
Genomic Investigation of Desert Streptomyces huasconensis D23 Reveals Its Environmental Adaptability and Antimicrobial Activity
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Bacterial Strain Isolation
2.2. Screening of the Antibacterial Activity
2.3. Screening UV-C Radiation-Resistance and H2O2 Tolerance of the Strain
2.4. Whole-Genome Sequencing and Annotation Analysis
3. Results
3.1. Taxonomic Studies of Streptomyces sp. D23
3.2. Screening for the Potential Activity
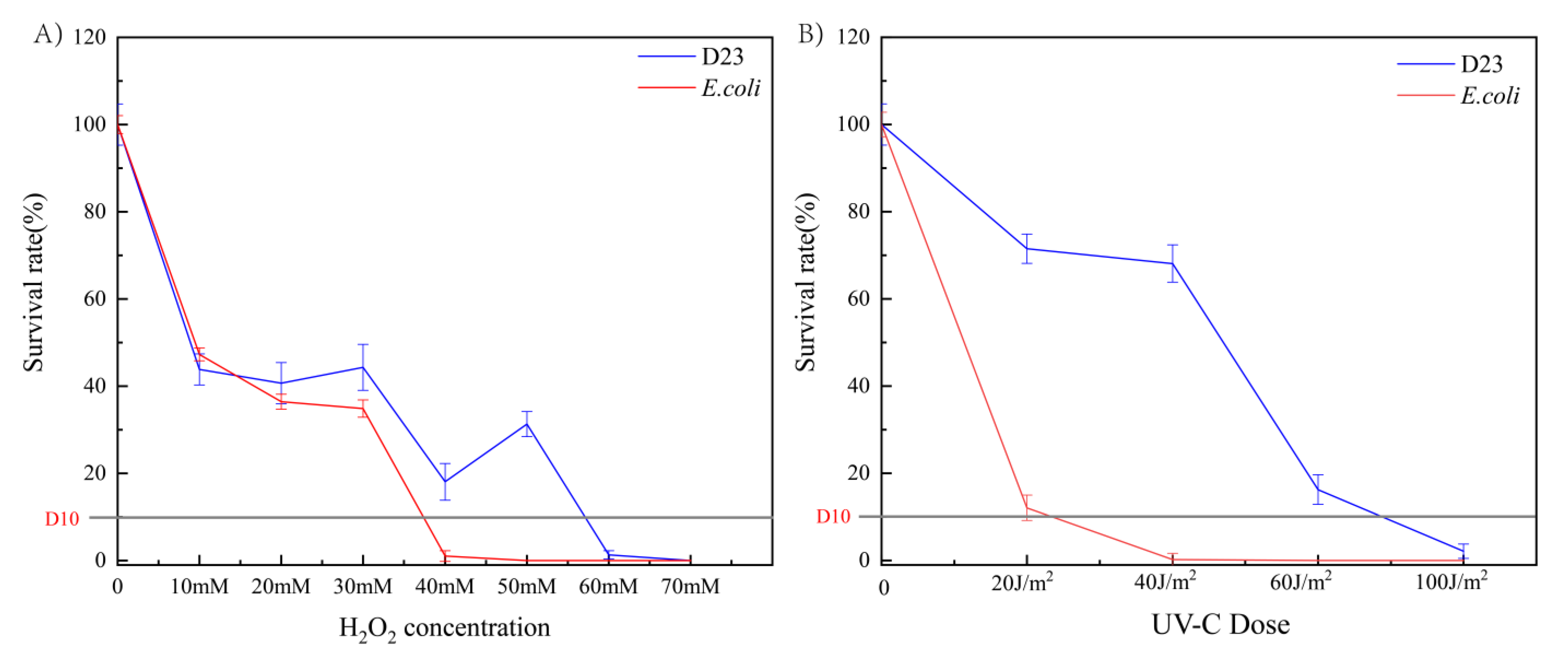
3.3. Survival Rates after Exposure to UV-C and H2O2 Tolerance
3.4. General Genome Features of S. huasconensis D23
3.5. Genetic Basis for Secondary Metabolites
3.6. Analysis of the Predicted Proteins
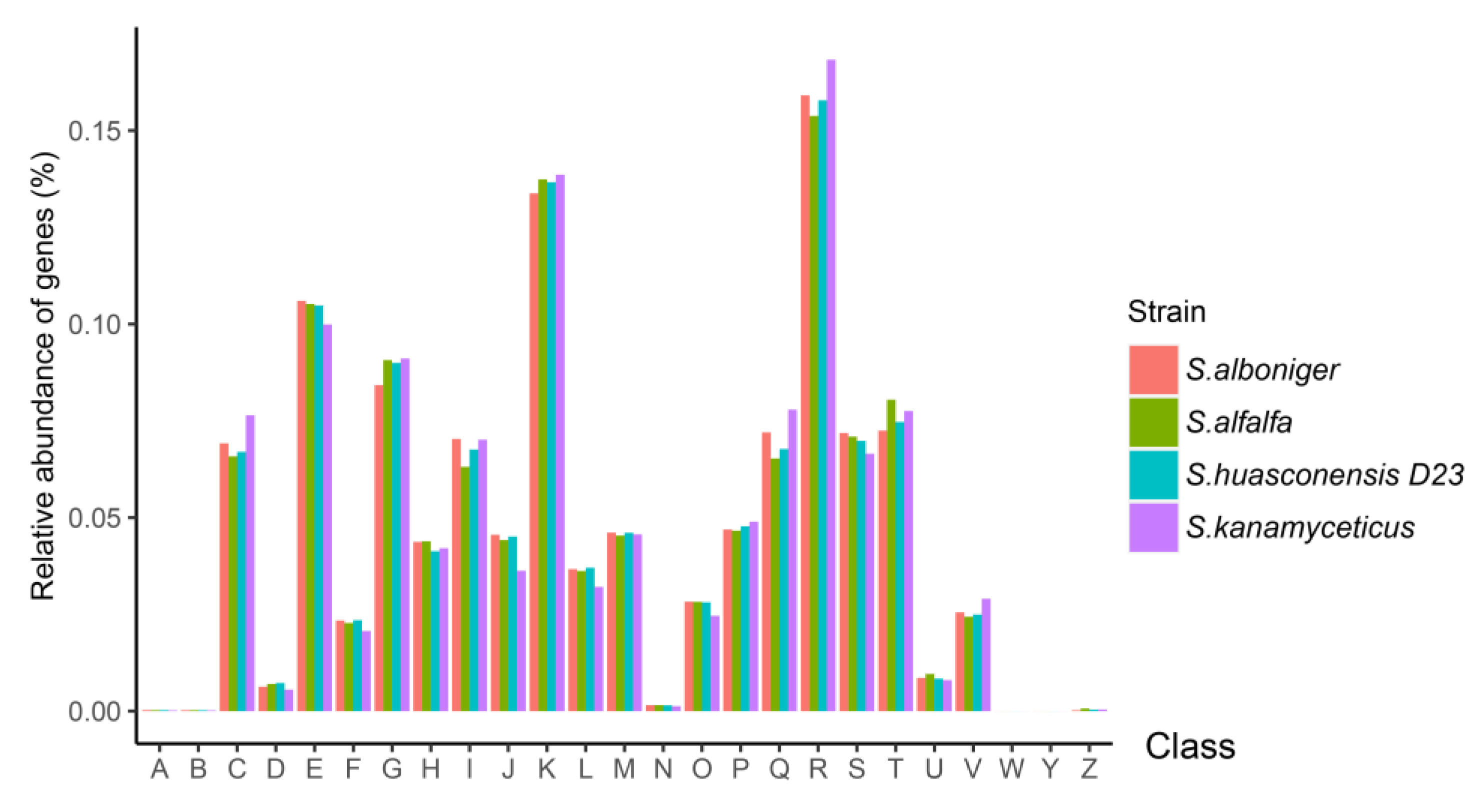
3.7. Cluster of Orthologous Groups (COG) Annotation
3.8. Adaptation to Environment and Stress Responses
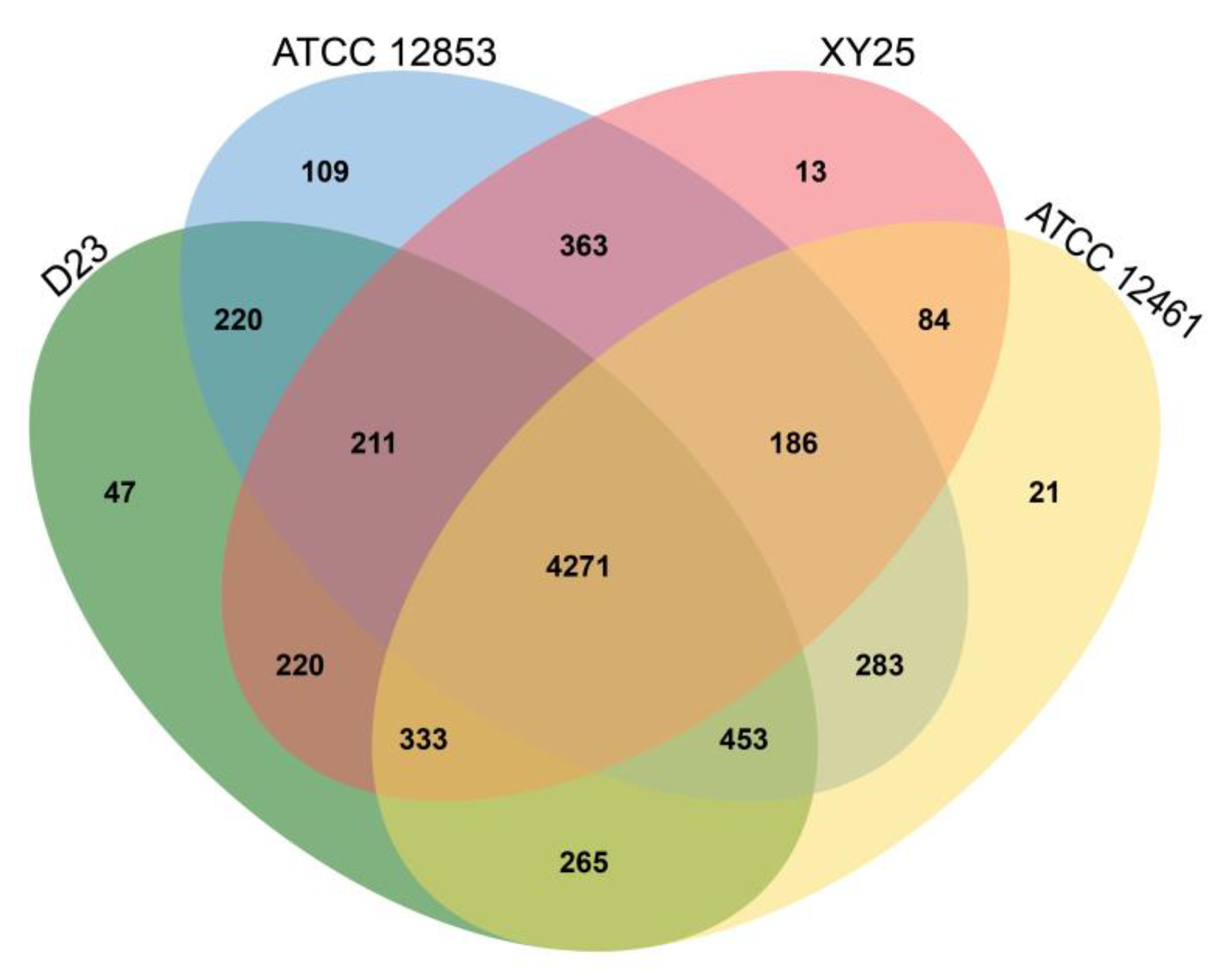
3.9. Core genome and Pan-Genome of S. huasconensis D23
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singer, A.C.; Shaw, H.; Rhodes, V.; Hart, A. Review of Antimicrobial Resistance in the Environment and Its Relevance to Environmental Regulators. Front. Microbiol. 2016, 7, 1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naughton, L.M.; Romano, S.; O’Gara, F.; Dobson, A.D.W. Identification of Secondary Metabolite Gene Clusters in the Pseudovibrio Genus Reveals Encouraging Biosynthetic Potential toward the Production of Novel Bioactive Compounds. Front. Microbiol. 2017, 8, 1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gyawali, R.; Ibrahim, S.A. Natural products as antimicrobial agents. Food Control. 2014, 46, 412–429. [Google Scholar] [CrossRef]
- Cragg, G.M.; Boyd, M.R.; Khanna, R.; Newman, D.J.; Sausville, E.A. Natural Product Drug Discovery and Development. J. Nat. Prod. 1997, 60, 52–60. [Google Scholar] [CrossRef]
- Ye, S.; Molloy, B.; Braña, A.F.; Zabala, D.; Olano, C.; Cortés, J.; Morís, F.; Salas, J.A.; Méndez, C. Identification by Genome Mining of a Type I Polyketide Gene Cluster from Streptomyces argillaceus Involved in the Biosynthesis of Pyridine and Piperidine Alkaloids Argimycins P. Front. Microbiol. 2017, 8, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Deng, Z.; Liu, T. Streptomyces species: Ideal chassis for natural product discovery and overproduction. Metab. Eng. 2018, 50, 74–84. [Google Scholar] [CrossRef]
