


The Diversity of Seed-Borne Fungi Associated with Soybean Grown in Southern Poland


Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Collection
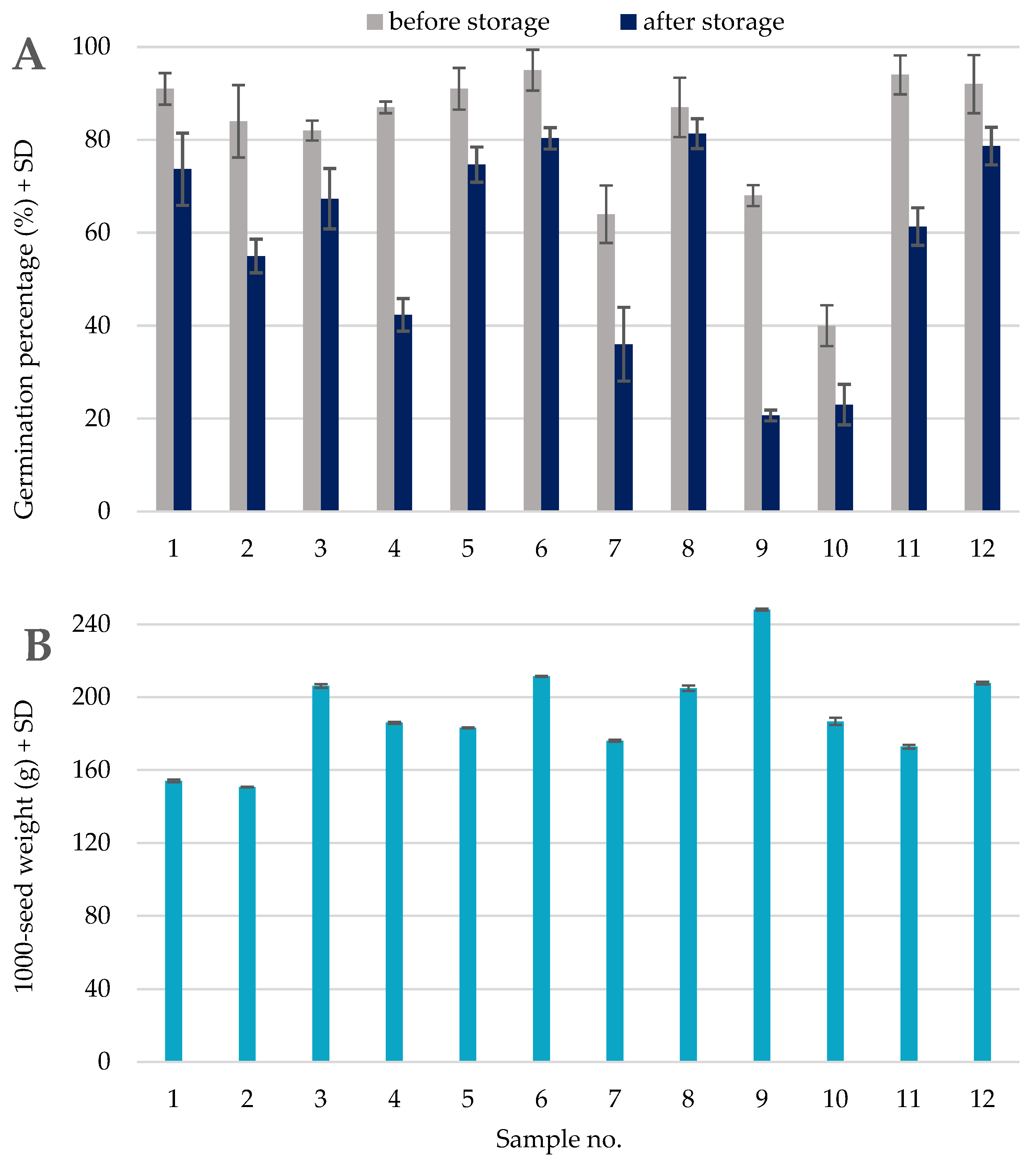
2.2. Germination Test
2.3. Isolation and Morphological Identification of Fungal Species
2.4. Molecular Species Identification of Fungal Isolates
2.5. Chemical Analysis of Soybean Seeds
2.6. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Statistic Data for Crop and Livestock and Food. Available online: https://www.fao.org/food-agriculture-statistics/data-release/en/ (accessed on 10 June 2024).
