Abstract
As a Gram-positive cocci existing in nature, Staphylococcus has a variety of species, such as Staphylococcus aureus and Staphylococcus epidermidis, etc. Growing evidence reveals that Staphylococcus is closely related to the occurrence and development of various cancers. On the one hand, cancer patients are more likely to suffer from bacterial infection and antibiotic-resistant strain infection compared to healthy controls. On the other hand, there exists an association between staphylococcal infection and carcinogenesis. Staphylococcus often plays a pathogenic role and evades the host immune system through surface adhesion molecules, α-hemolysin, PVL (Panton-Valentine leukocidin), SEs (staphylococcal enterotoxins), SpA (staphylococcal protein A), TSST-1 (Toxic shock syndrom toxin-1) and other factors. Staphylococcal nucleases (SNases) are extracellular nucleases that serve as genomic markers for Staphylococcus aureus. Interestingly, a human homologue of SNases, SND1 (staphylococcal nuclease and Tudor domain-containing 1), has been recognized as an oncoprotein. This review is the first to summarize the reported basic and clinical evidence on staphylococci and neoplasms. Investigations on the correlation between Staphylococcus and the occurrence, development, diagnosis and treatment of breast, skin, oral, colon and other cancers, are made from the perspectives of various virulence factors and SND1.
1. Introduction
Staphylococcus is a group of Gram-positive cocci that contains many different species, such as Staphylococcus aureus (S. aureus), Staphylococcus epidermidis (S. epidermidis), Staphylococcus saprophytics (S. saprophytics) [1,2,3,4]. As the most common pathogenic bacteria, S. aureus with different sequence types (STs) or spa types can cause inflammatory reactions in humans and animals [1,4,5]. The S. aureus-induced community and hospital-acquired infections may lead to adverse effects on the treatment and prognosis of patients [4]. With the widespread use of antibiotics in clinical practice, S. aureus has gradually become more drug-resistant, and the detection rate of methicillin-resistant Staphylococcus aureus (MRSA) also shows an upward trend [6]. Interestingly, Staphylococcus lugdunensis (S. lugdunensis) can secrete a polypeptide antibiotic called lugdunin to effectively restrain reproduction of and infection with MRSA [7]. As one of the main microorganisms on the skin’s surface, S. epidermidis plays an important role in the epidermal defense system of the body [8]. At present, more and more evidence supports the functional correlation between Staphylococcus and tumors, which is discussed in this review.
Staphylococcus in the host can play the role of inducing pathogenicity and escape from the host immune system through a variety of virulence factors, such as surface adhesion molecules, exotoxins and exoenzymes [9,10]. Various cell wall protein-anchored surface proteins, such as fibronectin-binding protein A/B (FnBPA/B), contribute to the adherence of Staphylococcus to host cells, which is the key to the staphylococcal pathogenesis [10,11,12,13]. As poreforming bacterial toxins, alpha-hemolysin and Panton-Valentine leukocidin (PVL) are considered to be the main virulence factors of severe infection caused by S. aureus infection [4,9,14]. A series of staphylococcal superantigens (SAg) produced by S. aureus can effectively activate the proliferation of T and B cells without any processing by antigen-presenting cells [9,15,16]. SpA (staphylococcal protein A) is one of the most important cell wall proteins in S. aureus, and has B cell superantigen activity [9]. SEs (staphylococcal enterotoxins) and TSST-1 (toxic shock syndrom toxin-1) function as potent inducers of cytotoxic T lymphocyte activity and cytokine production [15,16]. SEs include the Staphylococcus aureus enterotoxin A/B/C (SEA/B/C), and SEC is further divided into three subtypes (C1/2/3) [17,18]. TSST-1 can lead to toxic shock syndrome, and even multiple organ failure [19].
Extracellular nuclease is a secreted virulence factor and genetic marker for S. aureus. There exist two types of extracellular nuclease, staphylococcal nucleases (SNases) and thermonucleases (TNases) [20,21,22]. SND1 (staphylococcal nuclease and Tudor domain-containing 1) is the human homologue of Staphylococcus aureus nuclease, and can work as a member of RNA-induced silencing complex (RISC) that takes part in the cleavage of mRNA [23,24,25]. It is currently believed that human SND1 consists of four repeating staphylococcal nuclease-like (SN-like) domains [SN(1–4)] at the N terminus, and a SN5a-Tudor-SN5b (TSN) domain at the C terminus [25,26,27]. SND1 is a multifunctional protein that plays an important role in gene transcription regulation, pre-mRNA splicing, cell cycle, RNA metabolism and other biological processes [25,26,28,29,30,31,32,33]. Furthermore, a growing body of evidence reveals that SND1 with a recognizable nuclease domain is a kind of oncoprotein closely related to the occurrence and development of tumors, and which involves the potential nuclease activity [25,34,35,36,37].
In this study, we first conducted a retrieval from the Pubmed database using the search term: “(((((((((((Staphylococcus) or (Staphylococcus aureus)) or (Staphylococcus epidermidis)) OR (Staphylococcus saprophytics)) or (S. aureus)) or (S. epidermidis)) or (S. saprophytics)) or (Staphylococcus lugdunensis)) or (S. lugdunensis)) or (SND1)) or (staphylococcal nuclease)) AND ((((((carcinogenesis) or (cancer)) OR (cancers)) or (tumor)) or (tumors)) or (tumorigenesis))”. Then, the obtained literature was screened by reading the abstracts or full texts. Finally, we selected a total of 78 articles containing the scientific data between the presence of Staphylococcus and the occurrence, development, and treatment of different types of cancer. Table 1 summarizes the relevant clinical reports and basic experimental evidence, in terms of surface adhesion molecules, α-hemolysin, PVL, SEs, TSST-1, SpA, and SND1.

Table 1.
Summary of evidence on Staphylococcus and carcinogenesis.
2. Staphylococcus and Cancer-Related Clinical Reports
After the systematic literature research, a series of publications were retrieved regarding Staphylococcus and different clinical tumor diseases. For instance, when compared with negative controls, cancer patients tend to develop staphylococcal infections, and suffer from MRSA, which also greatly reduces the survival rate of patients with malignant tumors [40,61,86,91,109,113,116,117]. A 3-year retrospective study from a comprehensive cancer center reported that S. lugdunensis causes infection much less often than other coagulase-negative staphylococci species [81]. On the other hand, S. aureus is frequently detected in the oral cavity of most patients with malignant tumors undergoing chemotherapy and/or radiotherapy [47,58,86,87]. Maślak, E. et al. also observed the changes of Staphylococcus in the urine sample of prostate cancer patients treated with radiotherapy [112]. A study of an S. aureus bacteremia (SAB) case in a national database (n = 12,918) and a random population cohort (n = 117,465) analyzed the risk of primary cancer and discovered that SAB cases appeared more frequently in multiple myeloma, leukemia, sarcoma, cervical, liver, pancreatic, and urinary tract cancer, compared with a control group [100].
Microbiome sequencing and functional analysis for tumor and non-tumor patients will help to explore the correlation between staphylococcal system disorders and tumorigenesis prevention or treatment. Herein, we have gathered the scientific data on the functional relationship between staphylococci and several types of cancers.
2.1. Breast Cancer
Emerging evidence supports the links of Staphylococcus with breast diseases, especially breast cancer [99,118]. There are many clinical cases of breast cancer with MRSA [44]. Staphylococcus exhibits distinct distribution characteristics in different pathological tissues or states. For example, a relative abundance of Staphylococcus was detected in the breast tissues of women with breast cancer [78,82,97,98]. For instance, as the second most dominant bacterium, Staphylococcus (6.4% ± 9.4%) was prevalent in 22 out of 23 breast tissue samples of cases within black or white non-Hispanic cohorts of breast cancer [97]. Additionally, S. aureus and S. epidermidis are the common bacteria that cause infections around breast implants in cancer patients [72]. However, there are also reports with inconsistent conclusions. Breast microbiome profile data showed that the presence of Staphylococcus is negligible in the tissue of breast cancer [107], but An, J. et al. reported that the blood sample of healthy controls had a greater diversity of Staphylococcus than breast cancer patients [111].
2.2. Skin Cancer
In contrast to healthy skin, the presence of S. aureus DNA was strongly associated with squamous cell carcinoma [52]. Madhusudhan, N. et al. further reported that excessive S. aureus is significantly associated with an increased expression of human β-defensin-2 (HBD-2) in tumor samples from patients with cutaneous squamous cell carcinoma [93]. Cutaneous colonization of S. aureus is reportedly associated with the incidence of cutaneous T-cell lymphoma [69,119]. In response to adverse external stimuli, the expression microbiome of the body may become disorganized, such potentially suffering from a reduced level of the anti-tumor S. epidermidis population or a higher abundance of pathogenic S. aureus, which is associated with a high susceptibility to skin cancer [88,120,121]. When tumor patients are given specific clinical treatments, such as radiotherapy, chemotherapy, and probiotics, disorders of the skin microbiome are often observed [120,121].
2.3. Bladder Cancer
The altered abundance of Staphylococcus was detected in the tumor mucosa or urine samples of bladder cancer patients. For instance, Staphylococcus (cluster 2) was enriched in the microbial composition of tumor mucosa samples for bladder cancer [108]. Urine microbiota analysis of male bladder cancer patients in China indicated that various functional pathways were enriched in the cancer group, including S. aureus infection [85]. An abundance of Staphylococcus was significantly higher in urine samples of bladder cancer patients compared to benign prostatic hyperplasia controls [114].
2.4. Colon Cancer
In 2007, Noguchi, N. et al. first reported that tannin-producing S. ludunensis was more frequent in the swab samples of fecal and rectal for the advanced colon cancer group compared with the adenoma or normal group [48]. Furthermore, the genetic background investigation of the forty S. lugdunensis isolates from 288 rectal swabs indicated the links between the specific group D clone of S. lugdunensis and colon cancer [89].
