
Comprehensive Analysis of HIV-1 Integrase Resistance-Related Mutations in African Countries

 ,
,  ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Spatio-Temporal HIV-1 Subtype Epidemic Trend
2.2. Analysis of Polymorphisms and Integrase DRMs
2.3. Examination of Differences in Sequence Composition
3. Results
3.1. Distribution of Published HIV Integrase Sequences in SSA
3.2. HIV Integrase Amino Acid Diversity
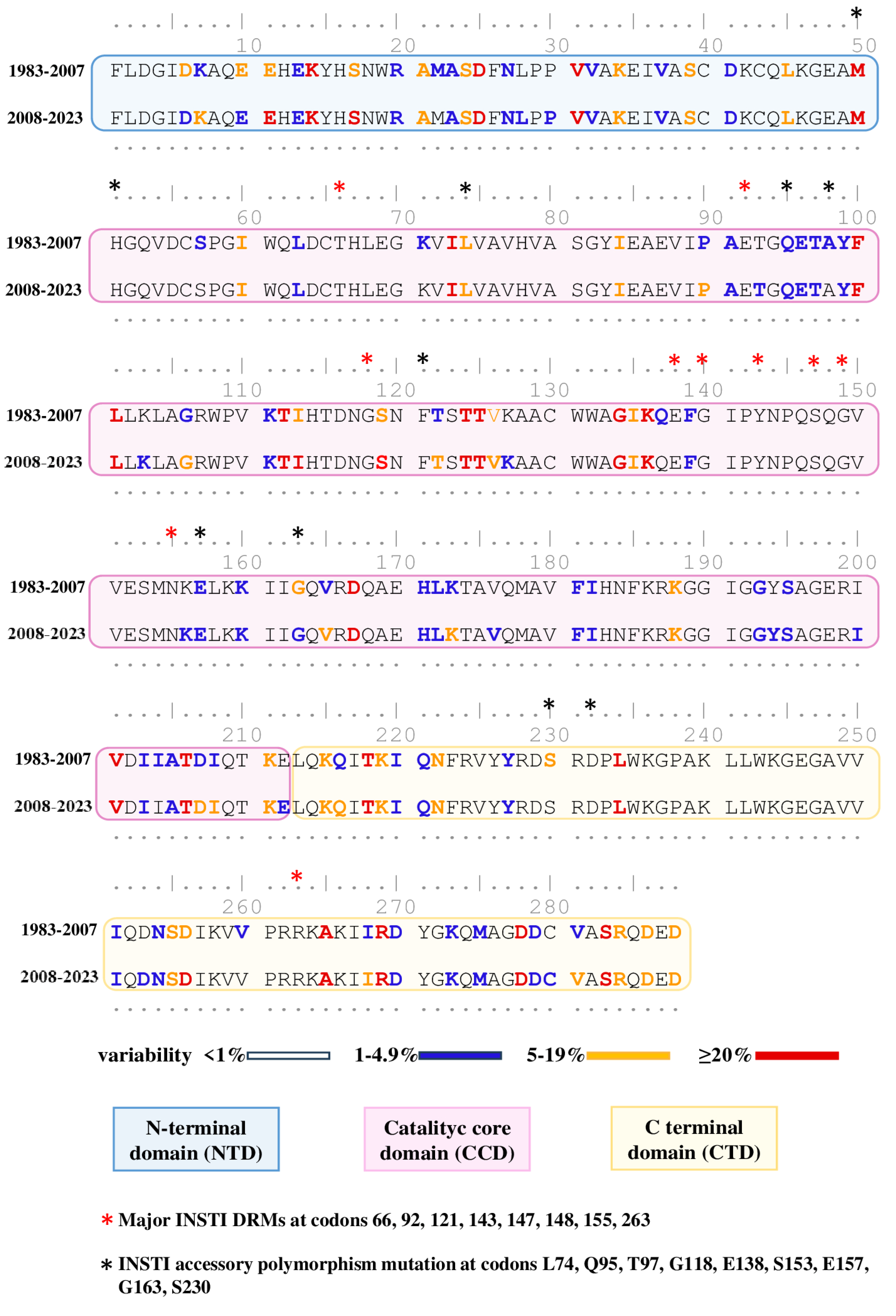
3.3. Natural Polymorphism Patterns and Conservation Analysis
3.4. Prevalence of INSTI Major DRMs
3.5. Prevalence of INSTI Accessory DRMs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Available online: https://www.afro.who.int/health-topics/hivaids (accessed on 10 October 2023).