- Schatz, A.; Bugle, E.; Waksman, S.A. Streptomycin, a Substance Exhibiting Antibiotic Activity Against Gram-Positive and Gram-Negative Bacteria. Exp. Biol. Med. 1944, 55, 66–69. [Google Scholar] [CrossRef]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bérdy, J. Bioactive Microbial Metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Park, B.S.; Bhatia, S.K.; Seo, H.-M.; Jeon, J.-M.; Kim, H.-J.; Yi, D.-H.; Lee, J.-H.; Choi, K.-Y.; Park, H.-Y.; et al. Production of Rapamycin in Streptomyces hygroscopicus from Glycerol-Based Media Optimized by Systemic Methodology. J. Microbiol. Biotechnol. 2014, 24, 1319–1326. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Giovanardi, D.; Stefani, E. Plant Growth Promoting and Biocontrol Activity of Streptomyces spp. as Endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, K.; Gupta, R.K. Rare actinomycetes: A potential storehouse for novel antibiotics. Crit. Rev. Biotechnol. 2011, 32, 108–132. [Google Scholar] [CrossRef] [PubMed]
- Neilson, J.W.; Califf, K.; Cardona, C.; Copeland, A.; van Treuren, W.; Josephson, K.L.; Knight, R.; Gilbert, J.A.; Quade, J.; Caporaso, J.G.; et al. Significant Impacts of Increasing Aridity on the Arid Soil Microbiome. mSystems 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Bahadur, A.; Li, T.; Sajjad, W.; Nasir, F.; Zia, M.A.; Wu, M.; Zhang, G.; Liu, G.; Chen, T.; Zhang, W. Transcriptional and biochemical analyses of Planomicrobium strain AX6 from Qinghai-Tibetan Plateau, China, reveal hydrogen peroxide scavenging potential. BMC Microbiol. 2022, 22, 265. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Neilson, J.W.; Kushwaha, P.; Maier, R.M.; Barberán, A. Life-history strategies of soil microbial communities in an arid ecosystem. ISME J. 2021, 15, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Sivakala, K.; Gutiérrez-García, K.; Jose, P.A.; Thinesh, T.; Anandham, R.; Barona-Gómez, F.; Sivakumar, N. Desert Environments Facilitate Unique Evolution of Biosynthetic Potential in Streptomyces. Molecules 2021, 26, 588. [Google Scholar] [CrossRef]
- Cortes-Abayay, C.; Dorador, C.; Schumann, P.; Andrews, B.; Asenjo, J.; Nouioui, I. Streptomyces huasconensis sp. nov., an haloalkalitolerant actinobacterium isolated from a high-altitude saline wetland at the Chilean Altiplano. Int. J. Syst. Evol. Microbiol. 2019, 69, 2315–2322. [Google Scholar] [CrossRef]
- Cortés-Albayay, C.; Jarmusch, S.A.; Trusch, F.; Ebel, R.; Andrews, B.A.; Jaspars, M.; Asenjo, J.A. Downsizing Class II Lasso Peptides: Genome Mining-Guided Isolation of Huascopeptin Containing the First Gly1-Asp7 Macrocycle. J. Org. Chem. 2020, 85, 1661–1667. [Google Scholar] [CrossRef]
