
The Influence of the Degree of Forest Management on Methylmercury and the Composition of Microbial Communities in the Sediments of Boreal Drainage Ditches

 , , , ,
, , , ,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
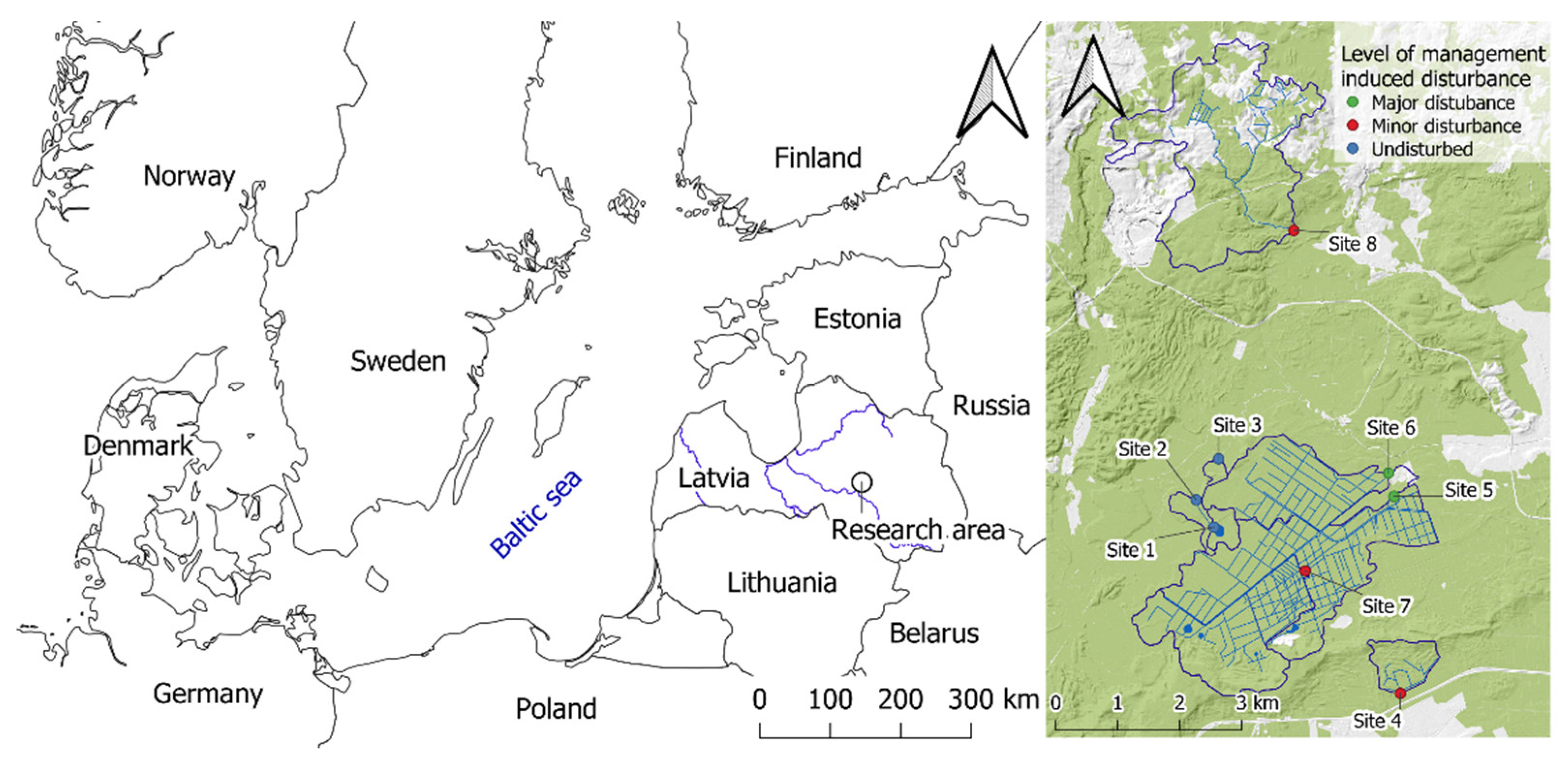
2.1. Sampling Sites
2.2. Sampling and Processing of Sediments
2.3. DNA Extraction, PCR Amplification and Sequencing
2.4. Sequencing Data Analysis
2.5. Chemical Analysis
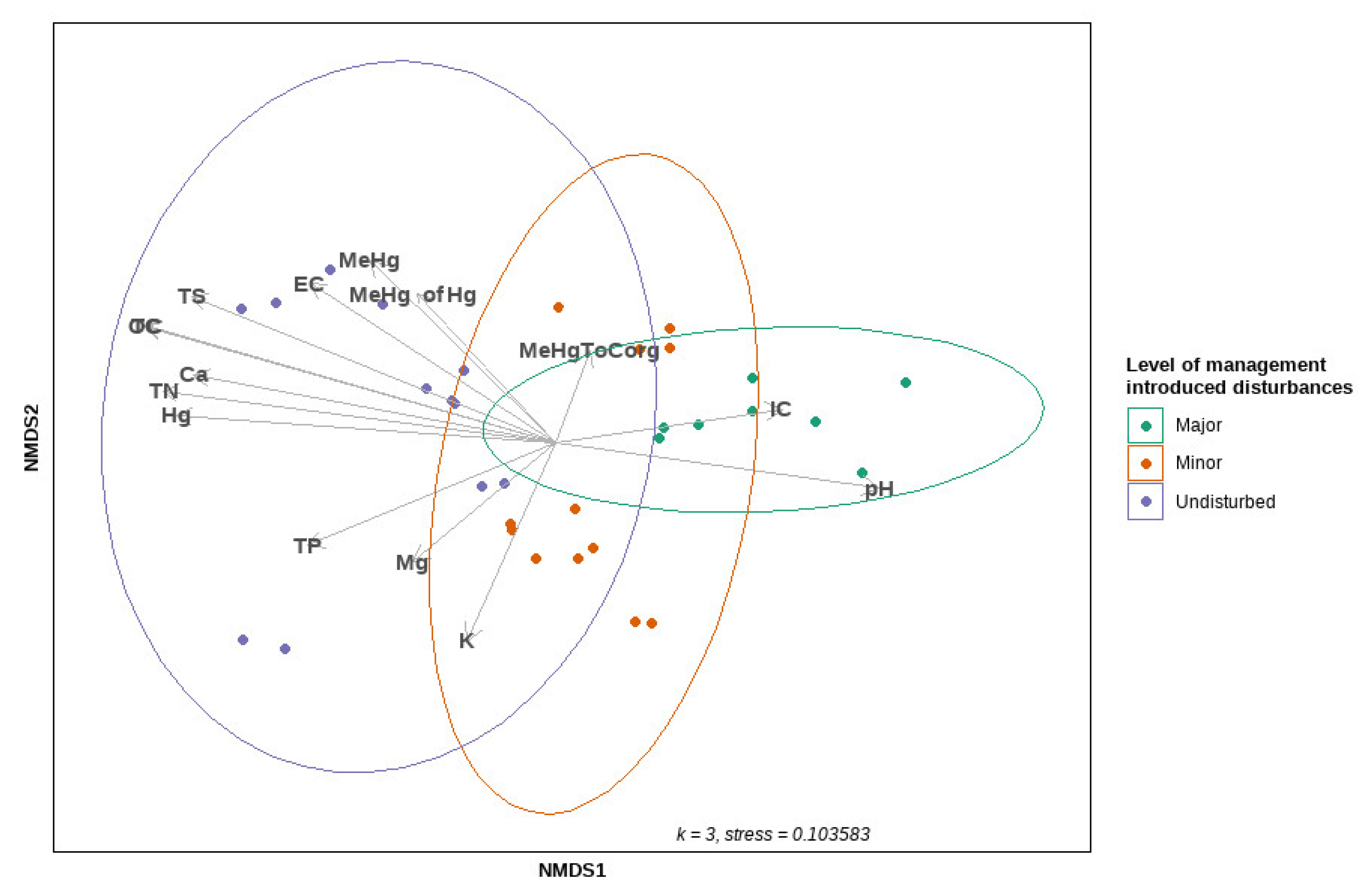
2.6. Bacterial and Archaeal Community and Statistical Analysis
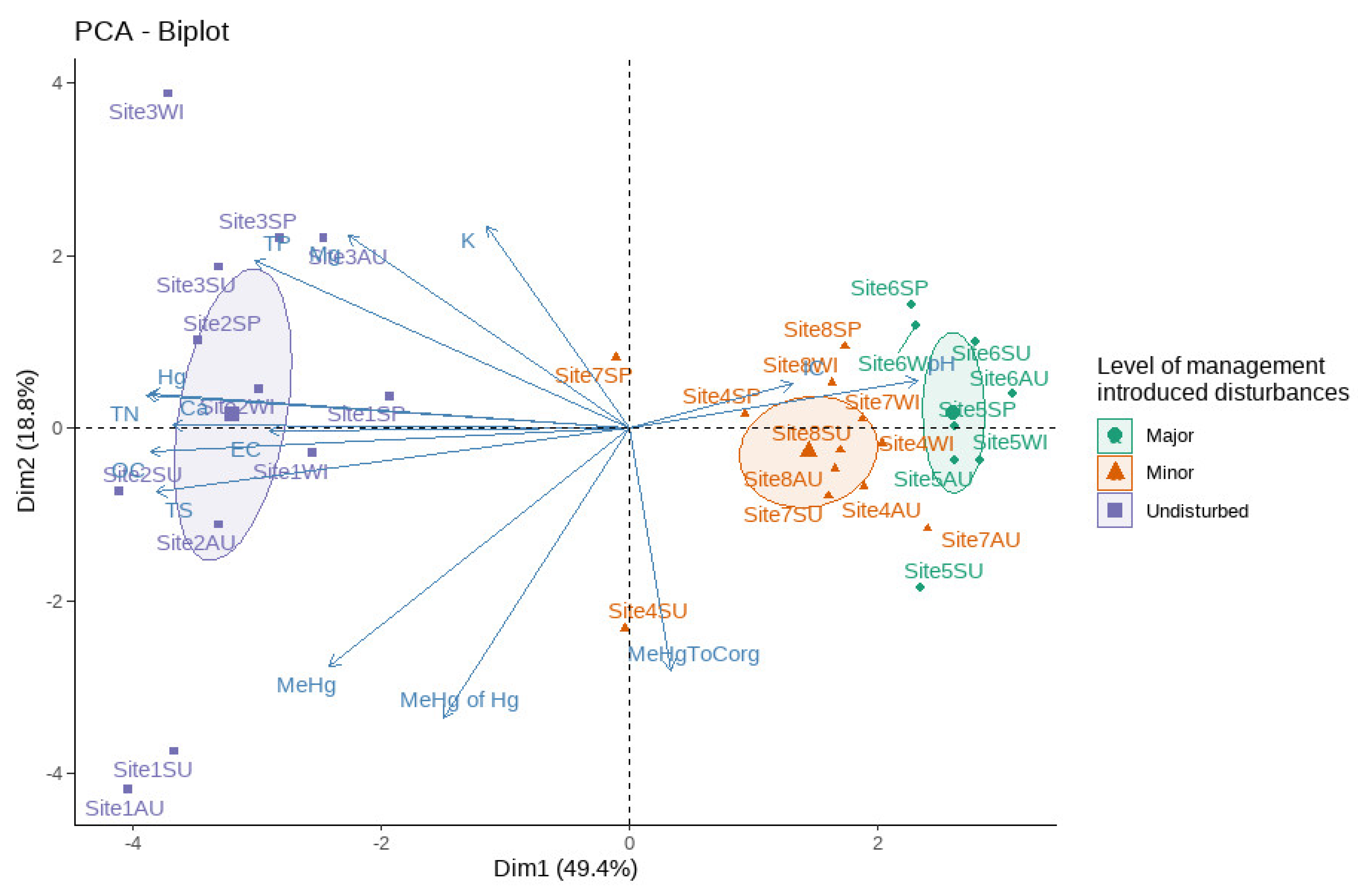
3. Results
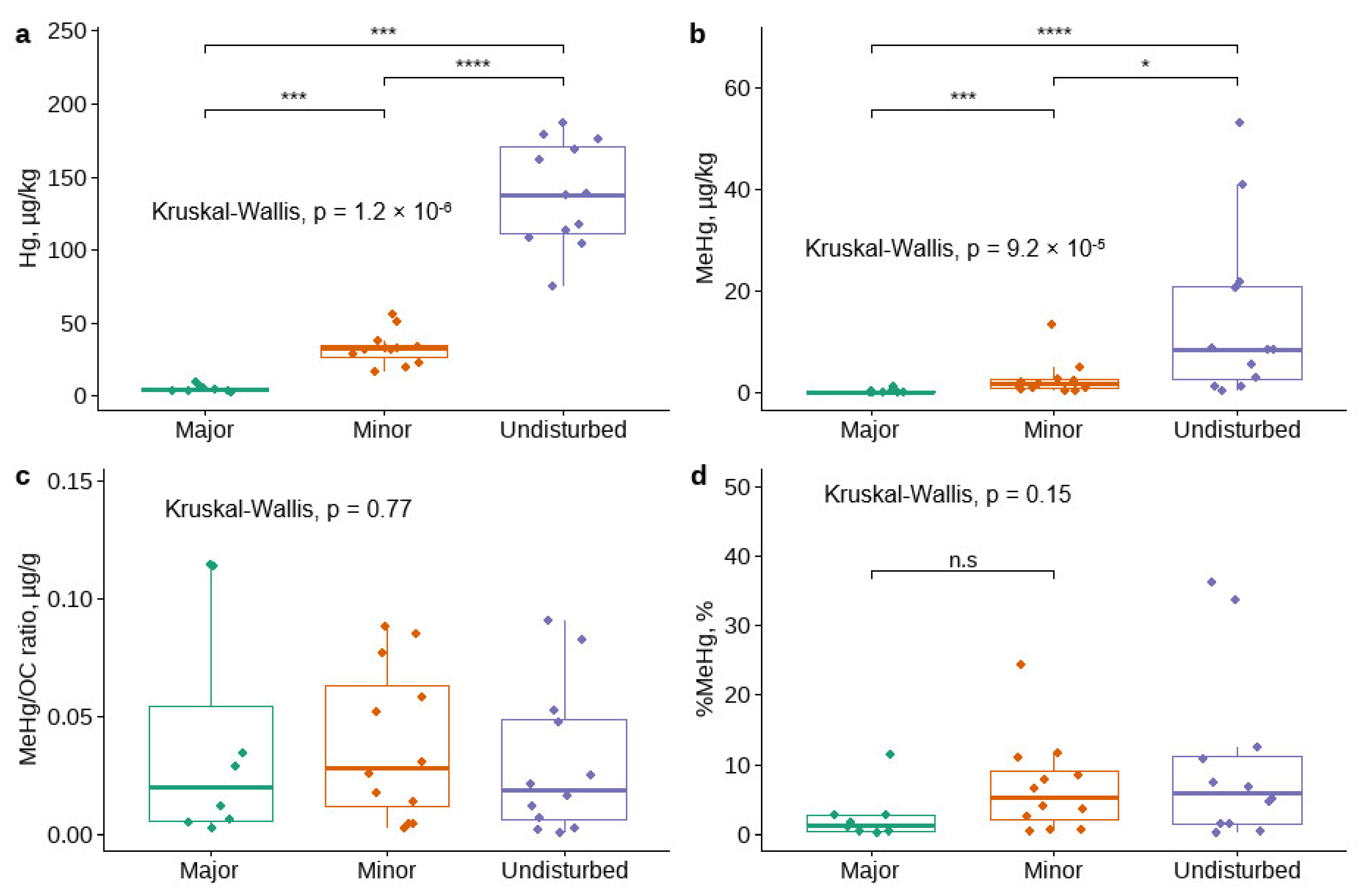
3.1. Sedimentary Hg Concentrations in Disturbed and Undisturbed Watercourses
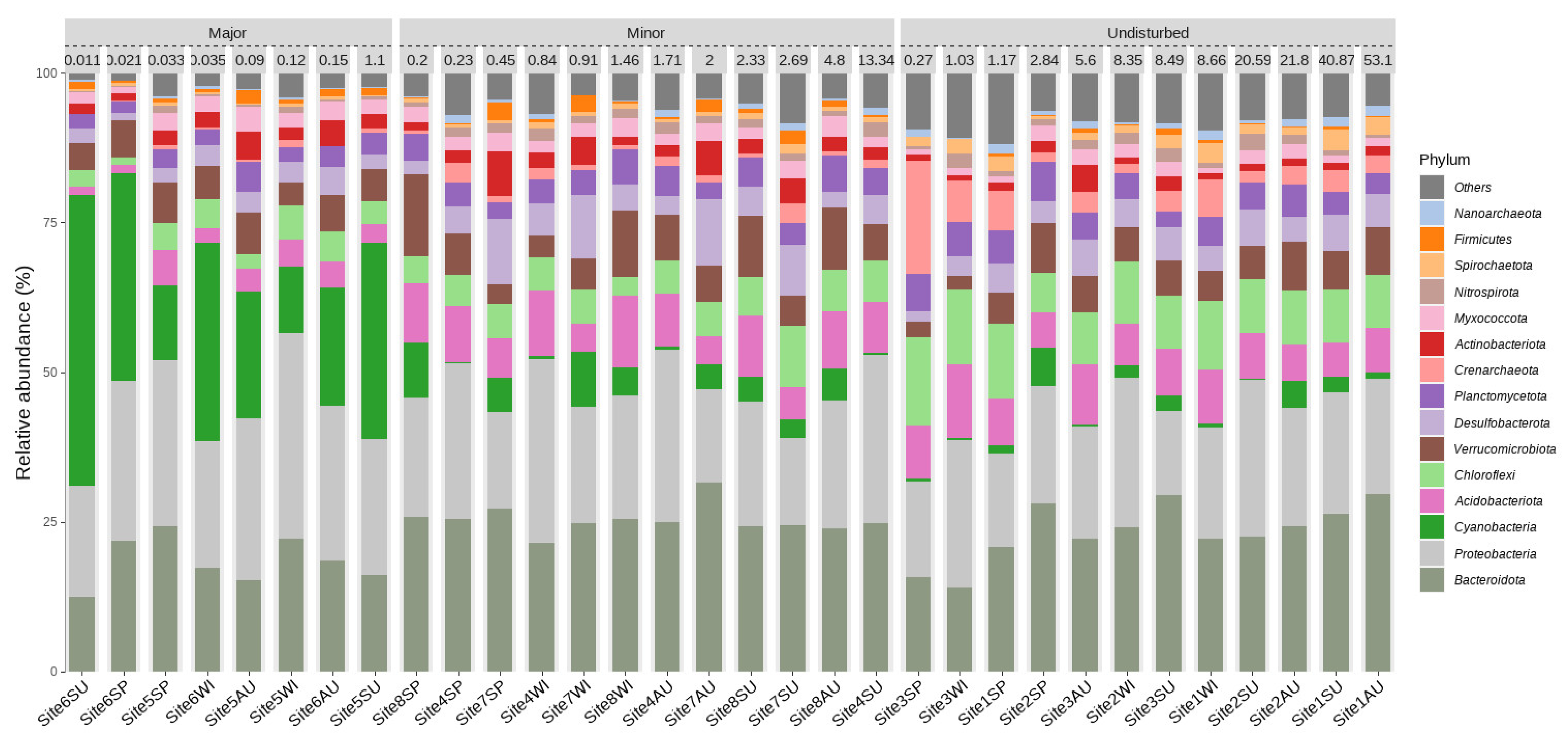
3.2. Microbial Composition and Diversity in Sediments of Disturbed and Undisturbed Water Courses
3.2.1. Bacterial and Archaeal Composition
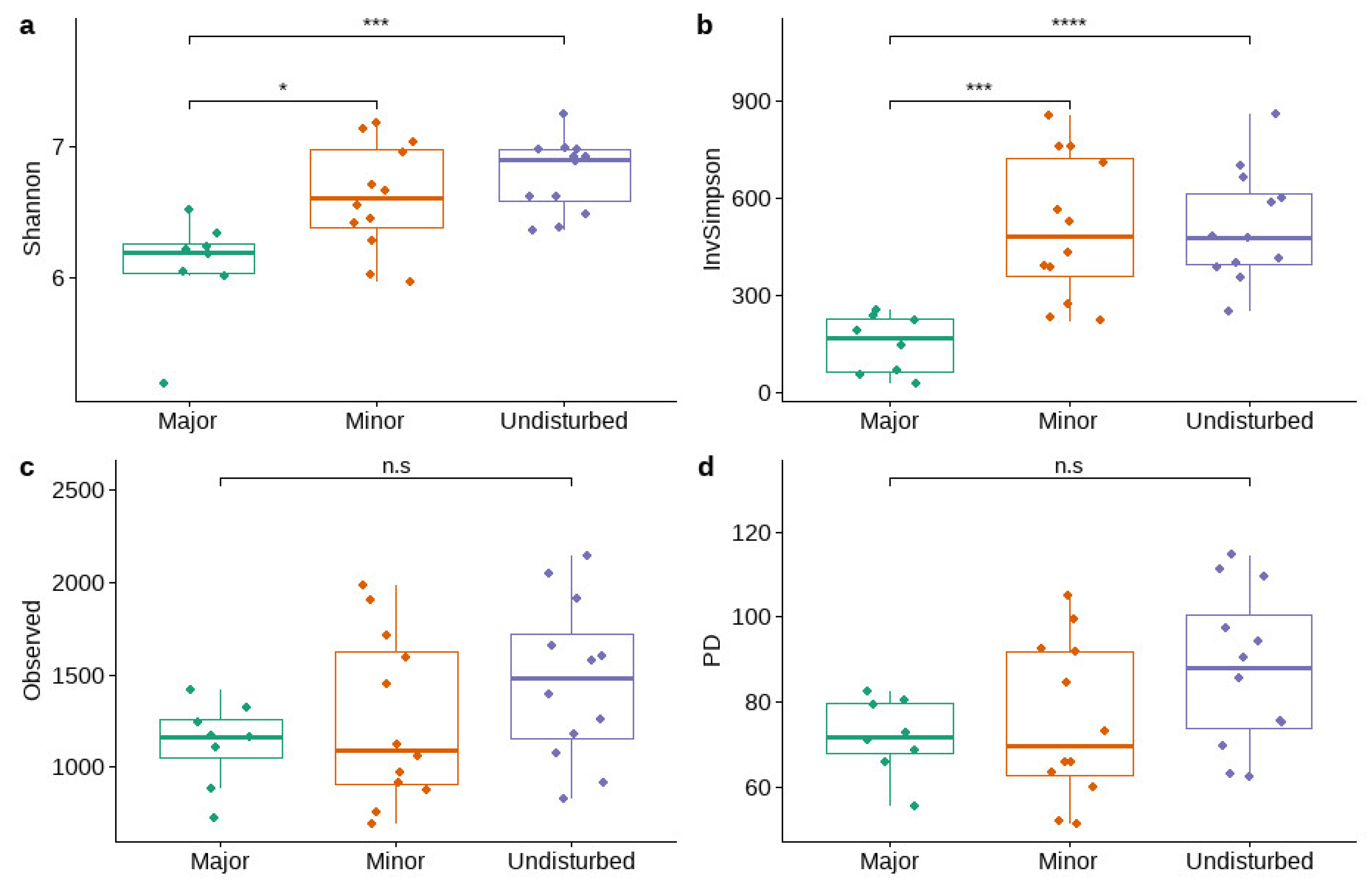
3.2.2. Alpha and Beta Diversity
3.2.3. Differential Abundance Testing and Indicator Analysis
3.3. Seasonal Variations in Sedimentary MeHg and Microbial Community Composition
4. Discussion
