006, 92, 152–159. [Google Scholar] [CrossRef]
- de Miranda, J.R.; Cordoni, G.; Budge, G. The Acute bee paralysis virus–Kashmir bee virus–Israeli acute paralysis virus complex. J. Invertebr. Pathol. 2010, 103, S30–S47. [Google Scholar] [CrossRef]
- Valles, S.M.; Strong, C.A.; Oi, D.H.; Porter, S.D.; Pereira, R.M.; Vander Meer, R.K.; Hashimoto, Y.; Hooper-Bùi, L.M.; Sánchez-Arroyo, H.; Davis, T.; et al. Phenology, distribution, and host specificity of Solenopsis invicta virus-1. J. Invertebr. Pathol. 2007, 96, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Berenyi, O.; Bakonyi, T.; Derakhshifar, I.; Koglberger, H.; Nowotny, N. Occurrence of Six Honeybee Viruses in Diseased Austrian Apiaries. Appl. Environ. Microbiol. 2006, 72, 2414–2420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tentcheva, D.; Gauthier, L.; Zappulla, N.; Dainat, B.; Cousserans, F.; Colin, M.E.; Bergoin, M. Prevalence and seasonal variations of six bee viruses in Apis mellifera L. and Varroa destructor mite populations in France. Appl. Environ. Microbiol. 2004, 70, 7185–7191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzei, M.; Cilia, G.; Forzan, M.; Lavazza, A.; Mutinelli, F.; Felicioli, A. Detection of replicative Kashmir Bee Virus and Black Queen Cell Virus in Asian hornet Vespa velutina (Lepelieter 1836) in Italy. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Cersini, A.; Bellucci, V.; Lucci, S.; Mutinelli, F.; Granato, A.; Porrini, C.; Felicioli, A.; Formato, G. First isolation of Kashmir bee virus (KBV) in Italy. J. Apic. Res. 2013, 52, 54–55. [Google Scholar] [CrossRef]
- Porrini, C.; Mutinelli, F.; Bortolotti, L.; Granato, A.; Laurenson, L.; Roberts, K.; Gallina, A.; Silvester, N.; Medrzycki, P.; Renzi, T.; et al. The Status of Honey Bee Health in Italy: Results from the Nationwide Bee Monitoring Network. PLoS ONE 2016, 11, e0155411. [Google Scholar] [CrossRef]
- Siede, R.; Büchler, R. First detection of Kashmir bee virus in Hesse, Germany. Berl. Munch. Tierarztl. Wochenschr. 2004, 117, 12–15. [Google Scholar]
- Ward, L.; Waite, R.; Boonham, N.; Fisher, T.; Pescod, K.; Thompson, H.; Chantawannakul, P.; Brown, M. First detection of Kashmir bee virus in the UK using real-time PCR. Apidologie 2007, 38, 181–190. [Google Scholar] [CrossRef] [Green Version]
- de Miranda, J.R.; Dainat, B.; Locke, B.; Cordoni, G.; Berthoud, H.; Gauthier, L.; Neumann, P.; Budge, G.E.; Ball, B.V.; Stoltz, D.B. Genetic characterization of slow bee paralysis virus of the honeybee (Apis mellifera L.). J. Gen. Virol. 2010, 91, 2524–2530. [Google Scholar] [CrossRef]
- Evans, J.D. Genetic Evidence for Coinfection of Honey Bees by Acute Bee Paralysis and Kashmir Bee Viruses. J. Invertebr. Pathol. 2001, 78, 189–193. [Google Scholar] [CrossRef] [Green Version]
- Pettis, J.; Van Engelsdorp, D.; Cox-Foster, D. Colony collapse disorder working group pathogen sub-group progress report. Am. Bee J. 2007, 103, 595–597. [Google Scholar]
