Abstract
Ticks are a significant group of arthropod vectors that transmit a large variety of pathogens responsible for human and animal diseases worldwide. Ticks are the second biggest transmitters of vector-borne diseases, behind mosquitoes. However, in West Africa, there is often only limited knowledge of tick-borne diseases. With the scarcity of appropriate diagnostic services, the prevalence of tick-borne diseases is generally underestimated in humans. In this review, we provide an update on tick-borne pathogens reported in people, animals and ticks in West Africa by microscopic, immunological and molecular methods. A systematic search was conducted in PubMed and Google Scholar. The selection criteria included all studies conducted in West Africa reporting the presence of Rickettsia, Borrelia, Anaplasma, Ehrlichia, Bartonella, Coxiella burnetii, Theileria, Babesia, Hepatozoon and Crimean–Congo haemorrhagic fever viruses in humans, animals or ticks. Our intention is to raise awareness of tick-borne diseases amongst human and animal health workers in West Africa, and also physicians working with tourists who have travelled to the region.
1. Introduction
Ticks are obligate blood-feeding parasites belonging to the phylum Arthropoda, the class Arachnida, and the order Acari [1]. They are divided into two main families: Ixodidae (hard ticks), comprising over 700 species worldwide, and Argasidae (soft ticks), comprising roughly 200 species [2]. Currently, in Africa, domestic animals can be infested with up to ten genera of ticks seven Ixodidae and three Argasidae [1]. A recent study reported that several species and subspecies of hard ticks, notably of the genera Hyalomma, Rhipicephalus, Ixodes, and Amblyomma, infest animals in different parts of the continent [3]. While ticks prefer animal hosts, many species will also feed on humans [4]. Globally, after mosquitoes, ticks are the second biggest group of vectors and reservoirs of animal and human viral, bacterial and protozoal pathogens [5]. More than 80% of the world’s cattle population is at risk of contracting tick-borne diseases (TBDs) [6] which are, particularly in sub-Saharan Africa, a major threat to the health, welfare and productivity of livestock. The impact ticks have on their hosts relates to their blood-feeding which can lead to both direct pathologies (e.g., anaemia, skin infections) and indirect pathologies (e.g., pathogen transmission, immunosuppression) [7]. Tick-borne diseases are strongly influenced by many factors, including host distribution, tick abundance and seasonality, pathogen virulence, and climate (temperature, precipitation, humidity, and vegetation cover), all of which contribute to the emergence and re-emergence of tick-borne diseases. Climate change may impact the incidence of tick-borne diseases by increasing tick populations, the rate of contact between livestock and ticks, and the rate of contact between livestock and wildlife [8]. In Europe, studies have been carried out on the impact of climate change on tick-borne diseases. It has been reported that climate change is responsible for the extension of the range of Ixodes ricinus in the north and at higher altitudes, leading to an increase in the prevalence of tick-borne encephalitis. Climate change is also partly responsible for changes in the distribution of D. reticulatus [9]. Over the last 20 years, the incidence of tick-borne diseases (Lyme disease, tick-borne encephalitis, and Crimean–Congo haemorrhagic fever) has increased in both Europe and America [10]. Although this increase could be partly caused by climate change, other factors could also contribute, as tick-borne disease systems are quite complex [10]. In Africa, diseases transmitted by ticks to humans are poorly studied, making it difficult to measure the impact of climate change on these diseases. However, there has been an increase in the incidence of tick-borne relapsing fever, one of the best-studied tick-borne bacterial diseases, in Senegal since the 1970s, and its range has extended over 350 km to north-west Morocco due to increased drought conditions [10].
The term “West Africa” is used to describe the land spanning the entire western part of sub-Saharan Africa. Roughly, this includes the coastal countries north of the Gulf of Guinea to the Senegal River, the countries covered by the Niger River Basin, and the countries of the Sahel hinterland [11]. This 6,140,000 km2 zone represents one-fifth of the area of the African continent and includes 16 countries, namely Benin, Burkina Faso, Cape Verde, Côte d’Ivoire, Gambia, Ghana, Guinea, Guinea Bissau, Liberia, Mali, Mauritania, Niger, Nigeria, Senegal, Sierra Leone, and Togo [11] (Figure 1).

Figure 1.
Tick vectors of the spotted fever group rickettsioses in West Africa. Above Am. variegatum, vector of R. africae, the agent of African tick bite fever, below Rh. sanguineus, the main vector of R. conorii subsp. conorii, the agent of Mediterranean spotted fever. (A,C) = females, (B,D) = males.
In this review, we use a “One Health” approach to describe human and animal TBDs in West Africa. The One Health concept encourages cooperation between professionals working on animal, environmental, and human health to develop integrated solutions for complex problems that impact on the health of animals, humans, and the planet [12]. To carry out this review, we searched the PubMed and Google Scholar databases using the keywords: ticks, tick-borne diseases, humans, animal, West Africa, Rickettsia, Borrelia, Anaplasma, Ehrlichia, Bartonella, Coxiella, Theileria, Babesia, Hepatozoon, and Crimean–Congo haemorrhagic fever virus. Papers that were published between 1970 and 2020, including molecular biology and serology studies, were selected for review and included in this review if they were written in English or with an English abstract.
2. Bacterial Diseases
2.1. Tick-Borne Spotted Fever Group Rickettsioses
Bacteria of the genus Rickettsia are obligate intracellular organisms that comprise 31 species of which 17, to date, are considered human and/or animal pathogens [13]. The spotted fever group (SFG) of Rickettsia comprises around 30 species that, with the exception of Rickettsia felis which is transmitted by fleas and possibly mosquitoes [14,15,16], are transmitted mainly by ticks and can cause tick-borne spotted fever group rickettsioses in animals and/or people. The typhus group comprises two species, Rickettsia typhi and Rickettsia prowazekii, which are most commonly transmitted by fleas and human body lice, respectively, but have occasionally been associated with ticks [13]. In humans, tick-borne spotted fever group (SFG) rickettsioses manifest mainly as fever, headache, muscle pain, and a rash which may be maculopapular but also vesicular or purpuric in severe cases. There might also be an inoculation eschar at the site where the tick was attached and an inconsistent regional lymphadenopathy [13]. Laboratory investigations may show thrombocytopaenia, hyponatraemia, elevated transaminases, and hyperbilirubinaemia, and the diagnosis can be made by serological (immunofluorescence) or molecular (quantitative PCR or standard PCR, particularly efficient on eschar swabbing samples) methods [13,17].
Five pathogenic SFG rickettsia have been reported in West Africa: Rickettsia africae, the agent of African tick-bite fever (ATBF), R. conorii conorii, the agent of Mediterranean spotted fever (MSF), R. sibirica mongolitimonae, the agent of lymphangitis-associated rickettsioses, and R. aeschlimannii and R. massiliae, which are agents of emerging rickettsioses in humans [13] (Figure 1).
Rickettsia africae is transmitted by ticks of the genus Amblyomma, in particular Amblyomma variegatum in West Africa (Figure 2). The infection rate of R. africae in these ticks, which feed very readily on people, is often very high, up to 100% [9]. After malaria, ATBF is the most commonly documented aetiology of fever in travellers returning from sub-Saharan Africa [18]. More than 350 travel-associated cases of ATBF have been reported in travellers from EU member countries, North America, Australia, Argentina, and Japan. Most of these travellers were infected in South Africa. In West Africa, cases of ATBF have only been reported in travellers who had stayed in Gambia [18]. As is often the case in the rest of Africa, in West Africa ATBF is often poorly recognised by health workers, perhaps due to lack of knowledge of the disease and diagnostic laboratories, but also because the disease is most often benign, and eschars and mild rashes are more difficult to detect in black skin [13]. However, exposure to R. africae is very common in West Africa, with a prevalence of reactive antibodies reported to be 19.6% among blood donors and patients from Mauritania [19], between 1.1% and 25.4% in Guinea [20], and between 20.6% and 45.6% in malaria-negative patients with a recent fever in the villages of Dielmo and Ndiop in Senegal, respectively [21]. In the only serological study of animals from West Africa, the prevalence of antibodies against R. africae in farm animals ranged from 0.6% to 18.8% in Guinea [20]. It is important to note, however, that serological diagnostic methods are limited by cross-reactivity with other species of SFG Rickettsia and thus infections with other species may have contributed to the results.
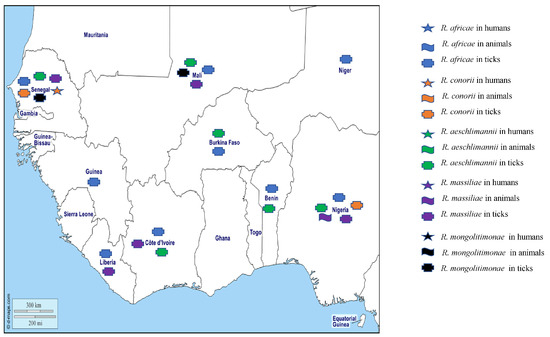
Figure 2.
Tick-borne rickettsiae detected by PCR in humans, animals, and ticks in West African countries.
In tick studies, DNA from R. africae has been found in Am. variegatum from Senegal, Benin, Mali, Côte d’Ivoire, Niger, Liberia, Guinea, Nigeria, and Burkina Faso [22,23,24,25,26,27,28,29,30,31,32,33], and in Amblyomma compressum from Liberia [26] (Table 1). Its DNA has also been found in a variety of other tick species including Hyalomma rufipes from Mali, Côte d’Ivoire, Guinea and Senegal [23,24,26,31], Hyalomma truncatum from Mali and Côte d’Ivoire [23,24], Hyalomma impeltatum from Nigeria [28], Hyalomma impressum from Côte d’Ivoire [24], Haemaphysalis paraleachi from Guinea [26], Rhipicephalus (Boophilus) annulatus from Senegal, Guinea, and Nigeria [21,26,30], Rhipicephalus evertsi evertsi from Mali, Senegal, and Nigeria [21,23,30], Rhipicephalus (Boophilus) decoloratus from Guinea and Nigeria, Rhipicephalus (Boophilus) geigyi, Rhipicephalus sanguineus s.l., and Rhipicephalus (Boophilus) microplus from Liberia, Nigeria, and Côte d’Ivoire, respectively [24,26,27,30], and recently in Rh. (B.) microplus from cattle from Mali [23] (Table 1). To date, only Amblyomma hebraeum and Am. variegatum have been shown to be vectors and reservoirs of R. africae [32]. The prevalence of R. africae in Am. variegatum is generally between 90% and 100%, due to the high rate of transovarian transmission and filial infection. The presence of R. africae DNA in other genera, often with varying prevalence (0.4% to 93%), does not prove that these ticks are also vectors, as they may have contained organisms in their digestive tract from blood meals taken from rickettsemic hosts, or from probable transovarial transmission [13]. Further studies are required to test the vector competence of these other tick species for R. africae.

Table 1.
Tick species found to be positive for tick-associated pathogens in different West African countries.
Rickettsia conorii conorii, the agent of MSF mainly transmitted by Rh. sanguineus s.l. [13], is one of the first disease agents which has been shown to be transmitted by arthropods. The disease is endemic in the Mediterranean region, where the majority of cases are encountered in the warmest months [75]. MSF is characterised by high fever, flu-like symptoms, a tache noir or ‘black spot’ representing an area of skin necrosis at the site of tick attachment, and a maculopapular rash. Neurological disorders and coagulopathies with multiple organ failure can occur in the severe forms of the disease [76]. In West Africa, R. conorii conorii has only been reported once, in a molecular study of a patient in Senegal who was negative for malaria but had a history of fever [21]. Bearing in mind the lack of specificity of serological tests (above), antibodies against R. conorii in indirect immunofluorescent antibody assays (IFAs) have been found in 13.5% of blood donors from Mauritania [19], and 38.2%, 31.5%, and 27% of apparently healthy adults (blood donors or volunteers) in Burkina Faso, Côte d’Ivoire, and Mali, respectively [77]. In the latter study, antibodies were detected using Western blots in 47.8%, 34.8%, and 30% of the patients. In ticks, DNA of R. conorii conorii was detected in 2/100 Rh. Sanguineus s.l. collected from rodents in Nigeria [70] and in 1/2360 Rh. E. evertsi collected in Senegal [21] (Table 1).
Rickettsia aeschlimannii was first isolated from Hyalomma marginatum, but several other tick species, particularly other Hyalomma spp., have now been reported as potential vectors [13,76]. The pathogenicity of this organism was demonstrated for the first time in France in a patient who travelled to Morocco in August 2000 and presented with a vesicular lesion of the ankle, a black spot with a fever of 39.5 °C, and a generalised maculopapular rash [78]. A second case was reported in a South African man who had been bitten by a Rhipicephalus appendiculatus during a hunt and was symptomless as he self-prescribed doxycycline immediately after removing the tick [76]. In West Africa, R. aeschlimannii has not been detected in humans and domestic animals. However, the DNA of R. aeschlimannii has been found in Hy. truncatum, Hy. rufipes, and Rh. sanguineus s.l. from Mali [23], Hy. rufipes collected in Côte d’Ivoire, Senegal, Burkina Faso, and Nigeria [21,24,31,33,52], Hy. truncatum collected in Côte d’Ivoire, Senegal, and Burkina Faso [21,24,33], Hy. impeltatum collected in Senegal and Nigeria [31,52], Rhipicephalus evertsi evertsi collected in Senegal [21,31], Am. variegatum, Rh. microplus, and Hy. rufipes collected in Benin [34], and Hy. rufipes, a Hy. impeltatum, Hy. truncatum, and Hy. dromedarii from Nigeria [49] (Table 1).
Rickettsia massiliae is transmitted by Rh. sanguineus s.l. and has been associated with fever, a palpable purpuric rash, and an eschar in humans [13]. Although human infections have been reported in South America and Europe, in West Africa the organism has only been detected in animal studies with the DNA of R. massiliae having been found in 3.5% of cattle studied in Nigeria [55]. In ticks, R. massiliae has been detected in Rhipicephalus sp. and Rh. senegalensis from Côte d’Ivoire [24,79], in Rh. sanguineus and Rh. turanicus from Nigeria [27,74], and in Rh. guilhoni and Rh. senegalensis from Senegal and Liberia, respectively [21,26] (Table 1).
Finally, R. sibirica mongolitimonae, which has been associated mainly with ticks of the genus Hyalomma, has been reported in humans in Southern Europe but also in travellers returning from North Africa (Algeria and Egypt) [13]. The clinical signs in patients include fever, maculopapular rash, one or more eschars, and regional lymphadenitis and lymphangitis that is characteristic of the disease and leads to it being named “rickettsiosis associated with lymphangitis” [76]. No cases of R. mongolitimonae rickettsiosis have been reported in West Africa in humans or animals. However, the DNA of R. mongolitimonae was detected in Hy. truncatum ticks collected from cattle in Senegal and Mali [21,23] (Table 1).
Two undescribed Rickettsia species have been reported from West Africa, including “Candidatus Rickettsia liberiensis”, genetically related to R. raoultii detected in Ixodes muniensis ticks collected from dogs in Liberia [26], and a Rickettsia species belonging to the Rickettsia rickettsii group in the tick Rh. evertsi in Nigeria [30]. Recently, the DNA of Rickettsia spp. was detected in Am. latum, Am. variegatum, Ixodes aulacodi, Rh. sanguineus, Hy. rufipes, Hy. truncatum, Rh. annulatus, and Rh. microplus collected from snakes, four-toed hedgehogs in Benin, cattle in Benin and Togo [34,36], and in Am. variegatum, Rh. sanguineus s.l., Rhipicephalus sp., Hy. truncatun, Hy. rufipes, Rh. evertsi, and Rhipicephalus (Boophilus) sp. in Ghana [35]. The different Rickettsia species detected in West Africa in humans, animals, and ticks are shown on the map in Figure 2.
2.2. Tick-Borne Relapsing Fevers
The tick-borne relapsing fevers (TBRFs) are acute febrile illnesses characterised by multiple recurrences of fever, headache, myalgia, and arthralgia. They are caused by spirochetes of the genus Borrelia (Borrelia crocidurae, Borrelia duttonii, B. recurrentis, and Borrelia hispanica), which are endemic in subtropical regions around the world [80]. Historically, TBRFs were considered to be transmitted by soft ticks of the genus Ornithodoros [81]. However, in 2011, the paradigm of TBRF transmission by soft ticks changed with the dis-covery in Russia in 1995 of Borrelia miyamotoi, which was reported to cause TBRFs and be transmitted by hard ticks (Ixodes genus); this was subsequently confirmed in Europe, Japan, and the USA [81]. They are a major cause of febrile illness in several regions of Africa [60], and in West Africa TBRF is caused by B. crocidurae transmitted by Ornithodoros sonrai [60]. There are also Borrelia that cause disease in animals—mainly Borrelia theileri causing bovine borreliosis, and other Borrelia of unknown pathogenicity which have also been detected in humans and ticks [82]. Tick-borne relapsing fever can be diagnosed by microscopy (microscopic detection of spirochetes in blood smears), culture, and animal inoculation, but the best and most widely available tools are molecular methods (PCR or qPCR in blood samples) that have been shown to be more sensitive and specific than blood smears during the acute febrile phase [17,81,83].