4.1. The Effect of Disturbances on the MeHg and THg Concentrations in the Sediment
4.2. The Effect of Disturbances on the Microbial Community in the Sediment
4.3. Temporal Variation in Sediment MeHg and Microbial Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lavoie, R.A.; Jardine, T.D.; Chumchal, M.M.; Kidd, K.A.; Campbell, L.M. Biomagnification of Mercury in Aquatic Food Webs. Environ. Sci. Technol. 2013, 47, 13385–13394. [Google Scholar] [CrossRef] [PubMed]
- Canuel, R.; Lucotte, M.; de Grosbois, S.B. Mercury Cycling and Human Health Concerns in Remote Ecosystems in the Americas. Sapiens 2009, 2, 1–13. [Google Scholar]
- WHO. Mercury and Health. 2017. Available online: https://www.who.int/news-room/fact-sheets/detail/mercury-and-health (accessed on 24 August 2022).
- WHO. 10 Chemicals of Public Health Concern. 2020. Available online: https://www.who.int/news-room/photo-story/photo-story-detail/10-chemicals-of-public-health-concern (accessed on 24 August 2022).
- Clayden, M.G.; Kidd, K.A.; Wyn, B.; Kirk, J.L.; Muir, D.C.G.; O’Driscoll, N.J. Mercury Biomagnification through Food Webs Is Affected by Physical and Chemical Characteristics of Lakes. Environ. Sci. Technol. 2013, 47, 12047–12053. [Google Scholar] [CrossRef] [PubMed]