- Global Report 2022. UNAIDS Report on the Global AIDS Epidemic 2020. UNAIDS Web Site. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 10 October 2023).
- Hemelaar, J.; Loganathan, S.; Elangovan, R.; Yun, J.; Dickson-Tetteh, L.; Kirtley, S. WHO-UNAIDS Network for HIV Country-level diversity of the HIV-1 pandemic between 1990 and 2015. J. Virol. 2020, 21, JVI.01580-20. [Google Scholar]
- Giovanetti, M.; Ciccozzi, M.; Parolin, C.; Borsetti, A. Molecular Epidemiology of HIV-1 in African Countries: A Comprehensive Overview. Pathogens 2020, 9, 1072. [Google Scholar] [CrossRef] [PubMed]
- WHO. HIV Drug Resistance Report 2021. 2021. Available online: https://www.who.int/publications/i/item/9789240038608 (accessed on 25 November 2023).
- WHO. Updated Recommendations on First-Line and Second-Line Antiretroviral Regimens and Post-Exposure Prophylaxis and Recommendations on Early Infant Diagnosis of HIV—Interim Guidance. 2018. Available online: https://www.who.int/hiv/pub/guidelines/ARV2018update/en/ (accessed on 10 October 2023).
- Zhao, A.V.; Crutchley, R.D.; Guduru, R.; Ton, K.; Lam, T.; Min, A.C. A clinical review of HIV integrase strand transfer inhibitors (INSTIs) for the prevention and treatment of HIV-1 infection. Retrovirology 2022, 19, 22. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.J.; Zhao, X.Z.; Passos, D.O.; Lyumkis, D.; Burke, T.R., Jr.; Hughes, S.H. Integrase Strand Transfer Inhibitors Are Effective Anti-HIV Drug. Viruses 2021, 13, 205. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.Y.; Liu, T.F.; Kiuchi, M.; Zioni, R.; Gifford, R.J.; Holmes, S.P.; Shafer, R.W. Natural variation of HIV-1 group M integrase: Implications for a new class of antiretroviral inhibitors. Retrovirology 2008, 5, 74. [Google Scholar] [CrossRef] [PubMed]
- Thierry, E.; Deprez, E.; Delelis, O. Different Pathways Leading to Integrase Inhibitors Resistance. Front. Microbiol. 2017, 7, 2165. [Google Scholar] [CrossRef]
- Tang, M.W.; Liu, T.F.; Shafer, R.W. The HIVdb System for HIV-1 Genotypic Resistance Interpretation. Intervirology 2012, 55, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Low, A.; Prada, N.; Topper, M.; Vaida, F.; Castor, D.; Mohri, H.; Hazuda, D.; Muesing, M.; Markowitz, M. Natural Polymorphisms of Human Immunodeficiency Virus Type 1 Integrase and Inherent Susceptibilities to a Panel of Integrase Inhibitors. Antimicrob. Agents Chemother. 2009, 53, 4275–4282. [Google Scholar] [CrossRef]
- Ngoufack Jagni Semengue, E.; Santoro, M.M.; Ndze, V.N.; Ka’e, A.C.; Yagai, B.; Nka, A.D.; Dambaya, B.; Takou, D.; Teto, G.; Fabeni, L.; et al. HIV-1 integrase resistance associated mutations and the use of dolutegravir in Sub-Saharan Africa: A systematic review and meta-analysis. PLoS Glob. Public Health 2022, 10, e0000826. [Google Scholar] [CrossRef]
- Schramm, B.; Temfack, E.; Descamps, D.; Nicholas, S.; Peytavin, G.; Bitilinyu-Bangoh, J.E.; Storto, A. Viral suppression and HIV-1 drug resistance 1 year after pragmatic transitioning to dolutegravir first-line therapy in Malawi: A prospective cohort study. Lancet HIV 2022, 9, e544–e553. [Google Scholar] [CrossRef]