2.5. Oral Cancer
Compared with healthy individuals, Staphylococcus was significantly more abundant in the oral squamous cell carcinomas group [110]. In 2004, Fujiki H. et al. found that tobacco tar-resistant S. aureus exists in the oral cavity of some individuals and has carcinogenic potential [42]. In addition, a study of 186 patients with oral squamous cell carcinoma reported a predominance of Gram-positive bacteria, including S. aureus and S. epidermidis, in the mouth of patients treated with chemotherapy and chemoradiotherapy [58].
2.6. Others
Apart from working to induce the discussed cancers, there are links between Staphylococcus and lung cancer, glioblastoma, and lymphoma. Fourdrain, A. et al. reported that the S. aureus carried in the nasal cavity before lung cancer surgery is related to an increased risk of health care-associated infection [94]. Similarly, S. epidermidis can also be detected in tissue samples taken from lung cancer patients during surgery [57]. In some glioblastoma multiforme cases, intracranial abscess complications caused by S. aureus have been observed [51]. Interestingly, some glioblastoma patients with staphylococcal intracranial infection after craniotomy displayed a relatively longer survival time [105]. However, the results are conflicting in breast implant-associated anaplastic large cell lymphoma (BIA-ALCL). It was reported that there was a high abundance of Staphylococcus in both breast implant-associated anaplastic and contralateral breast controls [90], but Hu H. et al. reported a lower abundance of Staphylococcus in the BIA-ALCL samples compared to that in the nontumor capsule specimens [79].
3. Staphylococcal Nuclease and Cancer
The presence or absence of S. aureus in samples can be determined by their diagnostic marker, staphylococcal nucleases [122]. Nucleases have long been recognized as potential biomarkers of cancer [36], however, no direct correlation between staphylococcal nucleases and cancer has been reported. The staphylococcal nuclease is a small globular protein containing 149 amino acid residues, and has been utilized to study the protein folding process [123]. As the staphylococcal nuclease purifies from a recombinant E. coli strain, micrococcal nuclease (Mnase) was applied in the chromatin immunoprecipitation assay or single-cell micrococcal nuclease sequencing of tumor samples [124,125]. SND1 is a conformed oncoprotein [25,34,35], which is the human homologue of SNases and contains four staphylococcal nuclease-like domains [23,24].
3.1. Structural Characteristics
Human SND1 protein (NP_055205.2; A0A140VK49_HUMAN), coded by the SND1 gene localized on chromosome 7q32.1 [34,126,127], consists of 910 amino acids. In 1997, Callebaut I. et al. first utilized the hydrophobic cluster analysis (HCA) method to initially resolve the structure of human SND1 protein and found that SND1 consists of four repetitive N-terminal SN and C-terminal Tudor domains [128]. In 2007, we first resolved the crystal structure of the TSN domain in human SND1 protein and found that TSN contains four α-helices, nine β-folds, and 14 linkage loops, in which the β (1~2) fold is involved in the composition of SN5a (679–703) [26]. Most of the α1-helices and β (3~6) fold to form a typical β-barrel Tudor (704–793) domain, and the β (7–9)-fold and α (2–4) helix are involved in the composition of SN5b (794–895) [26]. In 2008, Li, C. L. further reported that the SN3, SN4, Tudor and SN5 domains of human SND1 protein aggregate together to form a crescent-like structure [27]. The recessed basic surface formed by SN3 and SN4 serves as a binding site for citrate ions at the RNase active site, which can specifically bind with and degrade highly edited IU- and UI-containing double-stranded microRNA precursors [27]. Thus, staphylococcal nuclease-like domains of SND1 can bind to proteins and nucleic acids. This may involve a synergistic interaction between multiple SN structures.
3.2. Staphylococcal Nuclease Activity
The staphylococcal nuclease (SN) is a type of Ca2+-dependent enzyme that hydrolyzes the 5′-phosphodiester bond of single/double-stranded DNA and RNA [129,130]. It was initially thought that the SN domains of SND1 proteins lack key catalytic residues, like those of staphylococcal nucleases [24,128]. It was speculated that SND1 might have only nucleic acid binding ability, but no nuclease activities.
Nevertheless, emerging evidence suggests that the SND1 protein in multiple species can bind nucleic acids [27,131,132,133,134,135,136] and exhibits some nuclease activity [23,27,131,137,138,139,140,141,142,143,144]. For instance, Hannon et al. first discovered that the SND1 is a candidate of RISC and shows the nuclease activity in mammalian, Drosophila, and Caenorhabditis elegans, despite lacing a classical active site sequence [23,137]. In Plasmodium falciparum, the SND1 protein can degrade the RNA and single-stranded DNA, displaying Ca2+-dependent nuclease activity [131]. The nuclease activity of the SND1 protein was also detected in the species of Tick, Penaeus monodon, and Toxoplasma gondii [140,142,143,144]. In addition, the SND1 protein has some degradation ability for pri-miRNA/dsRNA and specific types of miRNAs after RNA editing which is supported by the crystal structure evidence [27]. SND1 protein degrades highly edited A to I pri-miR-142 [138]. Additionally, SND1 also specifically binds and degrades I-dsRNAs enriched in IU base pairs, without interacting with IU base pair-free dsRNAs [139].
3.3. SND1 and Cancer
The potential nuclease activity of the SN domain within SND1 may be closely linked to the oncogenic role of the SND1 protein [25,34,35,36,37]. SND1 plays a vital role in regulating several aspects of RNA metabolism through its nuclease activity. For instance, the binding of SND1 to the 3′UTR of PTPN23 (protein tyrosine phosphatase nonreceptor type 23) mRNA in human hepatocellular carcinoma (HCC) promotes its RNA degradation [37]. As a conventional staphylococcal nuclease inhibitor, pdTp (3′,5′-deoxythymidine bisphosphate) was reported to suppress the nuclease activity of SND1 [131,137]. In HCC cells, the remarkably enriched RISC activity of SND1 depends on the nuclease activity of highly expressed SND1, which can be affected by pdTp [56]. For the subcutaneous or in situ mouse models of HCC, the treatment of pdTp injection hinders the tumorigenesis of mice by affecting the nuclease activity of SND1 [84]. Scholarship generally concludes that the inhibition of SND1 nuclease activity by pdTp could be an effective intervention or therapeutic strategy for hepatocellular carcinoma.
4. Staphylococcus and Cancer Treatment
Clinical evidence indicates a correlation between the occurrence, development, and treatment of cancer and Staphylococcus [145]. In many cases, the predisposition to tumors is accompanied and facilitated by infection with specific staphylococci. Hattar, K. et al. reported that lipoteichoic acid, an inflammatory mediator from S. aureus, promotes the proliferation of lung cancer cell lines (A549 and H226) in vitro [83]. S. aureus infection was found to promote the lung metastasis of breast cancer cells through the formation of neutrophil extracellular traps [101]. Hence, some tumor-related interventions can be conducted, partly based on the pathogenesis of Staphylococcus. For instance, it may be possible to evade drug resistance in Staphylococcus and tumors by regulating intracellular reactive oxygen species [146].
Interestingly, there is continuous evidence that specific staphylococci have inhibitory effects on the proliferation, migration, and other biological behaviors of specific tumors [54,66]. For example, after intratumoral injection of S. aureus into the mouse model of orthotopic glioma, delayed glioma growth was observed, which may involve the anti-tumor effect of activated microglia [92].
4.1. Surface Adhesion Molecules
As a typical class of adhesion proteins from S. aureus, fibronectin-binding protein A/B (FnBPA/B) is associated with the adhesion and costimulatory signals of T lymphocytes [11,12]. The mice which were vaccinated with a recombinant Lactococcus lactis stain with cell surface-anchored FnBPA against S. aureus were better protected from the human papilloma virus (HPV)-induced cancer [76]. Aframomum melegueta extracts the display anti-adhesive abilities of S. aureus to lung carcinoma A549 cell line [106]. The extracellular adhesion protein (Eap) of S. aureus inhibited the bone metastasis of breast cancer cell line MDA-MB-231 [46]. In addition, some staphylococci were reported to adhere to bladder cancer cells. Szabados, F. et al. observed the internalization of S. saprophyticus ATCC 15305 into human urinary bladder carcinoma cell line 5637 in microscopy [49]. The treatment of metabolic glycoengineering with N-azidoacetyl-glucosamine (GlcNAz) leads to the reduced adherence of S. aureus to human T24 bladder carcinoma cells [64].
4.2. α-hemolysin
The α-hemolysin has certain anti-cancer effects and can also enhance the apoptosis of tumor cells induced by specific chemotherapy drugs [50,68,102]. For instance, a low toxic concentration of α-hemolysin can cause cell apoptosis through the mitochondrial pathway and improve the sensitivity of malignant pleural mesothelioma cells to cisplatin chemotherapy [50]. Additionally, researchers have tried to develop different bacterial delivery systems of α-hemolysin for the targeted killing of colorectal or breast cancer cells using Escherichia coli without the virulence factors [68,102].
4.3. Panton-Valentine leukocidin
As the S component of Panton-Valentine leukocidin, LukS-PV can induce mitochondria-mediated apoptosis and G0/G1 cell cycle arrest in human acute myeloid leukemia (AML) cell line (THP-1) [65], and effectively inhibit the tumorigenesis of HL-60 AML cells in severe combined immunodeficiency (SCID) mice [70]. This indicates that LukS-PV may be a multi-target drug candidate for the prevention and treatment of AML. For non-small-cell lung cancer (NSCLC) cells, LukS-PV promotes the apoptosis and cycle arrest of A549 and H460 cells through the P38/ERK MAPK signaling pathway [95]. For liver cancer, LukS-PV inhibits the migration of hepatocellular carcinoma cells by down-regulating histone deacetylase 6 (HDAC6) and increasing α-tubulin acetylation [115], and induces the apoptosis of HepG2 cells by regulating key proteins and metabolic pathways [96].