In West Africa, TBRF has mainly been studied in Senegal [84] where the presence of B. crocidurae has varied widely, depending on the study, date, and location. Between 1989 and 1990, there was only a low prevalence of 0.9% (12/1340) in children under the age of 14 with acute fever presenting at the Keur Moussa dispensary [85], while between 1990 and 2003 the average incidence of TBRF was 11 per 100 person-years [86]. The DNA of B. crocidurae was reported in 5.1% of patients with a fever in Dielmo in 1996 [87], 0.42% of suspected malaria patients examined in Dakar between October 1999 and October 2003 [88], 15% and 0.3% of patients who tested negative for malaria in 2011 in Dielmo and Ndiop, respectively [83], and 9.5% of febrile patients from Dielmo and Ndiop between February 2011 and January 2012 [89]. Further, TBRF has been reported in patients returning from Senegal who developed a fever [90]. Recently B. crocidurae DNA was found in 11.7% and 7.22% of febrile patients in Senegal [57,91]. In Togo, the DNA of B. crocidurae and B. duttonii was detected in 8.8% and 1.2% of febrile patients, respectively [92], while B. crocidurae was found in 3.4% of febrile patients who tested negative for malaria in Mali [93]. Borrelia crocudirae has also been detected by a variety of methods in asymptomatic small mammals in Senegal, Mali, and Mauritania [57,58,62,94], and by demonstrating B. crocidurae in blood smears in 17.6% of 740 rodents and 7.3% of 55 musk shrews in Senegal [95]. In Mali, 11.3% (82/726) of animals (rodents and shrews) were positive for antibodies against relapsing fever spirochetes, while 2.20% (16/724) were positive for spirochetes in blood smears [59]. In ticks, B. crocidurae has been reported in O. sonrai from Mauritania, Senegal, Mali, and Gambia [56,57,58,59,60] (Table 1).
The agent of bovine borreliosis, B. theileri, has been reported in Africa, Australia, and North and South America in cattle, goats, and sheep. In cattle, infections usually manifest as fever and anaemia [66]. In West Africa, the DNA of B. theileri has been detected in Rh. (B.) geigyi collected from cattle in Mali [66] (Table 1).
Other Borrelia species of unknown pathogenicity for humans and animals have been detected in ticks and cattle in West Africa. Borrelia spp. were detected in domestic animals in Ghana [96], in ticks collected from vegetation in Nigeria [30], and in Am. variegatum and Hy. truncatum collected in Mali [23] (Table 1). Three potential new Borrelia species (Candidatus Borrelia Africana, Candidatus Borrelia ivorensis, and Candidatus Borrelia kalaharica) have been detected in West Africa, the first two of which were detected in Am. variegatum from Côte d’Ivoire [24] and the other in Ornithodoros savignyi and livestock from Nigeria [97,98] (Table 1). The different Borrelia species detected in West Africa in humans, animals, and ticks are shown on the map in Figure 3.
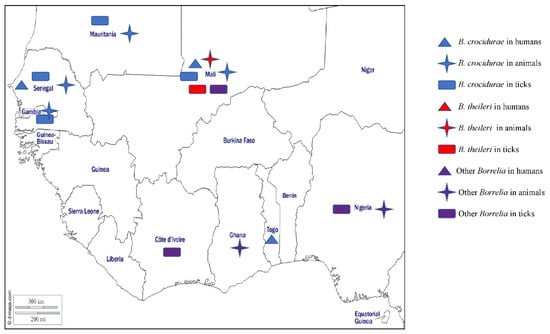
Figure 3.
Different species of Borrelia detected by PCR or microscopy methods in humans, animals, and ticks in West Africa.
2.3. Anaplasmosis
Anaplasmosis is a disease of humans and animals that is caused by Anaplasma species, all obligate intracellular gram-negative bacteria, that are mainly transmitted by ticks (Figure 4) [99,100]. In humans, A. phagocytophilum can be found in circulating neutrophils and is the agent of human granulocytic anaplasmosis (HGA), manifesting as lethargy, inappetence, weight loss, musculoskeletal pain, respiratory insufficiency, and severe gastrointestinal bleeding [99,100]. Thrombocytopaenia and liver enzyme alterations are the most common laboratory abnormality in HGA [99]. In animals, A. phagocytophilum also infects neutrophils and is the agent of tick-borne fever in sheep, pasture fever in cattle, canine granulocytic anaplasmosis, and equine granulocytic anaplasmosis [101]. Signs common to all species include fever, depression, and inappetence, with sheep and cattle also developing leukopaenia and severe secondary infections, horses developing icterus, ventral oedema, and petechiation [101], and dogs developing gastrointestinal and respiratory signs [102]. Diagnosis is based on clinical signs and symptoms, including thrombocytopaenia including leukopaenia and elevated transaminases, by microscopic identification of morulae in neutrophils on a blood smear, or in the buffy coat by serological and molecular methods [17,103]. In West Africa, there have been no reports of HGA, but the DNA of A. phagocytophilum has been detected in one healthy dog from Nigeria [104] and in two febrile sheep from Senegal [105]. Anaplasma phagocytophilum is mainly transmitted by ticks of the genus Ixodes, the tick species of which vary according to geographical area [106]. For example, I. pacificus and I. scapularis in the United States, I. ricinus, I. trianguliceps, I. hexagonus, and I. ventalloi in Europe, and I. persulcatus in Asia and Russia [106]. In addition, DNA of A. phagocytophilum has been detected in other tick species, but their vectorial competence and their role in the epidemiological cycle of this bacterium are still not clear [106]. There have been no reported studies on A. phagocytophilum in ticks in West Africa, despite numerous studies being carried out in this area.
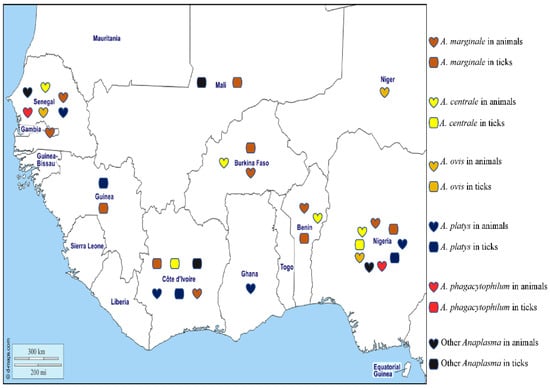
Figure 4.
Different species of Anaplasma detected by PCR in animals and ticks in West Africa.
Anaplasma marginale is found in the erythrocytes of ruminants and is the agent of bovine anaplasmosis. It is mainly transmitted by ticks belonging to the Rhipicephalus and Dermacentor genera [106] and has a wide distribution in tropical and subtropical areas. The distribution of the organism around the world is spreading rapidly, probably due to the transport of cattle from endemic to non-endemic areas and global warming, which favours tick survival [106]. In West Africa, A. marginale has been detected in cattle in various areas of Senegal [107,108,109] by microscopy of blood smears, a technique which has poor sensitivity and specificity. Similarly, A. marginale antigens have been detected in ELISA studies of cattle from Gambia [110]. The DNA of A. marginale has been reported in cattle from Nigeria [55,111,112,113,114], Senegal [68], Ghana [112], Côte d’Ivoire [115], Burkina Faso, and Benin [42,44,112]. In ticks from West Africa, the DNA of A. marginale has been reported in Rh. (B.) decoloratus collected in Nigeria and Burkina Faso [28,44], in Rh.(B.) microplus from Côte d’Ivoire, Mali, Benin, and Guinea [23,24,37,65], in Rh. geigyi from Guinea [65], and in Am. variegatum from Benin [39] (Table 1).
Anaplasma centrale is responsible for clinically benign bovine anaplasmosis and is principally transmitted by the African tick Rh. simus [106]. As the organism confers protective immunity against A. marginale and is less pathogenic, it is used as a live vaccine against A. marginale infections in cattle in many countries [106]. In animals from West Africa, the DNA of A. centrale has been reported in cattle in Nigeria [55,112], Senegal [68], Benin [112], and Burkina Faso [42,112]. In ticks, the DNA of A. centrale was found in Am. variegatum from Côte d’Ivoire and Rh. (B.) annulatus and Hy. impeltatum from Nigeria [24,30] (Table 1).
Anaplasma ovis, responsible for anaplasmosis in sheep, goats, and wild ruminants in tropical and subtropical regions, is an obligate intra-erythrocytic bacterium transmitted by Rhipicephalus spp. [106]. Anaplasma ovis has been reported in Africa, Asia, Europe, and the United States [106]. In West Africa, it has been detected by microscopy in the blood of small ruminants in Senegal [105,107,108,109]. Its DNA has been detected in dogs from Nigeria [104], and in sheep from Senegal and Niger [68,116]. Currently, A. ovis has not been reported in ticks collected in West Africa, despite numerous studies carried out in this area.
Anaplasma platys is found in platelets in dogs worldwide and is the agent of infectious canine cyclic thrombocytopenia (ICCT). It is mainly transmitted by Rh. sanguineus s.l. but can be detected in D. auratus, I. persulcatus, and Ha. longicornis, although these ticks are not known to be involved in its transmission [106]. While A. platys is mainly a canine pathogen, it can occasionally be found in other animals such as cats, foxes, Bactrian camels, deer, sika deer, cattle, and humans [106]. In West Africa, the DNA of A. platys has been detected in dogs, camels, and cattle from Nigeria [55,113,117,118,119], dogs from the Maio Island in the Cape Verde archipelago [120,121], dogs from Ghana and Côte d’Ivoire [71,73,118], and dogs, cattle, goats, and sheep from Senegal [68,105]. In ticks, the DNA of A. platys has been identified in Rh. sanguineus s.l. collected from dogs in Côte d’Ivoire [71], in Rh. microplus collected from cattle in Guinea [65], and in Hy. truncatum collected from cattle in Nigeria [55] (Table 1).
Other unidentified species of Anaplasma have been reported in ticks, including ‘Candidatus Anaplasma ivorensis’, which is similar to A. phagocytophilum and has been identified in Rh. (B.) microplus collected from cattle in Mali [23], and Am. variegatum in Côte d’Ivoire [24]. In O. sonrai collected from Senegal, the DNA of Anaplasma sp. have been detected [58] (Table 1). In Senegal, the DNA of a new potential species of Anaplasma named Candidatus Anaplasma africae, Candidatus (Ca) Anaplasma turritanum, Candidatus (Ca) Anaplasma cinensis, Candidatus (Ca) Anaplasma africanum, and Candidatus (Ca) Anaplasma boleense were detected in cattle, sheep, and goats [68,122], Anaplasma spp. (Badiuoré Ziguinchor) in sheep in Senegal and dogs and cattle in Nigeria [38,55,105], and Candidatus Anaplasma camelii in the blood of camels from Nigeria [49].
Finally, Anaplasma bovis is mainly found in the monocytes of cattle and buffalo, but also in many other domestic and wild animals [106]. Several species of ticks are suspected to be vectors of this bacterium in Africa, including Hyalomma sp., Am. variegatum, Rh. appendiculatus, Rh. sanguineus s.l., and Haemaphysalis spp. [106]. To date, A. bovis has not been reported in West Africa.
The different Anaplasma species detected in West Africa in humans, animals, and ticks are shown on the map in Figure 4.
2.4. Ehrlichiosis
Ehrlichioses are diseases of animals and humans caused by tick-borne obligate intracellular gram-negative bacteria of the genus Ehrlichia. The genus Ehrlichia comprises five species [123], four of which have been reported in West Africa, namely Ehrlichia canis, E. ewingii, E. ruminantium, and E. chaffeensis, along with unidentified Ehrlichia species [82] (Figure 5).
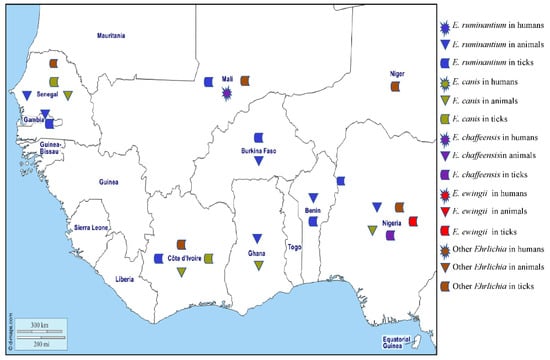
Figure 5.
Different species of Ehrlichia spp. detected by PCR in humans, animals, and ticks in West Africa.
Ehrlichia chaffeensis is found in circulating monocytes and is responsible for human monocytic ehrlichiosis (HME) that can have signs of fever, myalgia, headache, nausea, vomiting, acute renal failure, leukopaenia, thrombocytopaenia, and increased liver enzyme activity. The disease is regarded as an emerging zoonosis in the United States [124], with cases also reported in Europe and West Africa [125]. A person with HME has been described in Mali [126]. Antibodies against E. chaffeensis have been found in an asymptomatic blood donor in Burkina Faso, although serology assay cross reactions cannot be ruled out [127]. In the United States, E. chaffeensis is transmitted by Am. americanum, but DNA of the organism has been found in Hy. impeltatum in Nigeria [30] (Table 1). This study also identified three genotypes of Ehrlichia in Rh. e. evertsi that were phylogenetically close to E. chaffeensis.
Ehrlichia ewingii mainly infects granulocytes and is the aetiologic agent of granulocytic ehrlichiosis in humans and dogs in the United States where it is transmitted by Am. americanum [128]. In West Africa, no human or canine cases of granulocytic ehrlichiosis have been reported to date, but the DNA of E. ewingii has been detected in Rh. (B.) annulatus collected in Nigeria. Also, two other Ehrlichia genotypes that are closely related to E. ewingii were found in Am. variegatum and Hy. impeltatum [30] (Table 1).
Ehrlichia ruminantium, formerly Cowdria ruminantium, is the causative agent of heartwater in wild and domestic ruminants across Africa, where the main vectors are ticks of the genus Amblyomma [129]. Heartwater is a notifiable disease according to the World Organization for Animal Health (OIE) and a serious economic problem for pastoralists in sub-Saharan Africa. The disease occurs in most of sub-Saharan Africa, except the very dry south-west, and is present on the islands of Madagascar, Mauritius, Réunion, Grande Comore, and São Tomé. In the new world, E. ruminantium is present on the islands of Guadeloupe, Antigua, and Marie-Galante in the West Indies [129]. In West Africa, antibodies against E. ruminantium have been detected in small ruminants in Gambia [130], cattle in Côte d’Ivoire [131], and domestic ruminants in Ghana [132,133]. The DNA of E. ruminantium has been detected in lambs in Gambia [134], sheep in Senegal [105], cattle from Burkina Faso, Nigeria, Ghana, and Benin [40,55,112,135], and dogs in Nigeria [38]. The DNA of E. ruminantium has been found in Am. variegatum collected in Mali, Côte d’Ivoire, Burkina Faso, Gambia, Nigeria, and Benin [23,24,38,39,40,41,42]. Although it has also been detected in other species of ticks, including Rh. (B.) microplus collected on cattle in Mali, Côte d’Ivoire, and Burkina Faso [23,24,67], and Hy. truncatum, Hy. rufipes, and Rh. e. evertsi from Mali [23] (Table 1), the possible role, if any, that these ticks play in the epidemiology of E. ruminantium is undetermined.
Ehrlichia canis is transmitted by Rh. sanguineus s.l. and infects dogs worldwide, causing canine monocytic ehrlichiosis. Many infections are subclinical but there can be acute signs of fever, loss of appetite, lethargy, pallor, lymphadenomegaly, splenomegaly, and petechiation. In some chronically infected dogs, there is marked pancytopaenia with anaemia and secondary infections [123]. Antibodies against E. canis have only been reported in West Africa from dogs in Côte d’Ivoire [136,137]. The DNA of E. canis has been found in dogs from Nigeria [38,55,117,118,119,138,139,140,141], Senegal [68,142], Ghana [73], Cape Verde [120,121], and Côte d’Ivoire [143]. In ticks, the DNA of E. canis was detected in Rh. sanguineus s.l. collected from watchdogs in Côte d’Ivoire [72] and Rh. eversti eversti collected from sheep in Senegal [68] (Table 1).
The DNA of potentially new species of Ehrlichia, provisionally named “Candidatus Ehrlichia rustica” and “Candidatus Ehrlichia urmitei”, have been detected in Am. variegatum, Hy. truncatum, and Rh. (B.) microplus collected in Côte d’Ivoire and Mali [23,24]. Candidatus Neoehrlichia mikurensis has been identified in Rh. sanguineus s.l. in Nigeria [141], and Ehrlichia sp. (Omatjenne), which is phylogenetically close to Ehrlichia ruminantium, has been found in Hy. truncatum collected from cattle in Nigeria [55]. Two genotypes of Ehrlichia sp. (Erm58 and Eht224) from the E. canis group have been identified in Rh. muhsamae and Hy. truncatum collected in Mali and Niger, respectively [55] (Table 1). Recently, a potentially new species of Ehrlichia, Candidatus Ehrlichia senegalensis, and Ehrlichia sp., has been detected in rodents and O. sonrai, respectively, collected in Senegal [58,62]. The different Ehrlichia species detected in West Africa in humans, animals, and ticks are shown on the map in Figure 5.
2.5. Bartonellosis
The bartonelloses are caused by Bartonella species, which are fastidious haemotropic gram-negative organisms that can be transmitted by arthropods to people and a wide range of domestic and wild animals which can act as reservoir hosts [96]. At least 20 species of Bartonella are known to be responsible for specific intra-erythrocyte infections in their hosts [144]. Bartonella infection is diagnosed by serological, microbiological culture, and/or PCR methods [145]. In West Africa, several species of Bartonella have been reported in humans, animals, and arthropods (Figure 6).
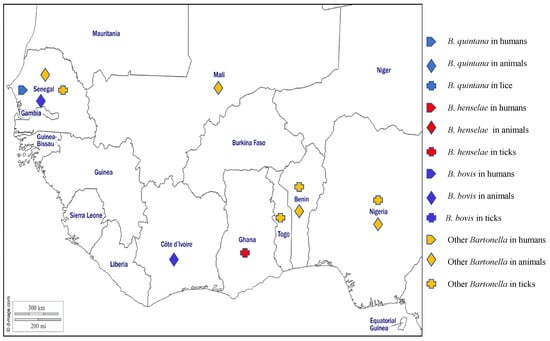
Figure 6.
Different species of Bartonella spp. detected by PCR in humans, animals, and ectoparasites in West Africa.