- Üstün, R.; Cat, A.; Catal, M.; Uzun, B. Identification of seedborne fungi on soybean (Glycine max. L.) seeds grown in Mediterranean region of Turkey. Turk. J. Agric. Res. 2021, 8, 367–373. [Google Scholar] [CrossRef]
- Kim, I.S.; Kim, C.H.; Yang, W.S. Physiologically active molecules and functional properties of soybean in human health—A current perspective. Int. J. Mol. Sci. 2021, 22, 4054. [Google Scholar] [CrossRef]
- Modgil, R.; Tanwar, B.; Goyal, A.; Kumar, V. Oilseeds: Health Attributes and Food Applications. Soybean (Glycine max); Springer: Singapore, 2021; pp. 1–46. [Google Scholar] [CrossRef]
- Shin, D.; Jeong, D. Korean traditional fermented soybean products: Jang. J. Ethn. Foods 2015, 2, 2–7. [Google Scholar] [CrossRef]
- Hartman, G.L.; West, E.D.; Herman, T.K. Crops that feed the world 2. Soybean-worldwide production, use, and constraints caused by pathogens and pests. Food Secur. 2011, 3, 5–17. [Google Scholar] [CrossRef]
- Roy, K.W.; Baird, R.E.; Abney, T.S. A review of soybean (Glycine max) seed, pod, and flower mycofloras in North America, with methods and a key for identification of selected fungi. Mycopathologia 2001, 150, 15–27. [Google Scholar] [CrossRef]
- Hartman, G.L.; Rupe, J.C.; Sikora, E.J.; Domier, L.L.; Davis, J.A.; Steffey, K.L. (Eds.) Compendium of Soybean Diseases and Pests, 5th ed.; APS Press: St. Paul, MN, USA, 2015. [Google Scholar]
- Christensen, C.M.; Meronuck, R.A. Quality Maintenance in Stored Grains and Seeds; University Avenue Southeast: Minneapolis, MN, USA, 1986. [Google Scholar]
- Carvalho, E.R.; Reis, L.V.; Rocha, D.K.; Penido, A.C.; da Rosa Mavaieie, D.P.; de Fátima Ferreira, V.; Oliveira, J.A. Incidence of fungal species in stored soybean seeds in relation to cooling before packing and to packing material. SCAP 2021, 44, 193–202. [Google Scholar] [CrossRef]
- Saxena, N.; Shiva Rani, S.K.; Deepika, M. Biodeterioration of soybean (Glycine max L.) seeds during storage by fungi. IJCMAS 2015, 4, 1118–1126. [Google Scholar]
- Park, J.-S.; Lee, K.-R.; Kim, J.-C.; Lim, S.-H.; Seo, J.-A.; Lee, Y.-W. A Hemorrhagic factor (Apicidin) produced by toxic Fusarium isolates from soybean seeds. Appl. Environ. Microbiol. 1999, 65, 126–130. [Google Scholar] [CrossRef]
- Barros, G.; Zanon, M.A.; Abod, A.; Oviedo, M.S.; Ramirez, M.L.; Reynoso, M.M.; Torres, A.; Chulze, S. Natural deoxynivalenol occurrence and genotype and chemotype determination of a field population of the Fusarium graminearum complex associated with soybean in Argentina. Food Addit. Contam. Part A 2012, 29, 293–303. [Google Scholar] [CrossRef]
- Olson, A.; Stenlid, J. Pathogenic fungal species hybrids infecting plants. Microbes Infect. 2002, 4, 1353–1359. [Google Scholar] [CrossRef]