4.4. Staphylococcal Superantigens
Currently, there are many S. aureus superantigens, such as SEA, SEB, SEC, TSST1, and SpA, which can exert anti-tumor effects by inducing immune cell death, tumor cell apoptosis and other mechanisms [147,148,149]. Several tumor-specific superantigens for cancer treatment are under development [39,150,151].
4.4.1. Staphylococcus Aureus Enterotoxin A
Enhanced SEA expression in tumor cells with poor immunogenicity increases immunogenicity as a vaccine [53]. In addition, SEA can be utilized in the design of fission superantigen fusion proteins for cancer immunotherapy [41,62,147,151]. For instance, Dohlsten M. et al. designed a C242Fab-SEA fusion protein to target SEA-reactive T cells against MHC-class II negative human colon cancer cells at nanomolar concentrations in vitro [41]. Additionally, an oncolytic adenovirus (PPE3-SEA) was reported to inhibit the growth of mice bladder cancer MB49 cells [62].
4.4.2. Staphylococcus Aureus Enterotoxin B
Like SEA, SEB has significant anti-tumor effects by activating T cells in tumor-bearing mice [38]. Akbari, A. et al. reported that SEB effectively down-regulated the expression of SMAD family members by 2/3 and reduced the proliferation of human primary glioblastoma cell line U87 [80]. Several publications reported the links between SEB and bladder cancer. SEB can activate T lymphocytes and inhibit bladder tumor cell growth in vitro and in vivo [152]. The anti-angiogenic effect of SEB was also observed in an experiment using a rat model of nonmuscle invasive bladder cancer [59]. SEB-stimulated peripheral blood mononuclear cells can lead to the apoptosis of transitional cell carcinoma cells [43]. Similarly, the corresponding modifications of SEB serve as efficient instruments of cancer therapy [60,147]. For instance, Gu L. et al. designed the SEB-H32Q/K173E mutant, which retains the properties of SAg, enhances the host immune response to tumor disease, and reduces the associated thermotoxicity [60].
4.4.3. Staphylococcus Aureus Enterotoxin C
Highly agglutinative staphylococcin (HAS), a mixture of S. aureus culture filtrate, plays a certain immunomodulatory role through the active SEC component in the clinical treatment of breast cancer, colon cancer, bladder cancer and other cancers [74,75,153]. As a result, HAS may reduce the side effects of radiotherapy or chemotherapy in specific tumors to a certain extent and improve the survival prognosis of patients [74,153]. In China, SEC2 and a series of mutants have commonly been used as antitumor immunotherapy agents [67,154,155].
4.4.4. Toxic Shock Syndrom Toxin-1
Superantigen TSST-1 was reported to stimulate T-cell activation and enhance the cytotoxic effect of T cells on colorectal cancer LoVo cells [63]. Jiang Y. Q. et al. reported that the fusion of protein TSST-1 with a 12-mer peptide was able to inhibit the hepatocellular carcinoma cell growth by activating T lymphocytes [45]. Additionally, LINC00847 lncRNA may serve as a therapeutic target of the staphylococcal enterotoxin TST gene in renal cell carcinoma [104].
4.4.5. Staphylococcal Protein A
As one of the most essential S. aureus cell wall proteins, SpA can be utilized in the clinical treatment of cancer [156]. Based on the cross-linking between SpA and the Fc region of an immunoglobulin, the immunoprecipitation assay of tumor-related protein molecular interactions can be performed, or the delivery system of anti-cancer antibodies or drugs can be prepared [157,158]. For instance, an alkyl vinyl sulfone/protein A-based immunostimulating complex was established to deliver the cancer drugs to trastuzumab-resistant HER2 (human epidermal growth factor receptor 2)-overexpressing breast HCC1954 cells [73].
4.5. Others
Other substances of Staphylococcus are found to have certain tumor-suppressive effects. First, a protein purified from Staphylococcus hominis strain MANF2 was found to have the ability to reduce the viability of colon cancer cell line (HT-29) and lung cancer cell line (A549) when associated with fermented food [103]. Second, the chemotaxis inhibitory protein of S. aureus can inhibit the mitochondrial peptide-induced migration of U87 glioblastoma cells [71]. Third, the peptidoglycan of infectious S. aureus can actively trigger the Toll-like receptor 2 to promote the invasiveness and adhesiveness of MDA-MB-231 cells in vitro [55]. Fifth, the S. epidermidis strain MO34 inhibited the melanoma growth by producing 6-n-hydroxyaminopurine [88,159]. Sixth, cytoplasmic fractions of Enterococcus faecalis and Staphylococcus hominis, isolated from human breast milk, can inhibit the proliferation of MCF-7 cells [77]. Lastly, S. aureus-derived extracellular vesicles enhance the efficacy of tamoxifen therapy in breast cancer cells (MCF7 and BT474) [111].
5. Conclusions
The treatment of clinical cancer patients is often complicated with Staphylococcus infection, and different tumor treatments are often accompanied by a change in the Staphylococcus spectrum. Other types of staphylococci have distinct and even opposite effects on the occurrence and development of specific tumors. Herein, we provided a bidirectional functional effect model of Staphylococcus on carcinogenesis, as shown in Figure 1.
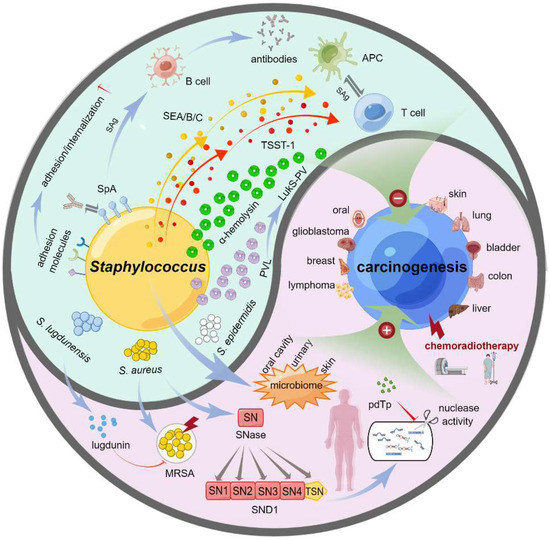
Figure 1.
Bidirectional functional effects of Staphylococcus on carcinogenesis. Staphylococcus has the bidirectional effects on carcinogenesis in various types of cancers, such as skin cancer, lung cancer, bladder cancer, colon cancer, liver cancer, lymphoma, breast cancer, glioblastoma, and oral cancer. On the one hand, the changes of staphylococcal flora in some tissues of the body, such as oral cavity, skin or urinary system, was linked to the predisposition to cancer or detected in cancer cases undergoing chemotherapy and/or radiotherapy. MRSA is often associated with a reduced survival rate of patients with malignant tumors. SNases work as the extracellular nucleases of S. aureus, and there exists a human homologue of SNases, SND1, which is closely related to the occurrence and development of different cancers. On the other hand, S. lugdunensis can secrete a lugdunin to curb the reproduction and infection of MRSA. S. aureus may play the role of tumor inhibition through the points of bacterial toxins (alpha-hemolysin, PVL or LurkS-PV), superantigens (SEA/B/C, TSST-1, SpA) of T/B cells, or adhesion molecules. Additionally, the inhibition of SND1 nuclease activity by pdTp may be an effective intervention or therapeutic strategy for liver cancer. This figure was drawn by Figdraw.
To treat cancer patients with bacterial infections, it is important to suppress their complications, starting with the pathogenic mechanism of specific Staphylococcus. Targeting the structures, secreted products, or artificial modifications of various virulence factors may result in great success when treating tumors. The accurate and efficient application of specific staphylococcal anti-tumor components also depends on basic experimental evidence, as well as the ongoing improvement of the system for the separation, purification, and presentation of active components.
In this review, we, for the first time, summarize the clinical reports, cellular and animal experimental evidence regarding the association between Staphylococcus and the diagnosis and treatment of tumors. Additionally, we systematically investigated the functional links between staphylococci and the occurrence, development, diagnosis, and treatment of breast, skin, oral, colon, and other types of cancers, in terms of surface adhesion molecules, α-hemolysin, PVL, SEs, TSST-1, SpA, and SND1, which provides novel insight into the functional relationship between bacterial infections and tumors.
Author Contributions
Conceptualisation: J.Y., Y.R., X.G.; data curation: X.Y., Y.W., E.S.; writing—original draft preparation: Y.W., X.G., Y.R.; writing—review and editing: E.S., J.Y.; project administration: J.Y.; funding acquisition: J.Y., X.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from Tianjin Natural Science Foundation (20JCYBJC00470 to X.J.); National Nature Science Foundation of China (32271201, 32070724 to J.Y.); Scientific Research Project of Tianjin Education Commission (Natural Science) (2019KJ171 to Y.R.); Excellent Talent Project of Tianjin Medical University (to J.Y.).
Data Availability Statement
Availability of published literature and correspondence should be addressed to the corresponding author.
Conflicts of Interest
The author declares no conflict of interest.
