Nattokinase, a Subtilisin-like Alkaline-Serine Protease, Reduces Mutacin Activity by Inactivating the Competence-Stimulating Peptide in Streptococcus mutans

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Isolation and Identification of S. mutans
2.3. Confirmation of Serotypes
2.4. Arbitrarily Primed PCR
2.5. PCR Screening of Mutacin Genes
2.6. Nattokinase
2.7. Mutacin Assay
2.8. RNA Extraction
2.9. Quantitative Reverse Transcription-PCR
2.10. Construction of comC Mutant
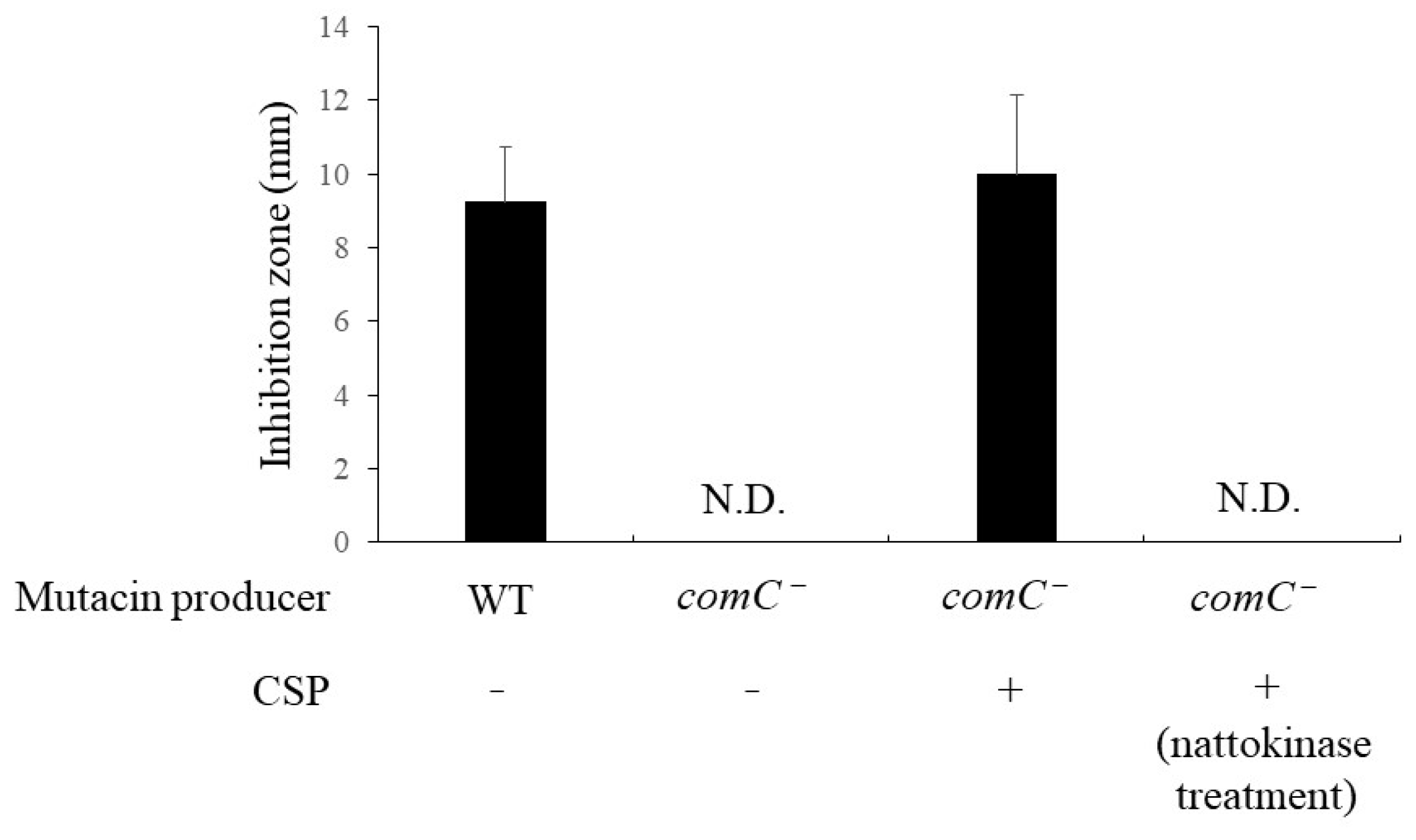
2.11. Effect of Nattokinase on CSP
3. Results
3.1. Characterization of the Clinical Isolate
3.2. Effect of Nattokinase on Mutacin Activity
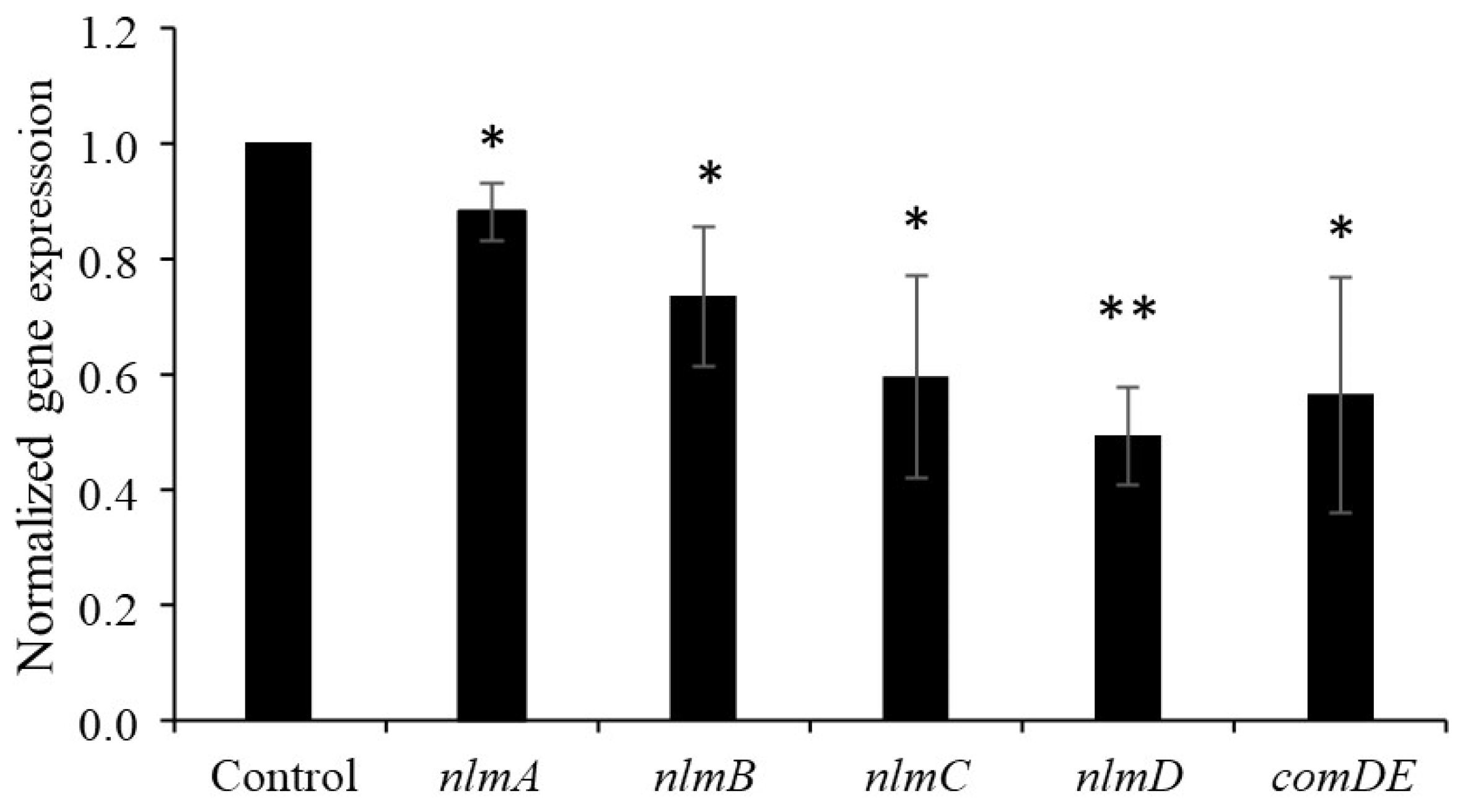
3.3. Effect of Nattokinase on Mutacin Structural Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuramitsu, H.K.; He, X.; Lux, R.; Anderson, M.H.; Shi, W. Interspecies interactions within oral microbial communities. Microbiol. Mol. Biol. Rev. 2007, 71, 653–670. [Google Scholar] [CrossRef]
- Merritt, J.; Qi, F. The mutacins of Streptococcus mutans: Regulation and ecology. Mol. Oral Microbiol. 2012, 27, 57–69. [Google Scholar] [CrossRef]
- Watanabe, A.; Kawada-Matsuo, M.; Le, M.N.-T.; Hisatsune, J.; Oogai, Y.; Nakano, Y.; Nakata, M.; Miyawaki, S.; Sugai, M.; Komatsuzawa, H. Comprehensive analysis of bacteriocins in Streptococcus mutans. Sci. Rep. 2021, 11, 12963. [Google Scholar] [CrossRef]
- Kamiya, R.U.; Höfling, J.F.; Gonçalves, R.B. Frequency and expression of mutacin biosynthesis genes in isolates of Streptococcus mutans with different mutacin-producing phenotypes. J. Med. Microbiol. 2008, 57, 626–635. [Google Scholar] [CrossRef][Green Version]
- Martin, B.; Quentin, Y.; Fichant, G.; Claverys, J.P. Independent evolution of competence regulatory cascades in streptococci? Trends Microbiol. 2006, 14, 339–345. [Google Scholar] [CrossRef]