- Reasoner, D.J.; Geldreich, E.E. A new medium for the enumeration and subculture of bacteria from potable water. Appl. Environ. Microbiol. 1985, 49, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kim, O.-S.; Cho, Y.-J.; Lee, K.; Yoon, S.-H.; Kim, M.; Na, H.; Park, S.-C.; Jeon, Y.S.; Lee, J.-H.; Yi, H.; et al. Introducing EzTaxon-e: A prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species. Int. J. Syst. Evol. Microbiol. 2012, 62, 716–721. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, T.; Li, J.; Wu, M.; Liu, G.; Zhang, W.; Zhang, B.; Zhang, S.; Zhang, G. High Proportions of Radiation-Resistant Strains in Culturable Bacteria from the Taklimakan Desert. Biology 2022, 11, 501. [Google Scholar] [CrossRef] [PubMed]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Haft, D.H.; DiCuccio, M.; Badretdin, A.; Brover, V.; Chetvernin, V.; O’Neill, K.; Li, W.; Chitsaz, F.; Derbyshire, M.K.; Gonzales, N.R.; et al. RefSeq: An update on prokaryotic genome annotation and curation. Nucleic Acids Res. 2017, 46, D851–D860. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; O’Neill, K.R.; Haft, D.H.; DiCuccio, M.; Chetvernin, V.; Badretdin, A.; Coulouris, G.; Chitsaz, F.; Derbyshire, M.K.; Durkin, A.S.; et al. RefSeq: Expanding the Prokaryotic Genome Annotation Pipeline reach with protein family model curation. Nucleic Acids Res. 2020, 49, D1020–D1028. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhu, Z.; Fu, L.; Niu, B.; Li, W. WebMGA: A customizable web server for fast metagenomic sequence analysis. BMC Genom. 2011, 12, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Dong, Z.; Fang, L.; Luo, Y.; Wei, Z.; Guo, H.; Zhang, G.; Gu, Y.Q.; Coleman-Derr, D.; Xia, Q.; et al. OrthoVenn2: A web server for whole-genome comparison and annotation of orthologous clusters across multiple species. Nucleic Acids Res. 2019, 47, W52–W58. [Google Scholar] [CrossRef] [Green Version]
- Chaudhari, N.M.; Gupta, V.; Dutta, C. BPGA- an ultra-fast pan-genome analysis pipeline. Sci. Rep. 2016, 6, 24373. [Google Scholar] [CrossRef] [Green Version]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A Comprehensive, Accurate, and Fast Distance-Based Phylogeny Inference Program. Mol. Biol. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef]
