
Evaluating Methods of Preserving Aquatic Invertebrates for Microbiome Analysis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Collection and Processing
2.2. DNA Extraction, Amplification, and Sequencing
2.3. Statistical Analyses
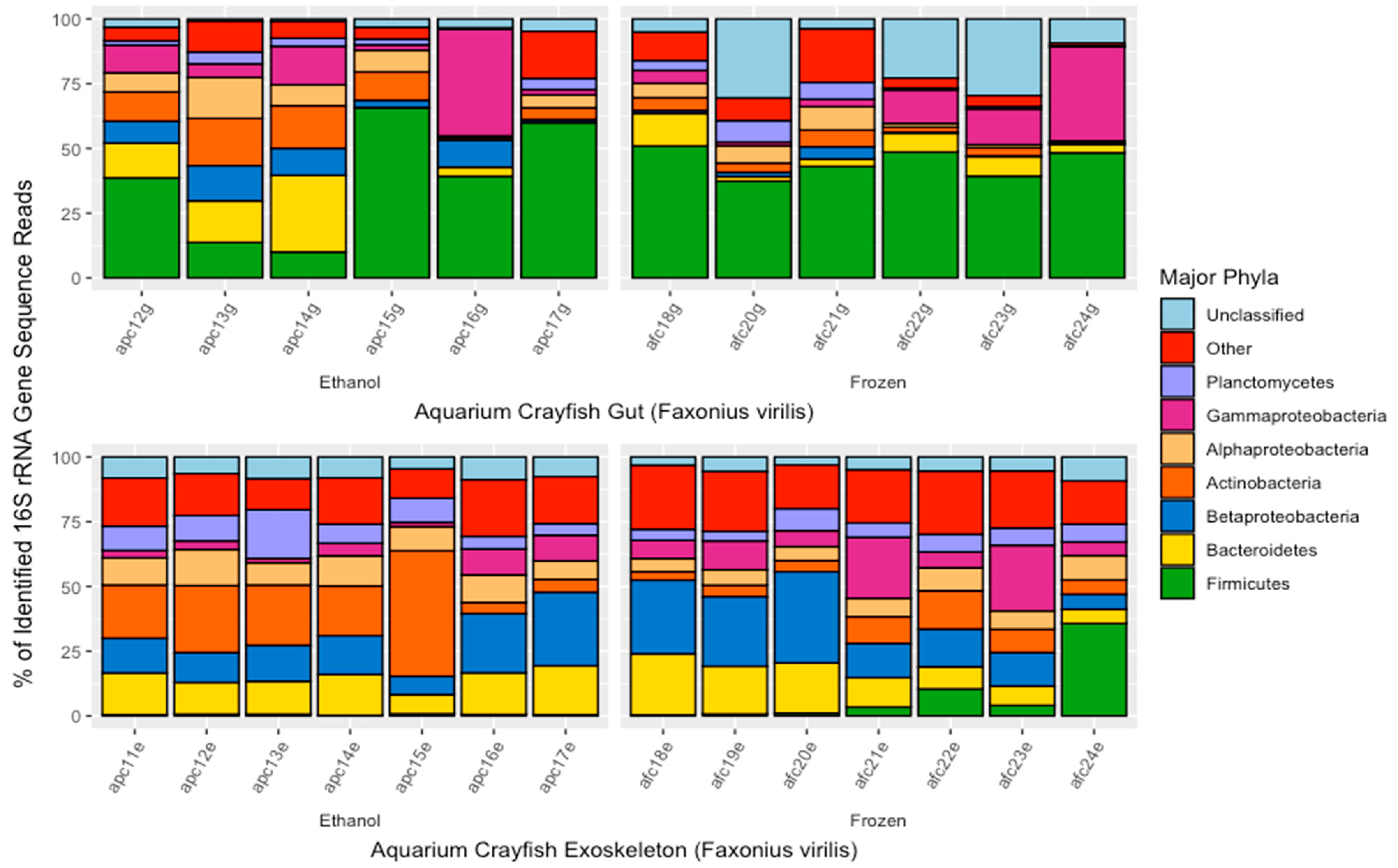
3. Results
3.1. Sequence Counts
3.2. Differences in the Crayfish Microbiome between Sample Types and Preservation Method
3.3. Dominant ASVs by Sample Type and Preservation Method
3.4. Patterns in the Dragonfly Nymph Microbiome by Species and Preservation Method
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cani, P.D. Human Gut Microbiome: Hopes, threats and promises. Gut 2018, 67, 1716–1725. [Google Scholar] [CrossRef] [PubMed]
- Clooney, A.G.; Fouhy, F.; Sleator, R.D.; O’Driscroll, A.; Stanton, C. Comparing Apples and Oranges?: Next Generation Sequencing and Its Impact on Microbiome Analysis. PLoS ONE 2016, 11, e0148028. [Google Scholar] [CrossRef]
- Colston, T.J.; Jackson, C.R. Microbiome evolution along divergent branches of the vertebrate tree of life: What is known and unknown. Mol. Ecol. 2016, 25, 3776–3800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Sheng, L.; Hong, Y.; Xi, C.; Gu, Y.; Zheng, N.; Li, M.; Chen, L.; Wu, G.; Li, Y.; et al. Variations of Gut Microbiome Profile Under Different Storage Conditions and Preservation Periods: A Multi-Dimensional Evaluation. Front. Microbiol. 2020, 11, 972. [Google Scholar] [CrossRef] [PubMed]
- Greay, T.L.; Gofton, A.W.; Paparini, A.; Ryan, U.M.; Oskam, C.L.; Irwin, P.J. Recent insights into the tick microbiome gained through next-generation sequencing. Parasites Vectors 2018, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Ghanbari, M.; Kneifel, W.; Domig, K.J. A new view of the fish gut microbiome: Advances from next-generation sequencing. Aquaculture 2015, 448, 464–475. [Google Scholar] [CrossRef]
- Foster, J.A.; Bunge, J.; Gilbert, J.A.; Moore, J.H. Measuring the microbiome: Perspectives on advances in DNA-based techniques for exploring microbial life. Brief. Bioinform. 2012, 13, 420–429. [Google Scholar] [CrossRef] [Green Version]
- Anslan, S.; Li, H.; Kunzel, S.; Vences, M. Microbiomes from feces vs. gut in tadpoles: Distinct community compositions between substrates and preservation methods. Salamandra 2021, 57, 96–104. [Google Scholar]
- Majumder, R.; Sutcliffe, B.; Taylor, P.W.; Chapman, T.A. Next-Generation Sequencing reveals relationship between the larval microbiome and food substrate in the polyphagous Queensland fruit fly. Sci. Rep. 2019, 9, 14292. [Google Scholar] [CrossRef] [Green Version]
