



Immunization with Live-Attenuated RHΔhad2a Strain Confers Partial Protective Immunity against Acute and Chronic Infection of Toxoplasma gondii in Mice
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Ethics Statement
2.2. Parasite Culture
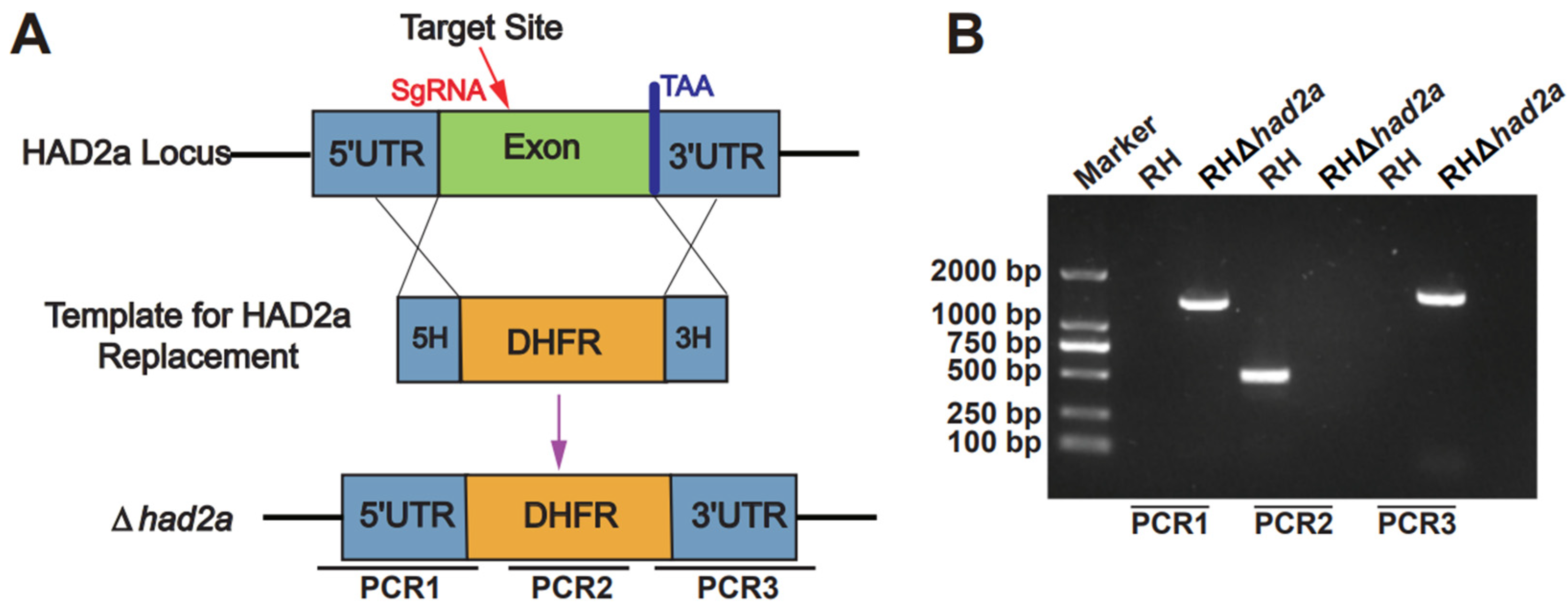
2.3. Construction of had2a Gene Knockout Strain
2.4. Immunofluorescence Assay (IFA)
2.5. Plaque Assay
2.6. Intracellular Replication Assay
2.7. Virulence Assessment in Mice
2.8. Antibodies Assessment in Immunized Mice
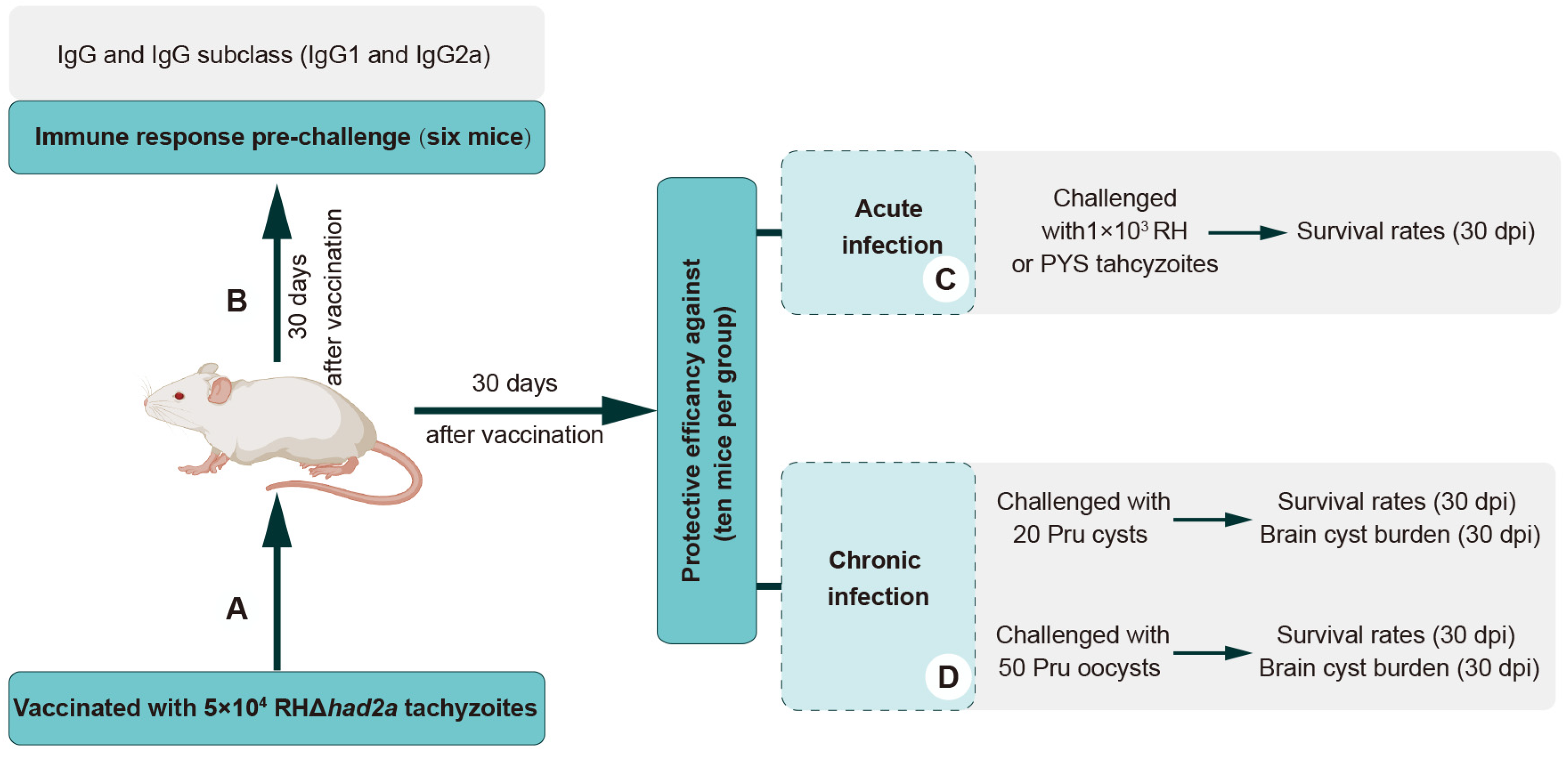
2.9. Protection of RHΔhad2a Immunization against T. gondii Infection
2.10. Statistical Analysis
3. Results
3.1. Successful Construction of RHΔhad2a
3.2. Knockout of had2a Gene Severely Attenuated T. gondii Virulence in Mice
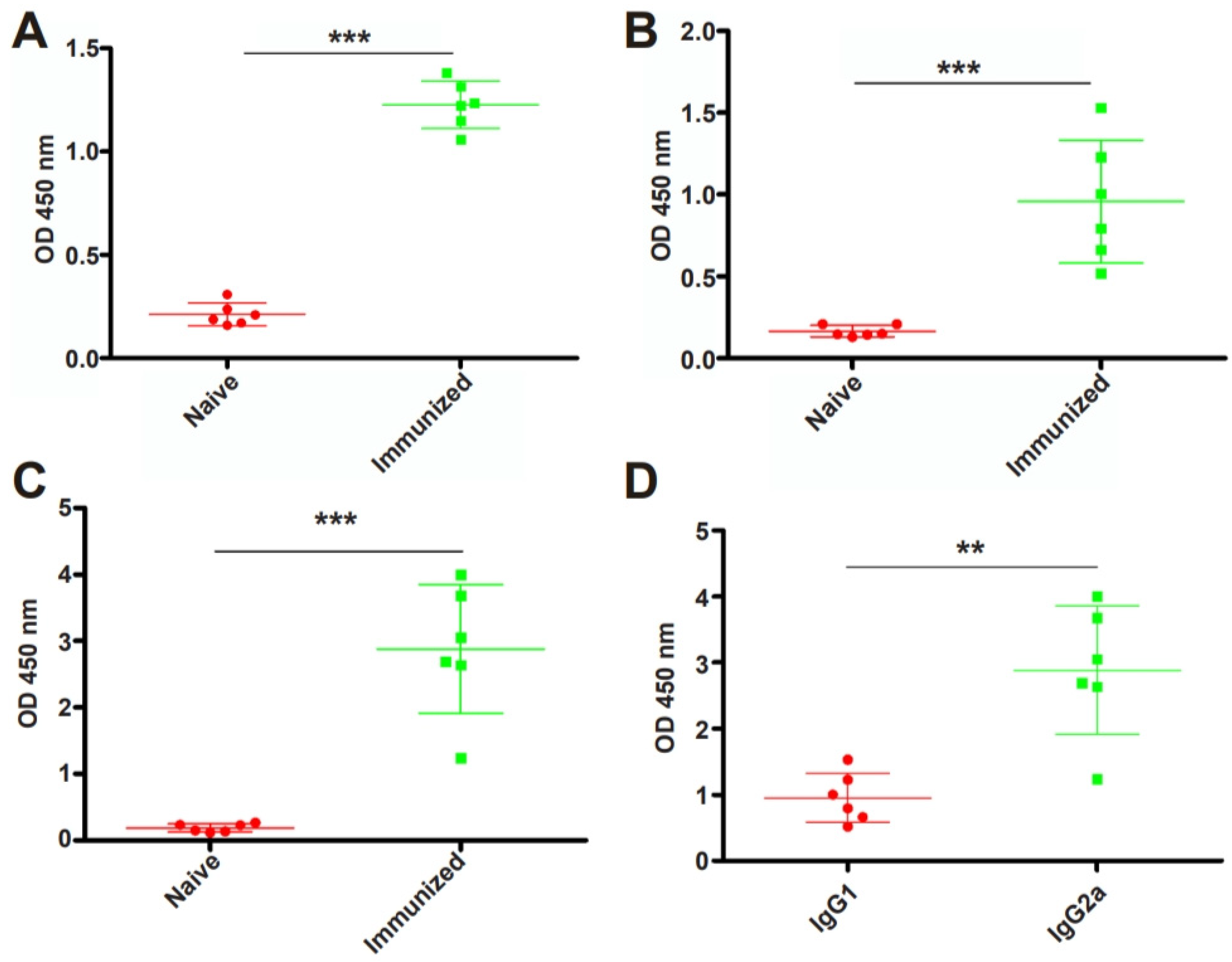
3.3. RHΔhad2a Vaccination Induces a Th1-Dominated Immune Response
3.4. Vaccination of RHΔhad2a Provides Partial Protection in Mice against Acute Infection of Virulent Strains
3.5. RHΔhad2a Vaccination Confers Partial Protection against Chronic Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Z.D.; Wang, S.C.; Liu, H.H.; Ma, H.Y.; Li, Z.Y.; Wei, F.; Zhu, X.Q.; Liu, Q. Prevalence and burden of Toxoplasma gondii infection in HIV-infected people: A systematic review and meta-analysis. Lancet HIV 2017, 4, e177–e188. [Google Scholar] [CrossRef]
- Robert-Gangneux, F.; Dardé, M.L. Epidemiology of and diagnostic strategies for toxoplasmosis. Clin. Microbiol. Rev. 2012, 25, 264–296. [Google Scholar] [CrossRef]
- Mendez, O.A.; Koshy, A.A. Toxoplasma gondii: Entry, association, and physiological influence on the central nervous system. PLoS Pathog. 2017, 13, e1006351. [Google Scholar] [CrossRef]
- Elmore, S.A.; Jones, J.L.; Conrad, P.A.; Patton, S.; Lindsay, D.S.; Dubey, J.P. Toxoplasma gondii: Epidemiology, feline clinical aspects, and prevention. Trends Parasitol. 2010, 26, 190–196. [Google Scholar] [CrossRef]