- Azad, A.M.; Frantzen, S.; Bank, M.S.; Johnsen, I.A.; Tessier, E.; Amouroux, D.; Madsen, L.; Maage, A. Spatial Distribution of Mercury in Seawater, Sediment, and Seafood from the Hardangerfjord Ecosystem, Norway. Sci. Total Environ. 2019, 667, 622–637. [Google Scholar] [CrossRef]
- Bravo, A.G.; Cosio, C. Biotic Formation of Methylmercury: A Bio–Physico–Chemical Conundrum. Limnol. Oceanogr. 2020, 65, 1010–1027. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, T.W.; Magos, L. The Toxicology of Mercury and Its Chemical Compounds. Crit. Rev. Toxicol. 2006, 36, 609–662. [Google Scholar] [CrossRef]
- Fitzgerald, W.F.; Clarkson, T.W. Mercury and Monomethylmercury: Present and Future Concerns. Environ. Health Perspect. 1991, 96, 159–166. [Google Scholar] [CrossRef]
- Rafati-Rahimzadeh, M.; Rafati-Rahimzadeh, M.; Kazemi, S.; Moghadamnia, A.A. Current Approaches of the Management of Mercury Poisoning: Need of the Hour. DARU J. Pharm. Sci. 2014, 22, 46. [Google Scholar] [CrossRef]
- Hong, Y.S.; Kim, Y.M.; Lee, K.E. Methylmercury Exposure and Health Effects. J. Prev. Med. Public Health 2012, 45, 353–363. [Google Scholar] [CrossRef]
