
Cytauxzoonosis in North America

 ,
,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Etiologic Agent
3. Affected Species
3.1. Domestic Cats
3.2. Bobcats
3.3. Puma and Florida Panthers
3.4. Other Wild and Exotic Felids
4. Life Cycle and Pathogenesis
5. Clinical Signs
6. Diagnosis
7. Epizootiology
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Wagner, J.E. A fatal cytauxzoonosis-like disease in cats. J. Am. Vet. Med. Assoc. 1976, 168, 585–588. [Google Scholar]
- Bendele, R.; Schwartz, W.; Jones, L. Cytauxzoonosis-like disease in Texas cats. Southwest. Vet. 1976, 23, 244–245. [Google Scholar]
- Kier-Schroeder, A.B. The Etiology and Pathogenesis of Feline Cytauxzoonosis; University of Missouri: Columbia, MO, USA, 1979. [Google Scholar]
- André, M.R.; Herrera, H.M.; de Jesus Fernandes, S.; de Sousa, K.C.M.; Gonçalves, L.R.; Domingos, I.H.; de Macedo, G.C.; Machado, R.Z. Tick-borne agents in domesticated and stray cats from the city of Campo Grande, state of Mato Grosso do Sul, midwestern Brazil. Ticks Tick-Borne Dis. 2015, 6, 779–786. [Google Scholar] [CrossRef]
- Peixoto, P.; Soares, C.; Scofield, A.; Santiago, C.; Franca, T.; Barros, S. Fatal cytauxzoonosis in captive-reared lions in Brazil. Vet. Parasitol. 2007, 145, 383–387. [Google Scholar] [CrossRef] [PubMed]
- André, M.R.; Adania, C.H.; Machado, R.Z.; Allegretti, S.M.; Felippe, P.A.; Silva, K.F.; Nakaghi, A.C.; Dagnone, A.S. Molecular detection of Cytauxzoon spp. in asymptomatic Brazilian wild captive felids. J. Wildl. Dis. 2009, 45, 234–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaspari, M.M.F. Estudo Epidemiológico de Patógenos Circulantes nas Populações de onça-Pintada e Animais Domésticos em áreas Preservadas de três Biomas Brasileiros: Cerrado, Pantanal e Amazônia. Universidade de São Paulo. 2010. Available online: https://www.teses.usp.br/teses/disponiveis/10/10134/tde-05102012-134828/pt-br.php (accessed on 30 August 2021).
- Yabsley, M.J.; Murphy, S.M.; Cunningham, M.W. Molecular detection and characterization of Cytauxzoon felis and a Babesia species in cougars from Florida. J. Wildl. Dis. 2006, 42, 366–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penzhorn, B.L.; Oosthuizen, M.C. Babesia species of domestic cats: Molecular characterization has opened Pandora’s Box. Front. Vet. Sci. 2020, 7, 134. [Google Scholar] [CrossRef]
- Alvarado-Rybak, M.; Solano-Gallego, L.; Millán, J. A review of piroplasmid infections in wild carnivores worldwide: Importance for domestic animal health and wildlife conservation. Parasit. Vectors 2016, 9, 538. [Google Scholar] [CrossRef] [Green Version]
- Panait, L.C.; Mihalca, A.D.; Modrý, D.; Juránková, J.; Ionică, A.M.; Deak, G.; Gherman, C.M.; Heddergott, M.; Hodžić, A.; Veronesi, F. Three new species of Cytauxzoon in European wild felids. Vet. Parasit. 2021, 290, 109344. [Google Scholar] [CrossRef]
- Reichard, M.V.; Van Den Bussche, R.A.; Meinkoth, J.H.; Hoover, J.P.; Kocan, A.A. A new species of Cytauxzoon from Pallas’ cats caught in mongolia and comments on the systematics and taxonomy of piroplasmids. J. Parasitol. 2005, 91, 420–426. [Google Scholar] [CrossRef]
- Neitz, W.O.; Thomas, A.D. Cytauxzoon sylvicaprae gen. nov., spec. nov., a protozoon responsible for a hitherto undescribed disease in the duiker, Sylvicapra grimmia (Linne). Onderstepoort J. Vet. Sci. Anim. Ind. 1948, 23, 63–76. [Google Scholar] [PubMed]
- Gubbels, M.-J.; Keroack, C.D.; Dangoudoubiyam, S.; Worliczek, H.L.; Paul, A.S.; Bauwens, C.; Elsworth, B.; Engelberg, K.; Howe, D.K.; Coppens, I. Fussing about fission: Defining variety among mainstream and exotic apicomplexan cell division modes. Front. Cell. Infect. Microbiol. 2020, 10, 269. [Google Scholar] [CrossRef] [PubMed]
- Schreeg, M.E.; Marr, H.S.; Tarigo, J.L.; Cohn, L.A.; Bird, D.M.; Scholl, E.H.; Levy, M.G.; Wiegmann, B.M.; Birkenheuer, A.J. Mitochondrial genome sequences and structures aid in the resolution of Piroplasmida phylogeny. PLoS ONE. 2016, 11, e0165702. [Google Scholar] [CrossRef] [PubMed]
- Shock, B.C.; Birkenheuer, A.J.; Patton, L.L.; Olfenbuttel, C.; Beringer, J.; Grove, D.M.; Peek, M.; Butfiloski, J.W.; Hughes, D.W.; Lockhart, J.M. Variation in the ITS-1 and ITS-2 rRNA genomic regions of Cytauxzoon felis from bobcats and pumas in the eastern United States and comparison with sequences from domestic cats. Vet. Parasitol. 2012, 190, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Pollard, D.A.; Reichard, M.V.; Cohn, L.A.; James, A.M.; Holman, P.J. Genetic variability of cloned Cytauxzoon felis ribosomal RNA ITS1 and ITS2 genomic regions from domestic cats with varied clinical outcomes from five states. Vet. Parasit. 