- Elsheikha, H.M.; Marra, C.M.; Zhu, X.Q. Epidemiology, pathophysiology, diagnosis, and management of cerebral toxoplasmosis. Clin. Microbiol. Rev. 2021, 34, e00115–e00119. [Google Scholar] [CrossRef]
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Pappas, G.; Roussos, N.; Falagas, M.E. Toxoplasmosis snapshots: Global status of Toxoplasma gondii seroprevalence and implications for pregnancy and congenital toxoplasmosis. Int. J. Parasitol. 2009, 39, 1385–1394. [Google Scholar] [CrossRef]
- Wang, J.L.; Zhang, N.Z.; Li, T.T.; He, J.J.; Elsheikha, H.M.; Zhu, X.Q. Advances in the development of anti-Toxoplasma gondii vaccines: Challenges, opportunities, and perspectives. Trends Parasitol. 2019, 35, 239–253. [Google Scholar] [CrossRef]
- Elsheikha, H.M. Congenital toxoplasmosis: Priorities for further health promotion action. Public Health 2008, 122, 335–353. [Google Scholar] [CrossRef]
- Buxton, D.; Innes, E.A. A commercial vaccine for ovine toxoplasmosis. Parasitology 1995, 110 Suppl, S11–S16. [Google Scholar] [CrossRef]
- Zhang, N.Z.; Chen, J.; Wang, M.; Petersen, E.; Zhu, X.Q. Vaccines against Toxoplasma gondii: New developments and perspectives. Expert. Rev. Vaccines 2013, 12, 1287–1299. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, H. Moving towards improved vaccines for Toxoplasma gondii. Expert. Opin. Biol. Ther. 2018, 18, 273–280. [Google Scholar] [CrossRef]
- Wang, J.L.; Liang, Q.L.; Li, T.T.; He, J.J.; Bai, M.J.; Cao, X.Z.; Elsheikha, H.M.; Zhu, X.Q. Toxoplasma gondii tkl1 deletion mutant is a promising vaccine against acute, chronic, and congenital toxoplasmosis in mice. J. Immunol. 2020, 204, 1562–1570. [Google Scholar] [CrossRef]
- Yang, J.; Yang, C.; Qian, J.; Li, F.; Zhao, J.; Fang, R. Toxoplasma gondii α-amylase deletion mutant is a promising vaccine against acute and chronic toxoplasmosis. Microb. Biotechnol. 2020, 13, 2057–2069. [Google Scholar] [CrossRef]