- UNEP Chemicals Branch. Global Mercury Assessment 2013: Sources, Emissions, Releases and Environmental Transport; UNEP Chemicals Branch: Geneva, Switzerland, 2013. [Google Scholar]
- Åkerblom, S.; Bignert, A.; Meili, M.; Sonesten, L.; Sundbom, M. Half a Century of Changing Mercury Levels in Swedish Freshwater Fish. Ambio 2014, 43, 91–103. [Google Scholar] [CrossRef]
- Levanoni, O.; Bishop, K.; Mckie, B.G.; Hartman, G.; Eklöf, K.; Ecke, F. Impact of Beaver Pond Colonization History on Methylmercury Concentrations in Surface Water. Environ. Sci. Technol. 2015, 49, 12679–12687. [Google Scholar] [CrossRef] [PubMed]
- Eklöf, K.; Lidskog, R.; Bishop, K. Managing Swedish Forestry’s Impact on Mercury in Fish: Defining the Impact and Mitigation Measures. Ambio 2016, 45, 163–174. [Google Scholar] [CrossRef] [PubMed]
- DIRECTIVE 2008/105/EC; European Parliament and of the Council of 16 December 2008 on Environmental Quality Standards in the Field of Water Policy, Amending and Subsequently Repealing Council Directives 82/176/EEC, 83/513/EEC, 84/156/EEC, 84/491/EEC. European Parliament: Bruxelles, Belgium, 2008.