- Cvitkovitch, D.G. Genetic competence and transformation in oral streptococci. Crit. Rev. Oral Biol. Med. 2001, 12, 217–243. [Google Scholar] [CrossRef]
- Iwamoto, A.; Nakamura, T.; Narisawa, N.; Kawasaki, Y.; Abe, S.; Torii, Y.; Senpuku, H.; Takenaga, F. The Japanese fermented food natto inhibits sucrose-dependent biofilm formation by cariogenic streptococci. Food Sci. Technol. Res. 2018, 24, 129–137. [Google Scholar] [CrossRef]
- Yokoyama, T.; Nakamura, T.; Kimijima, M.; Mandokoro, K.; Tokumaru, M.; Takatsuka, A.; Narisawa, N.; Kobayashi, R.; Takenaga, F. Subtilisin NAT, a subtilisin-like serine protease present in fermented soybean “natto” extract, inhibits Streptococcus mutans biofilm formation. Food Sci. Technol. Res. 2021, 27, 537–542. [Google Scholar] [CrossRef]
- Li, D.; Hou, L.; Hu, M.; Gao, Y.; Tian, Z.; Fan, B.; Li, S.; Wang, F. Recent advances in nattokinase-enriched fermented soybean foods: A review. Foods 2022, 11, 1867. [Google Scholar] [CrossRef]
- Chen, Z.; Saxena, D.; Caufield, P.W.; Ge, Y.; Wang, M.; Li, Y. Development of species-specific primers for detection of Streptococcus mutans in mixed bacterial samples. FEMS Microbiol. Lett. 2007, 272, 154–162. [Google Scholar] [CrossRef]
- Nakano, K.; Lapirattanakul, J.; Nomura, R.; Nemoto, H.; Alaluusua, S.; Grönroos, L.; Vaara, M.; Hamada, S.; Ooshima, T.; Nakagawa, I. Streptococcus mutans clonal variation revealed by multilocus sequence typing. J. Clin. Microbiol. 2007, 45, 2616–2625. [Google Scholar] [CrossRef]
- Valdez, R.M.A.; Duque, C.; Caiaffa, K.S.; Dos Santos, V.R.; Loesch, M.L.A.; Colombo, N.H.; Arthur, R.A.; Negrini, T.C.; Boriollo, M.F.G.; Delbem, A.C.B. Genotypic diversity and phenotypic traits of Streptococcus mutans isolates and their relation to severity of early childhood caries. BMC Oral Health 2017, 17, 115. [Google Scholar] [CrossRef]