- Liang, Q.L.; Sun, L.X.; Elsheikha, H.M.; Cao, X.Z.; Nie, L.B.; Li, T.T.; Li, T.S.; Zhu, X.Q.; Wang, J.L. RHΔgra17Δnpt1 strain of Toxoplasma gondii elicits protective immunity against acute, chronic and congenital Toxoplasmosis in mice. Microorganisms 2020, 8, 352. [Google Scholar] [CrossRef]
- Shen, Y.; Zheng, B.; Sun, H.; Wu, S.; Fan, J.; Ding, J.; Gao, M.; Kong, Q.; Lou, D.; Ding, H.; et al. A live attenuated RHΔompdcΔuprt mutant of Toxoplasma gondii induces strong protective immunity against toxoplasmosis in mice and cats. Infect. Dis. Poverty 2023, 12, 60. [Google Scholar] [CrossRef]
- Wu, M.; Liu, S.; Chen, Y.; Liu, D.; An, R.; Cai, H.; Wang, J.; Zhou, N.; Obed, C.; Han, M.; et al. Live-attenuated ME49Δcdpk3 strain of Toxoplasma gondii protects against acute and chronic toxoplasmosis. NPJ Vaccines 2022, 7, 98. [Google Scholar] [CrossRef]
- Wang, J.L.; Li, T.T.; Elsheikha, H.M.; Chen, K.; Cong, W.; Yang, W.B.; Bai, M.J.; Huang, S.Y.; Zhu, X.Q. Live attenuated Pru:Δcdpk2 strain of Toxoplasma gondii protects against acute, chronic, and congenital toxoplasmosis. J. Infect. Dis. 2018, 218, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Engelberg, K.; Ivey, F.D.; Lin, A.; Kono, M.; Lorestani, A.; Faugno-Fusci, D.; Gilberger, T.W.; White, M.; Gubbels, M.J. A MORN1-associated HAD phosphatase in the basal complex is essential for Toxoplasma gondii daughter budding. Cell. Microbiol. 2016, 18, 1153–1171. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Li, T.T.; Elsheikha, H.M.; Liang, Q.L.; Zhang, Z.W.; Wang, M.; Sibley, L.D.; Zhu, X.Q. The protein phosphatase 2A holoenzyme is a key regulator of starch metabolism and bradyzoite differentiation in Toxoplasma gondii. Nat. Commun. 2022, 13, 7560. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.W.; Li, T.T.; Wang, J.L.; Liang, Q.L.; Zhang, H.S.; Sun, L.X.; Zhu, X.Q. Functional characterization of two thioredoxin proteins of Toxoplasma gondii using the CRISPR-Cas9 system. Front. Vet. Sci. 2020, 7, 614759. [Google Scholar] [CrossRef]