- HELCOM. HELCOM Core Indicator Report; HELCOM: Helsinki, Finland, 2018. [Google Scholar]
- Soerensen, A.L.; Schartup, A.T.; Gustafsson, E.; Gustafsson, B.G.; Undeman, E.; Björn, E. Eutrophication Increases Phytoplankton Methylmercury Concentrations in a Coastal Sea—A Baltic Sea Case Study. Environ. Sci. Technol. 2016, 50, 11787–11796. [Google Scholar] [CrossRef]
- Mason, R.P.; Choi, A.L.; Fitzgerald, W.F.; Hammerschmidt, C.R.; Lamborg, C.H.; Soerensen, A.L.; Sunderland, E.M. Mercury Biogeochemical Cycling in the Ocean and Policy Implications. Environ. Res. 2012, 119, 101–117. [Google Scholar] [CrossRef]
- UNEP Chemicals Branch. The Global Atmospheric Mercury Assessment: Sources, Emissions and Transport; UNEP Chemicals Branch: Geneva, Switzerland, 2008. [Google Scholar]
- O’Connor, D.; Hou, D.; Ok, Y.S.; Mulder, J.; Duan, L.; Wu, Q.; Wang, S.; Tack, F.M.G.; Rinklebe, J. Mercury Speciation, Transformation, and Transportation in Soils, Atmospheric Flux, and Implications for Risk Management: A Critical Review. Environ. Int. 2019, 126, 747–761. [Google Scholar] [CrossRef]
- Xu, J.; Buck, M.; Eklöf, K.; Ahmed, O.O.; Schaefer, J.K.; Bishop, K.; Skyllberg, U.; Björn, E.; Bertilsson, S.; Bravo, A.G. Mercury Methylating Microbial Communities of Boreal Forest Soils. Sci. Rep. 2019, 9, 518. [Google Scholar] [CrossRef] [PubMed]
- Bishop, K.; Shanley, J.B.; Riscassi, A.; de Wit, H.A.; Eklöf, K.; Meng, B.; Mitchell, C.; Osterwalder, S.; Schuster, P.F.; Webster, J.; et al. Recent Advances in Understanding and Measurement of Mercury in the Environment: Terrestrial Hg Cycling. Sci. Total Environ. 2020, 721, 137647. [Google Scholar] [CrossRef] [PubMed]
- Morel, F.M.M.; Kraepiel, A.M.L.; Amyot, M. The Chemical Cycle and Bioaccumulation of Mercury. Annu. Rev. Ecol. Syst. 1998, 29, 543–566. [Google Scholar] [CrossRef]
- Bravo, A.G.; Bouchet, S.; Tolu, J.; Björn, E.; Mateos-Rivera, A.; Bertilsson, S. Molecular Composition of Organic Matter Controls Methylmercury Formation in Boreal Lakes. Nat. Commun. 2017, 8, 14255. [Google Scholar] [CrossRef]
- Åkerblom, S.; Nilsson, M.B.; Skyllberg, U.; Björn, E.; Jonsson, S.; Ranneby, B.; Bishop, K. Formation and Mobilization of Methylmercury across Natural and Experimental Sulfur Deposition Gradients. Environ. Pollut. 2020, 263, 114398. [Google Scholar] [CrossRef]
- Compeau, G.C.; Bartha, R. Principal Methylators. Microbiology 1985, 50, 498–502. [Google Scholar]
- Fleming, E.J.; Mack, E.E.; Green, P.G.; Nelson, D.C. Mercury Methylation from Unexpected Sources: Molybdate-Inhibited Freshwater Sediments and an Iron-Reducing Bacterium. Appl. Environ. Microbiol. 2006, 72, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Hamelin, S.; Amyot, M.; Barkay, T.; Wang, Y.; Planas, D. Methanogens: Principal Methylators of Mercury in Lake Periphyton. Environ. Sci. Technol. 2011, 45, 7693–7700. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, C.C.; Podar, M.; Bullock, A.L.; Graham, A.M.; Brown, S.D.; Somenahally, A.C.; Johs, A.; Hurt, R.A.; Bailey, K.L.; Elias, D.A. Mercury Methylation by Novel Microorganisms from New Environments. Environ. Sci. Technol. 2013, 47, 11810–11820. [Google Scholar] [CrossRef]
- Parks, J.M.; Johs, A.; Podar, M.; Bridou, R.; Hurt, R.A.; Smith, S.D.; Tomanicek, S.J.; Qian, Y.; Brown, S.D.; Brandt, C.C.; et al. The Genetic Basis for Bacterial Mercury Methylation. Science 2013, 339, 1332–1335. [Google Scholar] [CrossRef]
- Goñi-Urriza, M.; Corsellis, Y.; Lanceleur, L.; Tessier, E.; Gury, J.; Monperrus, M.; Guyoneaud, R. Relationships between Bacterial Energetic Metabolism, Mercury Methylation Potential, and HgcA/HgcB Gene Expression in Desulfovibrio Dechloroacetivorans BerOc1. Environ. Sci. Pollut. Res. 2015, 22, 13764–13771. [Google Scholar] [CrossRef]
- Braaten, H.F.V.; Lindholm, M.; de Wit, H.A. Five Decades of Declining Methylmercury Concentrations in Boreal Foodwebs Suggest Pivotal Role for Sulphate Deposition. Sci. Total Environ. 2020, 714, 136774. [Google Scholar] [CrossRef]
- Drott, A.; Lambertsson, L.; Björn, E.; Skyllberg, U. Do Potential Methylation Rates Reflect Accumulated Methyl Mercury in Contaminated Sediments? Environ. Sci. Technol. 2008, 42, 153–158. [Google Scholar] [CrossRef]
- Stamenkovic, J.; Gustin, M.S.; Dennett, K.E. Net Methyl Mercury Production versus Water Quality Improvement in Constructed Wetlands: Trade-Offs in Pollution Control. Wetlands 2005, 25, 748–757. [Google Scholar] [CrossRef]
- Yu, K.; Delaune, R.D.; Devai, I.; Tao, R.; Jugsujinda, A. Total and Methyl Mercury in Wetland Soils and Sediments of Louisiana’s Pontchartrain Basin (USA). J. Environ. Sci. Health-Part A Toxic/Hazard. Subst. Environ. Eng. 2008, 43, 1657–1662. [Google Scholar] [CrossRef]
- Selvendiran, P.; Driscoll, C.T.; Montesdeoca, M.R.; Choi, H.D.; Holsen, T.M. Mercury Dynamics and Transport in Two Adirondack Lakes. Limnol. Oceanogr. 2009, 54, 413–427. [Google Scholar] [CrossRef]
- Tjerngren, I.; Karlsson, T.; Björn, E.; Skyllberg, U. Potential Hg Methylation and MeHg Demethylation Rates Related to the Nutrient Status of Different Boreal Wetlands. Biogeochemistry 2012, 108, 335–350. [Google Scholar] [CrossRef]