- Tamura, S.; Yonezawa, H.; Motegi, M.; Nakao, R.; Yoneda, S.; Watanabe, H.; Yamazaki, T.; Senpuku, H. Inhibiting effects of Streptococcus salivarius on competence-stimulating peptide-dependent biofilm formation by Streptococcus mutans. Oral Microbiol. Immunol. 2009, 24, 152–161. [Google Scholar] [CrossRef]
- Vieira, J.; Messing, J. Production of single-stranded plasmid DNA. In Recombinant DNA Methodology; Wu, R., Grossman, L., Moldave, K., Eds.; Academic Press: San Diego, CA, USA, 1989; pp. 225–233. [Google Scholar]
- Shiroza, T.; Kuramitsu, H.K. Construction of a model secretion system for oral Streptococci. Infect. Immun. 1993, 61, 3745–3755. [Google Scholar] [CrossRef]
- Narisawa, N.; Kawarai, T.; Suzuki, N.; Sato, Y.; Ochiai, K.; Ohnishi, M.; Watanabe, H.; Senpuku, H. Competence-dependent endogenous DNA rearrangement and uptake of extracellular DNA give a natural variant of Streptococcus mutans without biofilm formation. J. Bacteriol. 2011, 193, 5147–5154. [Google Scholar] [CrossRef]
- Hossain, M.S.; Biswas, I. An extracelluar protease, SepM, generates functional competence-stimulating peptide in Streptococcus mutans UA159. J. Bacteriol. 2012, 194, 5886–5896. [Google Scholar] [CrossRef]
- Boles, B.R.; Thoendel, M.; Singh, P.K. Self-generated diversity produces “insurance Effects” in biofilm communities. Proc. Natl. Acad. Sci. USA 2004, 101, 16630–16635. [Google Scholar] [CrossRef]
- Boles, B.R.; Singh, P.K. Endogenous oxidative stress produces diversity and adaptability in biofilm communities. Proc. Natl. Acad. Sci. USA 2008, 105, 12503–12508. [Google Scholar] [CrossRef]
- Kreth, J.; Merritt, J.; Shi, W.; Qi, F. Co-ordinated bacteriocin production and competence development: A possible mechanism for taking up DNA from neighbouring species. Mol. Microbiol. 2005, 57, 392–404. [Google Scholar] [CrossRef]
- Kreth, J.; Merritt, J.; Zhu, L.; Shi, W.; Qi, F. Cell density- and ComE-dependent expression of a group of mutacin and mutacin-like genes in Streptococcus mutans. FEMS Microbiol. Lett. 2006, 265, 11–17. [Google Scholar] [CrossRef]