References
- Cvetnić, L.; Samardžija, M.; Duvnjak, S.; Habrun, B.; Cvetnić, M.; Jaki Tkalec, V.; Đuričić, D.; Benić, M. Multi Locus Sequence Typing and spa Typing of Staphylococcus Aureus Isolated from the Milk of Cows with Subclinical Mastitis in Croatia. Microorganisms 2021, 9, 725. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcus epidermidis—The ‘accidental’ pathogen. Nat. Reviews. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef]
- Raz, R.; Colodner, R.; Kunin, C.M. Who are you—Staphylococcus saprophyticus? Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2005, 40, 896–898. [Google Scholar] [CrossRef]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and virulence of Staphylococcus aureus. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef]
- Saidi, R.; Kaidi, R.; Khelef, D.; Solmaz, H.; Ergun, Y.; Mimoune, N.; Cantekin, Z. Investigation of the presence of slime production, VanA gene and antiseptic resistance genes in staphylococci isolated from bovine mastitis in Algeria. Vet. Stanica 2020, 52, 57–63. [Google Scholar] [CrossRef]
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus aureus: Molecular Characterization, Evolution, and Epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef]
- Wei, G.; He, Y. Antibacterial and Antibiofilm Activities of Novel Cyclic Peptides against Methicillin-Resistant Staphylococcus aureus. Int. J. Mol. Sci. 2022, 23, 8029. [Google Scholar] [CrossRef]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Reviews. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Tam, K.; Torres, V.J. Staphylococcus aureus Secreted Toxins and Extracellular Enzymes. Microbiol. Spectr. 2019, 7, 1–34. [Google Scholar] [CrossRef]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Höök, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Reviews. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef]
- Speziale, P.; Pietrocola, G. The Multivalent Role of Fibronectin-Binding Proteins A and B (FnBPA and FnBPB) of Staphylococcus aureus in Host Infections. Front. Microbiol. 2020, 11, 2054. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, Y.J.; Wann, E.R.; Fowler, T.; Duffield, E.; Höök, M.; McIntyre, B.W. Fibronectin binding protein A of Staphylococcus aureus can mediate human T lymphocyte adhesion and coactivation. J. Immunol. 2001, 166, 5129–5138. [Google Scholar] [CrossRef]
- Geoghegan, J.A.; Foster, T.J. Cell Wall-Anchored Surface Proteins of Staphylococcus aureus: Many Proteins, Multiple Functions. Curr. Top. Microbiol. Immunol. 2017, 409, 95–120. [Google Scholar] [CrossRef]
- von Hoven, G.; Qin, Q.; Neukirch, C.; Husmann, M.; Hellmann, N. Staphylococcus aureus α-toxin: Small pore, large consequences. Biol. Chem. 2019, 400, 1261–1276. [Google Scholar] [CrossRef] [PubMed]
- Abdurrahman, G.; Schmiedeke, F.; Bachert, C.; Bröker, B.M.; Holtfreter, S. Allergy-A New Role for T Cell Superantigens of Staphylococcus aureus? Toxins 2020, 12, 176. [Google Scholar] [CrossRef] [PubMed]
- Spaulding, A.R.; Salgado-Pabón, W.; Kohler, P.L.; Horswill, A.R.; Leung, D.Y.; Schlievert, P.M. Staphylococcal and streptococcal superantigen exotoxins. Clin. Microbiol. Rev. 2013, 26, 422–447. [Google Scholar] [CrossRef]
- Dinges, M.M.; Orwin, P.M.; Schlievert, P.M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34. [Google Scholar] [CrossRef]
- Marr, J.C.; Lyon, J.D.; Roberson, J.R.; Lupher, M.; Davis, W.C.; Bohach, G.A. Characterization of novel type C staphylococcal enterotoxins: Biological and evolutionary implications. Infect. Immun. 1993, 61, 4254–4262. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Qin, C.; Zhang, X.; Zhu, Y.; Li, A.; Wang, M.; Tang, Y.; Kreiswirth, B.N.; Chen, L.; Zhang, H.; et al. The tst gene associated Staphylococcus aureus pathogenicity island facilitates its pathogenesis by promoting the secretion of inflammatory cytokines and inducing immune suppression. Microb. Pathog. 2020, 138, 103797. [Google Scholar] [CrossRef]
- Tang, J.; Zhou, R.; Shi, X.; Kang, M.; Wang, H.; Chen, H. Two thermostable nucleases coexisted in Staphylococcus aureus: Evidence from mutagenesis and in vitro expression. FEMS Microbiol. Lett. 2008, 284, 176–183. [Google Scholar] [CrossRef]
- Hu, Y.; Xie, Y.; Tang, J.; Shi, X. Comparative expression analysis of two thermostable nuclease genes in Staphylococcus aureus. Foodborne Pathog. Dis. 2012, 9, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Bronner, S.; Monteil, H.; Prévost, G. Regulation of virulence determinants in Staphylococcus aureus: Complexity and applications. FEMS Microbiol. Rev. 2004, 28, 183–200. [Google Scholar] [CrossRef] [PubMed]
- Caudy, A.A.; Ketting, R.F.; Hammond, S.M.; Denli, A.M.; Bathoorn, A.M.; Tops, B.B.; Silva, J.M.; Myers, M.M.; Hannon, G.J.; Plasterk, R.H. A micrococcal nuclease homologue in RNAi effector complexes. Nature 2003, 425, 411–414. [Google Scholar] [CrossRef]
- Ponting, C.P. P100, a transcriptional coactivator, is a human homologue of staphylococcal nuclease. Protein Sci. A Publ. Protein Soc. 1997, 6, 459–463. [Google Scholar] [CrossRef]
- Gutierrez-Beltran, E.; Denisenko, T.V.; Zhivotovsky, B.; Bozhkov, P.V. Tudor staphylococcal nuclease: Biochemistry and functions. Cell Death Differ. 2016, 23, 1739–1748. [Google Scholar] [CrossRef]
- Shaw, N.; Zhao, M.; Cheng, C.; Xu, H.; Saarikettu, J.; Li, Y.; Da, Y.; Yao, Z.; Silvennoinen, O.; Yang, J.; et al. The multifunctional human p100 protein ‘hooks’ methylated ligands. Nat. Struct. Mol. Biol. 2007, 14, 779–784. [Google Scholar] [CrossRef]
- Li, C.L.; Yang, W.Z.; Chen, Y.P.; Yuan, H.S. Structural and functional insights into human Tudor-SN, a key component linking RNA interference and editing. Nucleic. Acids Res. 2008, 36, 3579–3589. [Google Scholar] [CrossRef]
- Välineva, T.; Yang, J.; Palovuori, R.; Silvennoinen, O. The transcriptional co-activator protein p100 recruits histone acetyltransferase activity to STAT6 and mediates interaction between the CREB-binding protein and STAT6. J. Biol. Chem. 2005, 280, 14989–14996. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Välineva, T.; Hong, J.; Bu, T.; Yao, Z.; Jensen, O.N.; Frilander, M.J.; Silvennoinen, O. Transcriptional co-activator protein p100 interacts with snRNP proteins and facilitates the assembly of the spliceosome. Nucleic. Acids research 2007, 35, 4485–4494. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zhao, X.; Zhu, Y.; He, J.; Shao, J.; Su, C.; Zhang, Y.; Zhang, W.; Saarikettu, J.; Silvennoinen, O.; et al. Tudor staphylococcal nuclease (Tudor-SN) participates in small ribonucleoprotein (snRNP) assembly via interacting with symmetrically dimethylated Sm proteins. J. Biol. Chem. 2012, 287, 18130–18141. [Google Scholar] [CrossRef]
- Su, C.; Zhang, C.; Tecle, A.; Fu, X.; He, J.; Song, J.; Zhang, W.; Sun, X.; Ren, Y.; Silvennoinen, O.; et al. Tudor staphylococcal nuclease (Tudor-SN), a novel regulator facilitating G1/S phase transition, acting as a co-activator of E2F-1 in cell cycle regulation. J. Biol. Chem. 2015, 290, 7208–7220. [Google Scholar] [CrossRef]
- Gao, X.; Shi, X.; Fu, X.; Ge, L.; Zhang, Y.; Su, C.; Yang, X.; Silvennoinen, O.; Yao, Z.; He, J.; et al. Human Tudor staphylococcal nuclease (Tudor-SN) protein modulates the kinetics of AGTR1-3’UTR granule formation. FEBS Lett. 2014, 588, 2154–2161. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Fu, X.; Song, J.; Zhang, Y.; Cui, X.; Su, C.; Ge, L.; Shao, J.; Xin, L.; Saarikettu, J.; et al. Poly(A)(+) mRNA-binding protein Tudor-SN regulates stress granules aggregation dynamics. FEBS J. 2015, 282, 874–890. [Google Scholar] [CrossRef]
- Cui, X.; Zhang, X.; Liu, M.; Zhao, C.; Zhang, N.; Ren, Y.; Su, C.; Zhang, W.; Sun, X.; He, J.; et al. A pan-cancer analysis of the oncogenic role of staphylococcal nuclease domain-containing protein 1 (SND1) in human tumors. Genomics 2020, 112, 3958–3967. [Google Scholar] [CrossRef] [PubMed]
- Jariwala, N.; Rajasekaran, D.; Srivastava, J.; Gredler, R.; Akiel, M.A.; Robertson, C.L.; Emdad, L.; Fisher, P.B.; Sarkar, D. Role of the staphylococcal nuclease and tudor domain containing 1 in oncogenesis (review). Int. J. Oncol. 2015, 46, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Balian, A.; Hernandez, F.J. Nucleases as molecular targets for cancer diagnosis. Biomark. Res. 2021, 9, 86. [Google Scholar] [CrossRef] [PubMed]
