Comparative Analysis of the Bacterial and Fungal Communities in the Gut and the Crop of Aedes albopictus Mosquitoes: A Preliminary Study


Abstract
:1. Introduction
2. Results and Discussion
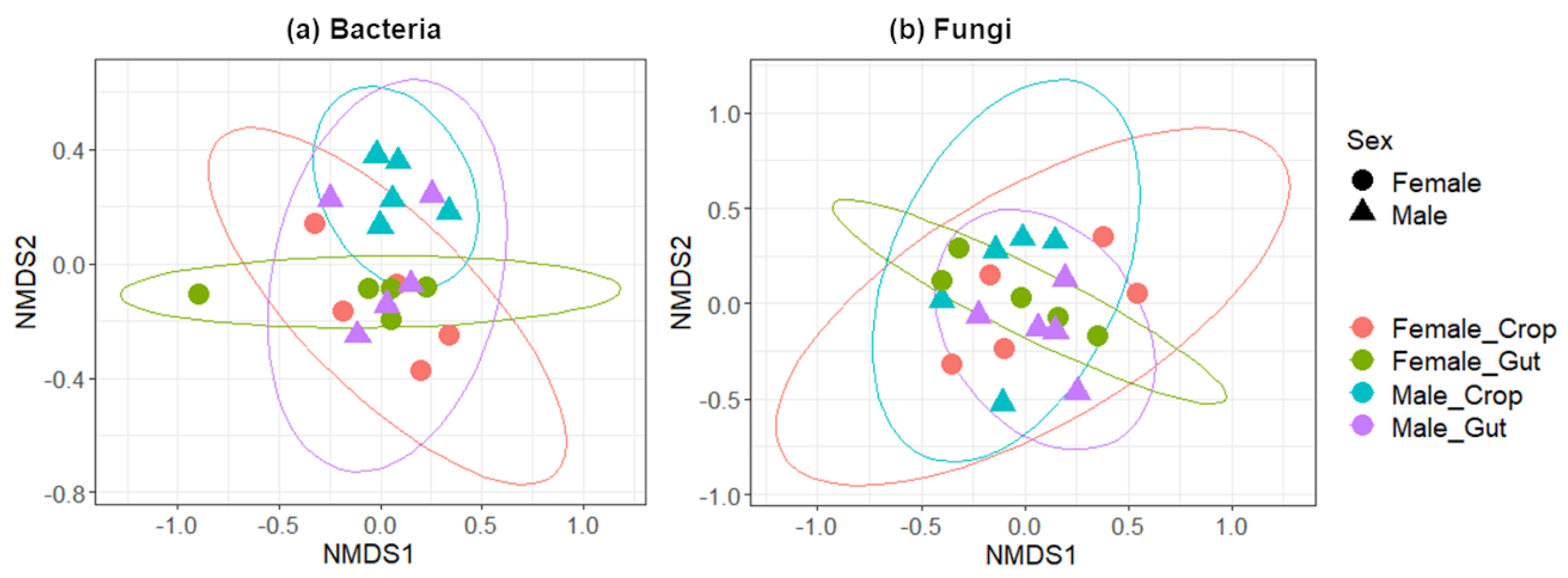
2.1. Microbial Diversity in the Gut and the Crop
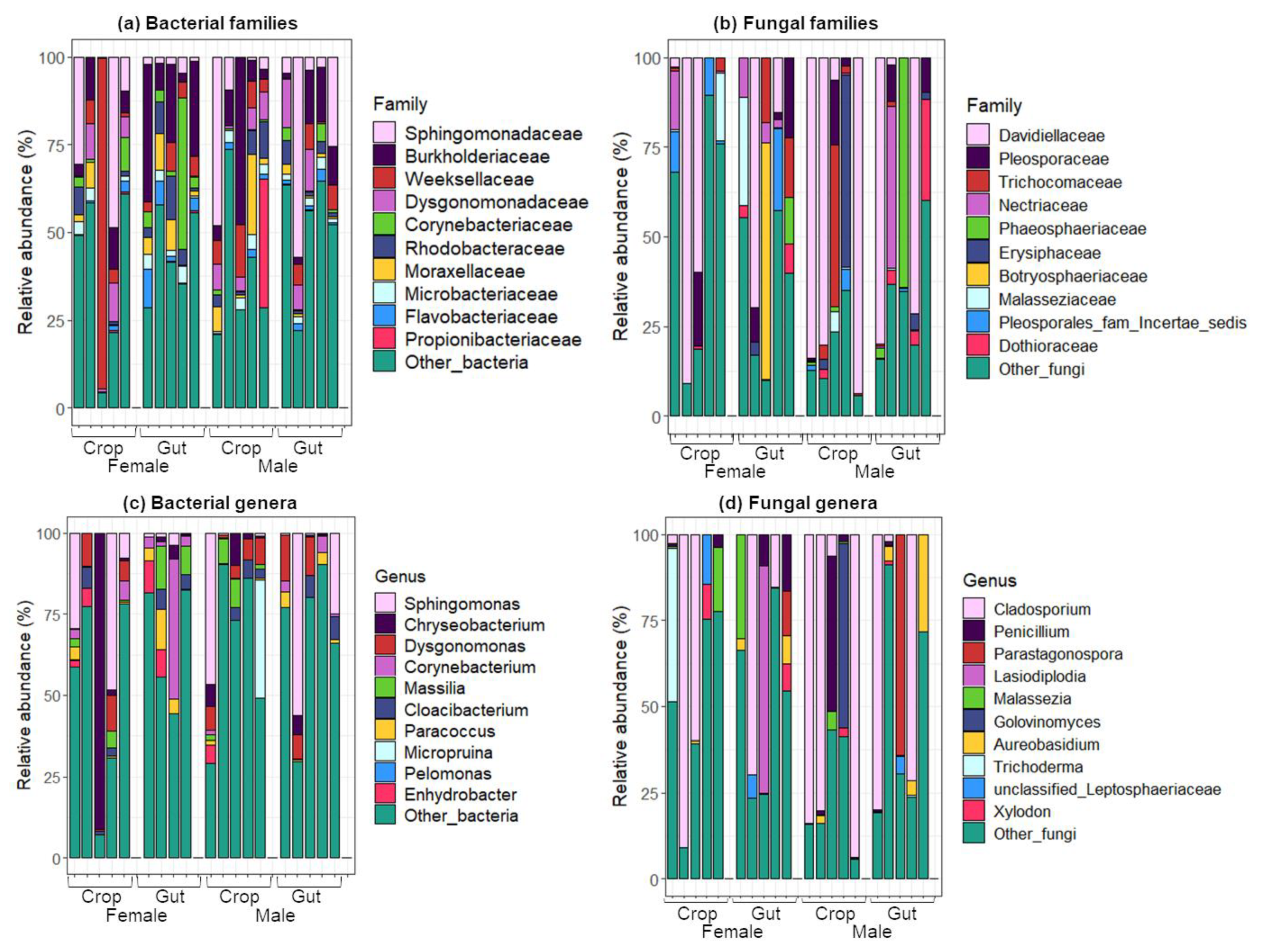
2.2. Taxonomic Composition of Bacterial and Fungal Communities in the Gut and the Crop
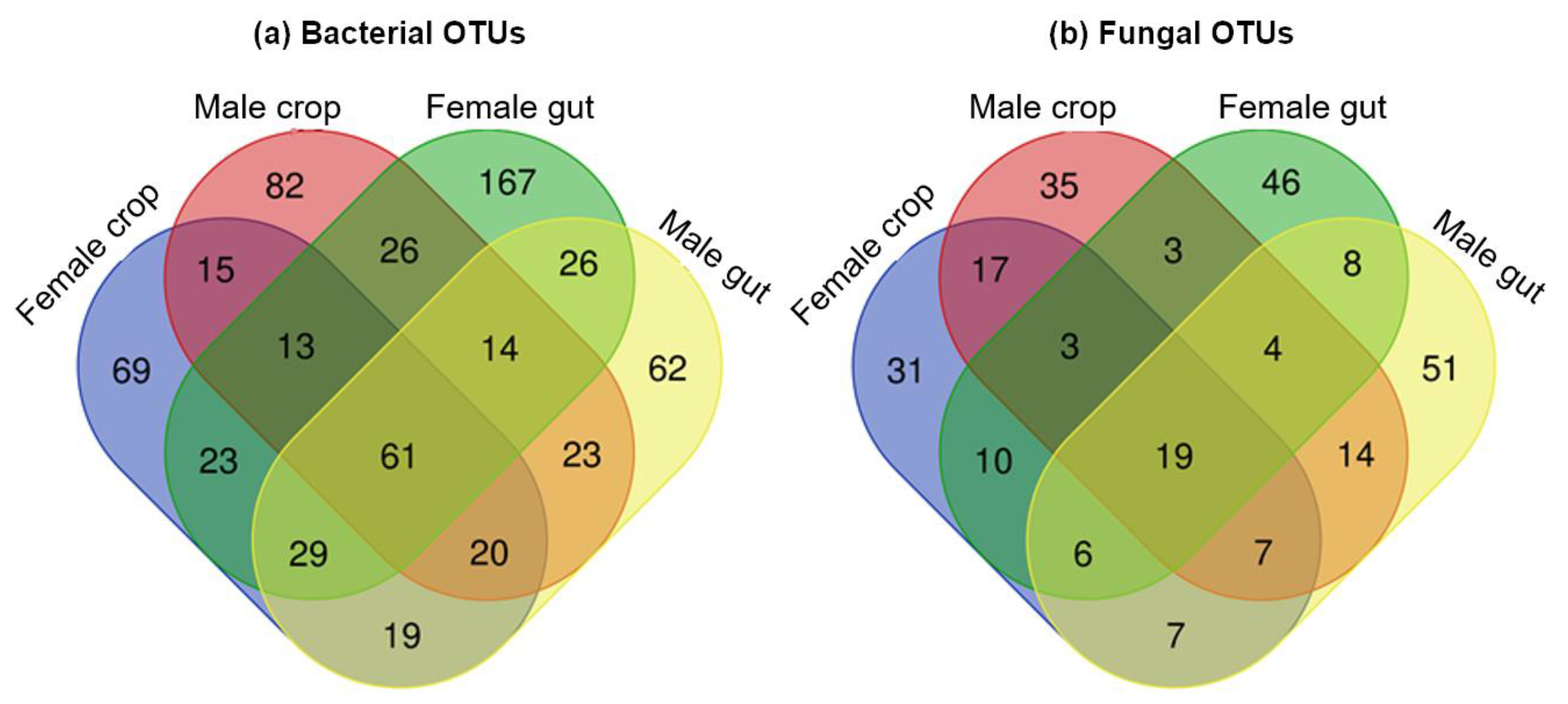
2.3. Shared Microbiota between the Gut and the Crop
3. Materials and Methods
3.1. Mosquito Colony and Rearing
3.2. DNA Extraction
3.3. DNA Library Preparation, MiSeq Illumina Sequencing and Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethical Agreement
References
- Bonizzoni, M.; Gasperi, G.; Chen, X.; James, A. The invasive mosquito species Aedes albopictus: Current knowledge and future perspectives. Trends Parasitol. 2013, 29, 460–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lwande, O.W.; Obanda, V.; Lindström, A.; Ahlm, C.; Evander, M.; Näslund, J.; Bucht, G. Globe-Trotting Aedes aegypti and Aedes albopictus: Risk Factors for Arbovirus Pandemics. Vector-Borne Zoonotic Dis. 2020, 20, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Global Invasive Species Database. 100 of the World’s Worst Invasive Alien Species. Available online: http://www.iucngisd.org/gisd/100_worst.php (accessed on 24 June 2020).
- Sherpa, S.; Blum, M.G.B.; Després, L. Cold adaptation in the Asian tiger mosquito’s native range precedes its invasion success in temperate regions. Evolution 2019, 73, 1793–1808. [Google Scholar] [CrossRef] [PubMed]
- Minard, G.; Mavingui, P.; Moro, C.V. Diversity and function of bacterial microbiota in the mosquito holobiont. Parasites Vectors 2013, 6, 146. [Google Scholar] [CrossRef] [Green Version]
- Dennison, N.J.; Jupatanakul, N.; Dimopoulos, G. The mosquito microbiota influences vector competence for human pathogens. Curr. Opin. Insect Sci. 2014, 3, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Guégan, M.; Zouache, K.; Démichel, C.; Minard, G.; Van, V.T.; Potier, P.; Mavingui, P.; Moro, C.V. The mosquito holobiont: Fresh insight into mosquito-microbiota interactions. Microbiome 2018, 6, 49. [Google Scholar] [CrossRef]