- Dufour, D.; Cordova, M.; Cvitkovitch, D.G.; Lévesque, C.M. Regulation of the competence pathway as a novel role associated with a streptococcal bacteriocin. J. Bacteriol. 2011, 193, 6552–6559. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.Y.; Deutch, A.; Hong, J.; Kuramitsu, H.K. Proteases of an early colonizer can hinder Streptococcus mutans colonization in vitro. J. Dent. Res. 2011, 90, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Jiang, W.; Zhang, M.; Zhang, L.; Shen, Y.; Huang, S.; Li, M.; Qiu, W.; Pan, Y.; Zhou, L.; et al. Affiliations expand the inhibitory effects of ficin on Streptococcus mutans biofilm formation. Biomed. Res. Int. 2021, 2021, 6692328. [Google Scholar] [CrossRef]
- Kada, S.; Ishikawa, A.; Ohshima, Y.; Yoshida, K.-I. Alkaline serine protease AprE plays an essential role in poly-γ-glutamate production during natto fermentation. Biosci. Biotechnol. Biochem. 2013, 77, 802–809. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, B.Y.; Kuramitsu, H.K. Interactions between oral bacteria: Inhibition of Streptococcus mutans bacteriocin production by Streptococcus Gordonii. Appl. Environ. Microbiol. 2005, 71, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Robson, C.L.; Wescombe, P.A.; Klesse, N.A.; Tagg, J.R. Isolation and partial characterization of the Streptococcus mutans type AII lantibiotic mutacin K8. Microbiology 2007, 153, 1631–1641. [Google Scholar] [CrossRef][Green Version]



{kind=link}
{kind=link}
| Patient No. | Isolation No. | Serotype | AP-PCR Genotype | Mutacin Genotype | Indicator Strains (mm) | ||||
|---|---|---|---|---|---|---|---|---|---|
| S. sanguinis | S. gordonii | S. mitis | S. sobrinus | S. salivarius | |||||
| C1 | 1 | c | 17 | 4 | 12.33 | 10.33 | 3.67 | N.D. | 7.00 |
| 2 | c | 17 | 4 | 2.00 | 4.33 | 3.33 | N.D. | 3.33 | |
| 3 | c | 17 | 4 | 1.33 | 4.33 | 3.50 | N.D. | 3.33 | |