- Jariwala, N.; Mendoza, R.G.; Garcia, D.; Lai, Z.; Subler, M.A.; Windle, J.J.; Mukhopadhyay, N.D.; Fisher, P.B.; Chen, Y.; Sarkar, D. Posttranscriptional Inhibition of Protein Tyrosine Phosphatase Nonreceptor Type 23 by Staphylococcal Nuclease and Tudor Domain Containing 1: Implications for Hepatocellular Carcinoma. Hepatol. Commun. 2019, 3, 1258–1270. [Google Scholar] [CrossRef] [PubMed]
- Newell, K.A.; Ellenhorn, J.D.; Bruce, D.S.; Bluestone, J.A. In vivo T-cell activation by staphylococcal enterotoxin B prevents outgrowth of a malignant tumor. Proc. Natl. Acad. Sci. USA 1991, 88, 1074–1078. [Google Scholar] [CrossRef] [PubMed]
- Dohlsten, M.; Hedlund, G.; Akerblom, E.; Lando, P.A.; Kalland, T. Monoclonal antibody-targeted superantigens: A different class of anti-tumor agents. Proc. Natl. Acad. Sci. USA 1991, 88, 9287–9291. [Google Scholar] [CrossRef]
- Jobbins, J.; Bagg, J.; Parsons, K.; Finlay, I.; Addy, M.; Newcombe, R.G. Oral carriage of yeasts, coliforms and staphylococci in patients with advanced malignant disease. J. Oral. Pathol. Med. 1992, 21, 305–308. [Google Scholar] [CrossRef]
- Dohlsten, M.; Lando, P.A.; Björk, P.; Abrahmsén, L.; Ohlsson, L.; Lind, P.; Kalland, T. Immunotherapy of human colon cancer by antibody-targeted superantigens. Cancer Immunol. Immunother. 1995, 41, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H.; Takeuchi, H.; Nishitani, N.; Yamanaka, H.; Suzuki, K.; Kurusu, M.; Suganuma, M. Carcinogenic potential of tobacco tar-resistant Staphylococcus aureus in buccal cavity. J. Cancer Res. Clin. Oncol. 2004, 130, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Perabo, F.G.; Willert, P.L.; Wirger, A.; Schmidt, D.H.; Von Ruecker, A.; Mueller, S.C. Superantigen-activated mononuclear cells induce apoptosis in transitional cell carcinoma. Anticancer. Res. 2005, 25, 3565–3573. [Google Scholar] [PubMed]
- Edey, A.J.; Bentley, P.G.; Garrett, J.P.; Liebmann, R.D. Ductal breast carcinoma presenting with methicillin-resistant Staphylococcus aureus mastitis. Breast J. 2005, 11, 491–492. [Google Scholar] [CrossRef]
- Jiang, Y.Q.; Wang, H.R.; Li, H.P.; Hao, H.J.; Zheng, Y.L.; Gu, J. Targeting of hepatoma cell and suppression of tumor growth by a novel 12mer peptide fused to superantigen TSST-1. Mol. Med. 2006, 12, 81–87. [Google Scholar] [CrossRef]
- Schneider, D.; Liaw, L.; Daniel, C.; Athanasopoulos, A.N.; Herrmann, M.; Preissner, K.T.; Nawroth, P.P.; Chavakis, T. Inhibition of breast cancer cell adhesion and bone metastasis by the extracellular adherence protein of Staphylococcus aureus. Biochem. Biophys. Res. Commun. 2007, 357, 282–288. [Google Scholar] [CrossRef]
- Napeñas, J.J.; Brennan, M.T.; Bahrani-Mougeot, F.K.; Fox, P.C.; Lockhart, P.B. Relationship between mucositis and changes in oral microflora during cancer chemotherapy. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2007, 103, 48–59. [Google Scholar] [CrossRef]
- Noguchi, N.; Ohashi, T.; Shiratori, T.; Narui, K.; Hagiwara, T.; Ko, M.; Watanabe, K.; Miyahara, T.; Taira, S.; Moriyasu, F.; et al. Association of tannase-producing Staphylococcus lugdunensis with colon cancer and characterization of a novel tannase gene. J. Gastroenterol. 2007, 42, 346–351. [Google Scholar] [CrossRef]
- Szabados, F.; Kleine, B.; Anders, A.; Kaase, M.; Sakinç, T.; Schmitz, I.; Gatermann, S. Staphylococcus saprophyticus ATCC 15305 is internalized into human urinary bladder carcinoma cell line 5637. FEMS Microbiol. Lett. 2008, 285, 163–169. [Google Scholar] [CrossRef]
- Johansson, D.; Johansson, A.; Behnam-Motlagh, P. alpha-Toxin of Staphylococcus aureus overcomes acquired cisplatin-resistance in malignant mesothelioma cells. Cancer Lett. 2008, 265, 67–75. [Google Scholar] [CrossRef]
- Kalita, O.; Kala, M.; Svebisova, H.; Ehrmann, J.; Hlobilkova, A.; Trojanec, R.; Hajduch, M.; Houdek, M. Glioblastoma multiforme with an abscess: Case report and literature review. J. Neurooncol. 2008, 88, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Kullander, J.; Forslund, O.; Dillner, J. Staphylococcus aureus and squamous cell carcinoma of the skin. Cancer Epidemiol. Biomark. Prev. 2009, 18, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Tian, R.; Xiu, B.; Yan, J.; Jia, R.; Zhang, L.; Chang, A.E.; Song, H.; Li, Q. Antitumor activity of T cells generated from lymph nodes draining the SEA-expressing murine B16 melanoma and secondarily activated with dendritic cells. Int. J. Biol. Sci. 2009, 5, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Walenkamp, A.M.; Boer, I.G.; Bestebroer, J.; Rozeveld, D.; Timmer-Bosscha, H.; Hemrika, W.; van Strijp, J.A.; de Haas, C.J. Staphylococcal superantigen-like 10 inhibits CXCL12-induced human tumor cell migration. Neoplasia 2009, 11, 333–344. [Google Scholar] [CrossRef]
- Xie, W.; Huang, Y.; Xie, W.; Guo, A.; Wu, W. Bacteria peptidoglycan promoted breast cancer cell invasiveness and adhesiveness by targeting toll-like receptor 2 in the cancer cells. PLoS ONE 2010, 5, e10850. [Google Scholar] [CrossRef]
- Yoo, B.K.; Santhekadur, P.K.; Gredler, R.; Chen, D.; Emdad, L.; Bhutia, S.; Pannell, L.; Fisher, P.B.; Sarkar, D. Increased RNA-induced silencing complex (RISC) activity contributes to hepatocellular carcinoma. Hepatology 2011, 53, 1538–1548. [Google Scholar] [CrossRef]
- Apostolou, P.; Tsantsaridou, A.; Papasotiriou, I.; Toloudi, M.; Chatziioannou, M.; Giamouzis, G. Bacterial and fungal microflora in surgically removed lung cancer samples. J. Cardiothorac. Surg. 2011, 6, 137. [Google Scholar] [CrossRef] [PubMed]
- Panghal, M.; Kaushal, V.; Kadayan, S.; Yadav, J.P. Incidence and risk factors for infection in oral cancer patients undergoing different treatments protocols. BMC Oral Health 2012, 12, 22. [Google Scholar] [CrossRef]
- Reis, L.O.; Ferreira, U.; Billis, A.; Cagnon, V.H.; Fávaro, W.J. Anti-angiogenic effects of the superantigen staphylococcal enterotoxin B and bacillus Calmette-Guérin immunotherapy for nonmuscle invasive bladder cancer. J. Urol. 2012, 187, 438–445. [Google Scholar] [CrossRef]
- Gu, L.; Yue, J.; Zheng, Y.; Zheng, X.; Wang, J.; Wang, Y.; Li, J.; Jiang, Y.; Jiang, H. Evaluation of a recombinant double mutant of staphylococcal enterotoxin B (SEB-H32Q/K173E) with enhanced antitumor activity effects and decreased pyrexia. PLoS ONE 2013, 8, e55892. [Google Scholar] [CrossRef]
- Alreshidi, M.A.; Alsalamah, A.A.; Hamat, R.A.; Neela, V.; Alshrari, A.S.; Atshan, S.S.; Alajlan, H.H.; Nor Shamsudin, M. Genetic variation among methicillin-resistant Staphylococcus aureus isolates from cancer patients in Saudi Arabia. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2013, 32, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Hao, L.; Chen, M.; Hu, J.; Shi, Z.; Zhang, Z.; Dong, B.; Fu, Y.; Pei, C.; Wu, Y. Target expression of Staphylococcus enterotoxin A from an oncolytic adenovirus suppresses mouse bladder tumor growth and recruits CD3+ T cell. Tumour. Biol. 2013, 34, 2863–2869. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Sun, X.; Lu, L.; Zheng, J.B.; Tian, Y.; Wang, W. Cytotoxicity of lymphocytes activated by superantigen toxic-shock-syndrome toxin-1 against colorectal cancer LoVo cells. Mol. Cell. Biochem. 2013, 376, 1–9. [Google Scholar] [CrossRef]
- Memmel, E.; Homann, A.; Oelschlaeger, T.A.; Seibel, J. Metabolic glycoengineering of Staphylococcus aureus reduces its adherence to human T24 bladder carcinoma cells. Chem. Commun. 2013, 49, 7301–7303. [Google Scholar] [CrossRef] [PubMed]
- Bu, S.; Xie, Q.; Chang, W.; Huo, X.; Chen, F.; Ma, X. LukS-PV induces mitochondrial-mediated apoptosis and G0/G1 cell cycle arrest in human acute myeloid leukemia THP-1 cells. Int. J. Biochem. Cell Biol. 2013, 45, 1531–1537. [Google Scholar] [CrossRef] [PubMed]