- Juttner, F.; Watson, S.B. Biochemical and Ecological Control of Geosmin and 2-Methylisoborneol in Source Waters. Appl. Environ. Microbiol. 2007, 73, 4395–4406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinert, D.J.; Balliano, G.; Schulz, G.E. Conversion of Squalene to the Pentacarbocyclic Hopene. Chem. Biol. 2004, 11, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Schulz, A.; Hermann, L.; Freibert, S.-A.; Bönig, T.; Hoffmann, T.; Riclea, R.; Dickschat, J.S.; Heider, J.; Bremer, E. Transcriptional regulation of ectoine catabolism in response to multiple metabolic and environmental cues. Environ. Microbiol. 2017, 19, 4599–4619. [Google Scholar] [CrossRef] [PubMed]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2013, 42, D206–D214. [Google Scholar] [CrossRef] [PubMed]
- Demain, A.L. Importance of microbial natural products and the need to revitalize their discovery. J. Ind. Microbiol. Biotechnol. 2014, 41, 185–201. [Google Scholar] [CrossRef]
- Zhou, Z.; Gu, J.; Du, Y.-L.; Li, Y.-Q.; Wang, Y. The -omics Era- Toward a Systems-Level Understanding of Streptomyces. Curr. Genom. 2011, 12, 404–416. [Google Scholar] [CrossRef] [Green Version]
- Wael, N.H.; Mohammed, I.A.A.; Ola, H.; Ahmed, S.M.; Michael, G.; Hozzein, W.N.; Ali, M.I.A.; Hammouda, O.; Mousa, A.S.; Goodfellow, M. Streptomyces sannurensis sp. nov., a new alkaliphilic member of the genus Streptomyces isolated from Wadi Sannur in Egypt. Afr. J. Microbiol. Res. 2011, 5, 1329–1334. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Li, Y.-Q.; Asem, M.D.; Lu, C.-Y.; Shi, X.-H.; Chu, X.; Zhang, W.-Q.; Di An, D.; Li, W.-J. Streptomyces xinjiangensis sp. nov., an actinomycete isolated from Lop Nur region. Arch. Microbiol. 2016, 198, 785–791. [Google Scholar] [CrossRef]
- Cortés-Albayay, C.; Dorador, C.; Schumann, P.; Schniete, J.K.; Herron, P.; Andrews, B.; Asenjo, J.; Nouioui, I. Streptomyces altiplanensis sp. nov., an alkalitolerant species isolated from Chilean Altiplano soil, and emended description of Streptomyces chryseus (Krasil’nikov et al. 1965) Pridham 1970. Int. J. Syst. Evol. Microbiol. 2019, 69, 2498–2505. [Google Scholar] [CrossRef]
- Cortés-Albayay, C.; Silber, J.; Imhoff, J.F.; Asenjo, J.A.; Andrews, B.; Nouioui, I.; Dorador, C. The Polyextreme Ecosystem, Salar de Huasco at the Chilean Altiplano of the Atacama Desert Houses Diverse Streptomyces spp. with Promising Pharmaceutical Potentials. Diversity 2019, 11, 69. [Google Scholar] [CrossRef]