- Giovanetti, M.; Farcomeni, S.; Sernicola, L.; Virtuoso, S.; Fontanelli Sulekova, L.; Maggiorella, M.T.; Buttò, S.; Taliani, G.; Ciccozzi, M.; Borsetti, A. Analysis of HIV-1 integrase genotypes and polymorphisms among integrase inhibitors-based antiretroviral treatment naïve patients in South Sudan. J. Med. Virol. 2022, 94, 3320–3327. [Google Scholar] [CrossRef] [PubMed]
- Mabeya, S.; Nyamache, A.; Ngugi, C.; Nyerere, A.; Lihana, R. Characterization of HIV-1 integrase gene and resistance associated mutations prior to roll out of integrase inhibitors by Kenyan National HIV-Treatment Program in Kenya. Ethiop. J. Health Sci. 2020, 30, 37–44. [Google Scholar] [PubMed]
- Arimide, D.A.; Szojka, Z.I.; Zealiyas, K.; Gebreegziabxier, A.; Adugna, F.; Sasinovich, S.; Björkman, P.; Medstrand, P. Pre-Treatment Integrase Inhibitor Resistance and Natural Polymorphisms among HIV-1 Subtype C Infected Patients in Ethiopia. Viruses 2022, 14, 729. [Google Scholar] [CrossRef] [PubMed]
- Semengue, E.N.J.; Armenia, D.; Inzaule, S.; Santoro, M.M.; Dambaya, B.; Takou, D.; Teto, G.; Nka, A.D.; Yagai, B.; Fabeni, L.; et al. Baseline integrase drug resistance mutations and conserved regions across HIV-1 clades in Cameroon: Implications for transition to dolutegravir in resource-limited settings. J. Antimicrob. Chemother. 2021, 76, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Mikasi, S.G.; Gichana, J.O.; van der Walt, C.; Brado, D.; Obasa, A.E.; Njenda, D.; Messembe, M.; Lyonga, E.; Assoumou, O.; Cloete, R.; et al. HIV-1 integrase diversity and resistance-associated mutations and polymorphisms among integrase strand transfer inhibitor-naive HIV-1 patients from Cameroon. AIDS Res. Hum. Retrov. 2020, 36, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Mikasi, S.G.; Isaacs, D.; Ikomey, G.M.; Shimba, H.; Cloete, R.; Jacobs, G.B. HIV-1 drug resistance mutation analyses of Cameroon-derived integrase sequences. AIDS Res. Hum. Retrov. 2021, 37, 54–56. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.S.; Mesplède, T.; Wainberg, M.A. Differences among HIV-1 subtypes in drug resistance against integrase inhibitors. Infect. Genet. Evol. 2016, 46, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Inzaule, S.C.; Hamers, R.L.; Noguera-Julian, M.; Casadellà, M.; Parera, M.; de Wit, T.F.R.; Paredes, R. Primary resistance to integrase strand transfer inhibitors in patients infected with diverse HIV-1 subtypes in sub-Saharan Africa. J. Antimicrob. Chemother. 2018, 73, 1167–1172. [Google Scholar] [CrossRef]
- Fofana, D.B.; Diarra, H.; Guindo, I.; Savadogo, M.K.; d’Almeida, M.; Diallo, F.I.; Baldé, A.; Soulié, C.; Kone, A.; Marcelin, A.-G.; et al. Prevalence of HIV-1 Natural Polymorphisms and Integrase-Resistance-Associated Mutations in African Children. Viruses 2023, 15, 546. [Google Scholar] [CrossRef]
- Update on the Transition to Dolutegravir-Based Antiretroviral Therapy: Report of a WHO Meeting. 29–30 March 2022; World Health Organization: Geneva, Switzerland, 2022.