- Terman, D.S.; Serier, A.; Dauwalder, O.; Badiou, C.; Dutour, A.; Thomas, D.; Brun, V.; Bienvenu, J.; Etienne, J.; Vandenesch, F.; et al. Staphylococcal entertotoxins of the enterotoxin gene cluster (egcSEs) induce nitrous oxide- and cytokine dependent tumor cell apoptosis in a broad panel of human tumor cells. Front. Cell. Infect. Microbiol. 2013, 3, 38. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Liu, L.; Xu, M.; Zhang, H.; Zhang, Y.; Zhang, C. T-cell proliferation and antitumour activities of a truncated mutant of staphylococcal enterotoxin C2 with decreased cytokine secretion. J. Med. Microbiol. 2013, 62, 451–456. [Google Scholar] [CrossRef] [PubMed]
- St Jean, A.T.; Swofford, C.A.; Panteli, J.T.; Brentzel, Z.J.; Forbes, N.S. Bacterial delivery of Staphylococcus aureus alpha-hemolysin causes regression and necrosis in murine tumors. Mol. Ther. 2014, 22, 1266–1274. [Google Scholar] [CrossRef] [PubMed]
- Krejsgaard, T.; Willerslev-Olsen, A.; Lindahl, L.M.; Bonefeld, C.M.; Koralov, S.B.; Geisler, C.; Wasik, M.A.; Gniadecki, R.; Kilian, M.; Iversen, L.; et al. Staphylococcal enterotoxins stimulate lymphoma-associated immune dysregulation. Blood 2014, 124, 761–770. [Google Scholar] [CrossRef]
- Shan, W.; Bu, S.; Zhang, C.; Zhang, S.; Ding, B.; Chang, W.; Dai, Y.; Shen, J.; Ma, X. LukS-PV, a component of Panton-Valentine leukocidin, exerts potent activity against acute myeloid leukemia in vitro and in vivo. Int. J. Biochem. Cell Biol. 2015, 61, 20–28. [Google Scholar] [CrossRef]
- Boer, J.C.; van Marion, D.M.; Joseph, J.V.; Kliphuis, N.M.; Timmer-Bosscha, H.; van Strijp, J.A.; de Vries, E.G.; den Dunnen, W.F.; Kruyt, F.A.; Walenkamp, A.M. Microenvironment involved in FPR1 expression by human glioblastomas. J. Neurooncol. 2015, 123, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, R.; Pesce, M.; Franchelli, S.; Baldelli, I.; De Maria, A.; Marchese, A. Phenotypic and genotypic characterization of staphylococci causing breast peri-implant infections in oncologic patients. BMC Microbiol. 2015, 15, 26. [Google Scholar] [CrossRef]
- Rodríguez-Serrano, F.; Mut-Salud, N.; Cruz-Bustos, T.; Gomez-Samblas, M.; Carrasco, E.; Garrido, J.M.; López-Jaramillo, F.J.; Santoyo-Gonzalez, F.; Osuna, A. Functionalized immunostimulating complexes with protein A via lipid vinyl sulfones to deliver cancer drugs to trastuzumab-resistant HER2-overexpressing breast cancer cells. Int. J. Nanomed. 2016, 11, 4777–4785. [Google Scholar] [CrossRef]
- Yu, Q.T.; Meng, Z.B. Treatment of advanced breast cancer with a combination of highly agglutinative staphylococcin and vinorelbine-based chemotherapy. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 3465–3468. [Google Scholar] [PubMed]
- Mu, P.; Dong, B.; Liang, P.; Yu, X.; Su, L.; Zhang, J. Clinical research on ultrasonically guided intrahepatic injections of HAS in interventional treatment of liver carcinomas. J. BU ON Off. J. Balk. Union Oncol. 2016, 21, 1394–1397. [Google Scholar]
- Almeida, J.F.; Breyner, N.M.; Mahi, M.; Ahmed, B.; Benbouziane, B.; Boas, P.C.; Miyoshi, A.; Azevedo, V.; Langella, P.; Bermúdez-Humarán, L.G.; et al. Expression of fibronectin binding protein A (FnBPA) from Staphylococcus aureus at the cell surface of Lactococcus lactis improves its immunomodulatory properties when used as protein delivery vector. Vaccine 2016, 34, 1312–1318. [Google Scholar] [CrossRef]
- Hassan, Z.; Mustafa, S.; Rahim, R.A.; Isa, N.M. Anti-breast cancer effects of live, heat-killed and cytoplasmic fractions of Enterococcus faecalis and Staphylococcus hominis isolated from human breast milk. In Vitro Cell. Dev. Biol. Anim. 2016, 52, 337–348. [Google Scholar] [CrossRef]
- Urbaniak, C.; Gloor, G.B.; Brackstone, M.; Scott, L.; Tangney, M.; Reid, G. The Microbiota of Breast Tissue and Its Association with Breast Cancer. Appl. Environ. Microbiol. 2016, 82, 5039–5048. [Google Scholar] [CrossRef]
- Hu, H.; Johani, K.; Almatroudi, A.; Vickery, K.; Van Natta, B.; Kadin, M.E.; Brody, G.; Clemens, M.; Cheah, C.Y.; Lade, S.; et al. Bacterial Biofilm Infection Detected in Breast Implant-Associated Anaplastic Large-Cell Lymphoma. Plast. Reconstr. Surg. 2016, 137, 1659–1669. [Google Scholar] [CrossRef]
- Akbari, A.; Farahnejad, Z.; Akhtari, J.; Abastabar, M.; Mobini, G.R.; Mehbod, A.S. Staphylococcus aureus Enterotoxin B Down-Regulates the Expression of Transforming Growth Factor-Beta (TGF-β) Signaling Transducers in Human Glioblastoma. Jundishapur. J. Microbiol. 2016, 9, e27297. [Google Scholar] [CrossRef]
- Nesher, L.; Tarrand, J.; Chemaly, R.F.; Rolston, K.V. Staphylococcus lugdunensis infections, filling in the gaps: A 3-year retrospective review from a comprehensive cancer center. Support. Care Cancer 2017, 25, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Altemus, J.; Niazi, F.; Green, H.; Calhoun, B.C.; Sturgis, C.; Grobmyer, S.R.; Eng, C. Breast tissue, oral and urinary microbiomes in breast cancer. Oncotarget 2017, 8, 88122–88138. [Google Scholar] [CrossRef] [PubMed]
- Hattar, K.; Reinert, C.P.; Sibelius, U.; Gökyildirim, M.Y.; Subtil, F.S.B.; Wilhelm, J.; Eul, B.; Dahlem, G.; Grimminger, F.; Seeger, W.; et al. Lipoteichoic acids from Staphylococcus aureus stimulate proliferation of human non-small-cell lung cancer cells in vitro. Cancer Immunol. Immunother. 2017, 66, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Jariwala, N.; Rajasekaran, D.; Mendoza, R.G.; Shen, X.N.; Siddiq, A.; Akiel, M.A.; Robertson, C.L.; Subler, M.A.; Windle, J.J.; Fisher, P.B.; et al. Oncogenic Role of SND1 in Development and Progression of Hepatocellular Carcinoma. Cancer Res. 2017, 77, 3306–3316. [Google Scholar] [CrossRef]
- Wu, P.; Zhang, G.; Zhao, J.; Chen, J.; Chen, Y.; Huang, W.; Zhong, J.; Zeng, J. Profiling the Urinary Microbiota in Male Patients With Bladder Cancer in China. Front. Cell. Infect. Microbiol. 2018, 8, 167. [Google Scholar] [CrossRef]
- Mathews, J.; Patel, M. Bacterial endotoxins and microorganisms in the oral cavities of patients on cancer therapy. Microb Pathog 2018, 123, 190–195. [Google Scholar] [CrossRef]
- Villafuerte, K.R.V.; Martinez, C.J.H.; Dantas, F.T.; Carrara, H.H.A.; Dos Reis, F.J.C.; Palioto, D.B. The impact of chemotherapeutic treatment on the oral microbiota of patients with cancer: A systematic review. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2018, 125, 552–566. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chen, T.H.; Butcher, A.M.; Trzoss, L.L.; Nam, S.J.; Shirakawa, K.T.; Zhou, W.; Oh, J.; Otto, M.; Fenical, W.; et al. A commensal strain of Staphylococcus epidermidis protects against skin neoplasia. Sci. Adv. 2018, 4, eaao4502. [Google Scholar] [CrossRef]
- Noguchi, N.; Fukuzawa, M.; Wajima, T.; Yokose, K.; Suzuki, M.; Nakaminami, H.; Kawai, T.; Moriyasu, F.; Sasatsu, M. Specific clones of Staphylococcus lugdunensis may be associated with colon carcinoma. J. Infect. Public Health 2018, 11, 39–42. [Google Scholar] [CrossRef]
- Walker, J.N.; Hanson, B.M.; Pinkner, C.L.; Simar, S.R.; Pinkner, J.S.; Parikh, R.; Clemens, M.W.; Hultgren, S.J.; Myckatyn, T.M. Insights into the Microbiome of Breast Implants and Periprosthetic Tissue in Breast Implant-Associated Anaplastic Large Cell Lymphoma. Sci. Rep. 2019, 9, 10393. [Google Scholar] [CrossRef]
- Shehata, M.M.K.; Radwan, S.M.; Ali, S.A.M. Effects of gamma-irradiation on antibiotic resistance and diagnostic molecular markers of methicillin-resistant Staphylococcus aureus in Egyptian cancer patients. Int. J. Radiat. Biol. 2019, 95, 1728–1743. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhang, J.; Fang, S.; Zhang, M.; Liu, S.; Tian, Y.; Ma, M.; Liu, F.; Jin, G. Inflammatory activation of microglia by Staphylococcus aureus caused phenotypic alterations and affected glioblastoma growth. Cell Biochem. Funct. 2019, 37, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Madhusudhan, N.; Pausan, M.R.; Halwachs, B.; Durdević, M.; Windisch, M.; Kehrmann, J.; Patra, V.; Wolf, P.; Boukamp, P.; Moissl-Eichinger, C.; et al. Molecular Profiling of Keratinocyte Skin Tumors Links Staphylococcus aureus Overabundance and Increased Human β-Defensin-2 Expression to Growth Promotion of Squamous Cell Carcinoma. Cancers 2020, 12, 541. [Google Scholar] [CrossRef] [PubMed]
- Fourdrain, A.; Bouabdallah, I.; Gust, L.; Cassir, N.; Brioude, G.; Falcoz, P.E.; Alifano, M.; Le Rochais, J.P.; D’Annoville, T.; Trousse, D.; et al. Screening and topical decolonization of preoperative nasal Staphylococcus aureus carriers to reduce the incidence of postoperative infections after lung cancer surgery: A propensity matched study. Interact. Cardiovasc. Thorac. Surg. 2020, 30, 552–558. [Google Scholar] [CrossRef]