Bartonella henselae most commonly infects domestic and wild cats but can accidentally infect humans. It is the main causative agent of cat scratch disease, a generally benign disease of children and young adults. In immunocompromised individuals, however, B. henselae can cause life-threatening disorders including bacteraemia and bacillary angiomatosis [144]. Further, it is the second most common Bartonella species causing endocarditis, with cases having been reported around the world [144]. In West Africa, no cases of B. henselae in humans have been reported, but antibodies against the organism have been reported in the serum of cat from Ghana [146]. Around the world, B. henselae is mostly transmitted by cat fleas, although the DNA of B. henselae has also been found in ticks (I. pacificus, I. ricinus, and I. affinis) [144]. No flea studies have been reported from West Africa, but the DNA of B. henselae has been found in Rh. sanguineus s.l. collected from dogs, in cats’ blood, and ticks collected from cats in Ghana [73,147].
Bartonella quintana, the agent of trench fever, is transmitted by human body lice, although DNA of the organism was found in Rh. sanguineus s.l. [148]. The DNA of B. quintana has been detected by PCR in blood from febrile patients in rural areas in Senegal [89,149] and antibodies against B. quintana were found in the sera of asymptomatic humans who had close contact with fruit bats in Ghana [146].
Bartonella bovis is the aetiological agent of bovine bartonellosis, a mostly asymptomatic disease which can occasionally present as endocarditis with anorexia, weight loss, wasting, and abnormal cardiac auscultation [144]. The organism is thought to be transmitted by arthropods such as fleas, flies, lice, mites, and ticks that are found to be naturally infected [150]. Reported in Europe, USA, Asia, and Africa, the prevalence of B. bovis in cattle is generally high but varies widely across studies and countries [150]. In West Africa, B. bovis has only been reported in cattle from Côte d’Ivoire and Senegal [151,152].
Several Bartonella spp. have been found in small mammals in West Africa, including a Bartonella spp. closely related to Bartonella elizabethae, which was amplified from rodents from Benin [34,153], Nigeria [154], and recently found in Mali [155], a Bartonella spp. close to Bartonella rochalimae in Benin [153], a Bartonella spp. close to B. tribocorum in Benin and Nigeria [153,154], a Bartonella spp. Similar to B. grahamii in Nigeria [154], unidentified Bartonella spp. in rodents, Am. varieagatum, Am. latum, Hy. rufipes, I. aulacodi, Rh. muhsamae, Rh. sulcatus, and Rh. microplus from Benin and Togo [34,36,153], in rodents, bats, and their ectoparasites from Nigeria [154,156], and rodents from Mali [155]. In addition, two potentially new species of Bartonella have been described, namely Candidatus Bartonella davoustii in cattle in Senegal [151] and Candidatus Bartonella mastomydis in rodents in Benin [153] and Mali [155]. Finally, three potentially new genotypes have been identified in rodents from Senegal [62], and B. senegalensis and B. massiliensis, two newly recognised species, have been reported in O. sonrai collected in Senegal [61]. The different Bartonella species detected in West Africa in humans, animals, and ticks are shown on the map in Figure 6.
2.6. Coxiella burnetii Infection
Coxiella burnetii is a small gram-negative intracellular bacterium which is highly resistant to the environment and the causative agent of Q fever. This zoonosis is mainly transmitted to humans by aerosols generated from infected placentas and birth liquids [157]. The organism has been found in many tick species, suggesting these arthropods might also play a role in the transmission of the bacterium [157]. Q fever has been reported almost everywhere it has been investigated, with the exception of New Zealand. Most primary humain infections are asymptomatic, but there can be fever, malaise, headache, fatigue, and pneumonia hepatitis, resulting in acute Q fever [157]. Serious obstetric complications and foetal malformations may occur in pregnant women. About 5% of infections become persistent with endocarditis, lymphadenitis, and vascular and osteoarticular infections developing many years later [157]. In animals, C. burnetii infections are also mostly asymptomatic but there might be abortions and stillbirths in cats and domestic ruminants which are a source of infection for people. Coxiella burnetii infection can be diagnosed by several methods, including culture (only performed by reference laboratories), serology (the most commonly used method), pathology–immunohistochemistry, and PCR [157].
In West Africa, Q fever has been reported in a man returning from Guinea Bissau with acute lobar pneumonia and fever, headache, haematuria, and hepatitis [158]. It has also been reported in tourists who had stayed in Gambia, Côte d’Ivoire, and Burkina Faso [18]. Antibodies reactive to C. burnetii have been reported in Nigerian patients hospitalised for various acute medical conditions [159], apparently healthy adults from Mali, Burkina Faso, and Côte d’Ivoire [77], children aged between one month and five years in Niger [160], children and adults from Ghana [161], nomads in rural areas of northern Burkina Faso [162], in northern Togo [163], blood donors and patients in Mauritania [19], and in adults and children in Gambia [164,165]. Coxiella burnetii DNA was detected in febrile patients who tested negative for malaria with Paracheck® in Mali [46] and in human and various environmental samples from Senegal [43,69,89].
In animals from West Africa, antibodies against C. burnetii have been detected in sera and milk from cows and in sheep in Nigeria [166,167] as well as in sera from dogs in Senegal and Côte d’Ivoire [168], livestock in the Republic of Guinea [20], cattle, sheep, and goats in Togo, Côte d’Ivoire, Mali, and Ghana [163,169,170,171], small ruminants (goats and sheep) in Gambia [164,172], and rodents in Cape Verde [173]. The DNA of C. burnetii has been detected in milk offered for consumption and rodents in Senegal [174,175], rodents in Nigeria [70], and recently in rodents in Mali [155].
The DNA of C. burnetii has been found in several species of ticks, although no specific tick vector has been found to play a role in the epizootic cycle of C. burnetii and the presence of DNA in a tick does not indicate it plays a role in transmission. In Senegal, DNA of C. burnetii was detected in Am. variegatum, Rh. (B.) annulatus, Rh. (B.) decoloratus, Rh. e. evertsi, Rh. guilhoni, Hy. rufipes, Hy. truncatun, and O. sonrai [43] (Table 1). It has also been identified in Am. variegatum, Rh. (B.) annulatus, Rh. e. evertsi, Hy. truncatun, Hy. dromedarii, and Hy. impeltatum collected from cattle in Nigeria and Am. variegatum collected from cattle in Côte d’Ivoire [24,30,49] (Table 1). Recently, C. burnetii has been identified in Am. variegatum, Rh. (B.) microplus, Rh. sanguineus s.l., Rh. e. evertsi, Hy. rufipes, and Hy. truncatun collected from cattle, Haemaphysalis spp. from rodents and head lice from patients in Mali [23,155,176], in Am. variegatum, Rh. sanguineus s.l., Rhipicephalus sp., Hy. truncatun, and Hy. rufipes collected from cattle in Ghana [35], and in unspecified ticks collected from cats in Ghana [147] (Table 1).
3. Protozoal Diseases
3.1. Theileriosis
Theileriosis is a tropical disease caused by obligate intracellular haemoprotozoa of Theileria genus that infect both leukocytes and erythrocytes and are transmitted by hard tick genera, mainly Amblyomma, Haemaphysalis, Hyalomma, and Rhipicephalus [177]. Wild and domestic ruminants are the main hosts in which infections cause enlargement of the lymph nodes, fever, anaemia, jaundice, leukopaenia, pulmonary oedema, lethargy, thrombocytopenia, and death [178]. Differential diagnosis of theileriosis requires observation of clinical signs, the epidemiological context, and the use of pathological, microscopic, and molecular techniques to detect the parasite or its DNA [179]. A large number of species have been reported in domestic animals around the world, including Theileria parva, T. annulata, T. taurotragi, T. lestoquardi, T. orientalis, T. velifera, T. mutans, T. uilenbergi, T. lowenshuni, T. sinensis, T. ovis, and T. equi [178]. To date, the species of Theileria reported from West Africa are T. annulata, T. mutans, T. velifera, T. taurotragi, T. equi, and Theileria sp. (Figure 7).
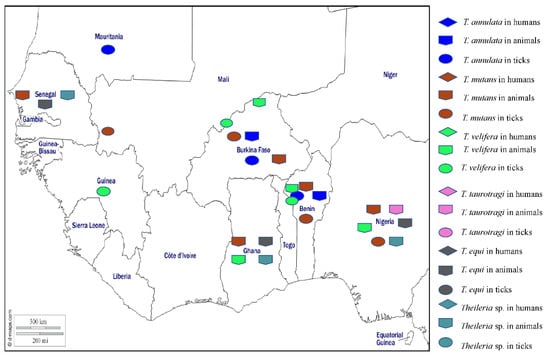
Figure 7.
Different species of Theileria detected by PCR in humans, animals, and ticks in West Africa.
Theileria annulata, the agent of tropical theileriosis manifested by fever and lymphadenomegaly, is transmitted by several species of Hyalomma and is present around the Mediterranean basin, northeast Africa, the Middle East, India, and South Asia. In West Africa, the DNA of T. annulata has been detected in cattle from Burkina Faso and Benin [42,44]. In ticks, the DNA of T. annulata was found in Am. variegatum, Hy. rufipes, Rh. decoloratus, Rh. geigyi, and Rh. microplus from Burkina Faso and Benin [42,44], and in Hy. dromadarii and Hy. rufipes from Mauritania [50] (Table 1).
Theileria mutans infects buffalo and cattle in Africa and is transmitted by Amblyomma spp. and infections are mostly benign and only very rarely associated with serious clinical illness. In West Africa, T. mutans has been detected by microscopy in blood smears from asymptomatic cattle from Senegal [107,108,109], Nigeria [55], and Ghana [96]. The DNA of T. mutans has been found in cattle blood from Burkina Faso and Benin [42,44]. In ticks, the DNA of T. mutans was detected in Hy. impeltatum and Rh. annulatus collected from cattle in Nigeria [30] and in Am. variegatum, Hy. rufipes, Rh. microplus, Rh. geigy, and Rh. decoloratus collected from cattle in Burkina Faso and Benin [39,42,44] (Table 1).
Theileria velifera infections in buffalo and cattle in Africa are benign and transmitted by Amblyomma spp. [178]. In West Africa, T. velifera has been detected in cattle from Nigeria, Ghana, Burkina Faso, and Benin [42,44,55,96]. In ticks, the DNA of T. velifera has been detected in Am. variegatum, Hy. rufipes, Rh. decoloratus, Rh. annulatus, and Rh. geigyi collected from cattle in Burkina, Benin, and Guinea [42,44,45] (Table 1).
Theileria taurotragi is a parasite that exists in African eland and cattle and is transmitted by Rhipicephalus spp. and infections are benign [178]. In West Africa, T. taurotragi has been reported in cattle from Nigeria [55] (Table 1).
Theileria equi infects horses, donkeys, and zebras in tropical and subtropical areas, causing equine piroplasmosis which can present as fever, anaemia, inappetence, oedema, icterus, haepatomegaly, splenomegaly, and death in some cases. The disease can cause serious economic losses in the equine industry and is transmitted by several tick genera including Hyalomma, Dermacentor, Rhipicephalus, and Amblyomma [178]. In West Africa, antibodies against T equi have been detected in horses and donkeys from Nigeria [180,181]. Theileria equi DNA has been detected in horses, donkeys, and asymptomatic dogs from Nigeria [38,51,182,183,184], and in horses from Ghana [185] and Senegal [186]. In ticks, T. equi DNA was detected in Rh. e. evertsi, Hy. dromedarii, and Hy. truncatum collected from horses and donkeys in Nigeria [51].
In addition, Theileria sp. (sable) and a Theileria sp. close to T. ovis have been identified in Nigerian dogs [38,141]. A Theileria sp. has been found in sheep and goats from Ghana [96]. In Senegal, a potentially new species of Theileria, provisionally named Theileria sp. “Africa”, has been found in a horse [186], and Candidatus Theileria senegalensis has been detected in a rodent [62]. The different Theileria species detected in West Africa in humans, animals, and ticks are shown on the map in Figure 7.
3.2. Babesiosis
Worldwide, there are more than 100 species of Babesia which are usually transmitted by hard ticks and infect the erythrocytes of a wide variety of domestic and wild animals. In Africa, animal babesiosis is most commonly due to B. bigemina and B. bovis in cattle, B. gibsoni and B. canis in dogs, and B. caballi in horses and donkeys [187]. Clinical signs result from haemolysis and include fever, depression, anaemia, splenomegaly, and jaundice. Only a few species infect humans [188], including Babesia microti, which mainly infects rodents and is transmitted by Ixodes in the northern hemisphere [189], and is the main causative agent of human babesiosis in the United States [189]. Babesia divergens is the major cause of human babesiosis in Europe and is transmitted by Ixodes ricinus, the only known vector [190]. Human babesiosis caused by B. microti and B. divergens is characterised by a fever that usually resolves spontaneously in immunocompetent patients. Sometimes there is haematuria and jaundice, depending on the degree of haemolysis [188]. Severe complications of babesiosis, such as splenic infarction and rupture, are more frequently observed in young, immunocompetent male patients [191,192,193]. Babesiosis can be diagnosed by observing abnormalities such as haemolytic anaemia, thrombocytopaenia, elevated transaminases, or by microscopic observation of the parasite on blood smears or detection of the DNA by PCR [17]. In West African humans, a single probable case of human babesiosis has been reported in an infant in Côte d’Ivoire [194]. Several species of Babesia have been reported in animals and ticks in West Africa (Figure 8). Babesia microti has been found in house rats collected in Nigeria [195] and B. divergens DNA has been detected in Am. variegatum ticks in Nigeria [28] (Table 1).
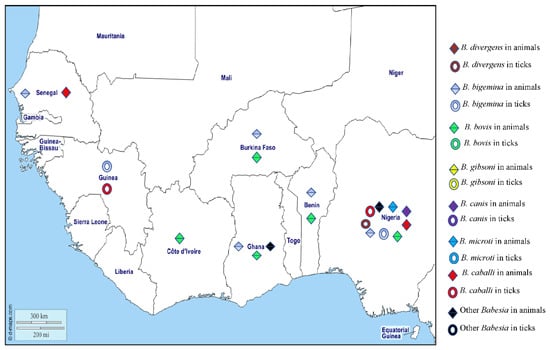
Figure 8.
Different species of Babasia spp. detected by PCR or microscopy in animals and ticks in West Africa.
Babesia bigemina is a parasite of cattle that has been reported on all continents, it is transmitted principally by Rh. (B.) microplus and Rh. (B.) annulatus [196]. In West Africa animals, B. bigemina has been detected by microscopy in bovine blood smears in Senegal [107,108,109] and Ghana [96]. The antibodies against B. bigemina have been found in cattle in Gambia [197] and Mali [198], and the DNA of B. bigemina has been detected in cattle in Nigeria, Burkina Faso, Ghana, Côte d’Ivoire, and Benin [55,112,115]. In ticks, the DNA of B. bigemina has been detected in Rh. (B.) decoloratus and Am. variegatum from Nigeria [28,64], and Rh. (B.) microplus from Guinea [65] (Table 1).
Babesia bovis, the agent of redwater fever in African cattle, is widespread in tropical and subtropical regions around the world and is transmitted by ticks, principally Rh. (B.) microplus and Rh. (B.) annulatus. In West Africa, antibodies against B. bovis have been found in cattle in Mali, Nigeria [199,200], and Côte d’Ivoire [201]. Studies have also reported the presence of B. bovis DNA in cattle from Nigeria, Burkina Faso, Côte d’Ivoire, Benin, and Ghana [42,44,55,112,115,202].
Babesia gibsoni mainly infects dogs in Asia, Europe, America, and North and East Africa, where it is probably transmitted by Rh. sanguineus s.l. [187]. Babesia gibsoni has only been detected in dogs from Cape Verde in West Africa [203].
There are three subspecies of Babesia canis, namely canis, vogeli, and rossi. The first two infect dogs around the world and are transmitted mainly by Rh. sanguineus s.l. and D. reticulatus. Babesia canis rossi infects dogs mainly in Africa, where it is transmitted by Haemaphysalis leachii [187] and causes a more severe disease than the two other subspecies. In West Africa, all three subspecies of B. canis have been detected in dogs from Nigeria [139,204,205,206].
Babesia caballi is also an agent of equine piroplasmosis (see theileriosis above) which occurs in most countries of the world where the competent Dermacentor, Rhipicephalus, and Hyalomma vectors are found [207]. The DNA of B. caballi has been detected in horses from Senegal [186] and Nigeria [183,184] and in Am. variegatum and Rh. decoloratus collected from cattle in Guinea [45,46] and Hyalomma spp. from Nigeria [49].
Babesia perroncitoi and B. trautmanni are responsible for swine babesiosis and have been reported in pigs in Nigeria and Ghana [208,209]. The different Babesia species detected in West Africa in humans, animals, and ticks are shown on the map in Figure 8.
3.3. Hepatozoonosis
Although there are hundreds of Hepatozoon species with a very wide variety of hosts and vectors [210], there are no human strains and only two species are of veterinary importance. Both species infect dogs, with Hepatozoon canis occurring around the world and Hepatozoon americanum infecting dogs in the United States [211]. Hepatozoon canis is transmitted by Rh. sanguineus s.l., with infections resulting in dogs being asymptomatic or developing serious and sometimes fatal signs including fever, lethargy, anaemia, cachexia, weight loss, and hind limb weakness [211]. In West Africa, H. canis DNA has been reported in dogs in Nigeria [117,118,140,212], Cape Verde [120,121,203], and Ghana [73,118]. Recently, H. canis and two Hepatozoon spp., closely related to Hepatozoon spp. from snakes in the north of Africa have been detected in rodents from Senegal [62].
In ticks, the DNA of H. canis was detected in Nigeria and Ghana [118,140].