- Song, S.J.; Amir, A.; Metcalf, J.L.; Amato, K.R.; Xu, Z.Z.; Humphrey, G.; Knight, R. Preservation methods differ in fecal microbiome stability, affecting suitability for field studies. mSystems 2016, 1, e00021-16. [Google Scholar] [CrossRef] [Green Version]
- Horng, K.R.; Ganz, H.H.; Eisen, J.A.; Marks, S.L. Effects of preservation method on canine (Canis lupus familiaris) fecal microbiota. PeerJ 2018, 6, e4827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Cock, M.; Virgilio, M.; Vandamme, P.; Augustinos, A.; Bourtzis, K.; Willems, A.; De Meyer, M. Impact of Sample Preservation and Manipulation on Insect Gut Microbiome Profiling. A Test Case with Fruit Flies (Diptera, Tephritidae). Front. Microbiol. 2019, 10, 2833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, T.J.; Dickerson, J.C.; Fierer, N. Evidence-based recommendations on storing and handling specimens for analyses of insect microbiota. PeerJ 2015, 3, e1190. [Google Scholar] [CrossRef] [Green Version]
- Simister, R.; Schmitt, S.; Taylor, M.W. Evaluating methods for the preservation and extraction of DNA and RNA for analysis of microbial communities in marine sponges. J. Exp. Mar. Biol. Ecol. 2011, 397, 38–43. [Google Scholar] [CrossRef]
- Rocha, J.; Coelho, F.; Peixe, L.; Gomes, N.C.M.; Calado, R. Optimization of preservation and processing of sea anemones for microbial community analysis using molecular tools. Sci. Rep. 2014, 4, 6986. [Google Scholar] [CrossRef] [PubMed]
- Blekhman, R.; Tang, K.; Archie, E.A.; Barreiro, L.B.; Johnson, Z.P.; Wilson, M.E.; Kohn, J.; Yuan, M.L.; Gesquiere, L.; Grieneisen, L.E.; et al. Common methods for fecal sample storage in field studies yield consistent signatures of individual identity in microbiome sequencing data. Sci. Rep. 2016, 6, 31519. [Google Scholar] [CrossRef] [Green Version]
- Lauber, C.L.; Zhou, N.; Gordon, J.I.; Knight, R.; Fierer, N. Effect of storage conditions on the assessment of bacterial community structure in soil and human-associated samples. FEMS Microbiol. Lett. 2010, 307, 80–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughn, C.C. Ecosystem services provided by freshwater mussels. Hydrobiologia 2018, 810, 15–27. [Google Scholar] [CrossRef]
- Weingarten, E.A.; Atkinson, C.L.; Jackson, C.R. The gut microbiome of freshwater Unionidae mussels is determined by host species and is selectively retained from filtered seston. PLoS ONE 2019, 14, e0224796. [Google Scholar] [CrossRef]
- Prather, C.M.; Pelini, S.L.; Laws, A.; Rivest, E.; Woltz, M.; Bloch, C.P.; Del Toro, I.; Ho, C.-K.; Kominoski, J.; Newbold, T.A.S.; et al. Invertebrates, ecosystem services and climate change. Biol. Rev. 2013, 88, 327–348. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, C.R.; Stone, B.W.G.; Tyler, H.L. Emerging perspectives on the natural microbiome of fresh produce vegetables. Agriculture 2015, 5, 170–187. [Google Scholar] [CrossRef] [Green Version]