- Wang, X.; Liu, T.; Wu, Y.; Zhong, D.; Zhou, G.; Su, X.; Xu, J.; Sotero, C.F.; Sadruddin, A.A.; Wu, K.; et al. Bacterial microbiota assemblage in Aedes albopictus mosquitoes and its impacts on larval development. Mol. Ecol. 2018, 27, 2972–2985. [Google Scholar] [CrossRef] [Green Version]
- Luis, P.; Vallon, L.; Tran, F.-H.; Hugoni, M.; Tran-Van, V.; Mavingui, P.; Minard, G.; Moro, C.V. Aedes albopictus mosquitoes host a locally structured mycobiota with evidence of reduced fungal diversity in invasive populations. Fungal Ecol. 2019, 39, 257–266. [Google Scholar] [CrossRef]
- Merritt, R.W.; Clements, A.N. The Biology of Mosquitoes, Volume 1: Development, Nutrition, and Reproduction. A. N. Clements. J. N. Am. Benthol. Soc. 1993, 12, 304–305. [Google Scholar] [CrossRef]
- Kenney, J.L.; Solberg, O.D.; Langevin, S.A.; Brault, A.C. Characterization of a novel insect-specific flavivirus from Brazil: Potential for inhibition of infection of arthropod cells with medically important flaviviruses. J. Gen. Virol. 2014, 95, 2796–2808. [Google Scholar] [CrossRef]
- Mancini, M.V.; Spaccapelo, R.; Damiani, C.; Accoti, A.; Tallarita, M.; Petraglia, E.; Rossi, P.; Cappelli, A.; Capone, A.; Peruzzi, G.; et al. Paratransgenesis to control malaria vectors: A semi-field pilot study. Parasites Vectors 2016, 9, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, W.A. Mosquito Sugar Feeding and Reproductive Energetics. Annu. Rev. Èntomol. 1995, 40, 443–474. [Google Scholar] [CrossRef] [PubMed]
- Barredo, E.; DeGennaro, M. Not Just from Blood: Mosquito Nutrient Acquisition from Nectar Sources. Trends Parasitol. 2020, 36, 473–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza-Neto, J.A.; Machado, F.; Lima, J.B.; Valle, D.; Ribolla, P. Sugar digestion in mosquitoes: Identification and characterization of three midgut α-glucosidases of the neo-tropical malaria vector Anopheles aquasalis (Diptera: Culicidae). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 147, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Eliason, D.A. Feeding Adult Mosquitoes on Solid Sugars. Nature 1963, 200, 289. [Google Scholar] [CrossRef] [PubMed]
- Dillon, R.; El Kordy, E. Carbohydrate Digestion in Sandflies: α-Glucosidase Activity in the Midgut of Phlebotomus langeroni. Comp. Biochem. Physiol. Part B: Biochem. Mol. Boil. 1997, 116, 35–40. [Google Scholar] [CrossRef]