- Cao, X.Z.; Wang, J.L.; Elsheikha, H.M.; Li, T.T.; Sun, L.X.; Liang, Q.L.; Zhang, Z.W.; Lin, R.Q. Characterization of the role of amylo-alpha-1,6-glucosidase protein in the infectivity of Toxoplasma gondii. Front. Cell. Infect. Microbiol. 2019, 9, 418. [Google Scholar] [CrossRef]
- Zheng, X.N.; Wang, J.L.; Elsheikha, H.M.; Wang, M.; Zhang, Z.W.; Sun, L.X.; Wang, X.C.; Zhu, X.Q.; Li, T.T. Functional characterization of 15 novel dense granule proteins in Toxoplasma gondii using the CRISPR-Cas9 system. Microbiol. Spectr. 2023, 11, e0307822. [Google Scholar] [CrossRef] [PubMed]
- Li, T.T.; Zhao, D.Y.; Liang, Q.L.; Elsheikha, H.M.; Wang, M.; Sun, L.X.; Zhang, Z.W.; Chen, X.Q.; Zhu, X.Q.; Wang, J.L. The antioxidant protein glutaredoxin 1 is essential for oxidative stress response and pathogenicity of Toxoplasma gondii. FASEB J. 2023, 37, e22932. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.N.; Sun, L.X.; Elsheikha, H.M.; Li, T.T.; Gao, J.; Wu, X.J.; Zhang, Z.W.; Wang, M.; Fu, B.Q.; Zhu, X.Q.; et al. A newly characterized dense granule protein (GRA76) is important for the growth and virulence of Toxoplasma gondii. Int. J. Parasitol. 2023, 54, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Bai, M.J.; Elsheikha, H.M.; Liang, Q.L.; Li, T.T.; Cao, X.Z.; Zhu, X.Q. Novel roles of dense granule protein 12 (GRA12) in Toxoplasma gondii infection. FASEB J. 2020, 34, 3165–3178. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.L.; Nie, L.B.; Li, T.T.; Elsheikha, H.M.; Sun, L.X.; Zhang, Z.W.; Zhao, D.Y.; Zhu, X.Q.; Wang, J.L. Functional characterization of 17 protein serine/threonine phosphatases in Toxoplasma gondii using CRISPR-Cas9 system. Front. Cell Dev. Biol. 2021, 9, 738794. [Google Scholar] [CrossRef]
- Gao, J.; Wu, X.J.; Zheng, X.N.; Li, T.T.; Kou, Y.J.; Wang, X.C.; Wang, M.; Zhu, X.Q. Functional characterization of eight zinc finger motif-containing proteins in Toxoplasma gondii type I RH strain using the CRISPR-Cas9 system. Pathogens 2023, 12, 1232. [Google Scholar] [CrossRef]