4. Viral Diseases
4.1. Crimean–Congo Haemorrhagic Fever (CCHF)
Crimean-Congo hemorrhagic fever (CCHF) is a zoonotic hemorrhagic disease with a high mortality rate in humans, caused by a virus of the Bunyaviridae family. While CCHF occurs worldwide, it is considered endemic in certain countries in Asia, Europe, and Africa. The virus is transmitted to humans either by ticks (especially of the Hyalomma genus) or by contact with the blood or secretions of infected animals [213]. The disease manifests as fever, chills, headache, dizziness, neck pain, nuchal rigidity, photophobia, retro-orbital pain, myalgia, arthralgia, nausea, vomiting, diarrhoea, and abdominal pain. It is associated with coagulopathies manifested by petechia, bruising, haematemesis, and melena, often associated with thrombocytopaenia and leukopaenia [213]. The diagnosis of CCHF is generally made by culture (virus isolation), serology (search for specific IgG and IgM antibodies), or molecular tools (detection of virus RNA) [214].
In West Africa, the first human case of CCHF was reported in 1983 in a febrile patient in southern Mauritania [215]. The CCHF virus has been isolated from a deceased patient, and antibodies against the virus have been found in hospitalised patients with signs of haemorrhagic fever in southwestern Mauritania [216]. Between February and August 2003, 38 people were diagnosed with the CCHF virus in Mauritania, 35 of whom resided in Nouakchott [217], and recently two human cases were diagnosed by ELISA and real-time reverse transcription PCR in Mauritania [218]. Anti-CCHF antibodies have been reported in febrile patients from Nigeria [219,220], a febrile patient with conjunctival jaundice, bleeding gums, and haematemesis from Senegal [221], people working in a slaughterhouse in Ghana [47], and Malian patients who had tested negative for Plasmodium falciparum and yellow fever, but who had a history of fever, and haemorrhagic, diarrhoeal, or icteric syndromes [222]. One imported case has been reported, in a woman returning to France from Senegal [223]. The CCHF virus RNA has been detected in humans in Nigeria and Mali [219,224].
In animals, antibodies against the CCHF virus have been detected in cattle from northern Nigeria [225], cattle, rodents, sheep, and goats from Mauritania [53,216,217,226], birds from Senegal [54], and cattle from Mali [227].
In ticks, three different genotypes of the CCHF virus have been identified in Hy. rufipes, Am. variegatum, Rh. guilhoni, and Rh. e. evertsi collected from cattle and goats in Senegal [48] (Table 1). The virus has been isolated from the immature stages of Hy. rufipes collected from a hornbill in Senegal [48]. It has also been detected in Hy. rufipes collected from camels and cattle in Mauritania [53], Hyalomma collected from cattle in Mali [228], and recently in Hy. excavatum and Am. variegatum collected from cattle in Ghana [47] (Table 1). Hyalomma spp. ticks are considered to be the only known vectors of the CCHF virus, so the discovery of virus RNA in Amblyomma spp. and Rhipicephalus spp. requires further investigation before these ticks can be considered as vectors.
4.2. African Swine Virus
African swine fever (ASF) is caused by the African swine fever virus (ASFV), which is the only member of the Asfarviridae family. ASF is the only DNA virus transmitted by arthropods, notably soft ticks of Ornithodoros genus [229]. In Africa and Europe, Ornithodoros moubata and O. erraticus are involved in the sylvatic transmission cycle [229]. First discovered in Kenya in 1921, ASF initially affected sub-Saharan African countries. In 2007, it was introduced from East Africa, then spread widely in Europe, and in 2018, the virus was introduced into China via Russia [229]. ASF can manifest itself clinically in domestic pigs in several forms, including perinatal, acute, subacute, or chronic forms. Clinical signs vary according to the form of the disease, and are characterised by high fever (i.e., a body temperature of 41–42 °C), loss of appetite, inactivity, dyspnoea and skin hyperaemia, lethargy, anorexia, inactivity, respiratory distress and severe pulmonary oedema, and sometimes sudden death can also be observed without signs of disease [229]. Diagnosis of ASF is based on culture (virus isolation on porcine macrophages), serology (enzyme-linked immunosorbent assays (ELISA), immunoblotting, and indirect immunostaining techniques), or qPCR (detection of virus DNA) [230].
In West Africa, there is little published data on ASF except in Senegal and Nigeria [231]. ASF was first reported in Senegal in 1957 and the disease is known to be endemic, with over 54 outbreaks reported since 1986 [231], then spread to many West African countries, including Côte d’Ivoire in 1996, Togo in 1998, Ghana in 2000, Burkina Faso in 2003, Niger in 2008, Benin in 2009, Liberia 2010, Gambia 2011, and Mali in 2016 [231,232]. In animals, antibodies against ASF have been detected in pigs in Senegal [233,234], Benin [235], Burkina Faso [235], Ghana [235], and Nigeria [236,237]. The DNA of ASFV was detected in pigs from Benin [235,238], Burkina Faso [235,239,240], Côte d’Ivoire [241], Ghana [235,242], Mali [239], Nigeria [235,243,244,245,246,247,248], Senegal [239], and in Togo [235]. In Nigeria, the DNA of ASFV was found in red river hogs (Potamochoerus porcus) [249].
In ticks, the DNA of ASFV has been found only in O. sonrai collected from pigsties and warthog burrows in Senegal [63].
5. Tick-Borne Diseases in Children and Pregnant Women
The incidence of tick-borne diseases in children has been studied little or not at all in many West African countries. In Senegal, studies have reported that the incidence of tick-borne relapsing fever in children can vary from 0.5% to 4.2% in children under 4 and over 10 years old, respectively [87], and 56% in children under 10 years old [83]. In Togo, TBRF DNA has also been found in children aged between 1 and 14 [92]. No studies on tick-borne diseases have been carried out among pregnant women in West Africa. In Tanzania, an East African country, it has been reported that TBRF during pregnancy can lead to the need for early and effective prevention and management of TBR [250]. However, as in many West African countries, access to health services remains a serious problem, especially for rural populations who are most exposed to tick-borne diseases. In most cases, treatment of tick-borne bacterial diseases is based on antibiotics such as doxycycline, clarithromycin, azithromycin, cefuroxime, and azithromycin [13,17]. Pregnant women and children represent a special population, and treatment of tick-borne diseases must follow specific recommendations. For example, doxycycline is not recommended for the treatment of Lyme disease in pregnant women and children under the age of eight, due to the availability of alternatives such as amoxicillin or cefuroxime. However, in cases of ehrlichiosis and anaplasmosis, doxycycline can be used in pregnant women and children, as the benefit of doxycycline is greater than the risk in these patients [17].
6. Conclusions
Tick-borne diseases are generally ignored, neglected and underestimated in West Africa. In febrile autochthonous patients or travellers returning from West Africa, the most commonly reported diseases are African tick-bite fever, tick-borne relapsing fever, Q fever and Crimean–Congo haemorrhagic fever. However, cases of co-infection between different tick-borne micro-organisms have not been described in West Africa as they have been in Europe [251]. Human co-infections can manifest themselves as seropositivity without clinical symptoms or co-disease, i.e., the simultaneous clinical expression of infections by two tick-borne micro-organisms. In most cases, co-infection has no impact on the severity of the disease [251]. However, antibodies against a far broader variety of tick-borne pathogens have been found in healthy blood donors in West Africa and many other tick-borne pathogens can be detected in wild and domestic animals and their ticks. Health professionals working with patients returning from West Africa should be aware they might have been infected with a broad range of tick-borne pathogens. Many tick-borne diseases have non-specific signs, and it is only with careful history taking on travel and possible tick-bites that health workers can be alerted to the possibility of tick-borne diseases in their patients. Diagnosis of tick-borne diseases will be carried out using the simplest techniques, and without doubt the most useful for the future will be molecular biology (qPCR and standard PCR). Ticks are, therefore, of major epidemiological interest for surveillance in areas where there are no specialised diagnostic laboratories testing for infections in humans and animals. Matrix-assisted desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS), which has recently revolutionised tick identification, could be a technique to facilitate the identification of African ticks, considering the low number of tick morphology identification specialists on this continent.
Author Contributions
A.Z.D. wrote the initial draft of the manuscript; P.P., B.D. and P.K. added their contributions and comments. All authors have read and agreed to the published version of the manuscript.
Funding
This work was conducted with the support of the A*MIDEX project (no. ANR-10-IAHU-03), funded by the French government’s “Investissements d’Avenir” programme, managed by the French National Research Agency (ANR).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank the Vecteurs Infections Tropicale et Méditerranéennes unit, the IHU Méditerranée-Infection of Marseille for library resources and support during the writing of this review, and Jean-Michel Berenger for providing the tick pictures.
Conflicts of Interest
The authors declare that they have no competing interest.
Abbreviations
| ATBF | African tick-bite fever |
| ASFV | African swine fever virus |
| ASF | African swine fever |
| CCHF | Crimean–Congo haemorrhagic fever |
| DNA | Deoxyribonucleic acid |
| EU | European Union |
| HGA | Human granulocytic anaplasmosis |
| HME | Human monocytic ehrlichiosis |
| MALDI-TOF MS | Matrix-assisted desorption/ionization time-of-flight mass spectrometry |
| ICCT | Infectious canine cyclic thrombocytopaenia |
| MSF | Mediterranean spotted fever |
| PCR | Polymerase chain reaction |
| RNA | Ribonucleic Acid |
| SFG | Spotted fever group |
| TBD | Tick-borne diseases |
| TBRF | Tick-borne relapsing fevers |
| TG | Typhus group |
| WOAH | World Organization for Animal Health |
References
- Walker, A.R.; Bouattour, A.; Camicas, J.L.; Estrada-Peña, A.; Horak, I.G.; Latif, A.A.; Pegram, R.G.; Preston, P.M. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; The University of Edinburgh: Edinburgh, UK, 2003; pp. 52–228. [Google Scholar]
- Kernif, T.; Leulmi, H.; Raoult, D.; Parola, P. Emerging tick-borne bacterial pathogens. Microbiol. Spectr. 2016, 4, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Onyiche, T.E.; MacLeod, E.T. Hard ticks (Acari: Ixodidae) and tick-borne diseases of sheep and goats in Africa: A review. Ticks Tick Borne Dis. 2023, 14, 102232. [Google Scholar] [CrossRef]
- Parola, P.; Raoult, D. Ticks and tickborne bacterial diseases in humans: An emerging infectious threat. Clin. Infect. Dis. 2001, 32, 897–928. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, J.; Estrada-Pena, A.; Venzal, J.M.; Kocan, K.M.; Sonenshine, D.E. Overview: Ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 2008, 13, 6938–6946. [Google Scholar] [CrossRef]
- Ghosh, S.; Azhahianambi, P.; Yadav, M.P. Upcoming and future strategies of tick control: A review. J. Vector Borne Dis. 2007, 44, 79–89. [Google Scholar]
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, S3–S14. [Google Scholar] [CrossRef]
- Kivaria, F.M. Climate change and the epidemiology of tick-borne diseases of cattle in Africa. Vet. J. 2010, 184, 7–8. [Google Scholar] [CrossRef]
- Gray, J.S.; Dautel, H.; Estrada-Pena, A.; Kahl, O.; Lindgren, E. Effects of climate change on ticks and tick-borne diseases in europe. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 593232. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, L. The Impacts of Climate Change on Ticks and Tick-Borne Disease Risk. Annu. Rev. Entomol. 2021, 66, 373–388. [Google Scholar] [CrossRef]
- Communauté économique des états de l’Afrique de l’ouest. Available online: https://fr.wikipedia.org/wiki/Afrique_de_l%27Ouest (accessed on 16 March 2021).
- Mackenzie, J.S.; Jeggo, M. The one health approach-why is it so important? Trop. Med. Infect. Dis. 2019, 4, 88. [Google Scholar] [CrossRef]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef]
- Abdad, M.Y.; Abou, A.R.; Fournier, P.E.; Stenos, J.; Vasoo, S. A concise review of the épidemiology and diagnostics of rickettsioses: Rickettsia and Orientia spp. J. Clin. Microbiol. 2018, 56, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Bermudez, C.S.E.; Troyo, A. A review of the genus Rickettsia in central America. Res. Rep. Trop. Med. 2018, 9, 103–112. [Google Scholar]
- Dieme, C.; Bechah, Y.; Socolovschi, C.; Audoly, G.; Berenger, J.M.; Faye, O.; Raoult, D.; Parola, P. Transmission potential of Rickettsia felis infection by Anopheles gambiae mosquitoes. Proc. Natl. Acad. Sci. USA 2015, 112, 8088–8093. [Google Scholar] [CrossRef]
- Pace, E.J.; O’Reilly, M. Tickborne Diseases: Diagnosis and management. Am. Fam. Physician 2020, 101, 530–540. [Google Scholar] [PubMed]
- Delord, M.; Socolovschi, C.; Parola, P. Rickettsioses and Q fever in travelers (2004–2013). Travel Med. Infect. Dis. 2014, 12, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Niang, M.; Parola, P.; Tissot-Dupont, H.; Baidi, L.; Brouqui, P.; Raoult, D. Prevalence of antibodies to Rickettsia conorii, Ricketsia africae, Rickettsia typhi and Coxiella burnetii in Mauritania. Eur. J. Epidemiol. 1998, 14, 817–818. [Google Scholar] [CrossRef]
- Kalivogi, S.; Buaro, M.E.; Konstantinov, O.K.; Plotnikova, L.F. The immune structure against q fever and tick-bite spotted fever group rickettsioses in the population and domestic animals of the Republic of Guinea. Med. Parazitol. 2013, 1, 28–30. [Google Scholar]
- Mediannikov, O.; Diatta, G.; Fenollar, F.; Sokhna, C.; Trape, J.F.; Raoult, D. Tick-borne rickettsioses, neglected emerging diseases in rural Senegal. PLoS Negl. Trop. Dis. 2010, 4, e821. [Google Scholar] [CrossRef]