2017, 244, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.M.; Berghaus, R.D.; Latimer, K.S.; Britt, J.O.; Rakich, P.M.; Peterson, D.S. Genetic variability of Cytauxzoon felis from 88 infected domestic cats in Arkansas and Georgia. J. Vet. Diagn. Investig. 2009, 21, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Brown, H.M.; Lockhart, J.M.; Latimer, K.S.; Peterson, D.S. Identification and genetic characterization of Cytauxzoon felis in asymptomatic domestic cats and bobcats. Vet. Parasitol. 2010, 172, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.M.; Modaresi, S.M.; Cook, J.L.; Latimer, K.S.; Peterson, D.S. Genetic variability of archived Cytauxzoon felis histologic specimens from domestic cats in Georgia, 1995–2007. J. Vet. Diagn. Investig. 2009, 21, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Schreeg, M.E.; Marr, H.S.; Tarigo, J.; Cohn, L.A.; Levy, M.G.; Birkenheuer, A.J. Pharmacogenomics of Cytauxzoon felis cytochrome b: Implications for atovaquone and azithromycin therapy in domestic cats with cytauxzoonosis. J. Clin. Microbiol. 2013, 51, 3066–3069. [Google Scholar] [CrossRef] [Green Version]
- Schreeg, M.E.; Marr, H.S.; Tarigo, J.L.; Cohn, L.A.; Levy, M.G.; Birkenheuer, A.J. Rapid high-resolution melt analysis of Cytauxzoon felis cytochrome b to aid in the prognosis of cytauxzoonosis. J. Clin. Microbiol. 2015, 53, 2517–2524. [Google Scholar] [CrossRef] [Green Version]
- Hartley, A.N.; Marr, H.S.; Birkenheuer, A.J. Cytauxzoon felis cytochrome b gene mutation associated with atovaquone and azithromycin treatment. J. Vet. Intern. Med. 2020, 34, 2432–2437. [Google Scholar] [CrossRef] [PubMed]
- Kier, A.B.; Wightman, S.R.; Wagner, J.E. Interspecies transmission of Cytauxzoon felis. Am. J. Vet. Res. 1982, 43, 102–105. [Google Scholar] [PubMed]
- Meinkoth, J.H.; Kocan, A.A. Feline cytauxzoonosis. Vet. Clin. N. Am. Small Anim. Pract. 2005, 35, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Sherrill, M.K.; Cohn, L.A. Cytauxzoonosis: Diagnosis and treatment of an emerging disease. J. Feline Med. Surg. 2015, 17, 940–948. [Google Scholar] [CrossRef]
- Haber, M.D.; Tucker, M.D.; Marr, H.S.; Levy, J.K.; Burgess, J.; Lappin, M.R.; Birkenheuer, A.J. The detection of Cytauxzoon felis in apparently healthy free-roaming cats in the USA. Vet. Parasitol. 2007, 146, 316–320. [Google Scholar] [CrossRef]
- Brown, H.M.; Latimer, K.S.; Erikson, L.E.; Cashwell, M.E.; Britt, J.O.; Peterson, D.S. Detection of persistent Cytauxzoon felis infection by polymerase chain reaction in three asymptomatic domestic cats. J. Vet. Diagn. Investig. 2008, 20, 485–488. [Google Scholar] [CrossRef] [Green Version]
- Meinkoth, J.; Kocan, A.A.; Whitworth, L.; Murphy, G.; Fox, J.C.; Woods, J.P. Cats surviving natural infection with Cytauxzoon felis: 18 cases (1997–1998). J. Vet. Intern. Med. 2000, 14, 521–525. [Google Scholar] [CrossRef]
- Rizzi, T.E.; Reichard, M.V.; Cohn, L.A.; Birkenheuer, A.J.; Taylor, J.D.; Meinkoth, J.H. Prevalence of Cytauxzoon felis infection in healthy cats from enzootic areas in Arkansas, Missouri, and Oklahoma. Parasit. Vectors 2015, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Wikander, Y.M.; Anantatat, T.; Kang, Q.; Reif, K.E. Prevalence of Cytauxzoon felis infection-carriers in eastern Kansas domestic cats. Pathogens 2020, 9, 854. [Google Scholar] [CrossRef]
- Duran, D.M. Determination of Ownership Practices and Cytauxzoon felis Prevalence in Domestic Felines of Northwest Arkansas. Animal Science Undergraduate Honors Theses. 2021. Available online: https://scholarworks.uark.edu/anscuht/44 (accessed on 30 August 2021).
- Cohn, L.A.; Birkenheuer, A.J. Cytauxzoonosis. In Infectious Diseases of the Dog and Cat, 4th ed.; Greene, C.E., Ed.; Elsevier: St. Louis, MO, USA, 2012; pp. 764–771. [Google Scholar]
- Ferris, D. A progress report on the status of a new disease of American cats: Cytauxzoonosis. Comp. Immunol. Microbiol. Infect. Dis. 1979, 1, 269–276. [Google Scholar] [CrossRef]
- Mueller, E.K.; Baum, K.A.; Papes, M.; Cohn, L.A.; Cowell, A.K.; Reichard, M.V. Potential ecological distribution of Cytauxzoon felis in domestic cats in Oklahoma, Missouri, and Arkansas. Vet. Parasitol. 2013, 192, 104–110. [Google Scholar] [CrossRef]
- Greene, C.; Latimer, K.; Hopper, E.; Shoeffler, G.; Lower, K.; Cullens, F. Administration of diminazene aceturate or imidocarb dipropionate for treatment of cytauxzoonosis in cats. J. Am. Vet. Med. Assoc. 1999, 215, 497–500. [Google Scholar]
- MacNeill, A.L.; Barger, A.M.; Skowronski, M.C.; Lanka, S.; Maddox, C.W. Identification of Cytauxzoon felis infection in domestic cats from southern Illinois. J. Feline Med. Surg. 2015, 17, 1069–1072. [Google Scholar] [CrossRef]
- Cytauxzoonosis in Southern Indiana. Available online: https://vet.purdue.edu/addl/news/Cytauxzoonosis%20in%20Southern%20Indiana.php (accessed on 31 August 2021).