- Gürtler, H.; Pedersen, R.; Anthoni, U.; Christophersen, C.; Nielsen, P.H.; Wellington, E.M.H.; Pedersen, C.; Bock, K. Albaflavenone, a sesquiterpene ketone with a zizaene skeleton produced by a streptomycete with a new rope morphology. J. Antibiot. 1994, 47, 434–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moody, S.C.; Zhao, B.; Lei, L.; Nelson, D.R.; Mullins, J.G.L.; Waterman, M.R.; Kelly, S.L.; Lamb, D.C. Investigating conservation of the albaflavenone biosynthetic pathway and CYP170 bifunctionality in streptomycetes. FEBS J. 2012, 279, 1640–1649. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-F.; Xun, H.; Gao, Q.; Tang, F.; Qi, F.-F.; Sun, J. Two new sesquiterpenoids from Dictyophora indusiata. J. Asian Nat. Prod. Res. 2022, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhou, X.; Liu, J.; Bao, K.; Zhang, G.; Tu, G.; Kieser, T.; Deng, Z. ‘Streptomyces nanchangensis’, a producer of the insecticidal polyether antibiotic nanchangmycin and the antiparasitic macrolide meilingmycin, contains multiple polyketide gene clusters. Microbiology 2002, 148, 361–371. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhou, X.; Dong, H.; Tu, G.; Wang, M.; Wang, B.; Deng, Z. A Complete Gene Cluster from Streptomyces nanchangensis NS3226 Encoding Biosynthesis of the Polyether Ionophore Nanchangmycin. Chem. Biol. 2003, 10, 431–441. [Google Scholar] [CrossRef] [Green Version]
- Shao, M.; Ma, J.; Li, Q.; Ju, J. Identification of the Anti-Infective Aborycin Biosynthetic Gene Cluster from Deep-Sea-Derived Streptomyces sp. SCSIO ZS0098 Enables Production in a Heterologous Host. Mar. Drugs 2019, 17, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, L.; Huang, H.; Li, H.; Wang, S.; Ju, J.; Li, W. Overexpression of a type III PKS gene affording novel violapyrones with enhanced anti-influenza A virus activity. Microb. Cell Factories 2018, 17, 61. [Google Scholar] [CrossRef] [Green Version]
- Kirst, H.A.; Dorman, D.E.; Occolowitz, J.L.; Jones, N.D.; Paschal, J.W.; Hamill, R.L.; Szymanski, E.F. The structure of A201A, a novel nucleoside antibiotic. J. Antibiot. 1985, 38, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Barrasa, M.I.; Tercero, J.A.; Lacalle, R.A.; Jimenez, A. The Ard1 Gene from Streptomyces capreolus Encodes a Polypeptide of the ABC-Transporters Superfamily Which Confers Resistance to the Aminonucleoside Antibiotic A201A. Eur. J. Biochem. 1995, 228, 562–569. [Google Scholar] [CrossRef]
- Barrasa, M.I.; Tercero, J.A.; Jimenez, A. The aminonucleoside antibiotic A201A is inactivated by a phosphotransferase activity from Streptomyces capreolus NRRL 3817, the producing organism-Isolation and molecular characterization of the relevant encoding gene and its DNA flanking regions. Eur. J. Biochem. 1997, 245, 54–63. [Google Scholar] [CrossRef]