- Kouamou, V.; Inzaule, S.; Manasa, J. Dolutegravir drug-resistance monitoring in Africa. Lancet 2021, 8, e664–e666. [Google Scholar] [CrossRef]
- Taylor, G. Rolling out HIV antiretroviral therapy in sub-Saharan Africa: 2003–2017. Can. Commun. Dis. Rep. 2018, 44, 68–70. [Google Scholar] [CrossRef] [PubMed]
- Tzou, P.L.; Rhee, S.Y.; Descamps, D.; Clutter, D.S.; Hare, B.; Mor, O.; Grude, M.; Parkin, N.; Jordan, M.R.; Bertagnolio, S.; et al. Integrase strand transfer inhibitor (INSTI)-resistance mutations for the surveillance of transmitted HIV-1 drug resistance. J. Antimicrob. Chemother. 2020, 75, 170–182. [Google Scholar] [CrossRef]
- Rhee, S.-Y.; Tzou, P.; Shafer, R. Temporal Trends in HIV-1 Mutations Used for the Surveillance of Transmitted Drug Resistance. Viruses 2021, 13, 879. [Google Scholar] [CrossRef] [PubMed]
- Depatureaux, A.; Mesplède, T.; Quashie, P.; Oliveira, M.; Moisi, D.; Plantier, J.C.; Brenner, B.; Wainberg, M.A. HIV-1 group O resistance against integrase inhibitors. J. Acquir. Immune Defic. Syndr. 2015, 70, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Lataillade, M.; Chiarella, J.; Kozal, M.J. Natural Polymorphism of the HIV-1 Integrase Gene and Mutations associated with Integrase Inhibitor Resistance. Antivir. Ther. 2007, 12, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Boerma, R.S.; Sigaloff, K.C.E.; Akanmu, A.S.; Inzaule, S.; van Hensbroek, M.B.; de Wit, T.F.R.; Calis, J.C. Alarming increase in pretreatment HIV drug resistance in children living in sub-Saharan Africa: A systematic review and meta-analysis. J. Antimicrob. Chemother. 2016, 72, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Madyadi, A.; Dhoro, M.; Shamu, T.; Washaya, T.; Kouamou, M.V.; Chimukangara, B.; Katzenstein, D.; Manasa, J. HIV-1 Genetic Diversity and Natural Polymorphisms of the Integrase Gene in Integrase Inhibitor-Naive Patients in Harare, Zimbabwe. AIDS Res. Hum. Retrov. 2021, 37, 954–961. [Google Scholar] [CrossRef]