- Gusmão, D.S.; Santos, A.V.; Marini, D.C.; Russo, E.D.S.; Peixoto, A.M.D.; Júnior, M.B.; Berbert-Molina, M.A.; Lemos, F.J.A. First isolation of microorganisms from the gut diverticulum of Aedes aegypti (Diptera: Culicidae): New perspectives for an insect-bacteria association. Memórias do Instituto Oswaldo Cruz 2007, 102, 919–924. [Google Scholar] [CrossRef] [Green Version]
- Gusmão, D.S.; Santos, A.V.; Marini, D.C.; Bacci, M.; Berbert-Molina, M.A.; Lemos, F.J. Culture-dependent and culture-independent characterization of microorganisms associated with Aedes aegypti (Diptera: Culicidae) (L.) and dynamics of bacterial colonization in the midgut. Acta Trop. 2010, 115, 275–281. [Google Scholar] [CrossRef]
- Terenius, O.; Lindh, J.M.; Eriksson-Gonzales, K.; Bussière, L.F.; Laugen, A.T.; Bergquist, H.; Titanji, K.; Faye, I. Midgut bacterial dynamics in Aedes aegypti. FEMS Microbiol. Ecol. 2012, 80, 556–565. [Google Scholar] [CrossRef] [Green Version]
- Kalappa, D.M.; Subramani, P.A.; Basavanna, S.K.; Ghosh, S.K.; Sundaramurthy, V.; Uragayala, S.; Tiwari, S.; Anvikar, A.R.; Valecha, N. Influence of midgut microbiota in Anopheles stephensi on Plasmodium berghei infections. Malar. J. 2018, 17, 385. [Google Scholar] [CrossRef] [Green Version]
- Moro, C.V.; Tran, F.H.; Raharimalala, F.N.; Ravelonandro, P.; Mavingui, P. Diversity of culturable bacteria including Pantoea in wild mosquito Aedes albopictus. BMC Microbiol. 2013, 13, 70. [Google Scholar] [CrossRef]
- Bennett, K.L.; Gómez-Martínez, C.; Chin, Y.; Saltonstall, K.; McMillan, W.O.; Rovira, J.R.; Loaiza, J.R. Dynamics and diversity of bacteria associated with the disease vectors Aedes aegypti and Aedes albopictus. Sci. Rep. 2019, 9, 12160. [Google Scholar] [CrossRef] [PubMed]
- White, D.C.; Sutton, S.D.; Ringelberg, D.B. The genus Sphingomonas: Physiology and ecology. Curr. Opin. Biotechnol. 1996, 7, 301–306. [Google Scholar] [CrossRef]