- Bodaly, R.A.; Jansen, W.A.; Majewski, A.R.; Fudge, R.J.P.; Strange, N.E.; Derksen, A.J.; Green, D.J. Postimpoundment Time Course of Increased Mercury Concentrations in Fish in Hydroelectric Reservoirs of Northern Manitoba, Canada. Arch. Environ. Contam. Toxicol. 2007, 53, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Roy, V.; Amyot, M.; Carignan, R. Seasonal Methylmercury Dynamics in Water Draining Three Beaver Impoundments of Varying Age. J. Geophys. Res. Biogeosciences 2009, 114, G2. [Google Scholar] [CrossRef]
- Ecke, F.; Levanoni, O.; Audet, J.; Carlson, P.; Eklöf, K.; Hartman, G.; McKie, B.; Ledesma, J.; Segersten, J.; Truchy, A.; et al. Meta-Analysis of Environmental Effects of Beaver in Relation to Artificial Dams. Environ. Res. Lett. 2017, 12, 113002. [Google Scholar] [CrossRef]
- Herrero Ortega, S.; Catalán, N.; Björn, E.; Gröntoft, H.; Hilmarsson, T.G.; Bertilsson, S.; Wu, P.; Bishop, K.; Levanoni, O.; Bravo, A.G. High Methylmercury Formation in Ponds Fueled by Fresh Humic and Algal Derived Organic Matter. Limnol. Oceanogr. 2018, 63, S44–S53. [Google Scholar] [CrossRef]
- Eklöf, K.; Bishop, K.; Bertilsson, S.; Björn, E.; Buck, M.; Skyllberg, U.; Osman, O.A.; Kronberg, R.M.; Bravo, A.G. Formation of Mercury Methylation Hotspots as a Consequence of Forestry Operations. Sci. Total Environ. 2018, 613–614, 1069–1078. [Google Scholar] [CrossRef]
- Lidskog, R.; Bishop, K.; Eklöf, K.; Ring, E.; Sandström, C. From Wicked Problem to Governable Entity? The Effects of Forestry on Mercury in Qyatic Systems. For. Policy Econ. 2018, 90, 90–96. [Google Scholar] [CrossRef]
- Kronberg, R.M.; Drott, A.; Jiskra, M.; Wiederhold, J.G.; Björn, E.; Skyllberg, U. Forest Harvest Contribution to Boreal Freshwater Methyl Mercury Load. Global Biogeochem. Cycles 2016, 30, 825–843. [Google Scholar] [CrossRef]
- Bravo, A.G.; Loizeau, J.L.; Dranguet, P.; Makri, S.; Björn, E.; Ungureanu, V.G.; Slaveykova, V.I.; Cosio, C. Persistent Hg Contamination and Occurrence of Hg-Methylating Transcript (HgcA) Downstream of a Chlor-Alkali Plant in the Olt River (Romania). Environ. Sci. Pollut. Res. 2016, 23, 10529–10541. [Google Scholar] [CrossRef]
- Bravo, A.G.; Peura, S.; Buck, M.; Ahmed, O.; Mateos-Rivera, A.; Ortega, S.H.; Schaefer, J.K.; Bouchet, S.; Tolu, J.; Björn, E.; et al. Methanogens and Iron-Reducing Bacteria: The Overlooked Members of Mercury-Methylating Microbial Communities in Boreal Lakes. Appl. Environ. Microbiol. 2018, 84, e01774-18. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.R.; Johs, A.; Bi, L.; Lu, X.; Hu, H.W.; Sun, D.; He, J.Z.; Gu, B. Unraveling Microbial Communities Associated with Methylmercury Production in Paddy Soils. Environ. Sci. Technol. 2018, 52, 13110–13118. [Google Scholar] [CrossRef]
- Liu, Y.R.; Delgado-Baquerizo, M.; Bi, L.; Zhu, J.; He, J.Z. Consistent Responses of Soil Microbial Taxonomic and Functional Attributes to Mercury Pollution across China. Microbiome 2018, 6, 183. [Google Scholar] [CrossRef] [PubMed]
- Hortellani, M.A.; Sarkisa, J.E.S.; Bonetti, J.; Bonetti, C. Evaluation of Mercury Contamination in Sediments from Santos—São Vicente Estuarine System, São Paulo State, Brazil. J. Braz. Chem. Soc. 2005, 16, 1140–1149. [Google Scholar] [CrossRef]
- EPA Indicators: Sediment Mercury. 2022. Available online: https://www.epa.gov/national-aquatic-resource-surveys/indicators-sediment-mercury (accessed on 24 August 2022).
- Kļaviņa, Z.; Bārdule, A.; Eklöf, K.; Bitenieks, K.; Kļaviņš, I.; Lībiete, Z. Carbon, Nutrients and Methylmercury in Water from Small Catchments Affected by Various Forest Management Operations. Forests 2021, 12, 1278. [Google Scholar] [CrossRef]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every Base Matters: Assessing Small Subunit RRNA Primers for Marine Microbiomes with Mock Communities, Time Series and Global Field Samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef]
- Quince, C.; Lanzen, A.; Davenport, R.J.; Turnbaugh, P.J. Removing Noise from Pyrosequenced Amplicons. BMC Bioinform. 2011, 12, 38. [Google Scholar] [CrossRef]
- Větrovský, T.; Baldrian, P.; Morais, D. SEED 2: A User-Friendly Platform for Amplicon High-Throughput Sequencing Data Analyses. Bioinformatics 2018, 34, 2292–2294. [Google Scholar] [CrossRef]
- Anslan, S.; Bahram, M.; Hiiesalu, I.; Tedersoo, L. PipeCraft: Flexible Open-Source Toolkit for Bioinformatics Analysis of Custom High-Throughput Amplicon Sequencing Data. Mol. Ecol. Resour. 2017, 17, e234–e240. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Davison, A.C.; Hinkly, D. Bootstrap Methods and Their Application, 8th ed.; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- R Core Team. R Development Core Team. R A Lang. Environ. Stat. Comput. 2016, 55, 275–286. [Google Scholar]
- Davis, N.M.; Proctor, D.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple Statistical Identification and Removal of Contaminant Sequences in Marker-Gene and Metagenomics Data. Microbiome 2018, 6, 226. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 24 August 2022).