- Ndashimye, E.; Avino, M.; Olabode, A.S.; Poon, A.F.Y.; Gibson, R.M.; Li, Y.; Meadows, A.; Tan, C.; Reyes, P.S.; Kityo, C.M.; et al. Accumulation of integrase strand transfer inhibitor resistance mutations confers high-level resistance to dolutegravir in non-B subtype HIV-1 strains from patients failing raltegravir in Uganda. J. Antimicrob. Chemother. 2020, 75, 3525–3533. [Google Scholar] [CrossRef]
- Brado, D.; Obasa, A.; Ikomey, G.M.; Cloete, R.; Singh, K.; Engelbrecht, S.; Neogi, U.; Jacobs, G.B. Analyses of HIV-1 integrase sequences prior to South African national HIV-treatment program and available of integrase inhibitors in Cape Town, South Africa. Sci. Rep. 2018, 8, 4709. [Google Scholar] [CrossRef]
- Kiros, M.; Tefera, D.A.; Andualem, H.; Geteneh, A.; Tesfaye, A.; Woldemichael, T.S.; Kidane, E.; Alemayehu, D.H.; Maier, M.; Mihret, A.; et al. Low level of HIV-1 C integrase strand transfer inhibitor resistance mutations among recently diagnosed ART-naive Ethiopians. Sci. Rep. 2023, 13, 6546. [Google Scholar] [CrossRef]
- Abdullahi, A.; Kida, I.M.; Maina, U.A.; Ibrahim, A.H.; Mshelia, J.; Wisso, H.; Adamu, A.; Onyemata, J.E.; Edun, M.; Yusuph, H.; et al. Limited emergence of resistance to integrase strand transfer inhibitors (INSTIs) in ART-experienced participants failing dolutegravir-based antiretroviral therapy: A cross-sectional analysis of a Northeast Nigerian cohort. J. Antimicrob. Chemother. 2023, 8, 2000–2007. [Google Scholar] [CrossRef] [PubMed]
- Abram, M.E.; Ram, R.R.; Margot, N.A.; Barnes, T.L.; White, K.L.; Callebaut, C.; Miller, M.D. Lack of impact of pre-existing T97A HIV-1 integrase mutation on integrase strand transfer inhibitor resistance and treatment outcome. PLoS ONE 2017, 12, e0172206. [Google Scholar] [CrossRef] [PubMed]
- Wensing, A.M.; Calvez, V.; Ceccherini-Silberstein, F.; Charpentier, C.; Günthard, H.F.; Paredes, R.; Shafer, R.W.; Richman, D.D. Update of the drug resistance mutations in HIV-1. Top. Antivir. Med. 2022, 30, 559–574. [Google Scholar] [PubMed]