- Dong, Y.; Manfredini, F.; Dimopoulos, G. Implication of the Mosquito Midgut Microbiota in the Defense against Malaria Parasites. PLOS Pathog. 2009, 5, e1000423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coon, K.L.; Vogel, K.J.; Brown, M.R.; Strand, M.R. Mosquitoes rely on their gut microbiota for development. Mol. Ecol. 2014, 23, 2727–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guégan, M.; Minard, G.; Tran, F.-H.; Van, V.T.; Dubost, A.; Moro, C.V. Short-term impacts of anthropogenic stressors on Aedes albopictus mosquito vector microbiota. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.A.; Liu, R.M.; Bennett, S.N. RNA shotgun metagenomic sequencing of northern California (USA) mosquitoes uncovers viruses, bacteria, and fungi. Front. Microbiol. 2015, 6, 185. [Google Scholar] [CrossRef] [Green Version]
- Valzania, L.; Coon, K.L.; Vogel, K.J.; Brown, M.R.; Strand, M.R. Hypoxia-induced transcription factor signaling is essential for larval growth of the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2018, 115, 457–465. [Google Scholar] [CrossRef] [Green Version]
- Malassigné, S.; Moro, C.V.; Luis, P. Mosquito Mycobiota: An Overview of Non-Entomopathogenic Fungal Interactions. Pathogens 2020, 9, 564. [Google Scholar] [CrossRef]
- Correa, M.A.; Brackney, D.E.; Steven, B. Axenic Aedes aegypti develop without live bacteria, but exhibit delayed development and reduced oviposition. BioRxiv 2018, 264978. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gilbreath, T.M.; Kukutla, P.; Yan, G.; Xu, J. Dynamic Gut Microbiome across Life History of the Malaria Mosquito Anopheles gambiae in Kenya. PLoS ONE 2011, 6, e24767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linenberg, I.; Christophides, G.K.; Gendrin, M. Larval diet affects mosquito development and permissiveness to Plasmodium infection. Sci. Rep. 2016, 6, 38230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tchioffo, M.T.; Boissière, A.; Abate, L.; Nsango, S.E.; Bayibéki, A.N.; Awono-Ambene, P.H.; Christen, R.; Gimonneau, G.; Morlais, I. Dynamics of Bacterial Community Composition in the Malaria Mosquito’s Epithelia. Front. Microbiol. 2016, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancini, M.V.; Damiani, C.; Accoti, A.; Tallarita, M.; Nunzi, E.; Cappelli, A.; Bozić, J.; Catanzani, R.; Rossi, P.; Valzano, M.; et al. Estimating bacteria diversity in different organs of nine species of mosquito by next generation sequencing. BMC Microbiol. 2018, 18, 126. [Google Scholar] [CrossRef] [Green Version]