- Adjou Moumouni, P.F.; Terkawi, M.A.; Jirapattharasate, C.; Cao, S.; Liu, M.; Nakao, R.; Umemiya-Shirafuji, R.; Yokoyama, N.; Sugimoto, C.; Fujisaki, K.; et al. Molecular detection of spotted fever group rickettsiae in Amblyomma variegatum ticks from Benin. Ticks Tick Borne Dis. 2016, 7, 828–833. [Google Scholar] [CrossRef]
- Diarra, A.Z.; Almeras, L.; Laroche, M.; Berenger, J.M.; Kone, A.K.; Bocoum, Z.; Dabo, A.; Doumbo, O.; Raoult, D.; Parola, P. Molecular and MALDI-TOF identification of ticks and tick-associated bacteria in Mali. PLoS Negl. Trop. Dis. 2017, 11, e0005762. [Google Scholar] [CrossRef]
- Ehounoud, C.B.; Yao, K.P.; Dahmani, M.; Achi, Y.L.; Amanzougaghene, N.; Kacou, N.A.; N’Guessan, J.D.; Raoult, D.; Fenollar, F.; Mediannikov, O. Multiple pathogens including potential New species in tick vectors in Cote d’Ivoire. PLoS Negl. Trop. Dis. 2016, 10, e0004367. [Google Scholar] [CrossRef]
- Lorusso, V.; Gruszka, K.A.; Majekodunmi, A.; Igweh, A.; Welburn, S.C.; Picozzi, K. Rickettsia africae in Amblyomma variegatum ticks, Uganda and Nigeria. Emerg. Infect. Dis. 2013, 19, 1705–1707. [Google Scholar] [CrossRef]
- Mediannikov, O.; Diatta, G.; Zolia, Y.; Balde, M.C.; Kohar, H.; Trape, J.F.; Raoult, D. Tick-borne rickettsiae in Guinea and Liberia. Ticks Tick Borne Dis. 2012, 3, 43–48. [Google Scholar] [CrossRef]
- Nnabuife, H.E.; Matur, B.; Ogo, N.I.; Goselle, O.; Shittu, I.; Mkpuma, N.; Obishakin, E.; Chima, N.; Kamani, J. Rickettsia africae and Rickettsia massiliae in ixodid ticks infesting small ruminants in agro-pastoral settlements in Plateau State, Nigeria. Exp. Appl. Acarol. 2023, 89, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Ogo, N.I.; de Mera, I.G.; Galindo, R.C.; Okubanjo, O.O.; Inuwa, H.M.; Agbede, R.I.; Torina, A.; Alongi, A.; Vicente, J.; Gortázar, C.; et al. Molecular identification of tick-borne pathogens in Nigerian ticks. Vet. Parasitol. 2012, 187, 572–577. [Google Scholar] [CrossRef]
- Parola, P.; Inokuma, H.; Camicas, J.L.; Brouqui, P.; Raoult, D. Detection and identification of spotted fever group Rickettsiae and Ehrlichiae in African ticks. Emerg. Infect. Dis. 2001, 7, 1014–1017. [Google Scholar] [CrossRef] [PubMed]
- Reye, A.L.; Arinola, O.G.; Hubschen, J.M.; Muller, C.P. Pathogen prevalence in ticks collected from the vegetation and livestock in Nigeria. Appl. Environ. Microbiol. 2012, 78, 2562–2568. [Google Scholar] [CrossRef] [PubMed]
- Sambou, M.; Faye, N.; Bassene, H.; Diatta, G.; Raoult, D.; Mediannikov, O. Identification of rickettsial pathogens in ixodid ticks in northern Senegal. Ticks Tick Borne Dis. 2014, 5, 552–556. [Google Scholar] [CrossRef]
- Socolovschi, C.; Huynh, T.P.; Davoust, B.; Gomez, J.; Raoult, D.; Parola, P. Transovarial and trans-stadial transmission of Rickettsiae africae in Amblyomma variegatum ticks. Clin. Microbiol. Infect. 2009, 15, 317–318. [Google Scholar] [CrossRef] [PubMed]
- Tomassone, L.; De, M.D.; Adakal, H.; Rodighiero, P.; Pressi, G.; Grego, E. Detection of Rickettsia aeschlimannii and Rickettsia africae in ixodid ticks from Burkina Faso and Somali region of Ethiopia by new real-time PCR assays. Ticks Tick Borne Dis. 2016, 7, 1082–1088. [Google Scholar] [CrossRef] [PubMed]
- Yessinou, R.E.; Cazan, C.D.; Panait, L.C.; Mollong, E.; Biguezoton, A.S.; Bonnet, S.I.; Farougou, S.; Groschup, M.H.; Mihalca, A.D. New geographical records for tick-borne pathogens in ticks collected from cattle in Benin and Togo. Vet. Med. Sci. 2023, 9, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Nimo-Paintsil, S.C.; Mosore, M.; Addo, S.O.; Lura, T.; Tagoe, J.; Ladzekpo, D.; Addae, C.; Bentil, R.E.; Behene, E.; Dafeamekpor, C.; et al. Ticks and prevalence of tick-borne pathogens from domestic animals in Ghana. Parasit. Vectors 2022, 15, 86. [Google Scholar] [CrossRef] [PubMed]
- Yessinou, R.E.; Adehan, S.; Hedegbetan, G.C.; Cassini, R.; Mantip, S.E.; Farougou, S. Molecular characterization of Rickettsia spp., Bartonella spp., and Anaplasma phagocytophilum in hard ticks collected from wild animals in Benin, West Africa. Trop. Anim. Health Prod. 2022, 54, 306. [Google Scholar] [CrossRef] [PubMed]
- Adjou Moumouni, P.F.; Aplogan, G.L.; Katahira, H.; Gao, Y.; Guo, H.; Efstratiou, A.; Jirapattharasate, C.; Wang, G.; Liu, M.; Ringo, A.E.; et al. Prevalence, risk factors, and genetic diversity of veterinary important tick-borne pathogens in cattle from Rhipicephalus microplus-invaded and non-invaded areas of Benin. Ticks Tick Borne Dis. 2018, 9, 450–464. [Google Scholar] [CrossRef]
- Adamu, M.; Troskie, M.; Oshadu, D.O.; Malatji, D.P.; Penzhorn, B.L.; Matjila, P.T. Occurrence of tick-transmitted pathogens in dogs in Jos, Plateau State, Nigeria. Parasit. Vectors 2014, 7, 119. [Google Scholar] [CrossRef] [PubMed]
- Adjou Moumouni, P.F.; Guo, H.; Gao, Y.; Liu, M.; Ringo, A.E.; Galon, E.M.; Vudriko, P.; Umemiya-Shirafuji, R.; Inoue, N.; Suzuki, H.; et al. Identification and genetic characterization of Piroplasmida and Anaplasmataceae agents in feeding Amblyomma variegatum ticks from Benin. Vet. Parasitol. Reg. Stud. Rep. 2018, 14, 137–143. [Google Scholar] [CrossRef]
- Anifowose, O.I.; Takeet, M.I.; Talabi, A.O.; Otesile, E.B. Molecular detection of Ehrlichia ruminantium in engorged Amblyomma variegatum and cattle in Ogun State, Nigeria. J. Parasit. Dis. 2020, 44, 403–410. [Google Scholar] [CrossRef]
- Faburay, B.; Geysen, D.; Munstermann, S.; Taoufik, A.; Postigo, M.; Jongejan, F. Molecular detection of Ehrlichia ruminantium infection in Amblyomma variegatum ticks in Gambia. Exp. Appl. Acarol. 2007, 42, 61–74. [Google Scholar] [CrossRef]
- Ouedraogo, A.S.; Zannou, O.M.; Biguezoton, A.S.; Yao, K.P.; Belem, A.M.G.; Farougou, S.; Oosthuizen, M.; Saegerman, C.; Lempereur, L. Cross border transhumance involvement in ticks and tick-borne pathogens dissemination and first evidence of Anaplasma centrale in Burkina Faso. Ticks Tick Borne Dis. 2021, 12, 101781. [Google Scholar] [CrossRef] [PubMed]
- Mediannikov, O.; Fenollar, F.; Socolovschi, C.; Diatta, G.; Bassene, H.; Molez, J.F.; Sokhna, C.; Trape, J.F.; Raoult, D. Coxiella burnetii in humans and ticks in rural Senegal. PLoS Negl. Trop. Dis. 2010, 4, e654. [Google Scholar] [CrossRef]
- Ouedraogo, A.S.; Zannou, O.M.; Biguezoton, A.S.; Kouassi, P.Y.; Belem, A.; Farougou, S.; Oosthuizen, M.; Saegerman, C.; Lempereur, L. Cattle ticks and associated tick-borne pathogens in Burkina Faso and Benin: Apparent northern spread of Rhipicephalus microplus in Benin and first evidence of Theileria velifera and Theileria annulata. Ticks Tick Borne Dis. 2021, 12, 101733. [Google Scholar] [CrossRef]
- Kartashov, M.Y.; Naidenova, E.V.; Zakharov, K.S.; Yakovlev, S.A.; Skarnovich, M.O.; Boumbaly, S.; Nikiforov, K.A.; Plekhanov, N.A.; Kritzkiy, A.A.; Ternovoi, V.A.; et al. Detection of Babesia caballi, Theileria mutans and Th. velifera in ixodid ticks collected from cattle in Guinea in 2017–2018. Vet. Parasitol. Reg. Stud. Rep. 2021, 24, 100564. [Google Scholar] [CrossRef]
- Tomassone, L.; Pagani, P.; De, M.D. Detection of Babesia caballi in Amblyomma variegatum ticks (Acari: Ixodidae) collected from cattle in the Republic of Guinea. Parassitologia 2005, 47, 247–251. [Google Scholar] [PubMed]
- Akuffo, R.; Brandful, J.A.; Zayed, A.; Adjei, A.; Watany, N.; Fahmy, N.T.; Hughes, R.; Doman, B.; Voegborlo, S.V.; Aziati, D.; et al. Crimean-Congo hemorrhagic fever virus in livestock ticks and animal handler seroprevalence at an abattoir in Ghana. BMC Infect. Dis. 2016, 16, 324. [Google Scholar] [CrossRef]
- Zeller, H.G.; Cornet, J.P.; Diop, A.; Camicas, J.L. Crimean-Congo hemorrhagic fever in ticks (Acari: Ixodidae) and ruminants: Field observations of an epizootic in Bandia, Senegal (1989–1992). J. Med. Entomol. 1997, 34, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Onyiche, T.E.; Raileanu, C.; Tauchmann, O.; Fischer, S.; Vasic, A.; Schäfer, M.; Biu, A.A.; Ogo, I.N.; Thekisoe, O.; Silaghi, C. Prevalence and molecular characterization of ticks and tick-borne pathogens of one-humped camels (Camelus dromedarius) in Nigeria. Parasit. Vectors 2020, 13, 428. [Google Scholar] [CrossRef] [PubMed]
- d’Oliveira, C.; van der Weide, M.; Jacquiet, P.; Jongejan, F. Detection of Theileria annulata by the PCR in ticks (Acari:Ixodidae) collected from cattle in Mauritania. Exp. Appl. Acarol. 1997, 21, 279–291. [Google Scholar] [CrossRef]
- Onyiche, T.E.; Taioe, M.O.; Ogo, N.I.; Sivakumar, T.; Biu, A.A.; Mbaya, A.W.; Xuan, X.; Yokoyama, N.; Thekisoe, O. Molecular evidence of Babesia caballi and Theileria equi in equines and ticks in Nigeria: Prevalence and risk factors analysis. Parasitology 2020, 147, 1238–1248. [Google Scholar] [CrossRef]
- Kamani, J.; Baneth, G.; Apanaskevich, D.A.; Mumcuoglu, K.Y.; Harrus, S. Molecular detection of Rickettsia aeschlimannii in Hyalomma spp. ticks from camels (Camelus dromedarius) in Nigeria, West Africa. Med. Vet. Entomol. 2015, 29, 205–209. [Google Scholar] [CrossRef]
- Saluzzo, J.F.; Digoutte, J.P.; Camicas, J.L.; Chauvancy, G. Crimean-Congo haemorrhagic fever and Rift Valley fever in south-eastern Mauritania. Lancet 1985, 1, 116. [Google Scholar] [CrossRef]
- Zeller, H.G.; Cornet, J.P.; Camicas, J.L. Crimean-Congo haemorrhagic fever virus infection in birds: Field investigations in Senegal. Res. Virol. 1994, 145, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, V.; Wijnveld, M.; Majekodunmi, A.O.; Dongkum, C.; Fajinmi, A.; Dogo, A.G.; Thrusfield, M.; Mugenyi, A.; Vaumourin, E.; Igweh, A.C.; et al. Tick-borne pathogens of zoonotic and veterinary importance in Nigerian cattle. Parasit. Vectors 2016, 9, 217. [Google Scholar] [CrossRef] [PubMed]
- Elbir, H.; FotsoFotso, A.; Diatta, G.; Trape, J.F.; Arnathau, C.; Renaud, F.; Durand, P. Ubiquitous bacteria Borrelia crocidurae in Western African ticks Ornithodoros sonrai. Parasit. Vectors 2015, 8, 477. [Google Scholar] [CrossRef] [PubMed]
- Ndiaye, E.H.I.; Diouf, F.S.; Ndiaye, M.; Bassene, H.; Raoult, D.; Sokhna, C.; Parola, P.; Diatta, G. Tick-borne relapsing fever Borreliosis, a major public health problem overlooked in Senegal. PLoS Negl. Trop. Dis. 2021, 15, e0009184. [Google Scholar] [CrossRef] [PubMed]
- Ouarti, B.; Sall, M.; Ndiaye, E.H.I.; Diatta, G.; Diarra, A.Z.; Berenger, J.M.; Sokhna, C.; Granjon, L.; Le Fur, J.; Parola, P. Pathogen detection in Ornithodoros sonrai ticks and invasive house Mice Mus musculus domesticus in Senegal. Microorganisms 2022, 10, 2367. [Google Scholar] [CrossRef]
- Schwan, T.G.; Anderson, J.M.; Lopez, J.E.; Fischer, R.J.; Raffel, S.J.; McCoy, B.N.; Safronetz, D.; Sogoba, N.; Maïga, O.; Traoré, C.F. Endemic foci of the tick-borne relapsing fever spirochete Borrelia crocidurae in Mali, West Africa, and the potential for human infection. PLoS Negl. Trop. Dis. 2012, 6, e1924. [Google Scholar] [CrossRef]
- Trape, J.F.; Diatta, G.; Arnathau, C.; Bitam, I.; Sarih, M.; Belghyti, D.; Bouattour, A.; Elguero, E.; Vial, L.; Mané, Y.; et al. The epidemiology and geographic distribution of relapsing fever borreliosis in West and North Africa, with a review of the Ornithodoros erraticus complex (Acari: Ixodida). PLoS ONE 2013, 8, e78473. [Google Scholar] [CrossRef]
- Mediannikov, O.; Diatta, G.; Kasongo, K.; Raoult, D. Identification of Bartonellae in the soft tick species Ornithodoros sonrai in Senegal. Vector Borne Zoonotic Dis. 2014, 14, 26–32. [Google Scholar] [CrossRef]
- Dahmana, H.; Granjon, L.; Diagne, C.; Davoust, B.; Fenollar, F.; Mediannikov, O. Rodents as hosts of pathogens and related zoonotic disease risk. Pathogens 2020, 9, 202. [Google Scholar] [CrossRef]
- Vial, L.; Wieland, B.; Jori, F.; Etter, E.; Dixon, L.; Roger, F. African swine fever virus DNA in soft ticks, Senegal. Emerg. Infect. Dis. 2007, 13, 1928–1931. [Google Scholar] [CrossRef]
- Akinboade, O.A.; Dipeolu, O.O. Bovine babesiosis in Nigeria: Detection of Babesia organisms in salivary glands of Boophilus decoloratus collected on trade cattle. Zentralbl. Veterinarmed. B 1983, 30, 153–155. [Google Scholar] [CrossRef]
- Makenov, M.T.; Toure, A.H.; Korneev, M.G.; Sacko, N.; Porshakov, A.M.; Yakovlev, S.A.; Radyuk, E.V.; Zakharov, K.S.; Shipovalov, A.V.; Boumbaly, S.; et al. Rhipicephalus microplus and its vector-borne haemoparasites in Guinea: Further species expansion in West Africa. Parasitol. Res. 2021, 120, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- McCoy, B.N.; Maiga, O.; Schwan, T.G. Detection of Borrelia theileri in Rhipicephalus geigyi from Mali. Ticks Tick Borne Dis. 2014, 5, 401–403. [Google Scholar] [CrossRef]
- Biguezoton, A.; Noel, V.; Adehan, S.; Adakal, H.; Dayo, G.K.; Zoungrana, S.; Farougou, S.; Chevillon, C. Ehrlichia ruminantium infects Rhipicephalus microplus in West Africa. Parasit. Vectors 2016, 9, 354. [Google Scholar] [CrossRef]
- Dahmani, M.; Davoust, B.; Sambou, M.; Bassene, H.; Scandola, P.; Ameur, T.; Raoult, D.; Fenollar, F.; Mediannikov, O. Molecular investigation and phylogeny of species of the Anaplasmataceae infecting animals and ticks in Senegal. Parasit. Vectors 2019, 12, 495. [Google Scholar] [CrossRef]
- Ratmanov, P.; Bassene, H.; Fenollar, F.; Tall, A.; Sokhna, C.; Raoult, D.; Mediannikov, O. The correlation of Q fever and Coxiella burnetii DNA in household environments in rural Senegal. Vector Borne Zoonotic Dis. 2013, 13, 70–72. [Google Scholar] [CrossRef] [PubMed]
- Kamani, J.; Baneth, G.; Gutierrez, R.; Nachum-Biala, Y.; Mumcuoglu, K.Y.; Harrus, S. Coxiella burnetii and Rickettsia conorii: Two zoonotic pathogens in peridomestic rodents and their ectoparasites in Nigeria. Ticks Tick Borne Dis. 2018, 9, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Matei, I.A.; D’Amico, G.; Yao, P.K.; Ionica, A.M.; Kanyari, P.W.; Daskalaki, A.A.; Dumitrache, M.O.; Sándor, A.D.; Gherman, C.M.; Qablan, M.; et al. Molecular detection of Anaplasma platys infection in free-roaming dogs and ticks from Kenya and Ivory Coast. Parasit. Vectors 2016, 9, 157. [Google Scholar] [CrossRef]
- Socolovschi, C.; Gomez, J.; Marie, J.L.; Davoust, B.; Guigal, P.M.; Raoult, D.; Parola, P. Ehrlichia canis in Rhipicephalus sanguineus ticks in the Ivory Coast. Ticks Tick Borne Dis. 2012, 3, 411–413. [Google Scholar] [CrossRef]
- Clarke, L.L.; Ballweber, L.R.; Allen, K.; Little, S.E.; Lappin, M.R. Prevalence of select vector-borne disease agents in owned dogs of Ghana. J. S. Afr. Vet. Assoc. 2014, 85, 996. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Elelu, N.; Ola-Fadunsin, S.D.; Bankole, A.A.; Raji, M.A.; Ogo, N.I.; Cutler, S.J. Prevalence of tick infestation and molecular characterization of spotted fever Rickettsia massiliae in Rhipicephalus species parasitizing domestic small ruminants in north-central Nigeria. PLoS ONE 2022, 17, e0263843. [Google Scholar] [CrossRef] [PubMed]