- Gil, H.; Delgado, E.; Benito, S.; Moreno-Lorenzo, M.; Thomson, M.M.; Spanish Group for the Study of Antiretroviral Drug Resistance. Factors associated with HIV-1 resistance to integrase strand transfer inhibitors in Spain: Implications for dolutegravir-containing regimens. Front. Microbiol. 2022, 13, 1051096. [Google Scholar] [CrossRef] [PubMed]
- Wares, M.; Mesplède, T.; Quashie, P.K.; Osman, N.; Han, Y.; Wainberg, M.A. The M50 polymorphic substitution in association with the R263K mutation in HIV-1 subtype B integrase increases drug resistance but does not restore viral replicative fitness. Retrovirology 2014, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Venter, W.D.F.; Moorhouse, M.; Sokhela, S.; Fairlie, L.; Mashabane, N.; Masenya, M.; Serenata, C.; Akpomiemie, G.; Qavi, A.; Chandiwana, N.; et al. Dolutegravir plus two different prodrugs of tenofovir to treat HIV. N. Engl. J. Med. 2019, 381, 803–815. [Google Scholar] [CrossRef]
- El Bouzidi, K.; Kemp, S.A.; Datir, R.P.; Murtala-Ibrahim, F.; Aliyu, A.; Kwaghe, V.; Frampton, D.; Roy, S.; Breuer, J.; Sabin, A.C.; et al. High prevalence of integrase mutation L74I in West African HIV-1 subtypes prior to integrase inhibitor treatment. J. Antimicrob. Chemother. 2020, 75, 1575–1579. [Google Scholar] [CrossRef]
- Parolin, C.; Borsetti, A.; Choe, H.; Farzan, M.; Kolchinsky, P.; Heesen, M.; Ma, Q.; Gerard, C.; Palú, G.; Dorf, M.E.; et al. Use of murine CXCR-4 as a second receptor by some T-cell-tropic human immunodeficiency viruses. J. Virol. 1998, 72, 1652–1656. [Google Scholar] [CrossRef]
- Hardy, I.; Brenner, B.; Quashie, P.; Thomas, R.; Petropoulos, C.; Huang, W.; Moisi, D.; Wainberg, M.A.; Roger, M. Evolution of a novel pathway leading to dolutegravir resistance in a patient harbouring N155H and multiclass drug resistance. J. Antimicrob. Chemother. 2015, 70, 405–441. [Google Scholar] [CrossRef]
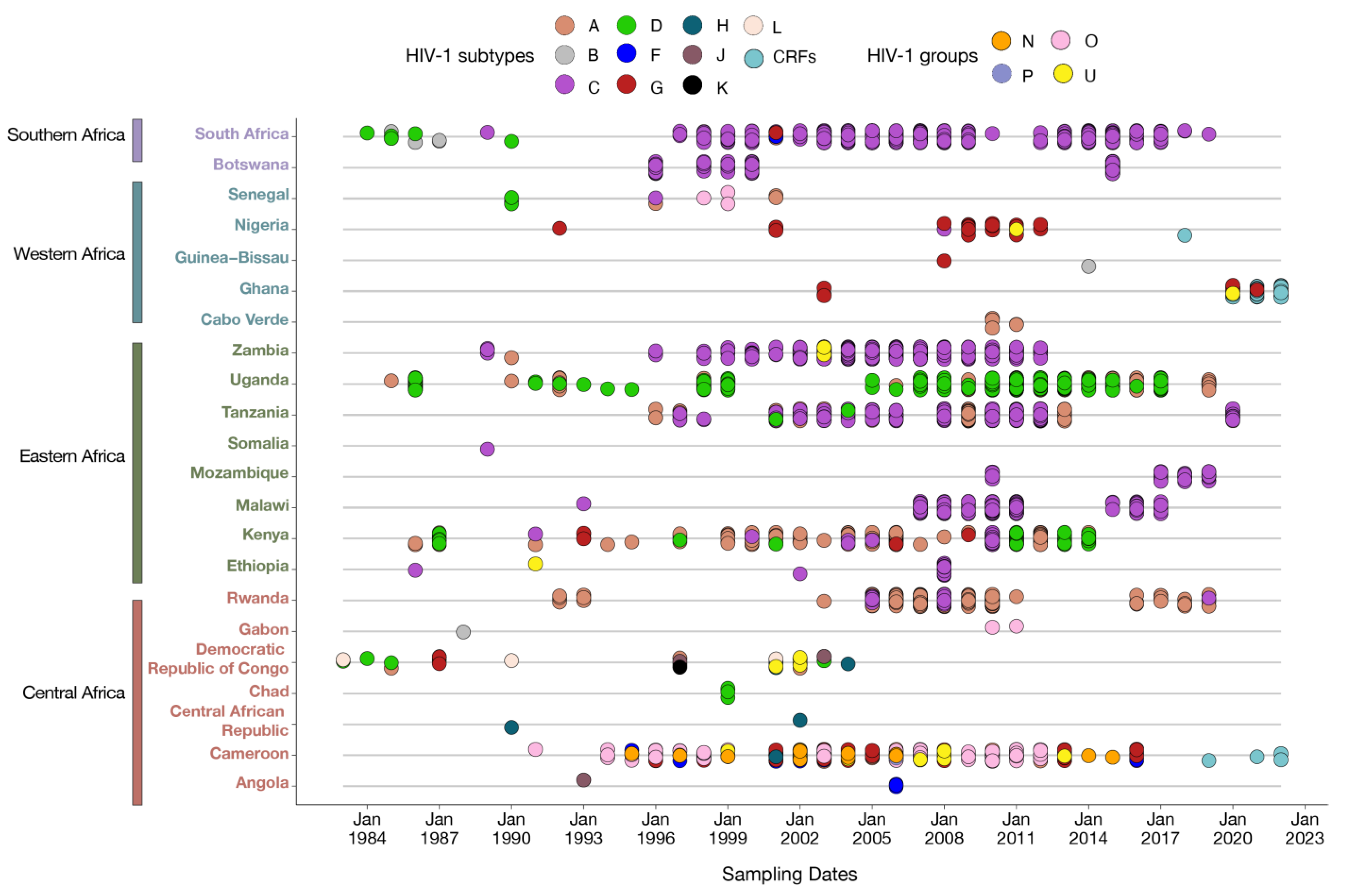

{kind=link}
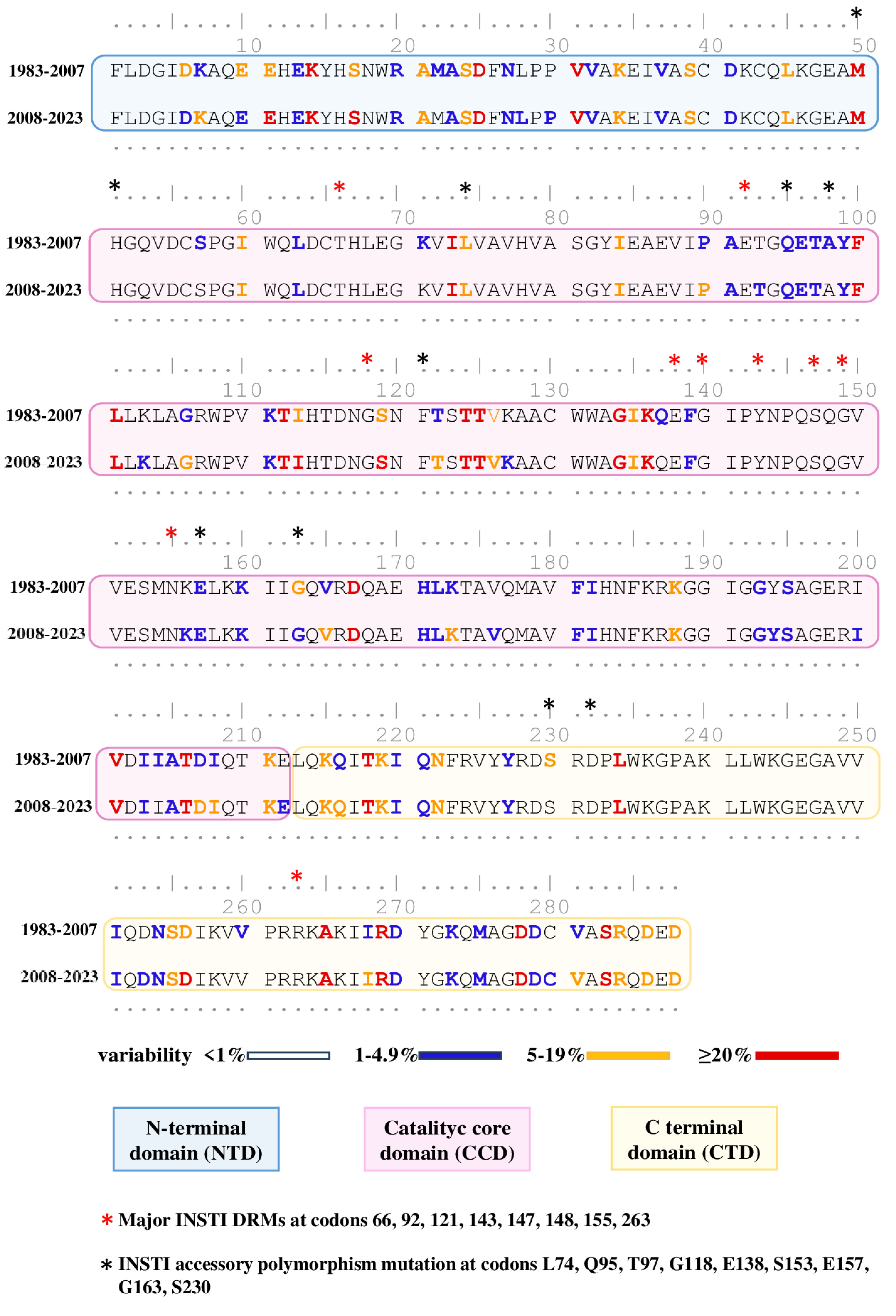{kind=link}
| Divergence | (1983–2007) | (2008–2023) | |
|---|---|---|---|
| Subtype A (618) vs. B | 0.06 ± 0.03 | Subtype A (1314) vs. B | 0.07 ± 0.04 |
| Subtype B (11) vs. B | 0.05 ± 0.03 | Subtype B (3) vs. B | 0.04 ± 0.03 |