- Nepomuceno, D.B.; Santos, V.C.; Araujo, R.; Pereira, M.H.; Sant’Anna, M.R.; Moreira, L.A.; Gontijo, N.F. pH control in the midgut of Aedes aegypti under different nutritional conditions. J. Exp. Boil. 2017, 220, 3355–3362. [Google Scholar] [CrossRef] [Green Version]
- Trembley, H.L. The Distribution of Certain Liquids in the Esophageal Diverticula and Stomach of Mosquitoes. Am. J. Trop. Med. Hyg. 1952, 1, 693–710. [Google Scholar] [CrossRef]
- Weaver, S.C.; Lorenz, L.H.; Scott, T.W. Distribution of Western Equine Encephalomyelitis Virus in the Alimentary Tract of Culex tarsalis (Diptera: Culicidae) Following Natural and Artificial Blood Meals. J. Med Èntomol. 1993, 30, 391–397. [Google Scholar] [CrossRef]
- Minard, G.; Tran, F.H.; Raharimalala, F.N.; Hellard, E.; Ravelonandro, P.; Mavingui, P.; Moro, C.V. Prevalence, genomic and metabolic profiles of Acinetobacter and Asaia associated with field-caught Aedes albopictus from Madagascar. FEMS Microbiol. Ecol. 2012, 83, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Minard, G.; Tran, F.-H.; Van, V.T.; Goubert, C.; Bellet, C.; Lambert, G.; Kim, K.L.H.; Thuy, T.H.T.; Mavingui, P.; Moro, C.V. French invasive Asian tiger mosquito populations harbor reduced bacterial microbiota and genetic diversity compared to Vietnamese autochthonous relatives. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Guégan, M.; Van, V.T.; Martin, E.; Minard, G.; Tran, F.; Fel, B.; Hay, A.; Simon, L.; Barakat, M.; Potier, P.; et al. Who is eating fructose within the Aedes albopictus gut microbiota? Environ. Microbiol. 2020, 22, 1193–1206. [Google Scholar] [CrossRef]
- Escudie, F.; Auer, L.; Bernard, M.; Cauquil, L.; Vidal, K.; Maman, S.; Mariadassou, M.; Hernandez-Raquet, G.; Pascal, G. FROGS: Find rapidly OTU with galaxy solution. Bioinformatics 2018, 34, 1287–1294. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2013, 42, D643–D648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- Anonymous. The R Project for Statistical Computing. Available online: http://www.r-project.org/ (accessed on 13 February 2012).