- Nagamori, Y.; Slovak, J.E.; Reichard, M.V. Prevalence of Cytauxzoon felis infection in healthy free-roaming cats in north-central Oklahoma and central Iowa. JFMS Open Rep. 2016, 2. [Google Scholar] [CrossRef] [PubMed]
- Meier, H.T.; Moore, L.E. Feline cytauxzoonosis: A case report and literature review. J. Am. Anim. Hosp. Assoc. 2000, 36, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.B.; Fisher, T. Fatal cytauxzoonosis in a Kentucky cat (Felis domesticus). Vet. Parasitol. 2006, 139, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.; Davis, C.D. Increasing frequency of feline cytauxzoonosis cases diagnosed in western Kentucky from 2001 to 2011. Vet. Parasitol. 2013, 198, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Hauck, W.; Snider 3rd, T.; Lawrence, J. Cytauxzoonosis in a native Louisiana cat. J. Am. Vet. Med. Assoc. 1982, 180, 1472–1474. [Google Scholar]
- Birkenheuer, A.J.; Le, J.A.; Valenzisi, A.M.; Tucker, M.D.; Levy, M.G.; Breitschwerdt, E.B. Cytauxzoon felis infection in cats in the mid-Atlantic states: 34 cases (1998–2004). J. Am. Vet. Med. Assoc. 2006, 228, 568–571. [Google Scholar] [CrossRef] [PubMed]
- Glenn, B.L.; Stair, E.L. Cytauxzoonosis in domestic cats: Report of two cases in Oklahoma, with a review and discussion of the disease. J. Am. Vet. Med. Assoc. 1984, 184, 822–825. [Google Scholar]
- Hoover, J.; Walker, D.; Hedges, J. Cytauxzoonosis in cats: Eight cases (1985–1992). J. Am. Vet. Med. Assoc. 1994, 205, 455–460. [Google Scholar]
- Reichard, M.V.; Baum, K.A.; Cadenhead, S.C.; Snider, T.A. Temporal occurrence and environmental risk factors associated with cytauxzoonosis in domestic cats. Vet. Parasitol. 2008, 152, 314–320. [Google Scholar] [CrossRef]
- Nietfeld, J.C.; Pollock, C. Fatal cytauxzoonosis in a free-ranging bobcat (Lynx rufus). J. Wildl. Dis. 2002, 38, 607–610. [Google Scholar] [CrossRef] [Green Version]
- Blouin, E.F.; Kocan, A.A.; Kocan, K.M.; Hair, J. Evidence of a limited schizogonous cycle for Cytauxzoon felis in bobcats following exposure to infected ticks. J. Wildl. Dis. 1987, 23, 499–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shock, B.C.; Murphy, S.M.; Patton, L.L.; Shock, P.M.; Olfenbuttel, C.; Beringer, J.; Prange, S.; Grove, D.M.; Peek, M.; Butfiloski, J.W. Distribution and prevalence of Cytauxzoon felis in bobcats (Lynx rufus), the natural reservoir, and other wild felids in thirteen states. Vet. Parasitol. 2011, 175, 325–330. [Google Scholar] [CrossRef]
- Birkenheuer, A.J.; Marr, H.S.; Warren, C.; Acton, A.E.; Mucker, E.M.; Humphreys, J.G.; Tucker, M.D. Cytauxzoon felis infections are present in bobcats (Lynx rufus) in a region where cytauxzoonosis is not recognized in domestic cats. Vet. Parasitol. 2008, 153, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Zieman, E.A.; Jimenez, F.A.; Nielsen, C.K. Concurrent examination of bobcats and ticks reveals high prevalence of Cytauxzoon felis in southern Illinois. J. Parasitol. 2017, 103, 343–348. [Google Scholar] [CrossRef]
- Zieman, E.A.; Nielsen, C.K.; Jiménez, F.A. Chronic Cytauxzoon felis infections in wild-caught bobcats (Lynx rufus). Vet. Parasitol. 2018, 252, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Glenn, B.; Rolley, R.; Kocan, A. Cytauxzoon-like piroplasms in erythrocytes of wild-trapped bobcats in Oklahoma. J. Am. Vet. Med. Assoc. 1982, 181, 1251–1253. [Google Scholar]
- Harvey, J.W.; Dunbar, M.R.; Norton, T.M.; Yabsley, M.J. Laboratory findings in acute Cytauxzoon felis infection in cougars (Puma concolor couguar) in Florida. J. Zoo Wildl. Med. 2007, 38, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Rotstein, D.S.; Taylor, S.K.; Harvey, J.W.; Bean, J. Hematologic effects of cytauxzoonosis in Florida panthers and Texas cougars in Florida. J. Wildl. Dis. 1999, 35, 613–617. [Google Scholar] [CrossRef]