- Qiang, Y.; Ma, F.; Wang, Z.; Nie, Z.; Xu, L.; Ding, P.; Ma, X. LukS-PV induces cell cycle arrest and apoptosis through p38/ERK MAPK signaling pathway in NSCLC cells. Biochem. Biophys. Res. Commun. 2020, 521, 846–852. [Google Scholar] [CrossRef]
- Zhao, C.C.; Yu, W.W.; Qi, Y.J.; Xu, L.F.; Wang, Z.R.; Qiang, Y.W.; Ma, F.; Ma, X.L. Quantitative proteomic analysis reveals that Luks-PV exerts antitumor activity by regulating the key proteins and metabolic pathways in HepG2 cells. Anti-Cancer Drugs 2020, 31, 223–230. [Google Scholar] [CrossRef]
- Thyagarajan, S.; Zhang, Y.; Thapa, S.; Allen, M.S.; Phillips, N.; Chaudhary, P.; Kashyap, M.V.; Vishwanatha, J.K. Comparative analysis of racial differences in breast tumor microbiome. Sci Rep 2020, 10, 14116. [Google Scholar] [CrossRef]
- Klann, E.; Williamson, J.M.; Tagliamonte, M.S.; Ukhanova, M.; Asirvatham, J.R.; Chim, H.; Yaghjyan, L.; Mai, V. Microbiota composition in bilateral healthy breast tissue and breast tumors. Cancer Causes Control 2020, 31, 1027–1038. [Google Scholar] [CrossRef]
- Chiba, A.; Bawaneh, A.; Velazquez, C.; Clear, K.Y.J.; Wilson, A.S.; Howard-McNatt, M.; Levine, E.A.; Levi-Polyachenko, N.; Yates-Alston, S.A.; Diggle, S.P.; et al. Neoadjuvant Chemotherapy Shifts Breast Tumor Microbiota Populations to Regulate Drug Responsiveness and the Development of Metastasis. Mol. Cancer Res. 2020, 18, 130–139. [Google Scholar] [CrossRef]
- Gotland, N.; Uhre, M.L.; Sandholdt, H.; Mejer, N.; Lundbo, L.F.; Petersen, A.; Larsen, A.R.; Benfield, T. Increased risk of incident primary cancer after Staphylococcus aureus bacteremia: A matched cohort study. Medicine 2020, 99, e19984. [Google Scholar] [CrossRef]
- Qi, J.L.; He, J.R.; Liu, C.B.; Jin, S.M.; Gao, R.Y.; Yang, X.; Bai, H.M.; Ma, Y.B. Pulmonary Staphylococcus aureus infection regulates breast cancer cell metastasis via neutrophil extracellular traps (NETs) formation. MedComm 2020, 1, 188–201. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, S.; Barzegari, A.; Esmaeili, A.; Omidi, Y. Designing a light-activated recombinant alpha hemolysin for colorectal cancer targeting. BioImpacts BI 2020, 10, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Khusro, A.; Aarti, C.; Mahizhaveni, B.; Dusthackeer, A.; Agastian, P.; Esmail, G.A.; Ghilan, A.M.; Al-Dhabi, N.A.; Arasu, M.V. Purification and characterization of anti-tubercular and anticancer protein from Staphylococcus hominis strain MANF2: In silico structural and functional insight of peptide. Saudi. J. Biol. Sci. 2020, 27, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Safarpour-Dehkordi, M.; Doosti, A.; Jami, M.S. Integrative Analysis of lncRNAs in Kidney Cancer to Discover A New lncRNA (LINC00847) as A Therapeutic Target for Staphylococcal Enterotoxin tst Gene. Cell J. 2020, 22, 101–109. [Google Scholar] [CrossRef]
- Chiappini, A.; Santos, A.N.; DE Trizio, I.; Croci, D.; Valci, L.; Reinert, M.; Marchi, F. Longer survival of glioblastoma complicated by bacterial infections after surgery: What is known today. J. Neurosurg. Sci. 2021, 65, 524–531. [Google Scholar] [CrossRef] [PubMed]
- El Dine, R.S.; Elfaky, M.A.; Asfour, H.; El Halawany, A.M. Anti-adhesive activity of Aframomum melegueta major phenolics on lower respiratory tract pathogens. Nat. Prod. Res. 2021, 35, 539–547. [Google Scholar] [CrossRef]
- Tzeng, A.; Sangwan, N.; Jia, M.; Liu, C.C.; Keslar, K.S.; Downs-Kelly, E.; Fairchild, R.L.; Al-Hilli, Z.; Grobmyer, S.R.; Eng, C. Human breast microbiome correlates with prognostic features and immunological signatures in breast cancer. Genome Med. 2021, 13, 60. [Google Scholar] [CrossRef]
- Parra-Grande, M.; Oré-Arce, M.; Martínez-Priego, L.; D’Auria, G.; Rosselló-Mora, R.; Lillo, M.; Sempere, A.; Lumbreras, B.; Sánchez-Hellín, V. Profiling the Bladder Microbiota in Patients With Bladder Cancer. Front. Microbiol. 2021, 12, 718776. [Google Scholar] [CrossRef]
- Li, Z.; Zhuang, H.; Wang, G.; Wang, H.; Dong, Y. Prevalence, predictors, and mortality of bloodstream infections due to methicillin-resistant Staphylococcus aureus in patients with malignancy: Systemic review and meta-analysis. BMC Infect. Dis. 2021, 21, 74. [Google Scholar] [CrossRef]
- Yang, J.; He, P.; Zhou, M.; Li, S.; Zhang, J.; Tao, X.; Wang, A.; Wu, X. Variations in oral microbiome and its predictive functions between tumorous and healthy individuals. J. Med. Microbiol. 2022, 71, 001568. [Google Scholar] [CrossRef]
- An, J.; Kwon, H.; Lim, W.; Moon, B.I. Staphylococcus aureus-Derived Extracellular Vesicles Enhance the Efficacy of Endocrine Therapy in Breast Cancer Cells. J. Clin. Med. 2022, 11, 2030. [Google Scholar] [CrossRef] [PubMed]
- Maślak, E.; Miśta, W.; Złoch, M.; Błońska, D.; Pomastowski, P.; Monedeiro, F.; Buszewski, B.; Mrochem-Kwarciak, J.; Bojarska, K.; Gabryś, D. A New Approach to Imaging and Rapid Microbiome Identification for Prostate Cancer Patients Undergoing Radiotherapy. Biomedicines 2022, 10, 1806. [Google Scholar] [CrossRef]
- Worku, M.; Belay, G.; Tigabu, A. Bacterial profile and antimicrobial susceptibility patterns in cancer patients. PLoS ONE 2022, 17, e0266919. [Google Scholar] [CrossRef] [PubMed]
- Mansour, B.; Monyók, Á.; Gajdács, M.; Stercz, B.; Makra, N.; Pénzes, K.; Vadnay, I.; Szabó, D.; Ostorházi, E. Bladder Tissue Microbiome Composition in Patients of Bladder Cancer or Benign Prostatic Hyperplasia and Related Human Beta Defensin Levels. Biomedicines 2022, 10, 1758. [Google Scholar] [CrossRef]
- Xu, X.; Ding, P.; Shi, L.; Wu, G.; Ma, X. LukS-PV inhibits hepatocellular carcinoma cells migration by downregulating HDAC6 expression. BMC Cancer 2022, 22, 630. [Google Scholar] [CrossRef]
- Abbasi Montazeri, E.; Khosravi, A.D.; Khazaei, S.; Sabbagh, A. Prevalence of methicillin resistance and superantigenic toxins in Staphylococcus aureus strains isolated from patients with cancer. BMC Microbiol. 2021, 21, 262. [Google Scholar] [CrossRef]
- Nanayakkara, A.K.; Boucher, H.W.; Fowler, V.G., Jr.; Jezek, A.; Outterson, K.; Greenberg, D.E. Antibiotic resistance in the patient with cancer: Escalating challenges and paths forward. CA A Cancer J. Clin. 2021, 71, 488–504. [Google Scholar] [CrossRef]
- Wang, K.; Nakano, K.; Naderi, N.; Bajaj-Elliott, M.; Mosahebi, A. Is the skin microbiota a modifiable risk factor for breast disease?: A systematic review. Breast 2021, 59, 279–285. [Google Scholar] [CrossRef]
- Fujii, K. Pathogenesis of cutaneous T cell lymphoma: Involvement of Staphylococcus aureus. J. Dermatol. 2022, 49, 202–209. [Google Scholar] [CrossRef]
- Squarzanti, D.F.; Zavattaro, E.; Pizzimenti, S.; Amoruso, A.; Savoia, P.; Azzimonti, B. Non-Melanoma Skin Cancer: News from microbiota research. Crit. Rev. Microbiol. 2020, 46, 433–449. [Google Scholar] [CrossRef]
- Richardson, B.N.; Lin, J.; Buchwald, Z.S.; Bai, J. Skin Microbiome and Treatment-Related Skin Toxicities in Patients With Cancer: A Mini-Review. Front. Oncol. 2022, 12, 924849. [Google Scholar] [CrossRef] [PubMed]
- Wagstaffe, S.J.; Hill, K.E.; Williams, D.W.; Randle, B.J.; Thomas, D.W.; Stephens, P.; Riley, D.J. Bispecific antibody-mediated detection of the Staphylococcus aureus thermonuclease. Anal. Chem. 2012, 84, 5876–5884. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Chan, S.C.; Leung, K.W.; Wu, J.M.; Fang, H.J.; Tsong, T.Y. Local stability identification and the role of key acidic amino acid residues in staphylococcal nuclease unfolding. FEBS J. 2005, 272, 3967–3974. [Google Scholar] [CrossRef] [PubMed]
- Lai, B.; Gao, W.; Cui, K.; Xie, W.; Tang, Q.; Jin, W.; Hu, G.; Ni, B.; Zhao, K. Principles of nucleosome organization revealed by single-cell micrococcal nuclease sequencing. Nature 2018, 562, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Zhang, L.; Ruan, H.; Li, G. Histone variant H2A.Z regulates nucleosome unwrapping and CTCF binding in mouse ES cells. Nucleic. Acids Res. 2020, 48, 5939–5952. [Google Scholar] [CrossRef] [PubMed]