- Sanglier, J.-J.; Quesniaux, V.; Fehr, T.; Hofmann, H.; Mahnke, M.; Memmert, K.; Schuler, W.; Zenke, G.; Gschwind, L.; Maurer, C.; et al. Sanglifehrins A, B, C and D, Novel Cyclophilin-binding Compounds Isolated from Streptomyces sp. A92-308110. I. Taxonomy, Fermentation, Isolation and Biological Activity. J. Antibiot. 1999, 52, 466–473. [Google Scholar] [CrossRef] [Green Version]
- Sokolskaja, E.; Sayah, D.M.; Luban, J. Target Cell Cyclophilin A Modulates Human Immunodeficiency Virus Type 1 Infectivity. J. Virol. 2004, 78, 12800–12808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umezawa, H.; Ueda, M.; Maeda, K.; Yagishita, K.; Kondo, S.; Okami, Y.; Utahara, R.; Osato, Y.; Nitta, K.; Takeuchi, T. Production and isolation of a new antibiotic: Kanamycin. J. Antibiot. 1957, 10, 181–188. [Google Scholar]
- Yanai, K.; Murakami, T. The kanamycin biosynthetic gene cluster from Streptomyces kanamyceticus. J. Antibiot. 2004, 57, 351–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chater, K.F.; Chandra, G. The evolution of development in Streptomyces analysed by genome comparisons. FEMS Microbiol. Rev. 2006, 30, 651–672. [Google Scholar] [CrossRef] [Green Version]
- Ensign, J.C. Formation, Properties, and Germination of Actinomycete Spores. Annu. Rev. Microbiol. 1978, 32, 185–219. [Google Scholar] [CrossRef]
- Gao, Q.; Garcia-Pichel, F. Microbial ultraviolet sunscreens. Nat. Rev. Genet. 2011, 9, 791–802. [Google Scholar] [CrossRef]
- McCarthy, A.J.; Williams, S.T. Actinomycetes as agents of biodegradation in the environment—A review. Gene 1992, 115, 189–192. [Google Scholar] [CrossRef]
- Bahadur, A.; Zhang, Z.; Sajjad, W.; Nasir, F.; Zia, M.A.; Liu, G.; Chen, T.; Zhang, W. Bacterial community structure and functions in microhabitats associated with black stones in Black Gobi desert, China. Ecol. Indic. 2022, 142, 109168. [Google Scholar] [CrossRef]
- Guesmi, S.; Nouioui, I.; Pujic, P.; Dubost, A.; Najjari, A.; Ghedira, K.; Igual, J.M.; Cherif, A.; Klenk, H.-P.; Sghaier, H.; et al. Draft genome sequence of Promicromonospora panici sp. nov., a novel ionizing-radiation-resistant actinobacterium isolated from roots of the desert plant Panicum turgidum. Extremophiles 2020, 25, 25–38. [Google Scholar] [CrossRef]