- Stone, B.W.G.; Jackson, C.R. Biogeographic patterns between bacterial phyllosphere communities of the Southern Magnolia (Magnolia grandiflora) in a small forest. Microb. Ecol. 2016, 71, 954–961. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- RStudio Team. RStudio: Integrated Development for R; RStudio PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 7 July 2020).
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. vegan: Community Ecology Package. Version 2.5–7 2008, 10, 631–637. [Google Scholar]
- Lahti, L.; Sudarshan, S.; Blake, T.; Salojarvi, J. Tools for microbiome analysis in R. Version 2017, 1, 28. [Google Scholar]
- Hale, V.L.; Tan, C.L.; Knight, R.; Amato, K.R. Effect of preservation method on spider monkey (Ateles geoffroyi) fecal microbiota over 8 weeks. J. Microbiol. Methods 2015, 113, 16–26. [Google Scholar] [CrossRef]
- Krogmann, L.; Holstein, J. Preserving and Specimen Handling: Insects and other Invertebrates. In Manual on Field Recording Techniques and Protocols for All Taxa Biodiversity Inventories; Eymann, J., Degreef, J., Hauser, C., Monje, J.C., Samyn, Y., VandenSpiegel, D., Eds.; ABC Taxa: Brussels, Belgium, 2010; Volume 1, pp. 463–479. [Google Scholar]
- Schiller, E.K.; Haring, E.; Daubl, B.; Gaub, L.; Szeiler, S.; Sattmann, H. Ethanol concentration and sample preservation considering diverse storage parameters: A survey of invertebrate wet collections of the Natural History Museum Vienna. Ann. Naturhist. Mus. Wien 2014, 116, 41–61. [Google Scholar]
- Moreau, C.S.; Wray, B.D.; Czekanski-Moir, J.E.; Rubin, B.E.R. DNA preservation: A test of commonly used preservatives for insects. Invert. Syst. 2013, 27, 81–86. [Google Scholar] [CrossRef]
- Nagy, Z.T. A hands-on overview of tissue preservation methods for molecular genetic analyses. Org. Divers. Evol. 2010, 10, 91–105. [Google Scholar] [CrossRef]
- Xavier, R.; Soares, M.C.; Silva, S.M.; Banha, F.; Gama, M.; Ribeiro, L.; Anastacio, P.; Cardoso, S.C. Environment and host-related factors modulate gut and carapace bacterial diversity of the invasive red swamp crayfish (Procambarus clarkii). Hydrobiologia 2021, 848, 1045–4057. [Google Scholar] [CrossRef]
- Chen, X.; Fan, L.; Qiu, L.; Dong, X.; Wang, Q.; Hu, G.; Meng, S.; Li, D.; Chen, J. Metagenomics Analysis Reveals Compositional and Functional Differences in the Gut Microbiota of Red Swamp Crayfish, Procambarus clarkii, Grown on Two Different Culture Environments. Front. Microbiol. 2021, 12, 3070. [Google Scholar] [CrossRef] [PubMed]