- Kohn, L.M. Mechanisms of fungal speciation. Annu. Rev. Phytopathol. 2005, 43, 279–308. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L. Global species numbers of fungi: Are tropical studies and molecular approaches contributing to a more robust estimate? Biodivers Conserv. 2012, 21, 2425–2433. [Google Scholar] [CrossRef]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal identification using molecular tools: A primer for the natural products research community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Kress, W.J.; Erickson, D.L. DNA barcodes: Methods and protocols. Methods Mol. Biol. 2012, 858, 3–8. [Google Scholar] [CrossRef]
- Herbert, P.D.; Gregory, T.R. The promise of DNA barcoding for taxonomy. Syst. Biol. 2005, 54, 852–859. [Google Scholar] [CrossRef]
- Yin, G.; Zhang, Y.; Pennerman, K.K.; Wu, G.; Hua, S.S.T.; Yu, J.; Jurick, W.M.; Guo, A.; Bennett, J.W. Characterization of Blue Mold Penicillium species isolated from stored Fruits using multiple highly conserved loci. J. Fungi 2017, 3, 12. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- O’Donnell, K.; Whitaker, B.K.; Laraba, I.; Proctor, R.H.; Brown, D.W.; Broders, K.; Kim, H.-S.; McCormick, S.P.; Busman, M.; Aoki, T.; et al. DNA sequence-based identification of Fusarium: A work in progress. Plant Dis. 2022, 106, 1597–1609. [Google Scholar] [CrossRef]
- Rogério, F.; Ciampi-Guillardi, M.; Barbieri, M.C.; Braganca, C.A.; Seixas, C.D.; Almeida, A.M.; Massola, N.S., Jr. Phylogeny and variability of Colletotrichum truncatum associated with soybean anthracnose in Brasil. J. Appl. Microbiol. 2017, 122, 402–415. [Google Scholar] [CrossRef]
- Dias, M.D.; Dias-Neto, J.J.; Santos, M.D.M.; Formento, A.N.; Bizerra, L.V.A.S.; Fonseca, M.E.N.; Boiteux, L.S.; Café-Filho, A.C. Current status of soybean anthracnose associated with Colletotrichum truncatum in Brazil and Argentina. Plants 2019, 8, 459. [Google Scholar] [CrossRef]
- Cortina, J.V.; Theodoro, G.F.; Walker, D.R. Identification of fungi on diseased soybean seeds harvested during a high rainfall period in Mato Grosso Do Sul, Brazil. Biosci. J. 2013, 29, 386–391. [Google Scholar]
- Chang, X.; Li, H.; Naeem, M.; Wu, X.; Yong, T.; Song, C.; Liu, T.; Chen, W.; Yang, W. Diversity of the seedborne fungi and pathogenicity of Fusarium species associated with intercropped soybean. Pathogens 2020, 9, 531. [Google Scholar] [CrossRef] [PubMed]
- Pedrozo, R. Characterization of Soybean Seedborne Fusarium spp. in the State of Kansas. USA; Kansas State University ProQuest Dissertations Publishing: Manhattan, KS, USA, 2017; Available online: https://krex.k-state.edu/bitstreams/bdc8ecbb-b208-44f6-9adb-fe857bdf628b/download (accessed on 3 September 2024).
- Olszak-Przybyś, H.; Korbecka-Glinka, G.; Czubacka, A.; Patkowska, E. Identification of fungi inhabiting underground plant parts of soybean [Glycine max (L.) Merrill] in two developmental stages. Acta Sci. Pol. Hortorum Cultus 2021, 20, 139–149. [Google Scholar] [CrossRef]
- ISTA-International Rules for Seed Testing 2022; The International Seed Testing Association: Bassersdorf, Switzerland, 2022; Chapter 5, i-5-56(64).