| C2 | 1 | c | 20 | 4 | 3.67 | 4.33 | 3.50 | N.D. | 4.00 |
| 2 | c | 11 | 4 | 3.33 | 4.33 | 3.00 | 1.00 | 3.67 | |
| 3 | c | 18 | 4 | 3.33 | 4.33 | 3.00 | N.D. | 3.33 | |
| C3 | 1 | e | 10 | 4 | 2.33 | 4.33 | 3.33 | N.D. | 5.00 |
| 2 | e | 10 | 4 | N.D. | 4.67 | 3.50 | N.D. | 4.33 | |
| 3 | e | 10 | 4 | 1.00 | 4.00 | 3.33 | N.D. | 7.67 | |
| C4 | 1 | c | 14 | 6 | 21.67 | 15.00 | 6.67 | 4.33 | 12.00 |
| 2 | c | 13 | 6 | 21.00 | 13.33 | 11.33 | 6.67 | 11.33 | |
| 3 | c | 13 | 6 | 19.33 | 12.67 | 9.33 | 6.67 | 12.00 | |
| C5 | 1 | c | 10 | 3 | 11.67 | 11.67 | 4.67 | 1.00 | 10.67 |
| 2 | c | 10 | 3 | 13.00 | 14.00 | 4.00 | N.D. | 12.00 | |
| 3 | c | 10 | 3 | 14.33 | 11.67 | 3.00 | N.D. | 9.00 | |
| C6 | 1 | c | 16 | 1 | 2.33 | 4.00 | 3.67 | N.D. | 3.33 |
| C7 | 1 | c | 15 | 3 | 2.33 | 4.00 | 4.33 | 1.33 | 3.33 |
| 2 | c | 12 | 3 | 1.00 | 3.67 | 2.00 | N.D. | 2.00 | |
| 3 | c | 12 | 3 | N.D. | 3.67 | N.D. | N.D. | 3.33 | |
| C8 | 1 | c | 22 | 2 | 8.33 | 9.33 | 8.67 | 6.00 | 10.00 |
| 2 | c | 22 | 9 | 12.67 | 10.00 | 8.33 | 4.00 | 9.67 | |
| 3 | c | 22 | 9 | 13.67 | 10.67 | 9.33 | 4.67 | 10.00 | |
| C9 | 1 | c | 4 | 4 | 0.00 | 6.00 | 2.33 | N.D. | 4.00 |
| 2 | c | 4 | 4 | 0.00 | 6.00 | 2.00 | N.D. | 4.67 | |
| 3 | c | 5 | 4 | N.D. | 5.67 | 3.00 | N.D. | 4.00 | |
| C10 | 1 | c | 8 | 3 | 12.67 | 12.00 | 4.00 | N.D. | 10.67 |
| C11 | 1 | c | 2 | 8 | 4.33 | 5.67 | 4.33 | 1.00 | 4.00 |
| 2 | c | 1 | 8 | 10.00 | 9.00 | 6.33 | 3.67 | 7.67 | |
| 3 | c | 1 | 8 | 10.33 | 8.67 | 6.33 | 3.67 | 7.00 | |
| C12 | 1 | e | 7 | 6 | 4.67 | 6.00 | 5.00 | 1.67 | 4.00 |
| 2 | e | 7 | 6 | 2.00 | 4.00 | 3.33 | N.D. | 3.00 | |
| 3 | e | 19 | 6 | 3.33 | 3.67 | 3.67 | N.D. | 4.33 | |
| C13 | 1 | c | 11 | 8 | 7.67 | 8.67 | 8.00 | 6.33 | 6.67 |
| 2 | c | 11 | 8 | 9.33 | 8.67 | 6.00 | 3.67 | 6.00 | |
| 3 | c | 11 | 8 | 7.33 | 7.00 | 5.67 | 3.67 | 5.67 | |
| C14 | 1 | c | 23 | 6 | 2.50 | 4.00 | 3.75 | 0.75 | 3.00 |