- Skelton, J.; Geyer, K.M.; Lennon, J.T.; Creed, R.P.; Brown, B.L. Multi-scale ecological filters shape the crayfish microbiome. Symbiosis 2016, 72, 159–170. [Google Scholar] [CrossRef]
- Dragicevic, P.; Bielen, A.; Petric, I.; Vuk, M.; Žucko, J.; Hudina, S. Microbiome of the successful freshwater invader, the signal crayfish, and its changes along the invasion range. Microbiol. Spectr. 2021, 9, e00389-21. [Google Scholar] [CrossRef] [PubMed]
- Nobles, S.; Jackson, C.R. Effects of Life Stage, Site, and Species on the Dragonfly Gut Microbiome. Microorganisms 2020, 8, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deb, R.; Nair, A.; Agashe, D. Host dietary specialization and neutral assembly shape gut bacterial communities of wild dragonflies. PeerJ 2019, 7, e8058. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aquarium Crayfish | ASV | Identification | Frequency a | Relative Abundance b | CI (+/−) |
|---|---|---|---|---|---|
| Gut Ethanol | ASV 1 | Firmicutes (Firmicutes) | 6/6 | 24.2% | 8.39% |
| ASV 4 | Flavobacterium (Bacteroidetes) | 6/6 | 7.10% | 2.55% | |
| ASV 9 | Mycobacterium (Actinobacteria) | 6/6 | 3.67% | 1.22% | |
| ASV 34 | Gemmobacter (Alpharoteobacteria) | 6/6 | 2.32% | 0.81% | |
| ASV 33 | Mycobacterium (Actinobacteria) | 6/6 | 1.77% | 0.60% | |
| ASV 69 | Dechloromonas (Betaproteobacteria) | 5/6 | 1.28% | 0.44% | |
| Gut Frozen | ASV 1 | Firmicutes (Firmicutes) | 6/6 | 21.8% | 5.01% |
| ASV 4 | Flavobacterium (Bacteroidetes) | 6/6 | 5.58% | 1.07% | |
| ASV 9 | Mycobacterium (Actinobacteria) | 6/6 | 5.23% | 0.63% | |
| ASV 27 | Hydromonas (Betaproteobacteria) | 6/6 | 3.93% | 1.48% | |
| ASV 25 | Citrobacter (Gammaproteobacteria) | 5/6 | 2.10% | 0.12% | |
| ASV 33 | Mycobacterium (Actinobacteria) | 6/6 | 1.82% | 0.25% | |
| Exoskeleton Ethanol | ASV 3 | Kineosporiaceae (Actinobacteria) | 7/7 | 15.5% | 1.93% |
| ASV 31 | Bacteroidetes (Bacteroidetes) | 7/7 | 2.14% | 0.18% | |
| ASV 19 | Phycisphaeraceae (Planctomycetes) | 6/7 | 1.98% | 0.60% | |
| ASV 21 | Pirellulaceae (Planctomycetes) | 7/7 | 1.84% | 0.13% | |
| ASV 28 | Fimbriiglobus (Planctomycetes) | 6/7 | 1.26% | 0.07% | |
| ASV 9 | Mycobacterium (Actinobacteria) | 7/7 | 1.11% | 0.10% | |
| Exoskeleton Frozen | ASV 1 | Firmicutes (Firmicutes) | 7/7 | 19.1% | 4.04% |
| ASV 4 | Flavobacterium (Bacteroidetes) | 7/7 | 4.81% | 0.89% | |
| ASV 9 | Mycobacterium (Actinobacteria) | 7/7 | 4.56% | 0.55% | |
| ASV 27 | Hydromonas (Betaproteobacteria) | 5/7 | 3.36% | 1.18% | |
| ASV 25 | Citrobacter (Gammaproteobacteria) | 6/7 | 1.81% | 0.24% | |
| ASV 33 | Mycobacterium (Actinobacteria) | 7/7 | 1.56% | 0.22% |