- Butt, M.T.; Bowman, D.; Barr, M.C.; Roelke, M.E. Iatrogenic transmission of Cytauxzoon felis from a Florida panther (Felix concolor coryi) to a domestic cat. J. Wildl. Dis. 1991, 27, 342–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakob, W.; Wesemeier, H.-H. A fatal infection in a Bengal tiger resembling cytauxzoonosis in domestic cats. J. Comp. Pathol. 1996, 114, 439–444. [Google Scholar] [CrossRef]
- Garner, M.; Lung, N.; Citino, S.; Greiner, E.; Harvey, J.; Homer, B. Fatal cytauxzoonosis in a captive-reared white tiger (Panthera tigris). Vet. Pathol. 1996, 33, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.M.; Cohn, L.A.; Downey, M.E.; Whitney, M.S.; Birkenheuer, A.J. Evaluation of Cytauxzoon felis infection status in captive-born wild felids housed in an area endemic for the pathogen. J. Am. Vet. Med. Assoc. 2012, 241, 1088–1092. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.M.; Cohn, L.A.; Birkenheuer, A.J. Lack of evidence for perinatal transmission of Cytauxzoon felis in domestic cats. Vet. Parasitol. 2012, 188, 172–174. [Google Scholar] [CrossRef] [PubMed]
- Reichard, M.V.; Edwards, A.C.; Meinkoth, J.H.; Snider, T.A.; Meinkoth, K.R.; Heinz, R.E.; Little, S.E. Confirmation of Amblyomma americanum (Acari: Ixodidae) as a vector for Cytauxzoon felis (Piroplasmorida: Theileriidae) to Domestic Cats. J. Med. Entomol. 2010, 47, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Reichard, M.V.; Meinkoth, J.H.; Edwards, A.C.; Snider, T.A.; Kocan, K.M.; Blouin, E.F.; Little, S.E. Transmission of Cytauxzoon felis to a domestic cat by Amblyomma americanum. Vet. Parasitol. 2009, 161, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Reichard, M.V.; Rugg, J.J.; Thomas, J.E.; Allen, K.E.; Barrett, A.W.; Murray, J.K.; Herrin, B.H.; Beam, R.A.; King, V.L.; Vatta, A.F. Efficacy of a topical formulation of selamectin plus sarolaner against induced infestations of Amblyomma americanum on cats and prevention of Cytauxzoon felis transmission. Vet. Parasitol. 2018, 270, S31–S37. [Google Scholar] [CrossRef]
- Reichard, M.V.; Thomas, J.E.; Arther, R.G.; Hostetler, J.A.; Raetzel, K.L.; Meinkoth, J.H.; Little, S.E. Efficacy of an imidacloprid 10%/flumethrin 4.5% collar (Seresto (R), Bayer) for preventing the transmission of Cytauxzoon felis to domestic cats by Amblyomma americanum. Parasitol. Res. 2013, 112, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Allen, K.E.; Thomas, J.E.; Wohltjen, M.L.; Reichard, M.V. Transmission of Cytauxzoon felis to domestic cats by Amblyomma americanum nymphs. Parasites Vectors 2019, 12, 28. [Google Scholar] [CrossRef] [Green Version]
- Blouin, E.F.; Kocan, A.A.; Glenn, B.L.; Kocan, K.M.; Hair, J.A. Transmission of Cytauxzoon felis Kier, 1979 from bobcats, Felis rufus (Schreber), to domestic cats by Dermacentor variabilis (Say). J. Wildl. Dis. 1984, 20, 241–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagamori, Y. Transmission of Cytauxzoon felis by Amblyomma americanum: Engorgement Weight of Nymphs and Attachment Time of Adults for Transmission to Domestic Cats; Oklahoma State University: Stillwater, OK, USA, 2016. [Google Scholar]
- Thomas, J.E.; Ohmes, C.M.; Payton, M.E.; Hostetler, J.A.; Reichard, M.V. Minimum transmission time of Cytauxzoon felis by Amblyomma americanum to domestic cats in relation to duration of infestation, and investigation of ingestion of infected ticks as a potential route of transmission. J. Feline Med. Surg. 2017, 20, 67–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kier, A.; Wagner, J.; Kinden, D. The pathology of experimental cytauxzoonosis. J. Comp. Pathol. 1987, 97, 415–432. [Google Scholar] [CrossRef]
- Snider, T.; Confer, A.; Payton, M. Pulmonary histopathology of Cytauxzoon felis infections in the cat. Vet. Pathol. 2010, 47, 698–702. [Google Scholar] [CrossRef]