- Gao, Q.; Zhang, N.Z.; Zhang, F.K.; Wang, M.; Hu, L.Y.; Zhu, X.Q. Immune response and protective effect against chronic Toxoplasma gondii infection induced by vaccination with a DNA vaccine encoding profilin. BMC Infect. Dis. 2018, 18, 117. [Google Scholar] [CrossRef]
- Villena, I.; Aubert, D.; Gomis, P.; Ferte, H.; Inglard, J.C.; Denis-Bisiaux, H.; Dondon, J.M.; Pisano, E.; Ortis, N.; Pinon, J.M. Evaluation of a strategy for Toxoplasma gondii oocyst detection in water. Appl. Environ. Microbiol. 2004, 70, 4035–4039. [Google Scholar] [CrossRef]
- Yang, W.B.; Wang, J.L.; Gui, Q.; Zou, Y.; Chen, K.; Liu, Q.; Liang, Q.L.; Zhu, X.Q.; Zhou, D.H. Immunization with a live-attenuated RH:ΔNPT1 strain of Toxoplasma gondii induces strong protective immunity against toxoplasmosis in mice. Front. Microbiol. 2019, 10, 1875. [Google Scholar] [CrossRef]
- Matta, S.K.; Rinkenberger, N.; Dunay, I.R.; Sibley, L.D. Toxoplasma gondii infection and its implications within the central nervous system. Nat. Rev. Microbiol. 2021, 19, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Dunay, I.R.; Gajurel, K.; Dhakal, R.; Liesenfeld, O.; Montoya, J.G. Treatment of toxoplasmosis: Historical perspective, animal models, and current clinical practice. Clin. Microbiol. Rev. 2018, 31, e00057-17. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Huang, S.Y.; Behnke, M.S.; Chen, K.; Shen, B.; Zhu, X.Q. The past, present, and future of genetic manipulation in Toxoplasma gondii. Trends Parasitol. 2016, 32, 542–553. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, A.M.; El-Tantawy, N.L.; Elsawey, A.M.; Abdelsalam, R.A.; Azab, M.S. The course of infection with Toxoplasma gondii RH strain in mice pre-vaccinated with gamma irradiated tachyzoites. Exp. Parasitol. 2019, 205, 107733. [Google Scholar] [CrossRef]