- McMurdie, P.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package, package version: 1.15-4; R Project Institute for Statistics and Mathematics, WU Wirtschaftsuniversität Wien: Wien, Austria, 2010; Available online: http://CRAN.R-project.org/package=vegan (accessed on 15 August 2011).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of Phylogenetics and Evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H.; Francois, R.; Henry, L.; Müller, K. RStudio dplyr: A Grammar of Data Manipulation. 2020. [Google Scholar]
- Slowikowski, K.; Schep, A.; Hughes, S.; Lukauskas, S.; Irisson, J.-O.; Kamvar, Z.N.; Ryan, T.; Christophe, D.; Hiroaki, Y.; Gramme, P. ggrepel: Automatically Position Non-Overlapping Text Labels with “ggplot2”. 2020. [Google Scholar]
- Wickham, H. The Split-Apply-Combine Strategy for Data Analysis. J. Stat. Softw. 2011, 40, 1–29. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Microorganism | Mosquito Sex | Organ | Shannon Index |
|---|---|---|---|
| Bacteria | Female | Crop | 2.74 ± 1.21 |
| Bacteria | Female | Gut | 3.17 ± 0.59 |
| Bacteria | Male | Crop | 3.42 ± 0.39 |
| Bacteria | Male | Gut | 3.17 ± 0.56 |
| Fungi | Female | Crop | 1.73 ± 0.82 |
| Fungi | Female | Gut | 1.22 ± 0.56 |
| Fungi | Male | Crop | 1.97 ± 0.64 |
| Fungi | Male | Gut | 1.65 ± 0.53 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guégan, M.; Martin, E.; Valiente Moro, C. Comparative Analysis of the Bacterial and Fungal Communities in the Gut and the Crop of Aedes albopictus Mosquitoes: A Preliminary Study. Pathogens 2020, 9, 628. https://doi.org/10.3390/pathogens9080628
Guégan M, Martin E, Valiente Moro C. Comparative Analysis of the Bacterial and Fungal Communities in the Gut and the Crop of Aedes albopictus Mosquitoes: A Preliminary Study. Pathogens. 2020; 9(8):628. https://doi.org/10.3390/pathogens9080628
Chicago/Turabian StyleGuégan, Morgane, Edwige Martin, and Claire Valiente Moro. 2020. "Comparative Analysis of the Bacterial and Fungal Communities in the Gut and the Crop of Aedes albopictus Mosquitoes: A Preliminary Study" Pathogens 9, no. 8: 628. https://doi.org/10.3390/pathogens9080628
APA StyleGuégan, M., Martin, E., & Valiente Moro, C. (2020). Comparative Analysis of the Bacterial and Fungal Communities in the Gut and the Crop of Aedes albopictus Mosquitoes: A Preliminary Study. Pathogens, 9(8), 628. https://doi.org/10.3390/pathogens9080628