| 2 | c | 23 | 5 | N.D. | 4.33 | N.D. | N.D. | 3.00 | |
| 3 | c | 23 | 5 | N.D. | 3.67 | N.D. | N.D. | 3.00 | |
| C15 | 1 | c/k | 24 | 3 | 10.75 | 18.00 | 4.50 | 1.50 | 11.75 |
| 2 | c | 21 | 7 | 6.50 | 7.25 | 6.00 | 4.00 | 6.00 | |
| C16 | 1 | c | 6 | 1 | 10.33 | 11.33 | 10.00 | 9.00 | 11.33 |
| 2 | c | 6 | 6 | 12.33 | 10.33 | 7.33 | 4.33 | 11.33 | |
| 3 | c | 6 | 6 | 11.33 | 10.67 | 8.33 | 5.00 | 11.33 | |
| C17 | 1 | c | 3 | 7 | 8.33 | 8.67 | 7.67 | 5.00 | 6.67 |
| 2 | c/k | 3 | 8 | 7.00 | 7.00 | 5.33 | 3.67 | 6.00 | |
| 3 | c | 3 | 8 | 6.67 | 7.33 | 5.33 | 3.67 | 6.33 | |
| UA159 | c | 9 | 4 | 13.00 | 12.67 | 4.00 | N.D. | 9.67 | |
| NBRC13955 | c | 12 | 4 | 17.33 | 22.67 | 7.00 | N.D. | 17.33 | |
| Mutacin Genotype | Positive Mutacin Genes | Number of Strains |
|---|---|---|
| 1 | mukA, smbAB, nlmABCD | 2 |
| 2 | mukA, nlmABCD | 1 |
| 3 | smbAB, nlmABCD | 8 |
| 4 | nlmABCD | 12 |
| 5 | nlmCD | 2 |
| 6 | smbAB, nlmCD | 9 |
| 7 | mukA, smbAB, nlmCD | 2 |
| 8 | mukA, nlmCD | 8 |
| 9 | muk, nlmABC | 2 |
| Patient No. | Isolation No. | Mutacin Activity to Commensal Oral Bacteria (mm) | ||||
|---|---|---|---|---|---|---|
| S. sanguinis | S. gordonii | S. mitis | S. sobrinus | S. salivarius | ||
| C1 | 1 | 9.00 | 7.33 | 2.00 | N.T. | 6.00 |
| 2 | 1.00 | 3.33 | 2.00 | N.T. | 2.67 | |
| 3 | 0.00 | 3.00 | 3.33 | N.T. | 3.33 | |
| C2 | 1 | 3.33 | 3.33 | 3.33 | N.T. | 4.00 |
| 2 | 2.00 | 3.33 | 3.33 | 0.00 | 3.67 | |
| 3 | 3.00 | 3.33 | 3.33 | N.T. | 3.67 | |
| C3 | 1 | 2.33 | 3.33 | 3.00 | N.T. | 5.33 |
| 2 | N.T. | 3.33 | 3.33 | N.T. | 4.33 | |
| 3 | 1.00 | 4.67 | 2.00 | N.T. | 7.33 | |
| C4 | 1 | 20.00 | 10.00 | 8.67 | 4.33 | 9.00 |
| 2 | 16.33 | 9.67 | 9.67 | 5.67 | 8.33 | |
| 3 | 11.33 | 10.00 | 9.00 | 6.67 | 7.33 | |
| C5 | 1 | 4.00 | 5.67 | 3.00 | 0.00 | 4.00 |
| 2 | 4.00 | 4.33 | 3.00 | N.T. | 3.67 | |
| 3 | 6.00 | 6.00 | 2.00 | N.T. | 5.00 | |
| C6 | 1 | 1.50 | 3.00 | 3.33 | N.T. | 2.00 |