- Mao, J.; Tang, Q.; Zhang, Z.; Wang, W.; Wei, D.; Huang, Y.; Liu, Z.; Shi, Y.; Goodfellow, M. Streptomyces radiopugnans sp. nov., a radiation-resistant actinomycete isolated from radiation-polluted soil in China. Int. J. Syst. Evol. Microbiol. 2007, 57, 2578–2582. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.M.; Bay, S.K.; Meier, D.V.; Chiri, E.; Cowan, D.A.; Gillor, O.; Woebken, D.; Greening, C. Energetic Basis of Microbial Growth and Persistence in Desert Ecosystems. mSystems 2020, 5, e00495-19. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Wang, Q.; Yang, M.; Chen, Z.; Li, J.; Wen, Y. Heat Shock Repressor HspR Directly Controls Avermectin Production, Morphological Development, and H 2 O 2 Stress Response in Streptomyces avermitilis. Appl. Environ. Microbiol. 2021, 87, AEM0047321. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Wu, C.; Zhu, L.; Chang, R.; Ma, W.; Deng, Y.; Chen, X. Bioactivity profiling of the extremolyte ectoine as a promising protectant and its heterologous production. 3 Biotech 2022, 12, 331. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Schröter, M.-A.; Hahn, M.B.; Solomun, T.; Sturm, H.; Kunte, H.J. Ectoine can enhance structural changes in DNA in vitro. Sci. Rep. 2017, 7, 7170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Liu, H.; Shi, M.; Jiang, M.; Li, L.; Zheng, Y. Microbial production of ectoine and hydroxyectoine as high-value chemicals. Microb. Cell Factories 2021, 20, 76. [Google Scholar] [CrossRef] [PubMed]
- Schrader, K.K.; Blevins, W.T. Effects of selected environmental conditions on biomass and geosmin production by Streptomyces halstedii. J. Microbiol. 1999, 37, 159–167. [Google Scholar]
- Sáenz, J.P.; Grosser, D.; Bradley, A.S.; Lagny, T.J.; Lavrynenko, O.; Broda, M.; Simons, K. Hopanoids as functional analogues of cholesterol in bacterial membranes. Proc. Natl. Acad. Sci. USA 2015, 112, 11971–11976. [Google Scholar] [CrossRef] [Green Version]
- Rizk, S.; Henke, P.; Santana-Molina, C.; Martens, G.; Gnädig, M.; Nguyen, N.A.; Devos, D.P.; Neumann-Schaal, M.; Saenz, J.P. Functional diversity of isoprenoid lipids in Methylobacterium extorquens PA1. Mol. Microbiol. 2021, 116, 1064–1078. [Google Scholar] [CrossRef]
- Goverde, R.L.J.; Huis in’t Veld, J.H.J.; Kusters, J.G.; Mooi, F.R. The psychrotrophic bacterium Yersinia enterocolitica requires expression of pnp, the gene for polynucleotide phosphorylase, for growth at low temperature (5 °C). Mol. Microbiol. 1998, 28, 555–569. [Google Scholar] [CrossRef]
- Hecker, M.; Völker, U. General stress response of Bacillus subtilis and other bacteria. Adv. Microb. Physiol. 2001, 44, 35–91. [Google Scholar] [CrossRef]