- Jalovecka, M.; Hajdusek, O.; Sojka, D.; Kopacek, P.; Malandrin, L. The complexity of piroplasms life cycles. Front. Cell. Infect. Microbiol. 2018, 8, 248. [Google Scholar] [CrossRef] [Green Version]
- Wightman, S.R. Clinical Features, Interspecies Susceptibility, and Biological Transmission of Experimentally Induced Feline Cytauxzoonosis; University of Missouri: Columbia, SC, USA, 1978. [Google Scholar]
- Bondy, P.J.; Cohn, L.A.; Tyler, J.W.; Marsh, A.E. Polymerase chain reaction detection of Cytauxzoon felis from field-collected ticks and sequence analysis of the small subunit and internal transcribed spacer 1 region of the ribosomal RNA gene. J. Parasitol. 2005, 91, 458–461. [Google Scholar] [CrossRef]
- Shock, B.C.; Moncayo, A.; Cohen, S.; Mitchell, E.A.; Williamson, P.C.; Lopez, G.; Garrison, L.E.; Yabsley, M.J. Diversity of piroplasms detected in blood-fed and questing ticks from several states in the United States. Ticks Tick-Borne Dis. 2014, 5, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.N.; Sundstrom, K.D.; Duncan, K.T.; Ientile, M.M.; Jordy, J.; Ghosh, P.; Little, S.E. Show us your ticks: A survey of ticks infesting dogs and cats across the USA. Parasites Vectors 2019, 12, 1–11. [Google Scholar] [CrossRef]
- Saleh, M.N.; Allen, K.E.; Lineberry, M.W.; Little, S.E.; Reichard, M.V. Ticks infesting dogs and cats in North America: Biology, geographic distribution, and pathogen transmission. Vet. Parasitol. 2021, 2021, 109392. [Google Scholar] [CrossRef] [PubMed]
- Duncan, K.T.; Saleh, M.N.; Sundstrom, K.D.; Little, S.E. Dermacentor variabilis is the predominant Dermacentor spp.(Acari: Ixodidae) feeding on fogs and cats throughout the United States. J. Med. Entomol. 2021, 58, 1241–1247. [Google Scholar] [CrossRef]
- Dahlgren, F.S.; Paddock, C.D.; Springer, Y.P.; Eisen, R.J.; Behravesh, C.B. Expanding range of Amblyomma americanum and simultaneous changes in the epidemiology of spotted fever group rickettsiosis in the United States. Am. J. Trop. Med. Hyg. 2016, 94, 35–42. [Google Scholar] [CrossRef]
- Raghavan, R.K.; Peterson, A.T.; Cobos, M.E.; Ganta, R.; Foley, D. Current and future distribution of the Lone Star tick, Amblyomma americanum (L.) (Acari: Ixodidae) in North America. PLoS ONE 2019, 14, e0209082. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Onofrio, V.C.; Barros-Battesti, D.M. The ticks (Acari: Ixodida: Argasidae, ixodidae) of Brazil. Syst. Appl. Acarol. 2009, 14, 30–46. [Google Scholar] [CrossRef]
- Wagner, J.E.; Ferris, D.H.; Kier, A.B.; Wightman, S.R.; Maring, E.; Morehouse, L.G.; Hansen, R.D. Experimentally induced cytauxzoonosis-like disease in domestic cats. Vet. Parasitol. 1980, 6, 305–311. [Google Scholar] [CrossRef]
- Clarke, L.L.; Rissi, D.R. Neuropathology of natural Cytauxzoon felis infection in domestic cats. Vet. Pathol. 2015, 52, 1167–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohn, L.A.; Birkenheuer, A.J.; Brunker, J.D.; Ratcliff, E.R.; Craig, A.W. Efficacy of atovaquone and azithromycin or imidocarb dipropionate in cats with acute cytauxzoonosis. J. Vet. Intern. Med. 2011, 25, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.M.; Cohn, L.A.; Marr, H.S.; Birkenheuer, A.J. Failure of efficacy and adverse events associated with dose-intense diminazene diaceturate treatment of chronic Cytauxzoon felis infection in five cats. J. Feline Med. Surg. 2014, 16, 157–163. [Google Scholar] [CrossRef]
- Tarigo, J.L.; Scholl, E.H.; Bird, D.M.; Brown, C.C.; Cohn, L.A.; Dean, G.A.; Levy, M.G.; Doolan, D.L.; Trieu, A.; Nordone, S.K.; et al. A novel candidate vaccine for cytauxzoonosis inferred from comparative apicomplexan genomics. PLoS ONE 2013, 8, e71233. [Google Scholar] [CrossRef]