| Subtype C (2099) vs. B | 0.06 ± 0.02 | Subtype C (2024) vs. B | 0.05 ± 0.02 |
| Subtype D (148) vs. B | 0.04 ± 0.02 | Subtype D (194) vs. B | 0.06 ± 0.05 |
| Subtype F (32) vs. B | 0.05 ± 0.02 | Subtype F (15) vs. B | 0.05 ± 0.03 |
| Subtype G (51) vs. B | 0.05 ± 0.01 | Subtype G (44) vs. B | 0.05 ± 0.02 |
| Subtypes (H, J, K, L) (17) vs. B | 0.06 ± 0.03 | Subtypes (H, J, K, L) vs. B | - |
| CRFs (453) vs. B | 0.06 ± 0.02 | CRFs (387) vs. B | 0.06 ± 0.02 |
| Groups (N, O, P, U) (62) vs. B | 0.12 ± 0.07 | Groups (N, O, P, U) (34) vs. B | 0.17 ± 0.19 |
| All groups (M, N, O, P, U) (3491) vs. B | 0.07 ± 0.04 | All groups (M, N, O, P, U) (4415) vs. B | 0.07 ± 0.04 |
| A 1983–2007 [618] | A 2008–2023 [1314] | C 1983–2007 [2099] | C 2008–2023 [2426] | D 1983–2007 [148] | CFRS 1983–2007 [453] | |
|---|---|---|---|---|---|---|
| T66A | 0.32 A(2) | 0.1 A(2) | 0.04 A(1) | 0.22 A(1) | ||
| E92G | 0.08 G(1) | |||||
| G118R | 0.12 R(3) | |||||
| E138K/A | 0.15K(2) | 0.05 A(1) | 0.04 K(1) | 0.6 K(1) | ||
| G140R | 0.12 R(1) | |||||
| Y143H | 0.1 H(2) | 0.04 H(1) | 0.68 H(1) | |||
| S147G | 0.1 G(2) | 0.04 G(1) | ||||
| Q148H | 0.05 H(1) | |||||
| N155H/T/D | 0.05 D(2) | 0.68 H(1) | 0.22 T(1) |
| A | A | C | C | D | D | F | F | G | G | HJIL | CFRS | CFRS | NOPU | NOPU | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1983- | 2008- | 1983- | 2008- | 1983- | 2008- | 1983- | 2008- | 1983- | 2008- | 1983- | 1983- | 2008- | 1983- | 2008- | |
| 2007 | 2023 | 2007 | 2023 | 2007 | 2023 | 2007 | 2023 | 2007 | 2023 | 2007 | 2007 | 2023 | 2007 | 2023 | |
| [618] | [1314] | [2099] | [2426] | [148] | [194] | [32] | [15] | [51] | [44] | [17] | [453] | [387] | [62] | [34] | |
| M50I | 18.28 | 9.36 | 47.97 | 48.8 | 0.67 | 2.58 | 18.75 | 26.6 | 11.74 | 18.18 | 17.66 | 19.9 | * 37.09 | * 85.29 | |
| I(113) | I(123) | I(1007) | I(1184) | I(1) | I(5) | I(6) | I(4) | I(6) | I(8) | I(80) | I(77) | I(23) | I(28) | ||
| L74M/I | 5.82 | 3.50 | 6.29 | 6.00 | 4.00 | 2.58 | 3.10 | 6.60 | 15.69 | 18.18 | 15.45 | 17.57 | * 59.67 | * 88.24 | |