- Todd, J.H.; De Miranda, J.R.; Ball, B.V. Incidence and molecular characterization of viruses found in dying New Zealand honey bee (Apis mellifera) colonies infested with Varroa destructor. Apidologie 2007, 38, 354–367. [Google Scholar] [CrossRef]
- Chen, Y.P.; Siede, R. Honey Bee Viruses. Adv. Virus Res. 2007, 70, 33–80. [Google Scholar]
- Shen, M.; Yang, X.; Cox-Foster, D.; Cui, L. The role of varroa mites in infections of Kashmir bee virus (KBV) and deformed wing virus (DWV) in honey bees. Virology 2005, 342, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Hung, A.C.F. PCR detection of Kashmir bee virus in honey bee excreta. J. Apic. Res. 2000, 39, 103–106. [Google Scholar] [CrossRef]
- Hung, A.C.F.; Shimanuki, H. A scientific note on the detection of Kashmir bee virus in individual honeybees and Varroa jacobsoni mites. Apidologie 1999, 30, 353–354. [Google Scholar] [CrossRef] [Green Version]
- Forzan, M.; Sagona, S.; Mazzei, M.; Felicioli, A. Detection of deformed wing virus in Vespa crabro. Bull. Insectology 2017, 70, 261–265. [Google Scholar]
- Mazzei, M.; Forzan, M.; Cilia, G.; Sagona, S.; Bortolotti, L.; Felicioli, A. First detection of replicative deformed wing virus (DWV) in Vespa velutina nigrithorax. Bull. Insectology 2018, 71, 211–216. [Google Scholar]
- Sébastien, A.; Lester, P.J.; Hall, R.J.; Wang, J.; Moore, N.E.; Gruber, M.A.M. Invasive ants carry novel viruses in their new range and form reservoirs for a honeybee pathogen. Biol. Lett. 2015, 11, 20150610. [Google Scholar] [CrossRef]
- Anderson, D.L. Kashmir bee virus: A relatively harmless virus of honey bee colonies. Am. Bee J. 1991, 131, 767–771. [Google Scholar]
- Quinn, O.; Gruber, M.A.M.; Brown, R.L.; Baty, J.W.; Bulgarella, M.; Lester, P.J. A metatranscriptomic analysis of diseased social wasps (Vespula vulgaris) for pathogens, with an experimental infection of larvae and nests. PLoS ONE 2018, 13, e0209589. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Levitt, A.L.; Rajotte, E.G.; Holmes, E.C.; Ostiguy, N.; vanEngelsdorp, D.; Lipkin, W.I.; dePamphilis, C.W.; Toth, A.L.; Cox-Foster, D.L. RNA Viruses in Hymenopteran Pollinators: Evidence of Inter-Taxa Virus Transmission via Pollen and Potential Impact on Non-Apis Hymenopteran Species. PLoS ONE 2010, 5, e14357. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D.; Schwarz, R.S. Bees brought to their knees: Microbes affecting honey bee health. Trends Microbiol. 2011, 19, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Hung, A.C.; Adams, J.R.; Shimanuki, H. Bee parasitic mite syndrome. (II). The role of Varroa mite and viruses. Am. Bee J. 1995, 135, 702–704. [Google Scholar]
- Ellis, J.D.; Pirk, C.W.W.; Hepburn, H.R.; Kastberger, G.; Elzen, P.J. Small hive beetles survive in honeybee prisons by behavioural mimicry. Naturwissenschaften 2002, 89, 326–328. [Google Scholar] [CrossRef]