- Melco, A.L.; Petrovitch, N.P.; Birkenheuer, A.J.; Yang, T.S.; Schreeg, M.E.; Savage, M. Pathognomonic thoracic radiographic findings are lacking in cats with acute cytauxzoonosis. Vet. Radiol. Ultrasound 2021, 62, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Birkenheuer, A.J.; Marr, H.; Alleman, A.R.; Levy, M.G.; Breitschwerdt, E.B. Development and evaluation of a PCR assay for the detection of Cytauxzoon felis DNA in feline blood samples. Vet. Parasitol. 2006, 137, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Schreeg, M.E.; Marr, H.S.; Griffith, E.H.; Tarigo, J.L.; Bird, D.M.; Reichard, M.V.; Cohn, L.A.; Levy, M.G.; Birkenheuer, A.J. PCR amplification of a multi-copy mitochondrial gene (cox3) improves detection of Cytauxzoon felis infection as compared to a ribosomal gene (18S). Vet. Parasitol. 2016, 225, 123–130. [Google Scholar] [CrossRef]
- Kao, Y.-F.; Peake, B.; Madden, R.; Cowan, S.R.; Scimeca, R.C.; Thomas, J.E.; Reichard, M.V.; Ramachandran, A.; Miller, C.A. A probe-based droplet digital polymerase chain reaction assay for early detection of feline acute cytauxzoonosis. Vet. Parasitol. 2021, 292, 109413. [Google Scholar] [CrossRef] [PubMed]
- Susta, L.; Torres-Velez, F.; Zhang, J.; Brown, C. An in situ hybridization and immunohistochemical study of cytauxzoonosis in domestic cats. Vet. Pathol. 2009, 46, 1197–1204. [Google Scholar] [CrossRef] [Green Version]
- Ogden, N.H.; Ben Beard, C.; Ginsberg, H.S.; Tsao, J.I. Possible effects of climate change on ixodid ticks and the pathogens they transmit: Predictions and observations. J. Med. Entomol. 2020, 58, 1536–1545. [Google Scholar] [CrossRef]
- Wikander, Y.M.; Kang, Q.; Reif, K.E. Acute Cytauxzoon felis cases in domestic cats from eastern Kansas, a retrospective case-control study. Vet. Sci. 2020, 7, 205. [Google Scholar] [CrossRef] [PubMed]
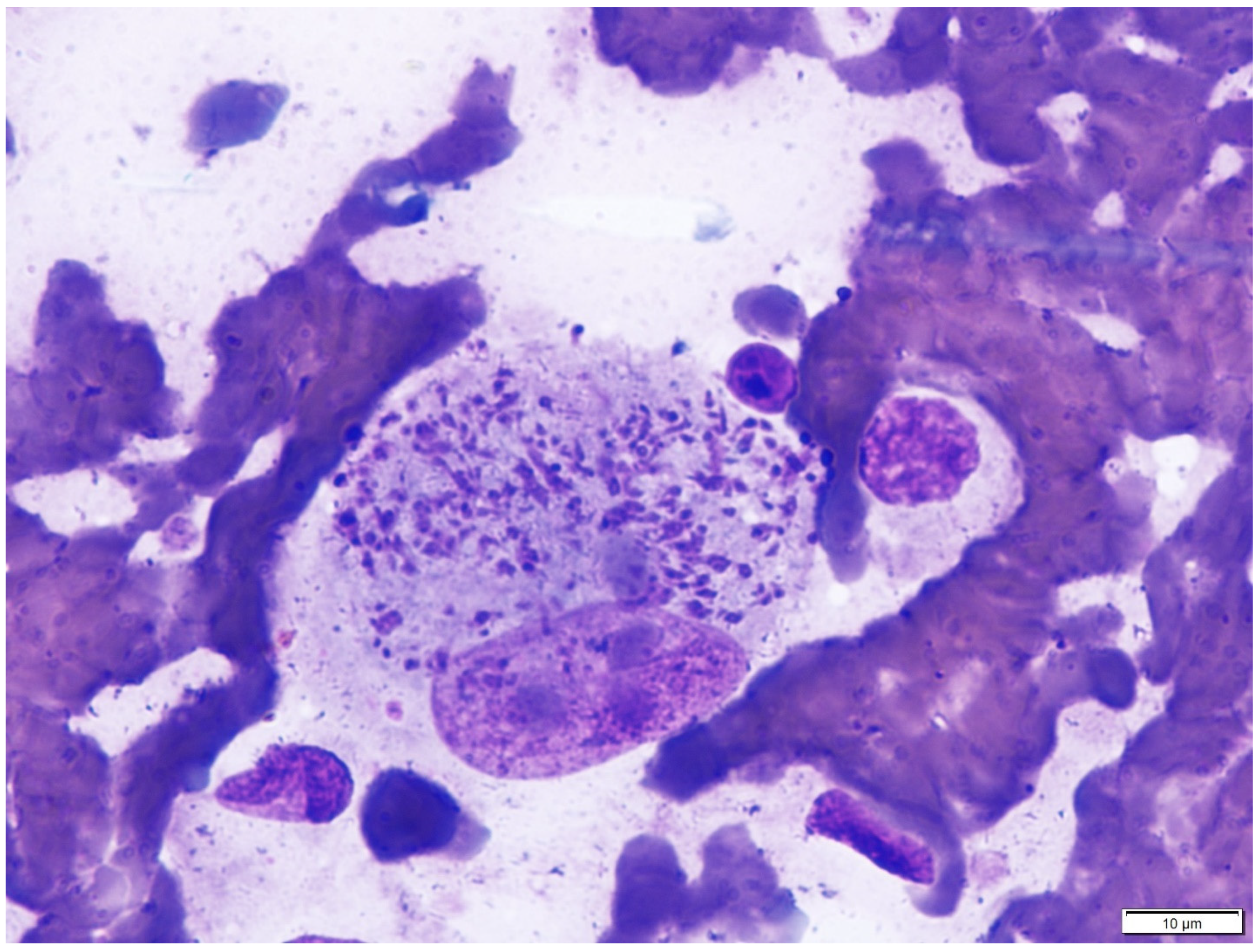






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| State | No. of Cats Tested | No. of Cats Infected | Prevalence; 95% Confidence Interval | Sample Period | Sample Type | Test Method | Reference |
|---|---|---|---|---|---|---|---|
| Alabama | NA | NA | NA | NA | NA | NA | [33] |
| Arkansas | NA | NA | NA | NA | NA | NA | [34] |
| NA | 18 | NA | 1997–1998 | blood | microscopy and PCR for 18S | [29] | |
| NA | 3 | NA | NA | blood | microscopy and PCR for ITS 1&2 | [28] | |
| NA | 57 | NA | 2005–2007 | blood | microscopy and PCR for ITS 1&2 | [18] | |
| 43 | 18 | 41.9%; 28.4–56.7% | NA | blood | microscopy and PCR for ITS 1&2 | [19] | |
| NA | 12 | NA | 1998–2011 | blood or other tissue | microscopy | [35] | |
| 161 | 25 | 15.5%; 10.7–22.0% | 2008–2012 | blood | PCR for18S | [30] | |
| 22 | 4 | 18.2%; 6.7–39.1% | 2020–2021 | blood | PCR for cox3 | [32] | |
| Florida | 494 | 1 | 0.2%; <0.1–1.3% | 1999–2000 | blood | PCR for18S | [27] |
| Georgia | NA | NA | NA | NA | NA | NA | [34] |
| NA | 9 | NA | NA | blood or other tissue | microscopy | [36] | |
| NA | 31 | NA | 2005–2007 | blood | microscopy and PCR for ITS 1&2 | [18] | |
| 46 | 9 | 19.6%; 10.4–33.4% | NA | blood | microscopy and PCR for ITS 1&2 | [19] | |
| Illinois | 59536 | 12 | NA | 2003–2012 | blood and spleen | microscopy and PCR for ITS 1&2 | [37] |
| Indiana | NA | NA | NA | NA | NA | NA | [38] |
| Iowa | 292 | 0 | 0.0%; 0.0–1.3% | 2012–2014 | blood | PCR for18S | [39] |
| Kansas | NA | 1 | NA | NA | lung and liver | microscopy | [40] |
| 1104 | 270 | 25.8%; 22.0–27.1% | 2018–2019 | blood | PCR: cox3 | [31] | |
| Kentucky | NA | 1 | NA | NA | brain, heart, lung, intestine, spleen, lymph node, and kidney | microscopy | [41] |
| NA | 56 | NA | 2001–2011 | blood or other tissue | microscopy | [42] | |
| Louisiana | NA | 1 | NA | NA | blood and other tissue | microscopy | [43] |
| Mississippi | NA | NA | NA | NA | NA | NA | [33] |
| Missouri | NA | 4 | NA | 1973–1975 | liver, lung, spleen, and lymph nodes | microscopy | [1] |
| NA | 68 | NA | 1998–2011 | blood or other tissue | microscopy | [35] | |