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Ames, IA, USA, 2006. [Google Scholar]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi; APS Press: St. Paul, MN, USA, 1988. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Hong, S.-B.; Go, S.-J.; Shin, H.-D.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Aspergillus fumigatus and related species. Mycologia 2006, 97, 1316–1329. [Google Scholar] [CrossRef]
- Lee, S.; Yamamoto, N. Accuracy of the high-throughput amplicon sequencing to identify species within the genus Aspergillus. Fungal Biol. 2015, 119, 1311–1321. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primers sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Torres-Cruz, T.J.; Whitaker, B.; Proctor, R.H.; Laraba, I.; Kim, H.-S.; Brown, D.W.; O’Donnell, K.; Geiser, D.M. FUSARIUM-ID v.3.0: An updated downloadable resource for Fusarium species identification. Plant Dis. 2022, 106, 1610–1616. [Google Scholar] [CrossRef]
- Kjeldahl, J.A. New Method for the Determination of Nitrogen in Organic Matter. Z. Anal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef]
- Escamilla, D.; Rosso, M.L.; Zhang, B. Identification of fungi associated with soybeans and effective seed disinfection treatments. Food Sci. Nutr. 2019, 7, 3194–3205. [Google Scholar] [CrossRef]
- Santos, H.O.; Carvalho, M.L.M.; Caldeira, C.M.; Coelho, S.V.B.; Von Pinho, E.V.R.; Oliveira, J.A. Physiological and biochemical aspects of castor beans seeds deterioration stored in different packaging conditions and temperatures. J. Seed Sci. 2016, 38, 241–247. [Google Scholar] [CrossRef]
- Kurdikeri, M.B.; Basavaraj, G.T.; Hiremath, N.V.; Aswathanarayan, S.C. Storability of soybean seed under ambient condition. Kar. J. Agric. Sci. 1996, 9, 552–554. [Google Scholar]
- Rabie, M.M.; Shady, T.S.M.; El-Shazly, A.E. Effect of growth Aspergillus niger and Penicillium spp. on the oil content and fatty acids composition of stored soybean seeds. J. Agric. Sci. Mansoura Univ. 2002, 27, 6051–6062. [Google Scholar]
- Dhingra, O.; Mizubuti, E.S.G.; Napoleao, I.T.; Jham, G. Free fatty acid accumulation and quality loss of stored soybean seeds invaded by Aspergillus ruber. Seed Sci. Technol. SST 2001, 29, 193–203. [Google Scholar]
- Sweeney, M.J.; Dobson, A.D.W. Mycotoxin production by Aspergillus, Fusarium and Penicillum species. Int. J. Food. Microbiol. 1998, 43, 141–158. [Google Scholar] [CrossRef]
- Rajenda, B.; Kakde, B.; Chavan, A.M. Deteriorative changes in oilseeds due to storage fungi and efficacy of botanicals. Curr. Bot. 2001, 2, 17–22. [Google Scholar]
- Medić-Pap, S.; Milošević, M.; Jasnić, S. Soybean seed-borne fungi in the Vojvodina province. Phytopathol. Pol. 2007, 45, 55–67. [Google Scholar]
- Bhattacharya, K.; Raha, S. Deteriorative changes of maize, groundnut and soybean seeds by fungi in storage. Mycopathologia 2002, 155, 135–141. [Google Scholar] [CrossRef]