| C7 | 1 | 2.00 | 3.67 | 3.67 | 0.00 | 2.00 |
| 2 | 1.00 | 3.33 | 2.00 | N.T. | 2.00 | |
| 3 | N.T. | 3.00 | 3.00 | N.T. | 2.00 | |
| C8 | 1 | 8.67 | 9.00 | 8.67 | 7.00 | 10.67 |
| 2 | 13.00 | 9.20 | 9.00 | 4.27 | 10.33 | |
| 3 | 14.10 | 9.50 | 9.30 | 4.97 | 10.67 | |
| C9 | 1 | N.T. | 4.67 | 2.00 | N.T. | 4.33 |
| 2 | N.T. | 4.67 | 2.00 | N.T. | 4.33 | |
| 3 | N.T. | 4.67 | 3.00 | N.T. | 4.33 | |
| C10 | 1 | 7.00 | 6.33 | 4.00 | N.T. | 5.67 |
| C11 | 1 | 3.67 | 5.00 | 4.00 | 1.00 | 3.67 |
| 2 | 9.00 | 8.33 | 6.33 | 4.00 | 7.00 | |
| 3 | 10.00 | 8.33 | 6.33 | 4.00 | 6.67 | |
| C12 | 1 | 3.00 | 4.67 | 4.33 | 2.00 | 4.33 |
| 2 | 2.33 | 3.33 | 3.33 | N.T. | 3.00 | |
| 3 | 3.33 | 3.33 | 3.33 | N.T. | 3.00 | |
| C13 | 1 | 6.33 | 7.67 | 6.33 | 5.33 | 5.33 |
| 2 | 8.67 | 9.00 | 6.00 | 4.00 | 6.33 | |
| 3 | 8.33 | 9.00 | 6.33 | 4.33 | 5.00 | |
| C14 | 1 | 1.50 | 3.25 | 3.75 | 0.00 | 1.75 |
| 2 | 2.00 | 3.00 | N.T. | N.T. | 2.00 | |
| 3 | N.T. | 3.33 | 3.33 | N.T. | 3.00 | |
| C15 | 1 | 5.25 | 7.75 | 4.00 | 0.75 | 6.50 |
| 2 | 4.25 | 5.75 | 5.75 | 3.75 | 4.75 | |
| C16 | 1 | 10.33 | 10.00 | 9.33 | 7.33 | 9.33 |
| 2 | 11.33 | 10.33 | 8.67 | 4.67 | 10.00 | |
| 3 | 10.67 | 9.67 | 8.00 | 4.67 | 9.33 | |
| C17 | 1 | 9.00 | 8.33 | 6.67 | 5.33 | 5.33 |
| 2 | 7.67 | 7.67 | 5.33 | 4.00 | 5.67 | |
| 3 | 7.00 | 8.30 | 5.80 | 3.25 | 5.67 | |
| UA159 | 6.67 | 5.33 | 4.00 | N.T. | 6.00 | |
| NBRC13955 | 10.67 | 10.33 | 4.67 | N.T. | 11.67 | |
| Mutacin Gene | Number of Positive/Negative Strains for Mutacin Gene | Average of Mutacin Activity against Commensal Oral Bacteria * | ||||
|---|---|---|---|---|---|---|
| S. sanguinis | S. gordonii | S. mitis | S. sobrinus | S. salivarius | ||
| nlmAB | Positive (n = 27) | 0.70 | 0.73 | 0.89 | 0.58 | 0.83 |
| single (n = 14) | 0.68 | 0.74 | 0.87 | 0.00 | 0.94 | |
| +smbAB (n = 8) | 0.57 | 0.61 | 0.85 | 0.17 | 0.57 | |
| +mukA (n = 3) | 1.03 | 0.93 | 1.03 | 1.10 | 1.07 | |