- Yarovinsky, F. Innate immunity to Toxoplasma gondii infection. Nat. Rev. Immunol. 2014, 14, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Pittman, K.J.; Knoll, L.J. Long-term relationships: The complicated interplay between the host and the developmental stages of Toxoplasma gondii during acute and chronic infections. Microbiol. Mol. Biol. Rev. 2015, 79, 387–401. [Google Scholar] [CrossRef]
- Sayles, P.C.; Gibson, G.W.; Johnson, L.L. B cells are essential for vaccination-induced resistance to virulent Toxoplasma gondii. Infect. Immun. 2000, 68, 1026–1033. [Google Scholar] [CrossRef]
- Tachibana, Y.; Hashizaki, E.; Sasai, M.; Yamamoto, M. Host genetics highlights IFN-γ-dependent Toxoplasma genes encoding secreted and non-secreted virulence factors in in vivo CRISPR screens. Cell Rep. 2023, 42, 112592. [Google Scholar] [CrossRef]
- Cerutti, A.; Blanchard, N.; Besteiro, S. The bradyzoite: A key developmental stage for the persistence and pathogenesis of toxoplasmosis. Pathogens 2020, 9, 234. [Google Scholar] [CrossRef]
- Jones, J.L.; Dubey, J.P. Foodborne toxoplasmosis. Clin. Infect. Dis. 2012, 55, 845–851. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | Use |
|---|---|---|
| SgHAD2a-KO-F | GCAATGTTGAAGGCGAAGCC | SgRNA of CRISPR plasmid for HAD2a knockout |
| SgRNA-R | AACTTGACATCCCCATTTAC | Construct the CRISPR plasmid for deleting HAD2a |
| SgHAD2a-KO-F | GCAATGTTGAAGGCGAAGCCGTTTTAGAGCTAGAAATAGC | Construct the CRISPR plasmid for deleting HAD2a |
| U5-HAD2a-F | GGTTTTCCCAGTCACGACGTTAACCGTCTGTGCCGGTATTG | Amplify the 5′ homologous arm of HAD2a |
| U5-HAD2a-R | GGATTTACAGCCTGGCGAAGCTTTACAAGCGGGACTTACCTTCA | Amplify the 5′ homologous arm of HAD2a |
| U3-HAD2a-F | CTATGCACTTGCAGGATGAATTCTAGAGGCACCGTTGTCAGCA | Amplify the 3′ homologous arm of HAD2a |
| U3-HAD2a-R | GAGCGGATAACAATTTCACAAAACGGCTACACTGCGTGGT | Amplify the 3′ homologous arms of HAD2a |
| DHFR-F | AAGCTTCGCCAGGCTGTAAATCC | Amplify the DHFR fragment |
| DHFR-R | GAATTCATCCTGCAAGTGCATAG | Amplify the DHFR fragment |
| pUC19-F | TGTGAAATTGTTATCCGCTC | Amplify the pUC19 fragment |
| pUC19-R | AACGTCGTGACTGGGAAAACC | Amplify the pUC19 fragment |