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R Tools for Integrating Phylogenies and Ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER V7; Primer-E Ltd.: Plymouth, UK, 2015; p. 296. [Google Scholar]
- Roberts, D.W. Labdsv: Ordination and Multivariate Analysis for Ecology 2019. Available online: https://CRAN.R-project.org/package=labdsv (accessed on 24 August 2022).
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef]
- Liu, C.; Cui, Y.; Li, X.; Yao, M. Microeco: An R Package for Data Mining in Microbial Community Ecology. FEMS Microbiol. Ecol. 2021, 97, fiaa255. [Google Scholar] [CrossRef]
- Kucheryavskiy, S. Mdatools—R Package for Chemometrics. Chemom. Intell. Lab. Syst. 2020, 198, 103937. [Google Scholar] [CrossRef]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-Regression: A Basic Tool of Chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Bergman, I.; Bishop, K.; Tu, Q.; Frech, W.; Åkerblom, S.; Nilsson, M. The Influence of Sulphate Deposition on the Seasonal Variation of Peat Pore Water Methyl Hg in a Boreal Mire. PLoS ONE 2012, 7, e45547. [Google Scholar] [CrossRef]
- Pierce, C.E.; Furman, O.S.; Nicholas, S.L.; Wasik, J.C.; Gionfriddo, C.M.; Wymore, A.M.; Sebestyen, S.D.; Kolka, R.K.; Mitchell, C.P.J.; Griffiths, N.A.; et al. Role of Ester Sulfate and Organic Disulfide in Mercury Methylation in Peatland Soils. Environ. Sci. Technol. 2022, 56, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- Coleman Wasik, J.K.; Engstrom, D.R.; Mitchell, C.P.J.; Swain, E.B.; Monson, B.A.; Balogh, S.J.; Jeremiason, J.D.; Branfireun, B.A.; Kolka, R.K.; Almendinger, J.E. The Effects of Hydrologic Fluctuation and Sulfate Regeneration on Mercury Cycling in an Experimental Peatland. J. Geophys. Res. Biogeosci. 2015, 120, 1697–1715. [Google Scholar] [CrossRef]
- Sun, G.; Riekerk, H.; Kornhak, L. Ground-Water-Table Rise after Forest Harvesting on Cypress-Pine Flatwoods in Florida. Wetlands 2000, 20, 101–112. [Google Scholar] [CrossRef]
- Fuhrmann, B.C.; Beutel, M.W.; O’Day, P.A.; Tran, C.; Funk, A.; Brower, S.; Pasek, J.; Seelos, M. Effects of Mercury, Organic Carbon, and Microbial Inhibition on Methylmercury Cycling at the Profundal Sediment-Water Interface of a Sulfate-Rich Hypereutrophic Reservoir. Environ. Pollut. 2021, 268, 115853. [Google Scholar] [CrossRef]
- Shade, A. Diversity Is the Question, Not the Answer. ISME J. 2017, 11, 1–6. [Google Scholar] [CrossRef]
- Frossard, A.; Donhauser, J.; Mestrot, A.; Gygax, S.; Bååth, E.; Frey, B. Long- and Short-Term Effects of Mercury Pollution on the Soil Microbiome. Soil Biol. Biochem. 2018, 120, 191–199. [Google Scholar] [CrossRef]
- Bravo, A.G.; Zopfi, J.; Buck, M.; Xu, J.; Bertilsson, S.; Schaefer, J.K.; Poté, J.; Cosio, C. Geobacteraceae Are Important Members of Mercury-Methylating Microbial Communities of Sediments Impacted by Waste Water Releases. ISME J. 2018, 12, 802–812. [Google Scholar] [CrossRef]
- Jones, D.S.; Johnson, N.W.; Mitchell, C.P.J.; Walker, G.M.; Bailey, J.V.; Pastor, J.; Swain, E.B. Diverse Communities of HgcAB+Microorganisms Methylate Mercury in Freshwater Sediments Subjected to Experimental Sulfate Loading. Environ. Sci. Technol. 2020, 54, 14265–14274. [Google Scholar] [CrossRef]
- Franco, M.W.; Mendes, L.A.; Windmöller, C.C.; Moura, K.A.F.; Oliveira, L.A.G.; Barbosa, F.A.R. Mercury Methylation Capacity and Removal of Hg Species from Aqueous Medium by Cyanobacteria. Water. Air. Soil Pollut. 2018, 229, 127. [Google Scholar] [CrossRef]
- Gionfriddo, C.M.; Wymore, A.M.; Jones, D.S.; Wilpiszeski, R.L.; Lynes, M.M.; Christensen, G.A.; Soren, A.; Gilmour, C.C.; Podar, M.; Elias, D.A. An Improved HgcAB Primer Set and Direct High-Throughput Sequencing Expand Hg-Methylator Diversity in Nature. Front. Microbiol. 2020, 11, 541554. [Google Scholar] [CrossRef] [PubMed]
- Hawinkel, S.; Mattiello, F.; Bijnens, L.; Thas, O. A Broken Promise: Microbiome Differential Abundance Methods Do Not Control the False Discovery Rate. Brief. Bioinform. 2019, 20, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Christensen, G.A.; Gionfriddo, C.M.; King, A.J.; Moberly, J.G.; Miller, C.L.; Somenahally, A.C.; Callister, S.J.; Brewer, H.; Podar, M.; Brown, S.D.; et al. Determining the Reliability of Measuring Mercury Cycling Gene Abundance with Correlations with Mercury and Methylmercury Concentrations. Environ. Sci. Technol. 2019, 53, 8649–8663. [Google Scholar] [CrossRef]
- Lei, P.; Zhang, J.; Zhu, J.; Tan, Q.; Kwong, R.W.M.; Pan, K.; Jiang, T.; Naderi, M.; Zhong, H. Algal Organic Matter Drives Methanogen-Mediated Methylmercury Production in Water from Eutrophic Shallow Lakes. Environ. Sci. Technol. 2021, 55, 10811–10820. [Google Scholar] [CrossRef] [PubMed]