- Díaz Arias, M.M.; Leandro, L.; Munkvold, G. Aggressiveness of Fusarium species and impact of root infection on growth and yield of soybean. Phytopathology 2013, 103, 822–832. [Google Scholar] [CrossRef]
- Pereira, P.; Nesci, A.; Etcheverry, M. Effects of biocontrol agents on Fusarium verticillioides count and fumonisin content in the maize agroecosystem. Impact on rhizospheric bacterial and fungal groups. Biol. Control 2007, 42, 281–287. [Google Scholar] [CrossRef]
- Meriles, J.M.; Lamarque, A.L.; Labuckas, D.O.; Maestri, D.M. Effect of fungal damage by Fusarium spp. and Diaporthe / Phomopsis complex on protein quantity and quality of soybean seed. J. Sci. Food Agric. 2004, 84, 1594–1598. [Google Scholar] [CrossRef]
- Răut, I.; Călin, M.; Capră, L.; Gurban, A.-M.; Doni, M.; Radu, N.; Jecu, L. Cladosporium sp. isolate as fungal plant growth promoting agent. Agronomy 2021, 11, 392. [Google Scholar] [CrossRef]
- Katati, B.; van Diepeningen, A.D.; Njapau, H.; Kachapulula, P.W.; Zwaan, B.J.; Schoustra, S.E. Niche partitioning association of fungal genera correlated with lower Fusarium and fumonisin-B1 levels in maize. BioControl 2024, 69, 185–197. [Google Scholar] [CrossRef]
- Rojas, E.C.; Jensen, B.; Jorgensen, H.J.L.; Latz, M.A.C.; Esteban, P.; Ding, Y.W.; Collinge, D.B. Selection of fungal endophytes with biocontrol potential against Fusarium head blight in wheat. Biol. Control 2020, 144, 9. [Google Scholar] [CrossRef]
- Martin, I.; Galvez, L.; Guasch, L.; Palmero, D. Fungal pathogens and seed storage in the dry state. Plants 2022, 11, 3167. [Google Scholar] [CrossRef] [PubMed]
- Araujo, R.S.L.; Araujo, M.E.V.; Gomes, F.A.; Barbosa, E.G.; Teixeira, I.R.; Correa, P.C. Ultraviolet-C radiation improves soybean physiological and sanitary quality. N. Z. J. Crop Hortic. Sci. 2020, 48, 213–227. [Google Scholar] [CrossRef]
- Santos, L.A.; Faria, C.; Marek, J.; Duhatschek, E.; Martinichen, D. Radiotherapy and thermotherapy as soybean treatments. Appl. Res. Agrotechnol. 2016, 9, 37–44. [Google Scholar] [CrossRef]
- Sayahi, K.; Sari, A.H.; Hamidi, A.; Nowruzi, B.; Hassani, F. Application of cold argon plasma on germination, root length, and decontamination of soybean cultivars. BMC Plant Biol. 2024, 24, 23. [Google Scholar] [CrossRef]
- Pastucha, A. Chitosan as a compound inhibiting the occurrence of soybean diseases. Acta Sci. Pol. Hortorum Cultus 2008, 7, 41–55. [Google Scholar]
- Al Yousef, S.A. Control of seed born mycobiota associated with Glycine max. L. Merr. Seeds by a combination of traditional medicinal plants extracts. Biocell 2021, 45, 1403–1411. [Google Scholar] [CrossRef]
- Krishnamurthy, Y.L.; Shashikala, J.; Shankar Naik, B. Antifungal potential of some natural against Aspergillus flavus in soybean seeds during storage. J. Stored Prod. Res. 2008, 44, 305–309. [Google Scholar] [CrossRef]
- Parikh, L.; Agindotan, B.O.; Burrows, M.E. Antifungal activity of plant-derived essential oils on pathogens of pulse crops. Plant Dis. 2021, 105, 1692–1701. [Google Scholar] [CrossRef] [PubMed]