| HAD2a-DHFR-F | AACCGTCTGTGCCGGTATTG | Amplify the 5H-DHFR-3H fragments |
| HAD2a-DHFR-R | AAACGGCTACACTGCGTGGT | Amplify the 5H-DHFR-3H fragments |
| PCR1-HAD2a-F | AATCAACGTAGCGCTGCTTGCTTC | Detect the insertion of 5′ homologous fragment in PCR1 |
| PCR1-HAD2a-R | GCCAAAGTAGAAAGGAATTAGCAT | Detect the insertion of 5′ homologous fragment in PCR1 |
| PCR2-HAD2a-F | GACAGCACAGAGAAACCCCT | Detect the deletion of HAD2a in PCR2 |
| PCR2-HAD2a-R | GGTGGTGATTGTCACGAACG | Detect the deletion of HAD2a in PCR2 |
| PCR3-HAD2a-F | TGACGCAGATGTGCGTGTATCCAC | Detect the insertion of 3′ homologous fragment in PCR3 |
| PCR3-HAD2a-R | CACACTGAGTCAGTAGCGTGTACA | Detect the insertion of 3′ homologous fragment in PCR3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.-S.; Cao, H.; Li, C.-X.; Zhang, Z.-W.; Wang, M.; Zhu, X.-Q.; Zheng, X.-N. Immunization with Live-Attenuated RHΔhad2a Strain Confers Partial Protective Immunity against Acute and Chronic Infection of Toxoplasma gondii in Mice. Pathogens 2024, 13, 121. https://doi.org/10.3390/pathogens13020121
Zhang H-S, Cao H, Li C-X, Zhang Z-W, Wang M, Zhu X-Q, Zheng X-N. Immunization with Live-Attenuated RHΔhad2a Strain Confers Partial Protective Immunity against Acute and Chronic Infection of Toxoplasma gondii in Mice. Pathogens. 2024; 13(2):121. https://doi.org/10.3390/pathogens13020121
Chicago/Turabian StyleZhang, Hai-Sheng, Hui Cao, Chen-Xu Li, Zhi-Wei Zhang, Meng Wang, Xing-Quan Zhu, and Xiao-Nan Zheng. 2024. "Immunization with Live-Attenuated RHΔhad2a Strain Confers Partial Protective Immunity against Acute and Chronic Infection of Toxoplasma gondii in Mice" Pathogens 13, no. 2: 121. https://doi.org/10.3390/pathogens13020121
APA StyleZhang, H.-S., Cao, H., Li, C.-X., Zhang, Z.-W., Wang, M., Zhu, X.-Q., & Zheng, X.-N. (2024). Immunization with Live-Attenuated RHΔhad2a Strain Confers Partial Protective Immunity against Acute and Chronic Infection of Toxoplasma gondii in Mice. Pathogens, 13(2), 121. https://doi.org/10.3390/pathogens13020121