- Ellis, J.D. Reviewing the confinement of small hive beetles (Aethina tumida) by western honey bees (Apis mellifera). Bee World 2005, 86, 56–62. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Wakefield, M.E.; Powell, M.E.; Marris, G.; Anderson, H.; Budge, G.E.; Mathers, J.J.; Blackburn, L.F.; Brown, M.A. The small hive beetle Aethina tumida: A review of its biology and control measures. Curr. Zool. 2013, 59, 644–653. [Google Scholar] [CrossRef]
- Chen, Y.P.; Pettis, J.S.; Collins, A.; Feldlaufer, M.F. Prevalence and Transmission of Honeybee Viruses. Appl. Environ. Microbiol. 2006, 72, 606–611. [Google Scholar] [CrossRef] [Green Version]
- Gusachenko, O.N.; Woodford, L.; Balbirnie-Cumming, K.; Ryabov, E.V.; Evans, D.J. Evidence for and against deformed wing virus spillover from honey bees to bumble bees: A reverse genetic analysis. Sci. Rep. 2020, 10, 16847. [Google Scholar] [CrossRef]
- Felicioli, A.; Forzan, M.; Sagona, S.; D’Agostino, P.; Baido, D.; Fronte, B.; Mazzei, M. Effect of oral administration of 1,3-1,6 β-glucans in DWV naturally infected newly emerged bees (Apis mellifera L.). Vet. Sci. 2020, 7, 52. [Google Scholar] [CrossRef]
- Cilia, G.; Garrido, C.; Bonetto, M.; Tesoriero, D.; Nanetti, A. Effect of Api-Bioxal® and ApiHerb® Treatments against Nosema ceranae Infection in Apis mellifera Investigated by Two qPCR Methods. Vet. Sci. 2020, 7, 125. [Google Scholar] [CrossRef]
- Dobbelaere, W.; de Graaf, D.C.; Peeters, J.E. Development of a fast and reliable diagnostic method for American foulbrood disease (Paenibacillus larvae subsp. larvae) using a 16S rRNA gene based PCR. Apidologie 2001, 32, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Roetschi, A.; Berthoud, H.; Kuhn, R.; Imdorf, A. Infection rate based on quantitative real-time PCR of Melissococcus plutonius, the causal agent of European foulbrood, in honeybee colonies before and after apiary sanitation. Apidologie 2008, 39, 362–371. [Google Scholar] [CrossRef]
- Xu, G.; Palmer-Young, E.; Skyrm, K.; Daly, T.; Sylvia, M.; Averill, A.; Rich, S. Triplex real-time PCR for detection of Crithidia mellificae and Lotmaria passim in honey bees. Parasitol. Res. 2018, 117, 623–628. [Google Scholar] [CrossRef]
- Chantawannakul, P.; Ward, L.; Boonham, N.; Brown, M. A scientific note on the detection of honeybee viruses using real-time PCR (TaqMan) in Varroa mites collected from a Thai honeybee (Apis mellifera) apiary. J. Invertebr. Pathol. 2006, 91, 69–73. [Google Scholar] [CrossRef]
- Kajobe, R.; Marris, G.; Budge, G.; Laurenson, L.; Cordoni, G.; Jones, B.; Wilkins, S.; Cuthbertson, A.G.S.; Brown, M.A. First molecular detection of a viral pathogen in Ugandan honey bees. J. Invertebr. Pathol. 2010, 104, 153–156. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Target | Pool (n = 30) | SHB 1 | SHB 2 | SHB 3 | SHB 4 | SHB 5 | SHB 6 | SHB 7 | SHB 8 | SHB 9 | SHB 10 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Crithidia. mellificae | POS | - | - | POS | - | POS | - | - | POS | - | - |