| 62 | 8 | 12.9%; 6.4–23.7% | 2008–2012 | blood | PCR for18S | [30] | |
| North Carolina | NA | 28 | NA | 1998–2004 | blood or other tissue | microscopy | [44] |
| 392 | 0 | 0.0%; 0.0–1.2% | 1999–2000 | Blood | PCR for18S | [27] | |
| Oklahoma | NA | 2 | NA | 1984 | blood or other tissue | microscopy | [45] |
| NA | 8 | NA | 1985–1992 | blood or other tissue | microscopy | [46] | |
| NA | 18 | NA | 1997–1998 | blood | microscopy and PCR for 18S | [29] | |
| NA | 232 | NA | 1995–2006 | blood or other tissue | microscopy | [47] | |
| NA | 130 | NA | 1998–2011 | blood or other tissue | microscopy | [35] | |
| 679 | 23 | 3.4%; 2.2–5.1% | 2008–2012 | blood | PCR for18S | [30] | |
| 380 | 3 | 0.79%; 0.2–2.4% | 2012–2014 | blood | PCR for18S | [39] | |
| South Carolina | NA | 3 | NA | 1998–2004 | blood or other tissue | microscopy | [44] |
| Tennessee | 75 | 1 | 1.3%; <0.1–7.9% | 2006 | blood | PCR for18S | [27] |
| Texas | NA | 2 | NA | NA | tissue | microscopy | [2] |
| Virginia | NA | 3 | NA | 1998–2004 | blood or other tissue | microscopy | [44] |
| State | No. of Bobcats Tested | No. of Bobcats Infected | Prevalence; 95% Confidence Interval | Sampling Period | Sample Tested | Test Method | Reference |
|---|---|---|---|---|---|---|---|
| Arkansas | 6 | NA | NA | NA | spleen | real-time PCR for 18S; PCR for ITS1 and ITS2 | [19] |
| California | 26 | 0 | 0.0%; 0.0–15.2% | 1999–2010 | blood or spleen | nested PCR for ITS1 | [50] |
| Colorado | 67 | 0 | 0.0%; 0.0–6.5% | 1999–2010 | blood or spleen | nested PCR for ITS1 | [50] |
| Florida | 45 | 16 | 35.6%; 23.2–50.2% | 1999–2010 | blood or spleen | nested PCR for ITS1 | [50] |
| 54 | NA | NA | NA | spleen | real-time PCR for 18S; PCR for ITS1 and ITS2 | [19] | |
| Georgia | 143 | 13 | 9.1%; 5.3–15.1% | 1999–2010 | blood or spleen | nested PCR for ITS1 | [50] |
| 73 | NA | NA | NA | spleen | real-time PCR for 18S; PCR for ITS1 and ITS2 | [19] | |
| Illinois | 125 | 88 | 70.4%; 61.9–77.7% | 2003–2015 | blood or spleen | nested PCR for 18S | [52] |
| Kansas | 39 | 12 | 30.8%; 18.5–46.5% | 1999–2010 | blood or spleen | nested PCR for ITS1 | [50] |
| 1 | 1 | NA | 2000 | microscopy | [48] | ||
| Kentucky | 74 | 41 | 55.4%; 44.1–66.2% | 1999–2010 | blood or spleen | nested PCR for ITS1 | [50] |
| Missouri | 39 | 31 | 79.5%; 64.2–89.5% | 1999–2010 | blood or spleen | nested PCR for ITS1 | [50] |
| North Carolina | 30 | 10 | 33.3%; 19.3–51.3% | 2004, 2005, 2006 | blood | PCR for18S | [51] |
| 8 | 5 | 62.5%; 30.4–86.5% | 1999–2010 | blood or spleen | nested PCR for ITS1 | [50] | |
| North Dakota | 172 | 3 | 1.7%; 0.4–5.2% | 1999–2010 | blood or spleen | nested PCR for ITS1 | [50] |
| Pennsylvania | 69 | 5 | 7.3%; 2.8–16.2% | 2002 | blood | PCR for 18S | [51] |
| Ohio | 19 | 0 | 0.0%; 0.0–19.8% | 1999–2010 | blood | nested PCR for ITS1 | [50] |
| Oklahoma | 10 | NA | NA | 1982–1984 | blood, liver, spleen, lung, or lymph nodes | microscopy | [49] |
| 20 | 13 | 65.0%; 43.2–82.0 | 1999–2010 | blood or spleen | nested PCR for ITS1 | [50] | |
| 26 | 13 | 50.0%; 32.1–67.9% | ~1982 | blood | microscopy | [54] | |
| South Carolina | 7 | 4 | 57.1%; 25.0–84.3% | 1999–2010 | blood or spleen | nested PCR for ITS1 | [50] |
| West Virginia | 37 | 0 | 0.0%; 0.0–11.2% | 1999–2010 | blood or spleen | nested PCR for ITS1 | [50] |
| State | Tick Species | Tick Life Stage | No. of Ticks or Tick Pools Tested | No. of Ticks or Tick Pools Infected | Prevalence or Minimum Infection Rate (95% Confidence Interval) | Reference |
|---|---|---|---|---|---|---|
| Georgia | Amblyomma americanum | NR * | 340 | 0 | 0.0% (0.0–1.4%) | [75] |
| Dermacentor variabilis | NR | 125 | 1 | 0.8% (<0.1–4.8%) | ||
| Amblyomma maculatum | NR | 16 | 0 | 0.0% (0.0–22.7%) | ||
| Ixodes scapularis | NR | 3 | 0 | 0.0% (0.0–61.8%) | ||
| Amblyomma spp. | NR | 2 | 0 | 0.0% (0.0–71.0%) | ||