- Kuchlan, P.; Kuchlan, M.K.; Ansari, M.M. Efficient application of Trichoderma viride on soybean Glycine max (L.) Merrill seed using thin layer polymer coating. Legume Res. 2019, 42, 260–264. [Google Scholar] [CrossRef]
- Pimentel, M.F.; Arnao, E.; Warner, E.; Rocha, L.F.; Subedi, A.; Elsharif, N.; Chilvers, M.; Matthiesen, R.; Robertson, A.E.; Bradley, C.A.; et al. Reduction of Phytium damping-off in soybean by biocontrol seed treatment. Plant Dis. 2022, 106, 2403–2414. [Google Scholar] [CrossRef]
- Kim, J.; Roy, M.; Ahn, S.H.; Shanmugam, G.; Yang, J.S.; Jung, H.W.; Jeon, J. Culturable endophytes associated with soybean seeds and their potential for suppressing seed-borne pathogens. Plant Pathol. J. 2022, 38, 313–322. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| No. | Cultivar | Province | Location # |
|---|---|---|---|
| 1 | Lajma | Dolnośląskie | Jarząbkowice |
| 2 | Favorit | Dolnośląskie | Mierczyce |
| 3 | Kapral | Dolnośląskie | Lubin |
| 4 | Pompei | Dolnośląskie | Jemielno |
| 5 | Pompei | Dolnośląskie | Żarów |
| 6 | Orpheus | Opolskie | Buszyce |
| 7 | Madlen | Podkarpackie | Koziarnia |
| 8 | Kapral | Dolnośląskie | Mierczyce |
| 9 | Atlanta | Podkarpackie | Tarnogóra |
| 10 | Kapral | Podkarpackie | Tarnogóra |
| 11 | Bilyavka | Lubelskie | Stężyca |
| 12 | Mavka | Lubelskie | Lubycza Królewska |
| Region | Genera of Fungi Amplified with This Region | Primer | Sequence (5′-3′) | Size (bp) | PCR Protocol Used in This Study | Ref. |
|---|---|---|---|---|---|---|
| Internal transcribed spacer (ITS) | All | ITS1 ITS4 | TCCGTAGGTGAACCTGCGG TCCTCCGCTTATTGATATGC | ~600 | 94 °C—5 min; 35 cycles (94 °C—1 min, 52 °C—1 min, 72 °C—1 min); 72 °C—10 min | [22] |
| Translation elongation factor 1-alpha (TEF1) | Fusarium | EF1 EF2 | ATGGGTAAGGARGACAAGAC GGARGTACCAGTSATCATG | ~700 | 94 °C—2 min; 35 cycles (94 °C—30 s, 56 °C—90 s, 68 °C—3 min); 68 °C—5 min | [23] |
| RNA polymerase second largest subunit (RPB2) | Fusarium | 5f2 7cr | GGGGWGAYCAGAAGAAGGC CCCATRGCTTGYTTRCCCAT | ~1700 | 94 °C—2 min; 35 cycles (94 °C—30 s, 56 °C—90 s, 68 °C—3 min); 68 °C—5 min | [23] |
| Calmodulin (CaM) | Aspergillus | CMD5 CMD6 | CCGAGTACAAGGAGGCCTTC CCGATAGAGGTCATAACGTGG | ~580 | 94 °C—1 min; 42 cycles (94 °C—1 min, 55 °C—30 s, 72 °C—90 s); 72 °C—10 min | [17] |
| Beta-tubulin (BenA) | Penicillium | Bt2a Bt2b | GGTAACCAAATCGGTGCTGCTTTC ACCCTCAGTGTAGTGACCCTTGGC | ~500 | 94 °C—5 min; 35 cycles (94 °C—45 s, 55 °C—45 s, 72 °C—1 min); 72 °C—7 min | [21] |
| Fungal Genus/Species # | Top BLAST Result | Number (and %) of Isolates within Genus | Number (and %) of Isolates within Species | |
|---|---|---|---|---|
| GenBank ID * | Percent Identity | |||
| Aspergillus spp. | 86 (24.2%) | |||
| -Aspergillus pseudoglaucus Blochwitz | LT671276CaM | 100% | 53 (14.9%) | |
| -Aspergillus flavus Link | OQ181323CaM | 99.8% | 26 (7.3%) | |
| -Aspergillus niger Tiegh. | LC794815CaM | 99.6% | 2 (0.6%) | |
| -Aspergillus proliferans G. Sm. | LT671146CaM | 99.1% | 2 (0.6%) | |