- Socolovschi, C.; Gaudart, J.; Bitam, I.; Huynh, T.P.; Raoult, D.; Parola, P. Why are there so few Rickettsia conorii conorii-infected Rhipicephalus sanguineus ticks in the wild? PLoS Negl. Trop. Dis. 2012, 6, e1697. [Google Scholar] [CrossRef] [PubMed]
- Brouqui, P.; Parola, P.; Fournier, P.E.; Raoult, D. Spotted fever rickettsioses in southern and eastern Europe. FEMS Immunol. Med. Microbiol. 2007, 49, 2–12. [Google Scholar] [CrossRef]
- Dupont, H.T.; Brouqui, P.; Faugere, B.; Raoult, D. Prevalence of antibodies to Coxiella burnetti, Rickettsia conorii, and Rickettsia typhi in seven African countries. Clin. Infect. Dis. 1995, 21, 1126–1133. [Google Scholar] [CrossRef]
- Raoult, D.; Fournier, P.E.; Abboud, P.; Caron, F. First documented human Rickettsia aeschlimannii infection. Emerg. Infect. Dis. 2002, 8, 748–749. [Google Scholar] [CrossRef]
- Berrelha, J.; Briolant, S.; Muller, F.; Rolain, J.M.; Marie, J.L.; Pages, F.; Raoult, D.; Parola, P. Rickettsia felis and Rickettsia massiliae in Ivory Coast, Africa. Clin. Microbiol. Infect. 2009, 15, 251–252. [Google Scholar] [CrossRef]
- Elbir, H.; Raoult, D.; Drancourt, M. Relapsing fever borreliae in Africa. Am. J. Trop. Med. Hyg. 2013, 89, 288–292. [Google Scholar] [CrossRef]
- Jakab, A.; Kahlig, P.; Kuenzli, E.; Neumayr, A. Tick borne relapsing fever—A systematic review and analysis of the literature. PLoS Negl. Trop. Dis. 2022, 16, e0010212. [Google Scholar] [CrossRef]
- Ehounoud, C.B.; Fenollar, F.; Dahmani, M.; N’Guessan, J.D.; Raoult, D.; Mediannikov, O. Bacterial arthropod-borne diseases in West Africa. Acta Trop. 2017, 171, 124–137. [Google Scholar] [CrossRef]
- Parola, P.; Diatta, G.; Socolovschi, C.; Mediannikov, O.; Tall, A.; Bassene, H.; Trape, J.F.; Raoult, D. Tick-borne relapsing fever borreliosis, rural senegal. Emerg. Infect. Dis. 2011, 17, 883–885. [Google Scholar] [CrossRef]
- Cutler, S.J.; Abdissa, A.; Trape, J.F. New concepts for the old challenge of African relapsing fever borreliosis. Clin. Microbiol. Infect. 2009, 15, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Trape, J.F.; Duplantier, J.M.; Bouganali, H.; Godeluck, B.; Legros, F.; Cornet, J.P.; Camicas, J.L. Tick-borne borreliosis in west Africa. Lancet 1991, 337, 473–475. [Google Scholar] [CrossRef] [PubMed]
- Vial, L.; Diatta, G.; Tall, A.; Bouganali, H.; Bouganali, H.; Durand, P.; Sokhna, C.; Rogier, C.; Renaud, F.; Trape, J.F. Incidence of tick-borne relapsing fever in west Africa: Longitudinal study. Lancet 2006, 368, 37–43. [Google Scholar] [CrossRef]
- Trape, J.F.; Godeluck, B.; Diatta, G.; Rogier, C.; Legros, F.; Albergel, J.; Pepin, Y.; Duplantier, J.M. The spread of tick-borne borreliosis in West Africa and its relationship to sub-Saharan drought. Am. J. Trop. Med. Hyg. 1996, 54, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Brahim, H.; Perrier-Gros-Claude, J.D.; Postic, D.; Baranton, G.; Jambou, R. Identifying relapsing fever Borrelia, Senegal. Emerg. Infect. Dis. 2005, 11, 474–475. [Google Scholar] [CrossRef]
- Sokhna, C.; Mediannikov, O.; Fenollar, F.; Bassene, H.; Diatta, G.; Tall, A.; Trape, J.F.; Drancourt, M.; Raoult, D. Point-of-care laboratory of pathogen diagnosis in rural Senegal. PLoS Negl. Trop. Dis. 2013, 7, e1999. [Google Scholar] [CrossRef]
- Patrat-Delon, S.; Drogoul, A.S.; Le, H.H.; Biziraguzenyuka, J.; Rabier, V.; Arvieux, C.; Michelet, C.; Chevrier, S.; Tattevin, P. Recurrent tick-borne fever: A possible diagnosis in patients returning from Senegal. Med. Mal. Infect. 2008, 38, 396–399. [Google Scholar] [CrossRef]
- El, H.I.; Diatta, G.; Diarra, A.Z.; Bassene, H.; Sokhna, C.; Parola, P. Quantitative polymerase chain ceaction from malaria rapid diagnostic tests to betect Borrelia crocidurae, the agent of tick-borne relapsing fever, in febrile patients in Senegal. Am. J. Trop. Med. Hyg. 2023, 108, 968–976. [Google Scholar]
- Nordstrand, A.; Bunikis, I.; Larsson, C.; Tsogbe, K.; Schwan, T.G.; Nilsson, M.; Bergström, S. Tick-borne relapsing fever diagnosis obscured by malaria, Togo. Emerg. Infect. Dis. 2007, 13, 117–123. [Google Scholar] [CrossRef]
- Toure, M.; Petersen, P.T.; Bathily, S.N.; Sanogo, D.; Wang, C.W.; Schioler, K.L.; Konradsen, F.; Doumbia, S.; Alifrangis, M. Molecular evidence of malaria and zoonotic diseases among rapid diagnostic test-negative febrile patients in low-transmission season, Mali. Am. J. Trop. Med. Hyg. 2017, 96, 335–337. [Google Scholar] [CrossRef] [PubMed]
- Diatta, G.; Duplantier, J.M.; Granjon, L.; Ba, K.; Chauvancy, G.; Ndiaye, M.; Trape, J.F. Borrelia infection in small mammals in West Africa and its relationship with tick occurrence inside burrows. Acta Trop. 2015, 152, 131–140. [Google Scholar] [CrossRef]
- Godeluck, B.; Duplantier, J.M.; Ba, K.; Trape, J.F. A longitudinal survey of Borrelia crocidurae prevalence in rodents and insectivores in Senegal. Am. J. Trop. Med. Hyg. 1994, 50, 165–168. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Koney, E.B.; Dogbey, O.; Walker, A.R. Incidence and prevalence of tick-borne haemoparasites in domestic ruminants in Ghana. Vet. Parasitol. 2004, 124, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Bankole, A.A.; Kumsa, B.; Mamo, G.; Ogo, N.I.; Elelu, N.; Morgan, W.; Cutler, S.J. Comparative analysis of tick-borne relapsing fever spirochaetes from Ethiopia and Nigeria. Pathogens 2023, 12, 81. [Google Scholar] [CrossRef]
- Cutler, S.J.; Idris, J.M.; Ahmed, A.O.; Elelu, N. Ornithodoros savignyi, the tick vector of “Candidatus Borrelia kalaharica” in Nigeria. J. Clin. Microbiol. 2018, 56, 10–1128. [Google Scholar] [CrossRef]
- Dumic, I.; Jevtic, D.; Veselinovic, M.; Nordstrom, C.W.; Jovanovic, M.; Mogulla, V.; Veselinovic, E.M.; Hudson, A.; Simeunovic, G.; Petcu, E.; et al. Human granulocytic anaplasmosis-a systematic review of published cases. Microorganisms 2022, 10, 1433. [Google Scholar] [CrossRef]
- Salinas-Estrella, E.; Amaro-Estrada, I.; Cobaxin-Cardenas, M.E.; Preciado de la Torre, J.F.; Rodriguez, S.D. Bovine anaplasmosis: Will there ever be an almighty effective vaccine? Front. Vet. Sci. 2022, 9, 946545. [Google Scholar] [CrossRef]
- Woldehiwet, Z. The natural history of Anaplasma phagocytophilum. Vet. Parasitol. 2010, 167, 108–122. [Google Scholar] [CrossRef]
- Carrade, D.D.; Foley, J.E.; Borjesson, D.L.; Sykes, J.E. Canine granulocytic anaplasmosis: A review. J. Vet. Intern. Med. 2009, 23, 1129–1141. [Google Scholar] [CrossRef]
- Sanchez, E.; Vannier, E.; Wormser, G.P.; Hu, L.T. Diagnosis, treatment, and prevention of Lyme disease, human granulocytic anaplasmosis, and babesiosis: A Review. JAMA 2016, 315, 1767–1777. [Google Scholar] [CrossRef]
- Aquino, L.C.; Kamani, J.; Haruna, A.M.; Paludo, G.R.; Hicks, C.A.; Helps, C.R.; Tasker, S. Analysis of risk factors and prevalence of haemoplasma infection in dogs. Vet. Parasitol. 2016, 221, 111–117. [Google Scholar] [CrossRef]
- Djiba, M.L.; Mediannikov, O.; Mbengue, M.; Thiongane, Y.; Molez, J.F.; Seck, M.T.; Fenollar, F.; Raoult, D.; Ndiaye, M. Survey of Anaplasmataceae bacteria in sheep from Senegal. Trop. Anim. Health Prod. 2013, 45, 1557–1561. [Google Scholar] [CrossRef]
- Battilani, M.; De, A.S.; Balboni, A.; Dondi, F. Genetic diversity and molecular epidemiology of Anaplasma. Infect. Genet. Evol. 2017, 49, 195–211. [Google Scholar] [CrossRef]
- Gueye, A.; Camicas, J.L.; Diouf, A.; Mbengue, M. Ticks and hemoparasitoses of livestock in Senegal. II. The Sahelian area. Rev. Elev. Med. Vet. Pays Trop. 1987, 40, 119–125. [Google Scholar]
- Gueye, A.; Mbengue, M.; Diouf, A. Ticks and hemoparasitic diseases in cattle in Senegal. IV. The southern Sudan area. Rev. Elev. Med. Vet. Pays Trop. 1990, 42, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Gueye, A.; Mbengue, M.; Diouf, A.; Sonko, M.L. Ticks and hemoparasitoses in livestock in Senegal. V. The northern Guinea area. Rev. Elev. Med. Vet. Pays Trop. 1993, 46, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, R.C.; Bah, M.; Kora, S.; Cassama, M.; Clifford, D.J. Susceptibility to different tick genera in Gambian N’Dama and Gobra zebu cattle exposed to naturally occurring tick infestations. Trop. Anim. Health Prod. 1995, 27, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Elelu, N.; Ferrolho, J.; Couto, J.; Domingos, A.; Eisler, M.C. Molecular diagnosis of the tick-borne pathogen Anaplasma marginale in cattle blood samples from Nigeria using qPCR. Exp. Appl. Acarol. 2016, 70, 501–510. [Google Scholar] [CrossRef]
- Heylen, D.J.A.; Kumsa, B.; Kimbita, E.; Frank, M.N.; Muhanguzi, D.; Jongejan, F.; Adehan, S.B.; Toure, A.; Aboagye-Antwi, F.; Ogo, N.I.; et al. Tick-borne pathogens and body condition of cattle in smallholder rural livestock production systems in East and West Africa. Parasit. Vectors 2023, 16, 117. [Google Scholar] [CrossRef]
- Kamani, J.; Schaer, J.; Umar, A.G.; Pilarshimwi, J.Y.; Bukar, L.; Gonzalez-Miguel, J.; Harrus, S. Molecular detection and genetic characterization of Anaplasma marginale and Anaplasma platys in cattle in Nigeria. Ticks Tick Borne Dis. 2022, 13, 101955. [Google Scholar] [CrossRef]
- Kamani, J.; Irene, S.; Qasim, A.M.M.M.; Olubade, T.A.; Abasiama, M.S.; Gajibo, A.; Balami, P.U.; Shands, M.; Harrus, S. Nucleotide sequence types (ntSTs) of Anaplasma marginale in cattle in Nigeria based on the major surface protein 5 (msp5) gene. Acta Trop. 2022, 233, 106544. [Google Scholar] [CrossRef] [PubMed]
- Toure, A.; Sanogo, M.; Sghiri, A.; Sahibi, H. Incidences of Rhipicephalus (Boophilus) microplus (Canestrini, 1888) transmitted pathogens in cattle in West Africa. Acta Parasitol. 2022, 67, 1282–1289. [Google Scholar] [CrossRef]
- Dahmani, M.; Marié, J.L.; Scandola, P.; Brah, S.; Davoust, B.; Mediannikov, O. Anaplasma ovis infects sheep in Niger. Small Rumin. Res. 2017, 151, 32–35. [Google Scholar] [CrossRef]
- Gruenberger, I.; Liebich, A.V.; Ajibade, T.O.; Obebe, O.O.; Ogbonna, N.F.; Wortha, L.N.; Unterköfler, M.S.; Fuehrer, H.P.; Ayinmode, A.B. Vector-borne pathogens in duard Dogs in Ibadan, Nigeria. Pathogens 2023, 12, 406. [Google Scholar] [CrossRef] [PubMed]
- Heylen, D.; Day, M.; Schunack, B.; Fourie, J.; Labuschange, M.; Johnson, S.; Githigia, S.M.; Akande, F.A.; Nzalawahe, J.S.; Tayebwa, D.S.; et al. A community approach of pathogens and their arthropod vectors (ticks and fleas) in dogs of African Sub-Sahara. Parasit. Vectors 2021, 14, 576. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, V.; Wijnveld, M.; Latrofa, M.S.; Fajinmi, A.; Majekodunmi, A.O.; Dogo, A.G.; Igweh, A.C.; Otranto, D.; Jongejan, F.; Welburn, S.C.; et al. Canine and ovine tick-borne pathogens in camels, Nigeria. Vet. Parasitol. 2016, 228, 90–92. [Google Scholar] [CrossRef]
- Gotsch, S.; Leschnik, M.; Duscher, G.; Burgstaller, J.P.; Wille-Piazzai, W.; Joachim, A. Ticks and haemoparasites of dogs from Praia, Cape Verde. Vet. Parasitol. 2009, 166, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Lauzi, S.; Maia, J.P.; Epis, S.; Marcos, R.; Pereira, C.; Luzzago, C.; Santos, M.; Puente-Payo, P.; Giordano, A.; Pajoro, M.; et al. Molecular detection of Anaplasma platys, Ehrlichia canis, Hepatozoon canis and Rickettsia monacensis in dogs from Maio Island of Cape Verde archipelago. Ticks Tick Borne Dis. 2016, 7, 964–969. [Google Scholar] [CrossRef]
- Zobba, R.; Murgia, C.; Dahmani, M.; Mediannikov, O.; Davoust, B.; Piredda, R.; Schianchi, E.; Scagliarini, A.; Pittau, M.; Alberti, A. Emergence of Anaplasma species related to A. phagocytophilum and A. platys in Senegal. Int. J. Mol. Sci. 2022, 24, 35. [Google Scholar] [CrossRef] [PubMed]
- Vieira, R.F.; Biondo, A.W.; Guimaraes, A.M.; dos Santos, A.P.; Dos Santos, R.P.; Dutra, L.H.; Diniz, P.P.; de Morais, H.A.; Messick, J.B.; Labruna, M.B.; et al. Ehrlichiosis in Brazil. Rev. Bras. Parasitol. Vet. 2011, 20, 1–12. [Google Scholar] [CrossRef]
- Wakeel, A.; Zhu, B.; Yu, X.J.; McBride, J.W. New insights into molecular Ehrlichia chaffeensis-host interactions. Microbes Infect. 2010, 12, 337–345. [Google Scholar] [CrossRef][Green Version]
- Rikihisa, Y. Molecular events involved in cellular invasion by Ehrlichia chaffeensis and Anaplasma phagocytophilum. Vet. Parasitol. 2010, 167, 155–166. [Google Scholar] [CrossRef]
- Uhaa, I.J.; MacLean, J.D.; Greene, C.R.; Fishbein, D.B. A case of human ehrlichiosis acquired in Mali: Clinical and laboratory findings. Am. J. Trop. Med. Hyg. 1992, 46, 161–164. [Google Scholar] [CrossRef]
- Brouqui, P.; Le, C.C.; Kelly, P.J.; Laurens, R.; Tounkara, A.; Sawadogo, S.; lo-Marcel, V.; Gondao, L.; Faugere, B.; Delmont, J. Serologic evidence for human ehrlichiosis in Africa. Eur. J. Epidemiol. 1994, 10, 695–698. [Google Scholar] [CrossRef]
- Thomas, R.J.; Dumler, J.S.; Carlyon, J.A. Current management of human granulocytic anaplasmosis, human monocytic ehrlichiosis and Ehrlichia ewingii ehrlichiosis. Expert Rev. Anti Infect. Ther. 2009, 7, 709–722. [Google Scholar] [CrossRef]
- Allsopp, B.A. Heartwater—Ehrlichia ruminantium infection. Rev. Sci. Tech. 2015, 34, 557–568. [Google Scholar] [CrossRef]
- Faburay, B.; Munstermann, S.; Geysen, D.; Bell-Sakyi, L.; Ceesay, A.; Bodaan, C.; Jongejan, F. Point seroprevalence survey of Ehrlichia ruminantium infection in small ruminants in Gambia. Clin. Diagn. Lab. Immunol. 2005, 12, 508–512. [Google Scholar] [CrossRef]
- Knopf, L.; Komoin-Oka, C.; Betschart, B.; Jongejan, F.; Gottstein, B.; Zinsstag, J. Seasonal epidemiology of ticks and aspects of cowdriosis in N’Dama village cattle in the Central Guinea savannah of Cote d’Ivoire. Prev. Vet. Med. 2002, 53, 21–30. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Koney, E.B.; Dogbey, O.; Sumption, K.J.; Walker, A.R.; Bath, A.; Jongejan, F. Detection by two enzyme-linked immunosorbent assays of antibodies to Ehrlichia ruminantium in field sera collected from sheep and cattle in Ghana. Clin. Diagn. Lab. Immunol. 2003, 10, 917–925. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Koney, E.B.; Dogbey, O.; Walker, A.R. Ehrlichia ruminantium seroprevalence in domestic ruminants in Ghana: I. Longitudinal survey in the Greater Accra Region. Vet. Microbiol. 2004, 100, 175–188. [Google Scholar] [CrossRef]