| Illinois | A. americanum | female | 57 | 8 | 14.0% (7.0–25.6%) | [52] |
| male | 60 | 10 | 16.7% (9.1–28.3%) | |||
| D. variabilis | female | 51 | 10 | 19.6% (10.8–32.6%) | ||
| male | 50 | 6 | 12.0% (5.3–24.1%) | |||
| Kentucky | A. americanum | NR | 61 | 0 | 0.0% (0.0–7.1%) | [75] |
| D. variabilis | NR | 42 | 0 | 0.0% (0.0–10.0%) | ||
| Missouri | A. americanum | adult | 210 | 0 | 0.0% (0.0–2.2%) | [74] |
| nymph | 16 | 3 † | 18.8% (5.8–43.8%) † | |||
| D. variabilis | adult | 79 | 0 | 0.0% (0.0–5.6%) | ||
| Rhipicephalus sanguineus | adult | 35 | 0 | 0.0% (0.0–11.8%) | ||
| Oklahoma | A. americanum | female | 49 | 3 | 1.9% (1.5–17.2%) | [62] |
| male | 46 | 1 | 0.7% (<0.1–12.4%) | |||
| nymph | 80 | 3 | 0.9% (0.8–10.9%) | |||
| D. variabilis | female | 23 | 0 | 0.0% (0.0–16.9%) | ||
| male | 28 | 0 | 0.0% (0.0–14.3%) | |||
| Pennsylvania | Ixodes scapularis | NR | 1 | 0 | 0.0% (0.0–83.3%) | [75] |
| Tennessee | A. americanum | NR | 184 | 0 | 0.0% (0.0–2.5%) | [75] |
| D. variabilis | NR | 442 | 8 | 1.8% (0.9–3.6%) | ||
| Texas | A. americanum | NR | 158 | 0 | 0.0% (0.0–2.9%) | [75] |
| D. variabilis | NR | 93 | 0 | 0.0% (0.0–4.8%) | ||
| Amblyomma cajennense | NR | 99 | 0 | 0.0% (0.0–4.5%) | ||
| Amblyomma spp. | NR | 64 | 0 | 0.0% (0.0–6.8%) | ||
| Ixodes woodi | NR | 1 | 0 | 0.0% (0.0–83.3%) |
| Method * | Host Sample | Gene Target | Amplicon Length | Primers | Reference |
|---|---|---|---|---|---|
| PCR | Blood, other tissues | 18S | 284 bp | F: 5′-GCGAATCGCATTGCTTTATGCT-3′ R: 5′-CAATTGATACTCCGGAAAGAG-3′ | [88] |
| PCR | Blood, other tissues | cytb | 1203 bp | F: 5′-AGGATACAGGGCTATAACCAAC-3′ R: 5′-GTACTCTGGCTATGTCAATTTCTAC-3′ | [22] |
| PCR | Blood, other tissues | ITS2, partial 5.8S and 28S | 431 bp | F: 5′-TGAACGTATTAGACACACCACCT-3′ R: 5′-TCCTCCCGCTTCACTCGCCG-3′ | [28] |
| PCR | Blood, other tissues | 18S | 82 bp | F: 5′-TGC ATC ATT TAT ATT CCT TAA TCG-3′ R: 5′-CAA TCT GGA TAA TCA TAC CGA AA-3′ | [19] |
| PCR | Blood, other tissues | ITS1 | 651 bp (domestic cats) | F: 5′-CGA TCG AGT GAT CCG GTG AAT TA-3′ R: 5′-GCT GCG TCC TTC ATC GAT GTG-3′ | [19] |
| PCR | Blood, other tissues | 746 bp (bobcats) | F: 5′-CGA TCG AGT GAT CCG GTG AAT TA-3′ R: 5′-GGA GTA CCA CAT GCA AGC AG-3′ | ||
| PCR | Blood, other tissues | ITS2 | 475 bp | F: 5′-AGC GAA TTG CGA TAA GCA TT 3′ R: 5′-TCA GCC GTT ACT AGG AGA-3′ | [19] |
| PCR | Blood, other tissues | 18S | Primary—700 bp | F: 5′-ACCTGGTTGATCCTGCCAGTAGTCATATGCTTG-3′ R5′-TCACCAGAAAAAGCCACAAC-3′ | [74] |
| Nested—289 bp | F: 5′-TCGCATTGCTTTATGCTGGCGATG-3′ R: 5′-GCCCTCCAATTGATACTCCGGAAA-3′ | [63] | |||
| ddPCR | Blood, other tissues | cox3 | 118 bp | F: 5′-CTACACTCTTTACACGTTTGTG -3′ R: 5′-AGGAGTATACTGGCATTTCG -3′ | [90] |
| ISH | Formalin-fixed, paraffin-embedded tissues | 16S-like | 600 bp | F: 5′-CATGTCTTAGTATAAGCTTTTATACAGAA-3′ R: 5′-AACGCTGCGGAAGCGAGATTAATGACAAGGCAG-3′ | [91] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reichard, M.V.; Sanders, T.L.; Weerarathne, P.; Meinkoth, J.H.; Miller, C.A.; Scimeca, R.C.; Almazán, C. Cytauxzoonosis in North America. Pathogens 2021, 10, 1170. https://doi.org/10.3390/pathogens10091170
Reichard MV, Sanders TL, Weerarathne P, Meinkoth JH, Miller CA, Scimeca RC, Almazán C. Cytauxzoonosis in North America. Pathogens. 2021; 10(9):1170. https://doi.org/10.3390/pathogens10091170
Chicago/Turabian StyleReichard, Mason V., Tiana L. Sanders, Pabasara Weerarathne, James H. Meinkoth, Craig A. Miller, Ruth C. Scimeca, and Consuelo Almazán. 2021. "Cytauxzoonosis in North America" Pathogens 10, no. 9: 1170. https://doi.org/10.3390/pathogens10091170
APA StyleReichard, M. V., Sanders, T. L., Weerarathne, P., Meinkoth, J. H., Miller, C. A., Scimeca, R. C., & Almazán, C. (2021). Cytauxzoonosis in North America. Pathogens, 10(9), 1170. https://doi.org/10.3390/pathogens10091170