- Peterson, B.D.; Mcdaniel, E.A.; Schmidt, A.G.; Lepak, R.F.; Janssen, S.E.; Tran, P.Q.; Marick, R.A.; Ogorek, J.M.; Dewild, J.F.; Krabbenhoft, D.P.; et al. Mercury Methylation Genes Identified across Diverse Anaerobic Microbial Guilds in a Eutrophic Sulfate-Enriched Lake. Environ. Sci. Technol. 2020, 54, 15840–15851. [Google Scholar] [CrossRef]
- Lázaro, W.L.; Díez, S.; Bravo, A.G.; da Silva, C.J.; Ignácio, Á.R.A.; Guimaraes, J.R.D. Cyanobacteria as Regulators of Methylmercury Production in Periphyton. Sci. Total Environ. 2019, 668, 723–729. [Google Scholar] [CrossRef]
- Villar, E.; Cabrol, L.; Heimbürger-Boavida, L.E. Widespread Microbial Mercury Methylation Genes in the Global Ocean. Environ. Microbiol. Rep. 2020, 12, 277–287. [Google Scholar] [CrossRef]
- Kearney, M.; Porter, W. Mechanistic Niche Modelling: Combining Physiological and Spatial Data to Predict Species’ Ranges. Ecol. Lett. 2009, 12, 334–350. [Google Scholar] [CrossRef]
- Hudelson, K.E.; Drevnick, P.E.; Wang, F.; Armstrong, D.; Fisk, A.T. Mercury Methylation and Demethylation Potentials in Arctic Lake Sediments. Chemosphere 2020, 248, 126001. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level of Management-Induced Disturbance | Sampling Sites | Sampling Sites Coordinates (Latitude; Longitude) | Description |
|---|---|---|---|
| Undisturbed | Site 1 | 56.689° N; 25.815° E | Natural stream from a peatland lake to river, close to the outlet |
| Site 2 | 56.693° N; 25.810° E | Natural stream from a peatland lake to river, 200 m downstream | |
| Site 3 | 56.699° N; 25.816° E | Natural stream with beaver site, nature reserve | |
| Minor | Site 4 | 56.664° N; 25.863° E | Drainage ditch, undisturbed for several decades |
| Site 7 | 56.682° N; 25.838° E | Drainage ditch, undisturbed for several decades | |
| Site 8 | 56.731° N; 25.839° E | Natural stream with slightly disturbed beaver site in recent years | |
| Major | Site 5 | 56.693° N; 25.862° E | Drainage ditch with beaver dam removal in 2017 |
| Site 6 | 56.696° N; 25.861° E | Drainage ditch, ditch cleaning in 2016 |
| Parameter, Unit | Value | Level of Management-Induced Disturbance of Watercourses | ||
|---|---|---|---|---|
| Undisturbed | Minor | Major | ||
| pH | mean ± S.E. | 6.2 ± 0.1 a | 6.5 ± 0.1 a | 7.0 ± 0.2 a |
| range | 5.8–6.7 | 5.7–7.1 | 6.3–7.6 | |
| EC, µS cm−1 | mean ± S.E. | 391 ± 49 a | 183 ± 48 ab | 54 ± 12 ac |
| range | 135–665 | 30–601 | 23–122 | |
| OC, g kg−1 | mean ± S.E. | 448 ± 20 a | 66 ± 10 b | 4 ± 1 c |
| range | 342–584 | 26–157 | 1–10 | |
| IC, g kg−1 | mean ± S.E. | <0.1 | <0.1 | 0.9 ± 0.5 |
| range | <0.1–<0.1 | <0.1–<0.1 | <0.1–4.0 | |
| TN, g kg−1 | mean ± S.E. | 22.3 ± 0.9 a | 4.2 ± 0.6 b | 0.4 ± 0.1 c |
| range | 18.3–28.9 | 2.3–9.3 | 0.1–1.1 | |
| TS, mg kg−1 | mean ± S.E. | 6568 ± 486 a | 781 ± 174 b | 0.5 ± 0.5 b |
| range | 4411–9644 | 196–2365 | <0.1–4.3 | |
| TP, g kg−1 | mean ± S.E. | 0.79 ± 0.096 a | 0.39 ± 0.04 a | 0.31 ± 0.06 a |
| range | 0.46–1.52 | 0.22–0.75 | 0.20–0.73 | |
| K, g kg−1 | mean ± S.E. | 1.16 ± 0.43 a | 0.58 ± 0.20 a | 0.33 ± 0.16 a |
| range | 0.11–4.11 | 0.17–2.53 | 0.08–1.42 | |
| Mg, g kg−1 | mean ± S.E. | 2.83 ± 0.35 a | 1.03 ± 0.15 a | 1.58 ± 0.48 a |
| range | 1.57–5.38 | 0.44–1.90 | 0.25–3.20 | |
| Ca, g kg−1 | mean ± S.E. | 30.9 ± 2.4 a | 6.4 ± 1.8 b | 4.9 ± 1.6 b |
| range | 7.8–38.4 | 0.5–23.3 | 0.2–10.6 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bitenieks, K.; Bārdule, A.; Eklöf, K.; Espenberg, M.; Ruņģis, D.E.; Kļaviņa, Z.; Kļaviņš, I.; Hu, H.; Lībiete, Z. The Influence of the Degree of Forest Management on Methylmercury and the Composition of Microbial Communities in the Sediments of Boreal Drainage Ditches. Microorganisms 2022, 10, 1981. https://doi.org/10.3390/microorganisms10101981
Bitenieks K, Bārdule A, Eklöf K, Espenberg M, Ruņģis DE, Kļaviņa Z, Kļaviņš I, Hu H, Lībiete Z. The Influence of the Degree of Forest Management on Methylmercury and the Composition of Microbial Communities in the Sediments of Boreal Drainage Ditches. Microorganisms. 2022; 10(10):1981. https://doi.org/10.3390/microorganisms10101981
Chicago/Turabian StyleBitenieks, Krišs, Arta Bārdule, Karin Eklöf, Mikk Espenberg, Dainis Edgars Ruņģis, Zane Kļaviņa, Ivars Kļaviņš, Haiyan Hu, and Zane Lībiete. 2022. "The Influence of the Degree of Forest Management on Methylmercury and the Composition of Microbial Communities in the Sediments of Boreal Drainage Ditches" Microorganisms 10, no. 10: 1981. https://doi.org/10.3390/microorganisms10101981
APA StyleBitenieks, K., Bārdule, A., Eklöf, K., Espenberg, M., Ruņģis, D. E., Kļaviņa, Z., Kļaviņš, I., Hu, H., & Lībiete, Z. (2022). The Influence of the Degree of Forest Management on Methylmercury and the Composition of Microbial Communities in the Sediments of Boreal Drainage Ditches. Microorganisms, 10(10), 1981. https://doi.org/10.3390/microorganisms10101981