| I(36) | I(46) | I(83) | I(146) | I(6) | I(5) | M(1) | I(1) | I(8) | I(8) | I(70) | I(68) | I(37) | I(30) | ||
| 0.97 | 1.67 | 0.76 | 0.08 | 0.67 | 1.96 | 2.27 | 6.00 | 5.42 | |||||||
| M(6) | M(22) | M(16) | M(2) | M(1) | M(1) | M(1) | M(27) | M(21) | |||||||
| Q95K | 0.32 | 0.15 | 0.14 | 0.16 | 0.51 | 2.27 | 5.88 | 0.44 | |||||||
| K(2) | K(2) | K(3) | K(4) | K(1) | K(1) | K(1) | K(2) | ||||||||
| T97A | * 15.04 | * 3.27 | 0.86 | 0.28 | 2.68 | 12.50 | 6.60 | 5.88 | 6.82 | 5.88 | 3.97 | 4.65 | 9.60 | ||
| A(93) | A(43) | A(18) | A(7) | A(4) | A(4) | A(1) | A(3) | A(3) | A(1) | A(18) | A(18) | A(6) | |||
| E157Q | 1.13 | 2.28 | 2.00 | 0.28 | 2.68 | 2.27 | 3.00 | 5.42 | |||||||
| Q(7) | Q(30) | Q(42) | Q(7) | Q(4) | Q(1) | Q(14) | Q(21) | ||||||||
| G163R/K | 0.16 | 0.07 | 0.05 | 0.16 | 0.67 | 0.25 | |||||||||
| R(1) | K(1) | R(1) | R(4) | R(1) | K(1) | ||||||||||
| 0.25 | |||||||||||||||
| R(1) | |||||||||||||||
| D232N | 0.19 | 1.35 | 0.22 | ||||||||||||
| N(4) | N(2) | N(1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Branda, F.; Giovanetti, M.; Sernicola, L.; Farcomeni, S.; Ciccozzi, M.; Borsetti, A. Comprehensive Analysis of HIV-1 Integrase Resistance-Related Mutations in African Countries. Pathogens 2024, 13, 102. https://doi.org/10.3390/pathogens13020102
Branda F, Giovanetti M, Sernicola L, Farcomeni S, Ciccozzi M, Borsetti A. Comprehensive Analysis of HIV-1 Integrase Resistance-Related Mutations in African Countries. Pathogens. 2024; 13(2):102. https://doi.org/10.3390/pathogens13020102
Chicago/Turabian StyleBranda, Francesco, Marta Giovanetti, Leonardo Sernicola, Stefania Farcomeni, Massimo Ciccozzi, and Alessandra Borsetti. 2024. "Comprehensive Analysis of HIV-1 Integrase Resistance-Related Mutations in African Countries" Pathogens 13, no. 2: 102. https://doi.org/10.3390/pathogens13020102
APA StyleBranda, F., Giovanetti, M., Sernicola, L., Farcomeni, S., Ciccozzi, M., & Borsetti, A. (2024). Comprehensive Analysis of HIV-1 Integrase Resistance-Related Mutations in African Countries. Pathogens, 13(2), 102. https://doi.org/10.3390/pathogens13020102