- Faburay, B.; Geysen, D.; Munstermann, S.; Bell-Sakyi, L.; Jongejan, F. Longitudinal monitoring of Ehrlichia ruminantium infection in Gambian lambs and kids by pCS20 PCR and MAP1-B ELISA. BMC Infect. Dis. 2007, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Adakal, H.; Gavotte, L.; Stachurski, F.; Konkobo, M.; Henri, H.; Zoungrana, S.; Huber, K.; Vachiery, N.; Martinez, D.; Morand, S.; et al. Clonal origin of emerging populations of Ehrlichia ruminantium in Burkina Faso. Infect. Genet. Evol. 2010, 10, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Davoust, B.; Bourry, O.; Gomez, J.; Lafay, L.; Casali, F.; Leroy, E.; Parzy, D. Surveys on seroprevalence of canine monocytic ehrlichiosis among dogs living in the Ivory Coast and Gabon and evaluation of a quick commercial test kit dot-ELISA. Ann. N. Y. Acad. Sci. 2006, 1078, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Davoust, B.; Parzy, D.; Demoncheaux, J.P.; Tine, R.; Diarra, M.; Marie, J.L.; Mediannikov, O. Usefulness of a rapid immuno-migration test for the detection of canine monocytic ehrlichiosis in Africa. Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 31–37. [Google Scholar] [CrossRef]
- Daramola, O.O.; Takeet, M.I.; Oyewusi, I.K.; Oyekunle, M.A.; Talabi, A.O. Detection and molecular characterisation of Ehrlichia canis in naturally infected dogs in South West Nigeria. Acta Vet. Hung. 2018, 66, 85–95. [Google Scholar] [CrossRef]
- Happi, A.N.; Toepp, A.J.; Ugwu, C.A.; Petersen, C.A.; Sykes, J.E. Detection and identification of blood-borne infections in dogs in Nigeria using light microscopy and the polymerase chain reaction. Vet. Parasitol. Reg. Stud. Rep. 2018, 11, 55–60. [Google Scholar] [CrossRef]
- Kamani, J.; Baneth, G.; Mumcuoglu, K.Y.; Waziri, N.E.; Eyal, O.; Guthmann, Y.; Harrus, S. Molecular detection and characterization of tick-borne pathogens in dogs and ticks from Nigeria. PLoS Negl. Trop. Dis. 2013, 7, e2108. [Google Scholar] [CrossRef]
- Kamani, J.; Lee, C.C.; Haruna, A.M.; Chung, P.J.; Weka, P.R.; Chung, Y.T. First detection and molecular characterization of Ehrlichia canis from dogs in Nigeria. Res. Vet. Sci. 2013, 94, 27–32. [Google Scholar] [CrossRef]
- Davoust, B.; Mediannikov, O.; Chene, J.; Massot, R.; Tine, R.; Diarra, M.; Demoncheaux, J.P.; Scandola, P.; Beugnet, F.; Chabanne, L. Study of ehrlichiosis in kennel dogs under treatment and prevention during seven months in Dakar (Senegal). Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 613–617. [Google Scholar] [CrossRef]
- Medkour, H.; Laidoudi, Y.; Athias, E.; Bouam, A.; Dizoe, S.; Davoust, B.; Mediannikov, O. Molecular and serological detection of animal and human vector-borne pathogens in the blood of dogs from Cote d’Ivoire. Comp. Immunol. Microbiol. Infect. Dis. 2020, 69, 101412. [Google Scholar] [CrossRef] [PubMed]
- Cheslock, M.A.; Embers, M.E. Human bartonellosis: An underappreciated public health problem? Trop. Med. Infect. Dis. 2019, 4, 69. [Google Scholar] [CrossRef] [PubMed]
- Shamekhi, A.F. Bartonellosis in chronic kidney disease: An unrecognized and unsuspected diagnosis. Ther. Apher. Dial. 2017, 21, 430–440. [Google Scholar] [CrossRef]
- Mannerings, A.O.; Osikowicz, L.M.; Restif, O.; Nyarko, E.; Suu-Ire, R.; Cunningham, A.A.; Wood, J.L.; Kosoy, M.Y. Exposure to bat-associated Bartonella spp. among humans and other animals, Ghana. Emerg. Infect. Dis. 2016, 22, 922–924. [Google Scholar] [CrossRef] [PubMed]
- Madder, M.; Day, M.; Schunack, B.; Fourie, J.; Labuschange, M.; van der Westhuizen, W.; Johnson, S.; Githigia, S.M.; Akande, F.A.; Nzalawahe, J.S.; et al. A community approach for pathogens and their arthropod vectors (ticks and fleas) in cats of sub-Saharan Africa. Parasit. Vectors 2022, 15, 321. [Google Scholar] [CrossRef]
- Angelakis, E.; Billeter, S.A.; Breitschwerdt, E.B.; Chomel, B.B.; Raoult, D. Potential for tick-borne bartonelloses. Emerg. Infect. Dis. 2010, 16, 385–391. [Google Scholar] [CrossRef]
- Diatta, G.; Mediannikov, O.; Sokhna, C.; Bassene, H.; Socolovschi, C.; Ratmanov, P.; Fenollar, F.; Raoult, D. Prevalence of Bartonella quintana in patients with fever and head lice from rural areas of Sine-Saloum, Senegal. Am. J. Trop. Med. Hyg. 2014, 91, 291–293. [Google Scholar] [CrossRef]
- Bai, Y.; Malania, L.; Alvarez, C.D.; Moran, D.; Boonmar, S.; Chanlun, A.; Suksawat, F.; Maruyama, S.; Knobel, D.; Kosoy, M. Global distribution of Bartonella infections in domestic bovine and characterization of Bartonella bovis strains using multi-locus sequence typing. PLoS ONE 2013, 8, e80894. [Google Scholar] [CrossRef]
- Dahmani, M.; Sambou, M.; Scandola, P.; Raoult, D.; Fenollar, F.; Mediannikov, O. Bartonella bovis and Candidatus Bartonella davousti in cattle from Senegal. Comp. Immunol. Microbiol. Infect. Dis. 2017, 50, 63–69. [Google Scholar] [CrossRef]
- Raoult, D.; La, S.B.; Kelly, P.J.; Davoust, B.; Gomez, J. Bartonella bovis in cattle in Africa. Vet. Microbiol. 2005, 105, 155–156. [Google Scholar]
- Martin-Alonso, A.; Houemenou, G.; Abreu-Yanes, E.; Valladares, B.; Feliu, C.; Foronda, P. Bartonella spp. in small mammals, Benin. Vector Borne Zoonotic Dis. 2016, 16, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Kamani, J.; Morick, D.; Mumcuoglu, K.Y.; Harrus, S. Prevalence and diversity of Bartonella species in commensal rodents and ectoparasites from Nigeria, West Africa. PLoS Negl. Trop. Dis. 2013, 7, e2246. [Google Scholar] [CrossRef] [PubMed]
- Diarra, A.Z.; Kone, A.K.; Doumbo, N.S.; Laroche, M.; Diatta, G.; Atteynine, S.A.; Coulibaly, M.; Sangare, A.K.; Kouriba, B.; Djimde, A.; et al. Molecular detection of microorganisms associated with small mammals and their ectoparasites in Mali. Am. J. Trop. Med. Hyg. 2020, 103, 2542–2551. [Google Scholar] [CrossRef] [PubMed]
- Kamani, J.; Baneth, G.; Mitchell, M.; Mumcuoglu, K.Y.; Gutierrez, R.; Harrus, S. Bartonella species in bats (Chiroptera) and bat flies (Nycteribiidae) from Nigeria, West Africa. Vector Borne Zoonotic Dis. 2014, 14, 625–632. [Google Scholar] [CrossRef]
- Eldin, C.; Melenotte, C.; Mediannikov, O.; Ghigo, E.; Million, M.; Edouard, S.; Mege, J.L.; Maurin, M.; Raoult, D. From Q Fever to Coxiella burnetii infection: A paradigm change. Clin. Microbiol. Rev. 2017, 30, 115–190. [Google Scholar] [CrossRef]
- Lyagoubi, M.; Fassin, D.; Rogeaux, O.; Gentilini, M. Q fever in Guinea-Bissau. 1 case. Bull. Soc. Pathol. Exot. Filiales 1989, 82, 575–577. [Google Scholar]
- Blondeau, J.; Yates, L.; Martin, R.; Marrie, T.; Ukoli, P.; Thomas, A. Q fever in Sokoto, Nigeria. Ann. N. Y. Acad. Sci. 1990, 590, 281–282. [Google Scholar] [CrossRef]
- Julvez, J.; Michault, A.; Kerdelhue, C. Serological study of rickettsia infections in Niamey, Niger. Med. Trop. 1997, 57, 153–156. [Google Scholar]
- Kobbe, R.; Kramme, S.; Kreuels, B.; Adjei, S.; Kreuzberg, C.; Panning, M.; Adjei, O.; Fleischer, B.; May, J. Q fever in young children, Ghana. Emerg. Infect. Dis. 2008, 14, 344–346. [Google Scholar] [CrossRef]
- Gidel, R.; Athawet, B. Serological survey of human brucellosis and rickettsial diseases in a group of a nomad population in the sahelian regions of Upper Volta. Ann. Soc. Belg. Med. Trop. 1975, 55, 77–83. [Google Scholar]
- Dean, A.S.; Bonfoh, B.; Kulo, A.E.; Boukaya, G.A.; Amidou, M.; Hattendorf, J.; Pilo, P.; Schelling, E. Epidemiology of brucellosis and q fever in linked human and animal populations in northern togo. PLoS ONE 2013, 8, e71501. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.; Hogerwerf, L.; Germeraad, E.A.; Roest, H.I.; Faye-Joof, T.; Jeng, M.; Nwakanma, D.; Secka, A.; Stegeman, A.; Goossens, B.; et al. Coxiella burnetii (Q fever) prevalence in associated populations of humans and small ruminants in Gambia. Trop. Med. Int. Health 2017, 22, 323–331. [Google Scholar] [CrossRef] [PubMed]
- van der Hoek, W.; Sarge-Njie, R.; Herremans, T.; Chisnall, T.; Okebe, J.; Oriero, E.; Versteeg, B.; Goossens, B.; van der Sande, M.; Kampmann, B.; et al. Short communication: Prevalence of antibodies against Coxiella burnetii (Q fever) in children in Gambia, West Africa. Trop. Med. Int. Health 2013, 18, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Adamu, S.G.; Kabir, J.; Umoh, J.U.; Raji, M.A. Seroprevalence of Coxiella burnetii in sheep flocks in Kaduna State, Northwestern Nigeria. Acta Vet. Hung. 2021, 69, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Adesiyun, A.A.; Jagun, A.G.; Kwaga, J.K.; Tekdek, L.B. Shedding of Coxiella burnetii in milk by Nigerian dairy and dual purposes cows. Int. J. Zoonoses 1985, 12, 1–5. [Google Scholar] [PubMed]
- Boni, M.; Davoust, B.; Tissot-Dupont, H.; Raoult, D. Survey of seroprevalence of Q fever in dogs in the southeast of France, French Guyana, Martinique, Senegal and the Ivory Coast. Vet. Microbiol. 1998, 64, 1–5. [Google Scholar] [CrossRef]
- Dione, M.M.; Sery, A.; Sidibe, C.A.K.; Wieland, B.; Fall, A. Exposure to multiple pathogens—Serological evidence for Rift Valley fever virus, Coxiella burnetii, Bluetongue virus and Brucella spp. in cattle, sheep and goat in Mali. PLoS Negl. Trop. Dis. 2022, 16, e0010342. [Google Scholar] [CrossRef]
- Folitse, R.D.; Opoku-Agyemang, T.; Amemor, E.; Opoku, E.D.; Bentum, K.E.; Emikpe, B.O. Serological evidence of Coxiella burnetii infection in slaughtered sheep and goats at Kumasi Abattoir, Ghana. J. Immunoassay Immunochem. 2020, 41, 152–157. [Google Scholar] [CrossRef]
- Kanoute, Y.B.; Gragnon, B.G.; Schindler, C.; Bonfoh, B.; Schelling, E. Reprint of “epidemiology of brucellosis, Q Fever and Rift Valley fever at the human and livestock interface in northern Cote d’Ivoire”. Acta Trop. 2017, 175, 121–129. [Google Scholar] [CrossRef]
- Klaasen, M.; Roest, H.J.; van der Hoek, W.; Goossens, B.; Secka, A.; Stegeman, A. Coxiella burnetii seroprevalence in small ruminants in Gambia. PLoS ONE 2014, 9, e85424. [Google Scholar] [CrossRef]
- Foronda, P.; Plata-Luis, J.; del Castillo-Figueruelo, B.; Fernandez-Alvarez, A.; Martin-Alonso, A.; Feliu, C.; Cabral, M.D.; Valladares, B. Serological survey of antibodies to Toxoplasma gondii and Coxiella burnetii in rodents in north-western African islands (Canary Islands and Cape Verde). Onderstepoort J. Vet. Res. 2015, 82, e1–e4. [Google Scholar] [CrossRef][Green Version]
- Breurec, S.; Poueme, R.; Fall, C.; Tall, A.; Diawara, A.; Bada-Alambedji, R.; Broutin, C.; Leclercq, A.; Garin, B. Microbiological quality of milk from small processing units in Senegal. Foodborne Pathog. Dis. 2010, 7, 601–604. [Google Scholar] [CrossRef]
- Mangombi-Pambou, J.; Granjon, L.; Labarrere, C.; Kane, M.; Niang, Y.; Fournier, P.E.; Delerce, J.; Fenollar, F.; Mediannikov, O. New genotype of Coxiella burnetii causing epizootic Q fever outbreak in rodents, northern Senegal. Emerg. Infect. Dis. 2023, 29, 1078–1081. [Google Scholar] [CrossRef]
- Amanzougaghene, N.; Fenollar, F.; Sangare, A.K.; Sissoko, M.S.; Doumbo, O.K.; Raoult, D.; Mediannikov, O. Detection of bacterial pathogens including potential new species in human head lice from Mali. PLoS ONE 2017, 12, e0184621. [Google Scholar] [CrossRef]
- Soosaraei, M.; Haghi, M.M.; Etemadifar, F.; Fakhar, M.; Teshnizi, S.H.; Hezarjaribi, H.Z.; Asfaram, S. Status of theileriosis among herbivores in Iran: A systematic review and meta-analysis. Vet. World 2018, 11, 332–341. [Google Scholar] [CrossRef]
- Morrison, W.I. The aetiology, pathogenesis and control of theileriosis in domestic animals. Rev. Sci. Tech. 2015, 34, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Clift, S.J.; Collins, N.E.; Oosthuizen, M.C.; Steyl, J.C.A.; Lawrence, J.A.; Mitchell, E.P. The pathology of pathogenic theileriosis in african wild artiodactyls. Vet. Pathol. 2020, 57, 24–48. [Google Scholar] [CrossRef]
- Kamani, J.; Bartova, E.; Kasparkova, N.; Mohammed, S.J.; Budikova, M.; Sedlak, K. Seroprevalence of Theileria equi, Babesia caballi, and Trichinella spp. infections in horses and donkeys from Nigeria, West Africa. Trop. Anim. Health Prod. 2021, 53, 338. [Google Scholar] [CrossRef] [PubMed]
- Onyiche, T.E.; Sivakumar, T.; Tuvshintulga, B.; Nugraha, A.B.; Ahedor, B.; Mofokeng, L.; Luka, J.; Mohammed, A.; Mbaya, A.W.; Biu, A.A.; et al. Serosurvey for equine piroplasms in horses and donkeys from north-western Nigeria using IFAT and ELISA. J. Immunoassay Immunochem. 2021, 42, 648–661. [Google Scholar] [CrossRef] [PubMed]
- Ehizibolo, D.O.; Kamani, J.; Ehizibolo, P.O.; Egwu, K.O.; Dogo, G.I.; Salami-Shinaba, J.O. Prevalence and significance of parasites of horses in some states of northern Nigeria. J. Equine Sci. 2012, 23, 1–4. [Google Scholar] [CrossRef]
- Mshelia, P.W.; Kappmeyer, L.; Johnson, W.C.; Kudi, C.A.; Oluyinka, O.O.; Balogun, E.O.; Richard, E.E.; Onoja, E.; Sears, K.P.; Ueti, M.W. Molecular detection of Theileria species and Babesia caballi from horses in Nigeria. Parasitol. Res. 2020, 119, 2955–2963. [Google Scholar] [CrossRef]
- Sunday Idoko, I.; Tirosh-Levy, S.; Leszkowicz, M.M.; Mohammed, A.B.; Sikiti, G.B.; Wesley, N.D.; Steinman, A. Genetic characterization of piroplasms in donkeys and horses from Nigeria. Animals 2020, 10, 324. [Google Scholar] [CrossRef]
- Kim, C.M.; Blanco, L.B.; Alhassan, A.; Iseki, H.; Yokoyama, N.; Xuan, X.; Igarashi, I. Diagnostic real-time PCR assay for the quantitative detection of Theileria equi from equine blood samples. Vet. Parasitol. 2008, 151, 158–163. [Google Scholar] [CrossRef]
- Dahmana, H.; Amanzougaghene, N.; Davoust, B.; Normand, T.; Carette, O.; Demoncheaux, J.P.; Mulot, B.; Fabrizy, B.; Scandola, P.; Chik, M.; et al. Great diversity of Piroplasmida in Equidae in Africa and Europe, including potential new species. Vet. Parasitol. Reg. Stud. Rep. 2019, 18, 100332. [Google Scholar] [CrossRef]
- Beugnet, F.; Moreau, Y. Babesiosis. Rev. Sci. Tech. 2015, 34, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Ord, R.L.; Lobo, C.A. Human babesiosis: Pathogens, prevalence, diagnosis and treatment. Curr. Clin. Microbiol. Rep. 2015, 2, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Westblade, L.F.; Simon, M.S.; Mathison, B.A.; Kirkman, L.A. Babesia microti: From mice to ticks to an increasing number of highly susceptible humans. J. Clin. Microbiol. 2017, 55, 2903–2912. [Google Scholar] [CrossRef]
- Yabsley, M.J.; Shock, B.C. Natural history of zoonotic Babesia: Role of wildlife reservoirs. Int. J. Parasitol. Parasites Wildl. 2013, 2, 18–31. [Google Scholar] [CrossRef]
- Dumic, I.; Patel, J.; Hart, M.; Niendorf, E.R.; Martin, S.; Ramanan, P. Splenic rupture as the first manifestation of Babesia Microti infection: Report of a case and review of literature. Am. J. Case Rep. 2018, 19, 335–341. [Google Scholar] [CrossRef]