- Liénard, P.; Rivière, M.; Van Vooren, P.; Szpirer, C.; Szpirer, J. Assignment of SND1, the gene encoding coactivator p100, to human chromosome 7q31.3 and rat chromosome 4q23 by in situ hybridization. Cytogenet. Cell Genet. 2000, 90, 253–254. [Google Scholar] [CrossRef]
- Ochoa, B.; Chico, Y.; Martínez, M.J. Insights Into SND1 Oncogene Promoter Regulation. Front. Oncol. 2018, 8, 606. [Google Scholar] [CrossRef]
- Callebaut, I.; Mornon, J.P. The human EBNA-2 coactivator p100: Multidomain organization and relationship to the staphylococcal nuclease fold and to the tudor protein involved in Drosophila melanogaster development. Biochem. J. 1997, 321 Pt 1, 125–132. [Google Scholar] [CrossRef]
- Hu, Y.; Meng, J.; Shi, C.; Hervin, K.; Fratamico, P.M.; Shi, X. Characterization and comparative analysis of a second thermonuclease from Staphylococcus aureus. Microbiol. Res. 2013, 168, 174–182. [Google Scholar] [CrossRef]
- Hynes, T.R.; Fox, R.O. The crystal structure of staphylococcal nuclease refined at 1.7 A resolution. Proteins 1991, 10, 92–105. [Google Scholar] [CrossRef]
- Hossain, M.J.; Korde, R.; Singh, S.; Mohmmed, A.; Dasaradhi, P.V.; Chauhan, V.S.; Malhotra, P. Tudor domain proteins in protozoan parasites and characterization of Plasmodium falciparum tudor staphylococcal nuclease. Int. J. Parasitol. 2008, 38, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.L.; Tian, L.; Kumamaru, T.; Hamada, S.; Okita, T.W. Multifunctional RNA Binding Protein OsTudor-SN in Storage Protein mRNA Transport and Localization. Plant Physiol. 2017, 175, 1608–1623. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Yan, Z.; Wang, Y.; Yan, X.; Han, Y. Tudor-SN, a component of stress granules, regulates growth under salt stress by modulating GA20ox3 mRNA levels in Arabidopsis. J. Exp. Bot. 2014, 65, 5933–5944. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.J.; Korde, R.; Singh, P.K.; Kanodia, S.; Ranjan, R.; Ram, G.; Kalsey, G.S.; Singh, R.; Malhotra, P. Plasmodium falciparum Tudor Staphylococcal Nuclease interacting proteins suggest its role in nuclear as well as splicing processes. Gene 2010, 468, 48–57. [Google Scholar] [CrossRef]
- Ascano, M.; Hafner, M.; Cekan, P.; Gerstberger, S.; Tuschl, T. Identification of RNA-protein interaction networks using PAR-CLIP. Wiley Interdiscip. Reviews. RNA 2012, 3, 159–177. [Google Scholar] [CrossRef]
- Baltz, A.G.; Munschauer, M.; Schwanhäusser, B.; Vasile, A.; Murakawa, Y.; Schueler, M.; Youngs, N.; Penfold-Brown, D.; Drew, K.; Milek, M.; et al. The mRNA-bound proteome and its global occupancy profile on protein-coding transcripts. Mol. Cell 2012, 46, 674–690. [Google Scholar] [CrossRef]
- Cuatrecasas, P.; Fuchs, S.; Anfinsen, C.B. The binding of nucleotides and calcium to the extracellular nuclease of Staphylococcus aureus. Studies by gel filtration. J. Biol. Chem. 1967, 242, 3063–3067. [Google Scholar] [CrossRef]
- Yang, W.; Chendrimada, T.P.; Wang, Q.; Higuchi, M.; Seeburg, P.H.; Shiekhattar, R.; Nishikura, K. Modulation of microRNA processing and expression through RNA editing by ADAR deaminases. Nat. Struct. Mol. Biol. 2006, 13, 13–21. [Google Scholar] [CrossRef]
- Scadden, A.D. The RISC subunit Tudor-SN binds to hyper-edited double-stranded RNA and promotes its cleavage. Nat. Struct. Mol. Biol. 2005, 12, 489–496. [Google Scholar] [CrossRef]
- Ayllón, N.; Naranjo, V.; Hajdušek, O.; Villar, M.; Galindo, R.C.; Kocan, K.M.; Alberdi, P.; Šíma, R.; Cabezas-Cruz, A.; Rückert, C.; et al. Nuclease Tudor-SN Is Involved in Tick dsRNA-Mediated RNA Interference and Feeding but Not in Defense against Flaviviral or Anaplasma phagocytophilum Rickettsial Infection. PLoS ONE 2015, 10, e0133038. [Google Scholar] [CrossRef]
- Elbarbary, R.A.; Miyoshi, K.; Myers, J.R.; Du, P.; Ashton, J.M.; Tian, B.; Maquat, L.E. Tudor-SN-mediated endonucleolytic decay of human cell microRNAs promotes G(1)/S phase transition. Science 2017, 356, 859–862. [Google Scholar] [CrossRef] [PubMed]
- Phetrungnapha, A.; Panyim, S.; Ongvarrasopone, C. Penaeus monodon Tudor staphylococcal nuclease preferentially interacts with N-terminal domain of Argonaute-1. Fish Shellfish Immunol. 2013, 34, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Phetrungnapha, A.; Panyim, S.; Ongvarrasopone, C. A Tudor staphylococcal nuclease from Penaeus monodon: cDNA cloning and its involvement in RNA interference. Fish Shellfish Immunol. 2011, 31, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Musiyenko, A.; Majumdar, T.; Andrews, J.; Adams, B.; Barik, S. PRMT1 methylates the single Argonaute of Toxoplasma gondii and is important for the recruitment of Tudor nuclease for target RNA cleavage by antisense guide RNA. Cell. Microbiol. 2012, 14, 882–901. [Google Scholar] [CrossRef] [PubMed]
- Sheweita, S.A.; Alsamghan, A.S. Molecular Mechanisms Contributing Bacterial Infections to the Incidence of Various Types of Cancer. Mediat. Inflamm. 2020, 2020, 4070419. [Google Scholar] [CrossRef] [PubMed]
- Dharmaraja, A.T. Role of Reactive Oxygen Species (ROS) in Therapeutics and Drug Resistance in Cancer and Bacteria. J. Med. Chem. 2017, 60, 3221–3240. [Google Scholar] [CrossRef]
- Shivaee, A.; Sedighi, M.; Imani Fooladi, A.A. Staphylococcal enterotoxins as good candidates for cancer immunotherapy: A systematic review. Ann. Di Ig. Med. Prev. E Di Comunita 2020, 32, 648–663. [Google Scholar] [CrossRef]
- Terman, D.S.; Bohach, G.; Vandenesch, F.; Etienne, J.; Lina, G.; Sahn, S.A. Staphylococcal superantigens of the enterotoxin gene cluster (egc) for treatment of stage IIIb non-small cell lung cancer with pleural effusion. Clin. Chest Med. 2006, 27, 321–334. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, X.; Rao, X. Apoptosis induced by Staphylococcus aureus toxins. Microbiol. Res. 2017, 205, 19–24. [Google Scholar] [CrossRef]
- Dohlsten, M.; Abrahmsén, L.; Björk, P.; Lando, P.A.; Hedlund, G.; Forsberg, G.; Brodin, T.; Gascoigne, N.R.; Förberg, C.; Lind, P.; et al. Monoclonal antibody-superantigen fusion proteins: Tumor-specific agents for T-cell-based tumor therapy. Proc. Natl. Acad. Sci. USA 1994, 91, 8945–8949. [Google Scholar] [CrossRef]
- Golob-Urbanc, A.; Rajčević, U.; Strmšek, Ž.; Jerala, R. Design of split superantigen fusion proteins for cancer immunotherapy. J. Biol. Chem. 2019, 294, 6294–6305. [Google Scholar] [CrossRef] [PubMed]
- Perabo, F.G.; Willert, P.L.; Wirger, A.; Schmidt, D.H.; Wardelmann, E.; Sitzia, M.; von Ruecker, A.; Mueller, S.C. Preclinical evaluation of superantigen (staphylococcal enterotoxin B) in the intravesical immunotherapy of superficial bladder cancer. Int. J. Cancer 2005, 115, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.L.; Yan, Z.; Chen, J.; Zhao, W.H.; Guo, W. Clinical application of highly agglutinative staphylococcin in cancer treatment updates of the literature. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2718–2725. [Google Scholar] [PubMed]
- Fu, X.; Xu, M.; Zhang, H.; Li, Y.; Li, Y.; Zhang, C. Staphylococcal Enterotoxin C2 Mutant-Directed Fatty Acid and Mitochondrial Energy Metabolic Programs Regulate CD8(+) T Cell Activation. J. Immunol. 2020, 205, 2066–2076. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, H.; Xu, M.; Liu, C.; Zhang, C. Biological analysis of the deletion mutants of Staphylococcal enterotoxin C2. Appl. Microbiol. Biotechnol. 2009, 83, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Solal-Celigny, P.; Simeon, J.; Herrera, A.; Boivin, P. Cancer treatment with Staphylococcus aureus protein A. Biomed. Pharmacother. Biomed. Pharmacother. 1985, 39, 177–186. [Google Scholar]
- Rigi, G.; Ghaedmohammadi, S.; Ahmadian, G. A comprehensive review on staphylococcal protein A (SpA): Its production and applications. Biotechnol. Appl. Biochem. 2019, 66, 454–464. [Google Scholar] [CrossRef]
- Chen, X.; Schneewind, O.; Missiakas, D. Engineered human antibodies for the opsonization and killing of Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2022, 119, e2114478119. [Google Scholar] [CrossRef]
- Kozmin, S.G.; Rogozin, I.B.; Moore, E.A.; Abney, M.; Schaaper, R.M.; Pavlov, Y.I. Comment on “A commensal strain of Staphylococcus epidermidis protects against skin neoplasia” by Nakatsuji et al. Sci. Adv. 2019, 5, eaaw3915. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).