- Holtmann, G.; Brigulla, M.; Steil, L.; Schütz, A.; Barnekow, K.; Völker, U.; Bremer, E.; Bremer, E. RsbV- Independent Induction of the SigB-Dependent General Stress Regulon of Bacillus subtilis during Growth at High Temperature. J. Bacteriol. 2004, 186, 6150–6158. [Google Scholar] [CrossRef] [PubMed]
- Dorey, A.; Marinho, C.; Piveteau, P.; O’Byrne, C. Role and regulation of the stress activated sigma factor sigma B (σB) in the saprophytic and host-associated life stages of Listeria monocytogenes. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2019; Volume 106, pp. 1–48. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B.c | B.th | S.a | E.c | P.p | L.l | M.l | S.ep | |
|---|---|---|---|---|---|---|---|---|
| CASM | 30 ± 3 | 15 ± 9 | 25 ± 1 | 14 ± 2 | 10 ± 9 | - | - | - |
| ISP2 | 9 ± 4 | - | - | - | - | 9 ± 1 | 12 ± 4 | - |
| R2A | 22 ± 4 | - | - | - | - | - | - | 19 ± 1 |
| Characteristics | D23 | S. alfalfae XY25 | S. alboniger ATCC12461 | S. kanamyceticus ATCC12853 |
|---|---|---|---|---|
| Size (bp) | 8,236,582 | 8,273,342 | 7,962,786 | 10,133,897 |
| GC Content (%) | 71.6 | 72.2 | 71.2 | 71.0 |
| Number of Contigs (with PEGs) | 1 | 21 | 1 | 1 |
| Number of Coding Sequences | 7131 | 7175 | 6840 | 8762 |
| Number of tRNA | 94 | 85 | 81 | 80 |
| Number of rRNA | 18 | 17 | 18 | 18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, Y.; Zhang, G.; Bahadur, A.; Xu, Y.; Liu, Y.; Tian, M.; Ding, W.; Chen, T.; Zhang, W.; Liu, G. Genomic Investigation of Desert Streptomyces huasconensis D23 Reveals Its Environmental Adaptability and Antimicrobial Activity. Microorganisms 2022, 10, 2408. https://doi.org/10.3390/microorganisms10122408
Wen Y, Zhang G, Bahadur A, Xu Y, Liu Y, Tian M, Ding W, Chen T, Zhang W, Liu G. Genomic Investigation of Desert Streptomyces huasconensis D23 Reveals Its Environmental Adaptability and Antimicrobial Activity. Microorganisms. 2022; 10(12):2408. https://doi.org/10.3390/microorganisms10122408
Chicago/Turabian StyleWen, Ying, Gaosen Zhang, Ali Bahadur, Yeteng Xu, Yang Liu, Mao Tian, Wei Ding, Tuo Chen, Wei Zhang, and Guangxiu Liu. 2022. "Genomic Investigation of Desert Streptomyces huasconensis D23 Reveals Its Environmental Adaptability and Antimicrobial Activity" Microorganisms 10, no. 12: 2408. https://doi.org/10.3390/microorganisms10122408
APA StyleWen, Y., Zhang, G., Bahadur, A., Xu, Y., Liu, Y., Tian, M., Ding, W., Chen, T., Zhang, W., & Liu, G. (2022). Genomic Investigation of Desert Streptomyces huasconensis D23 Reveals Its Environmental Adaptability and Antimicrobial Activity. Microorganisms, 10(12), 2408. https://doi.org/10.3390/microorganisms10122408