| Lotmaria passim | POS | - | POS | - | POS | - | POS | - | - | POS | - |
| KBV | POS * | POS * | - | - | - | - | - | - | - | - | POS * |
| DWV | POS * | POS * | POS * | POS * | - | POS * | POS * | - | POS * | POS * | - |
| Target | Primer Name | Sequence (5′-3′) | Reference |
|---|---|---|---|
| Paenibacillus larvae | AFB-F | CTTGTGTTTCTTTCGGGAGACGCCA | [82] |
| AFB-R | TCTTAGAGTGCCCACCTCTGCG | ||
| Melissococcus plutonius | MelissoF | CAGCTAGTCGGTTTGGTTCC | [83] |
| MelissoR | TTGGCTGTAGATAGAATTGACAAT | ||
| Crithida mellificae | Cmel_Cyt_b_F | TAAATTCACTACCTCAAATTCAATAACATAATCAT | [84] |
| Cmel_Cyt_b_R | ATTTATTGTTGTAATCGGTTTTATTGGATATGT | ||
| Lotmaria passim | Lp2F 459 | AGGGATATTTAAACCCATCGAA | [33] |
| Lp2R 459 | ACCACAAGAGTACGGAATGC |
| Target | Primer Name | Sequence (5′-3′) | Reference |
|---|---|---|---|
| KBV | KBV 83F | ACCAGGAAGTATTCCCATGGTAAG | [85] |
| KBV 161R | TGGAGCTATGGTTCCGTTCAG | ||
| DWV | DWV Fw 8450 | TGGCATGCCTTGTTCACCGT | [47] |
| DWV Rev 8953 | CGTGCAGCTCGATAGGATGCCA | ||
| ABPV | APV 95F | TCCTATATCGACGACGAAAGACAA | [85] |
| APV 159R | GCGCTTTAATTCCATCCAATTGA | ||
| IAPV | IAPV B4S0427_R130M | RCRTCAGTCGTCTTCCAGGT | [86] |
| IAPV B4S0427_L17M | CGAACTTGGTGACTTGARGG | ||
| BQCV | BQCV 9195F | GGTGCGGGAGATGATATGGA | [85] |
| BQCV 8265R | GCCGTCTGAGATGCATGAATAC | ||
| SBV | SBV 311F 79 | AAGTTGGAGGCGCGyAATTG | [85] |
| SBV 380R | CAAATGTCTTCTTACdAGAGGyAAGGATTG | ||
| CBPV | CPV 304F 79 | TCTGGCTCTGTCTTCGCAAA | [85] |
| CPV 371R | GATACCGTCGTCACCCTCATG | ||
| SPV major | SPV 8383F 81 | TGATTGGACTCGGCTTGCTA | [59] |
| SPV 8456R | CAAAATTTGCATAATCCCCAGTT | ||
| SPV minor | SPV Minor F1 | ATAGCGCTTTAGTTCAATTGCCAT | [38] |
| SPV Minor R1 | CTGGAATATGACCATCACGCAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nanetti, A.; Ellis, J.D.; Cardaio, I.; Cilia, G. Detection of Lotmaria passim, Crithidia mellificae and Replicative Forms of Deformed Wing Virus and Kashmir Bee Virus in the Small Hive Beetle (Aethina tumida). Pathogens 2021, 10, 372. https://doi.org/10.3390/pathogens10030372
Nanetti A, Ellis JD, Cardaio I, Cilia G. Detection of Lotmaria passim, Crithidia mellificae and Replicative Forms of Deformed Wing Virus and Kashmir Bee Virus in the Small Hive Beetle (Aethina tumida). Pathogens. 2021; 10(3):372. https://doi.org/10.3390/pathogens10030372
Chicago/Turabian StyleNanetti, Antonio, James D. Ellis, Ilaria Cardaio, and Giovanni Cilia. 2021. "Detection of Lotmaria passim, Crithidia mellificae and Replicative Forms of Deformed Wing Virus and Kashmir Bee Virus in the Small Hive Beetle (Aethina tumida)" Pathogens 10, no. 3: 372. https://doi.org/10.3390/pathogens10030372
APA StyleNanetti, A., Ellis, J. D., Cardaio, I., & Cilia, G. (2021). Detection of Lotmaria passim, Crithidia mellificae and Replicative Forms of Deformed Wing Virus and Kashmir Bee Virus in the Small Hive Beetle (Aethina tumida). Pathogens, 10(3), 372. https://doi.org/10.3390/pathogens10030372