| +mukA+smbAB (n = 2) | 0.82 | 0.82 | 0.92 | 0.81 | 0.71 | |
| Negative (n = 21) | 0.88 | 0.90 | 0.99 | 0.96 | 0.85 | |
| mukA | Positive (n = 15) | 0.95 | 0.96 | 0.98 | 1.03 | 0.91 |
| single (n = 8) | 0.97 | 1.02 | 0.99 | 1.04 | 0.92 | |
| +smbAB (n = 2) | 0.87 | 0.88 | 0.91 | 1.00 | 0.80 | |
| Negative (n = 33) | 0.70 | 0.74 | 0.91 | 0.60 | 0.80 | |
| smbAB | Positive (n = 21) | 0.75 | 0.75 | 0.93 | 0.72 | 0.70 |
| Negative (n = 27) | 0.83 | 0.85 | 0.93 | 0.97 | 0.94 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kimijima, M.; Narisawa, N.; Hori, E.; Mandokoro, K.; Ito, T.; Ota, Y.; Sashida, M.; Kawai, Y.; Takenaga, F. Nattokinase, a Subtilisin-like Alkaline-Serine Protease, Reduces Mutacin Activity by Inactivating the Competence-Stimulating Peptide in Streptococcus mutans. Pathogens 2024, 13, 286. https://doi.org/10.3390/pathogens13040286
Kimijima M, Narisawa N, Hori E, Mandokoro K, Ito T, Ota Y, Sashida M, Kawai Y, Takenaga F. Nattokinase, a Subtilisin-like Alkaline-Serine Protease, Reduces Mutacin Activity by Inactivating the Competence-Stimulating Peptide in Streptococcus mutans. Pathogens. 2024; 13(4):286. https://doi.org/10.3390/pathogens13040286
Chicago/Turabian StyleKimijima, Manami, Naoki Narisawa, Eiji Hori, Kengo Mandokoro, Tatsuro Ito, Yukina Ota, Momoko Sashida, Yasushi Kawai, and Fumio Takenaga. 2024. "Nattokinase, a Subtilisin-like Alkaline-Serine Protease, Reduces Mutacin Activity by Inactivating the Competence-Stimulating Peptide in Streptococcus mutans" Pathogens 13, no. 4: 286. https://doi.org/10.3390/pathogens13040286
APA StyleKimijima, M., Narisawa, N., Hori, E., Mandokoro, K., Ito, T., Ota, Y., Sashida, M., Kawai, Y., & Takenaga, F. (2024). Nattokinase, a Subtilisin-like Alkaline-Serine Protease, Reduces Mutacin Activity by Inactivating the Competence-Stimulating Peptide in Streptococcus mutans. Pathogens, 13(4), 286. https://doi.org/10.3390/pathogens13040286