| -Aspergillus montevidensis Talice & J.A. Mackinnon | OQ181343CaM | 100% | 1 (0.3%) | |
| -Aspergillus niveoglaucus Thom & Raper | LT671255CaM | 99.6% | 1 (0.3%) | |
| -Aspergillus repens (Corda) Sacc. | OR241663CaM | 100% | 1 (0.3%) | |
| Alternaria spp. | 81 (22.8%) | |||
| -Alternaria alternata (Fr.) Keissl. | MT373505ITS | 100% | 71 (20.0%) | |
| -Alternaria tenuissima (Kunze) Wiltshire | MT212224ITS | 99.8% | 3 (0.9%) | |
| -Alternaria infectoria E.G. Simmons | KR912323ITS | 99.7% | 1 (0.3%) | |
| -Alternaria spp. | 6 (1.7%) | |||
| Fusarium spp. | 58 (16.3%) | |||
| -Fusarium graminearum Schwabe | OR689618TEF1 | 98.8% | 19 (5.4%) | |
| -Fusarium flagelliforme J.W. Xia, L. Lombard, Sand.-Den., X.G. Zhang & Crous | ON292364TEF1 | 99.8% | 8 (2.3%) | |
| -Fusarium avenaceum (Fr.) Sacc. | MK185024TEF1 | 100% | 7 (2.0%) | |
| -Fusarium equiseti (Corda) Sacc. | DQ842058TEF1 | 100% | 7 (2.0%) | |
| -Fusarium tricinctum (Corda) Sacc. | MG990939TEF1 | 100% | 7 (2.0%) | |
| -Fusarium sambucinum Fuckel (syn. Fusarium cerealis) | MH582259TEF1 | 100% | 5 (1.4%) | |
| -Fusarium sporotrichioides Sherb. | MZ078869TEF1 | 100% | 3 (0.9%) | |
| -Fusarium fujikuroi Nirenberg | OR933683TEF1 | 100% | 1 (0.3%) | |
| -Fusarium redolens Wollenw. | HQ731067TEF1 | 99.3% | 1 (0.3%) | |
| Penicillium spp. | 25 (7.0%) | |||
| -Penicillium brevicompactum Dierckx | MK895703BenA | 100% | 7 (2.0%) | |
| -Penicillium aurantiogriseum Dierckx | MN031407BenA | 100% | 4 (1.1%) | |
| -Penicillium polonicum K.W. Zaleski | MK450898BenA | 100% | 4 (1.1%) | |
| -Penicillium griseofulvum Dierckx | LC682861BenA | 100% | 3 (0.9%) | |
| -Penicillium melinii Thom | KP016760BenA | 99.8% | 2 (0.6%) | |
| -Penicillium adametzii K.W. Zaleski | JN625959BenA | 100% | 1 (0.3%) | |
| -Penicillium bialowiezense K.W. Zaleski | MW980921BenA | 100% | 1 (0.3%) | |
| -Penicilium citrinum Thom | OR241785BenA | 100% | 1 (0.3%) | |
| -Penicillium freii Frisvad & Samson | OL631579BenA | 100% | 1 (0.3%) | |
| -Penicillium neoechinulatum (Frisvad, Filt. & Wicklow) Frisvad & Samson | MN969388BenA | 99.8% | 1 (0.3%) | |
| Cladosporium spp. | 21 (5.9%) | |||
| -Cladosporium cladosporioides (Fresen.) G.A. de Vries | MN966603ITS | 100% | 9 (2.5%) | |
| -Cladosporium cucumerinum Ellis & Arthur | OR008922ITS | 99.7% | 1 (0.3%) | |
| -Cladosporium pseudocladosporioides Bensch, Crous & U. Braun | MF473221ITS | 99.0% | 1 (0.3%) | |
| -Cladosporium ramotenellum K. Schub., Zalar, Crous & U. Braun | MT361323ITS | 99.9% | 1 (0.3%) | |
| -Cladosporium rectoides Bensch, H.D. Shin, Crous & U. Braun | OQ165257ITS | 99.8% | 1 (0.3%) | |
| -Cladosporium uredinicola Speg. | MW999339ITS | 99.7% | 1 (0.3%) | |
| -Cladosporium spp. | 7 (2.0%) | |||
| Botrytis spp. | 17 (4.8%) | |||
| -Botrytis cinerea Pers. | MT250963ITS | 100% | 17 (4.8%) | |
| Epicoccum spp. | 14 (3.9%) | |||
| -Epicoccum nigrum Link | MN523199ITS | 99.7% | 14 (3.9%) | |
| Diaporthe spp. | 12 (3.4%) | |||
| -Diaporthe novem J.M. Santos, Vrandečič & A.J.L. Philips | MZ066809ITS | 100% | 6 (1.7%) | |