| Field-Collected Crayfish | ASV | Identification | Frequency a | Relative Abundance b | CI (+/−) |
|---|---|---|---|---|---|
| Gut Ethanol | ASV 7 | Catenococcus (Gammaproteobacteria) | 3/3 | 16.6% | 2.96% |
| ASV 1 | Rhodobacter (Firmicutes) | 3/3 | 12.7% | 3.22% | |
| ASV 15 | Bacilli (Firmicutes) | 3/3 | 10.3% | 1.86% | |
| ASV 11 | Clostridium_XlVb (Firmicutes) | 3/3 | 7.72% | 1.81% | |
| ASV 22 | Firmicutes (Fimicutes) | 3/3 | 5.73% | 1.65% | |
| ASV 32 | Dysgonomonas (Bacteroidetes) | 3/3 | 4.73% | 0.75% | |
| Gut Frozen | ASV 17 | Firmicutes (Firmicutes) | 5/5 | 14.5% | 3.51% |
| ASV 1 | Firmicutes (Firmicutes) | 5/5 | 12.5% | 1.53% | |
| ASV 22 | Firmicutes (Firmicutes) | 3/5 | 4.77% | 1.03% | |
| ASV 11 | Clostridium_XlVb (Firmicutes) | 5/5 | 4.53% | 0.86% | |
| ASV 32 | Dysgonomonas (Bacteroidetes) | 3/5 | 3.41% | 1.01% | |
| ASV 15 | Bacilli (Firmicutes) | 3/5 | 2.09% | 0.39% | |
| Exoskeleton Ethanol | ASV 83 | Methylococcaceae (Gammaproteobacteria) | 3/3 | 3.78% | 1.44% |
| ASV 16 | Verrucomicrobium (Verrucomicrobia) | 3/3 | 3.37% | 1.24% | |
| ASV 68 | Kineosporiaceae (Actinobacteria) | 3/3 | 3.32% | 0.83% | |
| ASV 115 | Verrucomicrobiaceae (Verrucomicrobia) | 3/3 | 2.73% | 0.54% | |
| ASV 171 | Verrucomicrobia (Verrucomicrobia) | 3/3 | 1.62% | 0.13% | |
| ASV 193 | Micrococcales (Actinobacteria) | 3/3 | 1.05% | 0.08% | |
| Exoskeleton Frozen | ASV 8 | Comamonadaceae (Proteobacteria) | 5/5 | 9.49% | 1.06% |
| ASV 16 | Verrucomicrobium (Verrucomicrobia) | 5/5 | 4.87% | 0.76% | |
| ASV 24 | Methylobacter (Gammaproteobacteria) | 5/5 | 3.84% | 0.35% | |
| ASV 12 | Sphaerotilus (Betaproteobacteria) | 5/5 | 3.73% | 0.20% | |
| ASV 29 | Comamonadaceae (Betaproteobacteria) | 5/5 | 2.80% | 0.57% | |
| ASV 35 | Aquabacterium (Betaproteobacteria) | 5/5 | 2.73% | 0.19% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaughn, S.N.; Jackson, C.R. Evaluating Methods of Preserving Aquatic Invertebrates for Microbiome Analysis. Microorganisms 2022, 10, 811. https://doi.org/10.3390/microorganisms10040811
Vaughn SN, Jackson CR. Evaluating Methods of Preserving Aquatic Invertebrates for Microbiome Analysis. Microorganisms. 2022; 10(4):811. https://doi.org/10.3390/microorganisms10040811
Chicago/Turabian StyleVaughn, Stephanie N., and Colin R. Jackson. 2022. "Evaluating Methods of Preserving Aquatic Invertebrates for Microbiome Analysis" Microorganisms 10, no. 4: 811. https://doi.org/10.3390/microorganisms10040811
APA StyleVaughn, S. N., & Jackson, C. R. (2022). Evaluating Methods of Preserving Aquatic Invertebrates for Microbiome Analysis. Microorganisms, 10(4), 811. https://doi.org/10.3390/microorganisms10040811