- Dumic, I.; Madrid, C.; Rueda, P.L.; Nordstrom, C.W.; Taweesedt, P.T.; Ramanan, P. Splenic complications of Babesia microti infection in humans: A systematic review. Can. J. Infect. Dis. Med. Microbiol. 2020, 2020, 6934149. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.M.; Johnson, J.E.; Reece, R.; Mermel, L.A. Babesiosis-associated splenic rupture: Case series from a hyperendemic region. Clin. Infect. Dis. 2019, 69, 1212–1217. [Google Scholar] [CrossRef] [PubMed]
- Vermeil, C.; Menut, J.; Miegeville, M.; Cruziat, J.; Julienne, F.; Morin, O.; Roger, A.P.; Marjolet, M.; Bouillard, C. Babesiasis, pediatric malaria: Does confusion exist in Africa? Bull. Soc. Pathol. Exot. Filiales 1983, 76, 797–804. [Google Scholar]
- Akinboade, O.A.; Dipeolu, O.O.; Ogunji, F.O.; Adegoke, G.O. The parasites obtained and bacteria isolated from house rats (Rattus rattus Linnaeus, 1758) caught in human habitations in Ibadan, Nigeria. Int. J. Zoonoses 1981, 8, 26–32. [Google Scholar]
- Bock, R.; Jackson, L.; de Vos, A.; Jorgensen, W. Babesiosis of cattle. Parasitology 2004, 129, S247–S269. [Google Scholar] [CrossRef] [PubMed]
- Kuttler, K.L.; Clifford, D.J.; Touray, B.N. Prevalence of anaplasmosis and babesiosis in N’Dama cattle of the Gambia. Trop. Anim. Health Prod. 1988, 20, 37–41. [Google Scholar] [CrossRef]
- Miller, D.K.; Diall, O.; Craig, T.M.; Wagner, G.G. Serological prevalence of bovine babesiosis in Mali. Trop. Anim. Health Prod. 1984, 16, 71–77. [Google Scholar] [CrossRef]
- Ajayi, S.A.; Dipeolu, O.O. Prevalence of Anaplasma marginale, Babesia bigemina and B. bovis in Nigerian cattle using serological methods. Vet. Parasitol. 1986, 22, 147–149. [Google Scholar] [CrossRef]
- Akinboade, O.A.; Dipeolu, O.O. Comparison of blood smear and indirect fluorescent antibody techniques in detection of haemoparasite infections in trade cattle in Nigeria. Vet. Parasitol. 1984, 14, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Toure, A.; Sanogo, M.; Sghiri, A.; Sahibi, H. Diagnostic accuracy of an indirect enzyme linked immunosorbent assay (iELISA) for screening of Babesia bovis in cattle from West Africa. Life 2023, 13, 203. [Google Scholar] [CrossRef]
- Nagano, D.; Sivakumar, T.; De De Macedo, A.C.; Inpankaew, T.; Alhassan, A.; Igarashi, I.; Yokoyama, N. The genetic diversity of merozoite surface antigen 1 (MSA-1) among Babesia bovis detected from cattle populations in Thailand, Brazil and Ghana. J. Vet. Med. Sci. 2013, 75, 1463–1470. [Google Scholar] [CrossRef]
- Kirchner, M.; Brunner, A.; Edelhofer, R.; Joachim, A. Vector-borne parasites of dogs on the Islands of Cabo Verde. Wien Klin. Wochenschr. 2008, 120, 49–53. [Google Scholar] [CrossRef]
- Hirata, H.; Omobowale, T.; Adebayo, O.; Asanuma, N.; Haraguchi, A.; Murakami, Y.; Kusakisako, K.; Ikeda, K.; Asakawa, M.; Suzuki, K.; et al. Identification and phylogenetic analysis of Babesia parasites in domestic dogs in Nigeria. J. Vet. Med. Sci. 2022, 84, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Kamani, J.; Sannusi, A.; Dogo, A.G.; Tanko, J.T.; Egwu, K.O.; Tafarki, A.E.; Ogo, I.N.; Kemza, S.; Onovoh, E.; Shamaki, D.; et al. Babesia canis and Babesia rossi co-infection in an untraveled Nigerian dog. Vet. Parasitol. 2010, 173, 334–335. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Omobowale, O.; Tozuka, M.; Ohta, K.; Matsuu, A.; Nottidge, H.O.; Hirata, H.; Ikadai, H.; Oyamada, T. Molecular survey of Babesia canis in dogs in Nigeria. J. Vet. Med. Sci. 2007, 69, 1191–1193. [Google Scholar] [CrossRef] [PubMed]
- Wise, L.N.; Pelzel-McCluskey, A.M.; Mealey, R.H.; Knowles, D.P. Equine piroplasmosis. Vet. Clin. N. Am. Equine Pract. 2014, 30, 677–693. [Google Scholar] [CrossRef]
- Dipeolu, O.O.; Majaro, O.M.; Akinboade, O.A.; Nwufor, K.J. Studies on the blood parasites of pigs in Ibadan, Nigeria. Vet. Parasitol. 1982, 10, 87–90. [Google Scholar] [CrossRef]
- Permin, A.; Yelifari, L.; Bloch, P.; Steenhard, N.; Hansen, N.P.; Nansen, P. Parasites in cross-bred pigs in the Upper East region of Ghana. Vet. Parasitol. 1999, 87, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.J.; Kim, Y.H.; Oh, H.H.; Choi, U.S. First case of canine infection with Hepatozoon canis (Apicomplexa: Haemogregarinidae) in the Republic of Korea. Korean J. Parasitol. 2017, 55, 561–564. [Google Scholar] [CrossRef]
- O’Dwyer, L.H. Brazilian canine hepatozoonosis. Rev. Bras. Parasitol. Vet. 2011, 20, 181–193. [Google Scholar] [CrossRef]
- Sasaki, M.; Omobowale, O.; Ohta, K.; Tozuka, M.; Matsuu, A.; Hirata, H.; Nottidge, H.O.; Ikadai, H.; Oyamada, T. A PCR-based epidemiological survey of Hepatozoon canis in dogs in Nigeria. J. Vet. Med. Sci. 2008, 70, 743–745. [Google Scholar] [CrossRef][Green Version]
- Charrel, R.N.; Attoui, H.; Butenko, A.M.; Clegg, J.C.; Deubel, V.; Frolova, T.V.; Gould, E.A.; Gritsun, T.S.; Heinz, F.X.; Labuda, M.; et al. Tick-borne virus diseases of human interest in Europe. Clin. Microbiol. Infect. 2004, 10, 1040–1055. [Google Scholar] [CrossRef] [PubMed]
- Fillatre, P.; Revest, M.; Tattevin, P. Crimean-Congo hemorrhagic fever: An update. Med. Mal. Infect. 2019, 49, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Saluzzo, J.F.; Aubry, P.; McCormick, J.; Digoutte, J.P. Haemorrhagic fever caused by Crimean Congo haemorrhagic fever virus in Mauritania. Trans. R Soc. Trop. Med. Hyg. 1985, 79, 268. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, J.P.; LeGuenno, B.; Guillaud, M.; Wilson, M.L. A fatal case of Crimean-Congo haemorrhagic fever in Mauritania: Virological and serological evidence suggesting epidemic transmission. Trans. R Soc. Trop. Med. Hyg. 1990, 84, 573–576. [Google Scholar] [CrossRef]
- Nabeth, P.; Cheikh, D.O.; Lo, B.; Faye, O.; Vall, I.O.; Niang, M.; Wague, B.; Diop, D.; Diallo, M.; Diallo, B.; et al. Crimean-Congo hemorrhagic fever, Mauritania. Emerg. Infect. Dis. 2004, 10, 2143–2149. [Google Scholar] [CrossRef]
- Boushab, B.M.; Kelly, M.; Kebe, H.; Bollahi, M.A.; Basco, L.K. Crimean-Congo Hemorrhagic Fever, Mauritania. Emerg. Infect. Dis. 2020, 26, 817–818. [Google Scholar] [CrossRef]
- Bukbuk, D.N.; Dowall, S.D.; Lewandowski, K.; Bosworth, A.; Baba, S.S.; Varghese, A.; Watson, R.J.; Bell, A.; Atkinson, B.; Hewson, R. Serological and virological evidence of Crimean-Congo haemorrhagic fever virus circulation in the human population of Borno State, Northeastern Nigeria. PLoS Negl. Trop. Dis. 2016, 10, e0005126. [Google Scholar] [CrossRef]
- David-West, T.S.; Cooke, A.R.; David-West, A.S. Seroepidemiology of Congo virus (related to the virus of Crimean haemorrhagic fever) in Nigeria. Bull. World Health Organ. 1974, 51, 543–546. [Google Scholar]
- Nabeth, P.; Thior, M.; Faye, O.; Simon, F. Human Crimean-Congo hemorrhagic fever, Senegal. Emerg. Infect. Dis. 2004, 10, 1881–1882. [Google Scholar] [CrossRef]
- Safronetz, D.; Sacko, M.; Sogoba, N.; Rosenke, K.; Martellaro, C.; Traore, S.; Cissé, I.; Maiga, O.; Boisen, M.; Nelson, D.; et al. Vectorborne infections, Mali. Emerg. Infect. Dis. 2016, 22, 340–342. [Google Scholar] [CrossRef]
- Tarantola, A.; Nabeth, P.; Tattevin, P.; Michelet, C.; Zeller, H. Lookback exercise with imported Crimean-Congo hemorrhagic fever, Senegal and France. Emerg. Infect. Dis. 2006, 12, 1424–1426. [Google Scholar] [CrossRef]
- Baumann, J.; Knupfer, M.; Ouedraogo, J.; Traore, B.Y.; Heitzer, A.; Kane, B.; Maiga, B.; Sylla, M.; Kouriba, B.; Wölfel, R. Lassa and Crimean-Congo hemorrhagic fever viruses, Mali. Emerg. Infect. Dis. 2019, 25, 999–1002. [Google Scholar] [CrossRef]
- Umoh, J.U.; Ezeokoli, C.D.; Ogwu, D. Prevalence of antibodies to Crimean-haemorrhagic fever-Congo virus in cattle in northern Nigeria. Int. J. Zoonoses 1983, 10, 151–154. [Google Scholar] [PubMed]
- Sas, M.A.; Mertens, M.; Isselmou, E.; Reimer, N.; El Mamy, B.O.; Doumbia, B.; Groschup, M.H. Crimean-Congo hemorrhagic fever virus-specific antibody detection in cattle in Mauritania. Vector Borne Zoonotic Dis. 2017, 17, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Maiga, O.; Sas, M.A.; Rosenke, K.; Kamissoko, B.; Mertens, M.; Sogoba, N.; Traore, A.; Sangare, M.; Niang, M.; Schwan, T.G.; et al. Serosurvey of Crimean-Congo hemorrhagic fever virus in cattle, Mali, West Africa. Am. J. Trop. Med. Hyg. 2017, 96, 1341–1345. [Google Scholar] [CrossRef] [PubMed]
- Zivcec, M.; Maiga, O.; Kelly, A.; Feldmann, F.; Sogoba, N.; Schwan, T.G.; Feldmann, H.; Safronetz, D. Unique strain of Crimean-Congo hemorrhagic fever virus, Mali. Emerg. Infect. Dis. 2014, 20, 911–913. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chen, W.; Qiu, Z.; Li, Y.; Fan, J.; Wu, K.; Zhao, M.; Ding, H.; Fan, S.; Chen, J. African swine fever firus: A review. Life 2022, 12, 1255. [Google Scholar] [CrossRef]
- Blome, S.; Franzke, K.; Beer, M. African swine fever—A review of current knowledge. Virus Res. 2020, 287, 198099. [Google Scholar] [CrossRef]
- Penrith, M.L.; Vosloo, W.; Jori, F.; Bastos, A.D. African swine fever virus eradication in Africa. Virus Res. 2013, 173, 228–246. [Google Scholar] [CrossRef]
- Penrith, M.L.; Bastos, A.D.; Etter, E.M.C.; Beltran-Alcrudo, D. Epidemiology of African swine fever in Africa today: Sylvatic cycle versus socio-economic imperatives. Transbound Emerg. Dis. 2019, 66, 672–686. [Google Scholar] [CrossRef]
- Cubillos, C.; Gomez-Sebastian, S.; Moreno, N.; Nunez, M.C.; Mulumba-Mfumu, L.K.; Quembo, C.J.; Heath, L.; Etter, E.M.; Jori, F.; Escribano, J.M.; et al. African swine fever virus serodiagnosis: A general review with a focus on the analyses of African serum samples. Virus Res. 2013, 173, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Etter, E.M.; Seck, I.; Grosbois, V.; Jori, F.; Blanco, E.; Vial, L.; Akakpo, A.J.; Bada-Alhambedji, R.; Kone, P.; Roger, F.L. Seroprevalence of African swine fever in Senegal, 2006. Emerg. Infect. Dis. 2011, 17, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Kouakou, K.V.; Michaud, V.; Biego, H.G.; Gnabro, H.P.G.; Kouakou, A.V.; Mossoun, A.M.; Awuni, J.A.; Minoungou, G.L.; Aplogan, G.L.; Awoumé, F.K.; et al. African and classical swine fever situation in Ivory-Coast and neighboring countries, 2008–2013. Acta Trop. 2017, 166, 241–248. [Google Scholar] [CrossRef]
- Adedeji, A.J.; Atai, R.B.; Gyang, H.E.; Gambo, P.; Habib, M.A.; Weka, R.; Muwanika, V.B.; Masembe, C.; Luka, P.D. Live pig markets are hotspots for spread of African swine fever virus in Nigeria. Transbound Emerg. Dis. 2022, 69, e1526–e1540. [Google Scholar] [CrossRef] [PubMed]
- Asambe, A.; Sackey, A.K.B.; Tekdek, L.B. Prevalence of African swine fever virus and classical swine fever virus antibodies in pigs in Benue State, Nigeria. Trop. Anim. Health Prod. 2018, 50, 689–692. [Google Scholar] [CrossRef]
- Ohouko, O.F.H.; Koudouvo, K.; Dougnon, T.J.; Agbonon, A.; Karim, I.Y.A.; Farougou, S.; Gbeassor, M. African swine fever in Benin and prevalence of the disease in Southern Benin: A retrospective study (2014–2018). J. Adv. Vet. Anim. Res. 2020, 7, 464–470. [Google Scholar] [CrossRef]
- Minoungou, G.L.; Diop, M.; Dakouo, M.; Ouattara, A.K.; Settypalli, T.B.K.; Lo, M.M.; Sidibe, S.; Kanyala, E.; Kone, Y.S.; Diallo, M.S.; et al. Molecular characterization of African swine fever viruses in Burkina Faso, Mali, and Senegal 1989–2016: Genetic diversity of ASFV in West Africa. Transbound Emerg. Dis. 2021, 68, 2842–2852. [Google Scholar] [CrossRef]
- Sidi, M.; Zerbo, H.L.; Ouoba, B.L.; Settypalli, T.B.K.; Bazimo, G.; Ouandaogo, H.S.; Sie, B.N.; Guy, I.S.; Adama, D.D.; Savadogo, J.; et al. Molecular characterization of African swine fever viruses from Burkina Faso, 2018. BMC Vet. Res. 2022, 18, 69. [Google Scholar] [CrossRef]
- Couacy-Hymann, E.; Kouakou, K.V.; Achenbach, J.E.; Kouadio, L.; Koffi, Y.M.; Godji, H.P.; Adjé, K.E.; Oulaï, J.; Pell-Minhiaud, H.J.; Lamien, C.E. Re-emergence of genotype I of African swine fever virus in Ivory Coast. Transbound Emerg. Dis. 2019, 66, 882–896. [Google Scholar] [CrossRef]
- Spinard, E.; Rai, A.; Osei-Bonsu, J.; O’Donnell, V.; Ababio, P.T.; Tawiah-Yingar, D.; Arthur, D.; Zhou, Y.; Chepkwony, T.; Abel, L.; et al. The 2022 outbreaks of African swine fever dirus Demonstrate the first report of genotype II in Ghana. Viruses 2023, 15, 1722. [Google Scholar] [CrossRef]
- Adedeji, A.J.; Luka, P.D.; Atai, R.B.; Olubade, T.A.; Hambolu, D.A.; Ogunleye, M.A.; Muwanika, V.B.; Masembe, C. First-time presence of African swine fever virus genotype II in Nigeria. Microbiol. Resour. Announc. 2021, 10, e0035021. [Google Scholar] [CrossRef]
- Ambagala, A.; Goonewardene, K.; Lamboo, L.; Goolia, M.; Erdelyan, C.; Fisher, M.; Handel, K.; Lung, O.; Blome, S.; King, J.; et al. Characterization of a novel African swine fever virus p72 genotype II from Nigeria. Viruses 2023, 15, 915. [Google Scholar] [CrossRef]
- Awosanya, E.J.; Olugasa, B.O.; Gimba, F.I.; Sabri, M.Y.; Ogundipe, G.A. Detection of African swine fever virus in pigs in Southwest Nigeria. Vet. World 2021, 14, 1840–1845. [Google Scholar] [CrossRef]
- Luka, P.D.; Erume, J.; Yakubu, B.; Owolodun, O.A.; Shamaki, D.; Mwiine, F.N. Molecular detection of torque teno sus virus and coinfection with African swine fever virus in blood samples of pigs from some slaughterhouses in Nigeria. Adv. Virol. 2016, 2016, 6341015. [Google Scholar] [CrossRef]
- Luka, P.D.; Adedeji, A.J.; Jambol, A.R.; Ifende, I.V.; Luka, H.G.; Choji, N.D.; Weka, R.; Settypalli, T.B.K.; Achenbach, J.E.; Cattoli, G.; et al. Coinfections of African swine fever virus, porcine circovirus 2 and 3, and porcine parvovirus 1 in swine in Nigeria. Arch. Virol. 2022, 167, 2715–2722. [Google Scholar] [CrossRef] [PubMed]
- Tizhe, E.V.; Luka, P.D.; Adedeji, A.J.; Tanko, P.; Gurumyen, G.Y.; Buba, D.M.; Tizhe, U.D.; Bitrus, A.A.; Oragwa, A.O.; Shaibu, S.J.; et al. Laboratory diagnosis of a new outbreak of acute African swine fever in smallholder pig farms in Jos, Nigeria. Vet. Med. Sci. 2021, 7, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Luther, N.J.; Majiyagbe, K.A.; Shamaki, D.; Lombin, L.H.; Antiabong, J.F.; Bitrus, Y.; Owolodun, O. Detection of African swine fever virus genomic DNA in a Nigerian red river hog (Potamochoerus porcus). Vet. Rec. 2007, 160, 58–59. [Google Scholar] [CrossRef]
- Jongen, V.H.; van Roosmalen, J.; Tiems, J.; Van, H.J.; Wetsteyn, J.C. Tick-borne relapsing fever and pregnancy outcome in rural Tanzania. Acta Obstet. Gynecol. Scand. 1997, 76, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Boyer, P.H.; Lenormand, C.; Jaulhac, B.; Talagrand-Reboul, E. Human co-infections between Borrelia burgdorferi s.l. and other Ixodes-borne microorganisms: A systematic review. Pathogens 2022, 11, 282. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).