| -Diaporthe eres Nitschke | MT573478ITS | 100% | 4 (1.1%) | |
| -Diaporthe phaseolorum (Cooke & Ellis) Sacc. (syn. Diaporthe caulivora) | HM347679ITS | 100% | 1 (0.3%) | |
| -Diaporthe sp. | HE774492ITS | 99.5% | 1 (0.3%) | |
| Rhizopus spp. | 10 (2.8%) | |||
| -Rhizopus stolonifer (Ehrenb.) Vuill. | OP437906ITS | 100% | 10 (2.8%) | |
| Sarocladium spp. | 10 (2.8%) | |||
| -Sarocladium strictum (W. Gams) Summerb. | OR346314ITS | 99.8% | 7 (2.0%) | |
| -Sarocladium mali G.Y. Sun & Y.M. Hou | MF987662ITS | 99.8% | 3 (0.9%) | |
| Stemphylium sp. | 7 (2.0%) | |||
| -Stemphylium vesicarium (Wallr.) E.G. Simmons | MT629829ITS | 100% | 7 (2.0%) | |
| Boeremia spp. | 5 (1.4%) | |||
| Boeremia exigua (Desm.) Aveskamp, Gruyter & Verkley | MT397284ITS | 99.5% | 5 (1.4%) | |
| Periconia spp. | 5 (1.4%) | |||
| -Periconia byssoides Pers | MH859902ITS | 99.6% | 1 (0.3%) | |
| -Periconia pseudobyssoides Markovsk. & A. Kačergius | LC014587ITS | 99.8% | 1 (0.3%) | |
| -Periconia spp. | 3 (0.9%) | |||
| Geomyces spp. | 2 (0.6%) | |||
| -Geomyces sp. | JX270404ITS | 99.5% | 2 (0.6%) | |
| Marquandomyces (syn. Paecilomyces) sp. | 1 (0.3%) | |||
| Marquandomyces marquandii (Massee) Samson, Houbraken & Luangsa-ard (syn. Paecilomyces marquandii) | JN545822ITS | 99.3% | 1 (0.3%) | |
| Phialophora sp. | 1 (0.3%) | |||
| -Phialophora sp. | HQ713777ITS | 99.6% | 1 (0.3%) | |
| Total | 355 | 355 | ||
| Seed Features | Free Fatty Acid Content [%] | Total Oil Content [%] | Protein Content [%] | Nitrogen Content [%] | Moisture Content [%] | Germination [%] | 1000-Seed Weight [g] |
|---|---|---|---|---|---|---|---|
| All fungi | 0.25 | 0.33 | −0.55 | −0.55 | −0.41 | 0.06 | −0.80 ** |
| Aspergillus spp. | 0.59 * | 0.33 | −0.22 | −0.22 | 0.11 | −0.32 | −0.57 |
| Aspergillus pseudoglaucus | 0.76 ** | 0.54 | −0.56 | −0.56 | −0.21 | −0.23 | −0.40 |
| Fusarium spp. | 0.18 | 0.57 | −0.81 ** | −0.81 ** | −0.51 | 0.23 | −0.43 |
| Fusarium flagelliforme | 0.01 | 0.18 | −0.59 * | −0.59 * | −0.24 | −0.27 | −0.38 |
| Penicillium spp. | 0.65 * | 0.14 | −0.41 | −0.41 | 0.40 | −0.45 | −0.35 |
| Penicillium aurantiogriseum | 0.40 | 0.12 | −0.65 * | −0.65 * | 0.29 | −0.33 | −0.50 |
| Cladosporium spp. | 0.05 | −0.03 | 0.26 | 0.26 | −0.43 | 0.70 * | −0.07 |
| Sarocladium spp. | −0.36 | −0.48 | 0.59 * | 0.59 * | 0.08 | −0.21 | 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olszak-Przybyś, H.; Korbecka-Glinka, G. The Diversity of Seed-Borne Fungi Associated with Soybean Grown in Southern Poland. Pathogens 2024, 13, 769. https://doi.org/10.3390/pathogens13090769
Olszak-Przybyś H, Korbecka-Glinka G. The Diversity of Seed-Borne Fungi Associated with Soybean Grown in Southern Poland. Pathogens. 2024; 13(9):769. https://doi.org/10.3390/pathogens13090769
Chicago/Turabian StyleOlszak-Przybyś, Hanna, and Grażyna Korbecka-Glinka. 2024. "The Diversity of Seed-Borne Fungi Associated with Soybean Grown in Southern Poland" Pathogens 13, no. 9: 769. https://doi.org/10.3390/pathogens13090769