
Research Progress of the Gut Microbiome in Hybrid Fish


 ,
,
Abstract
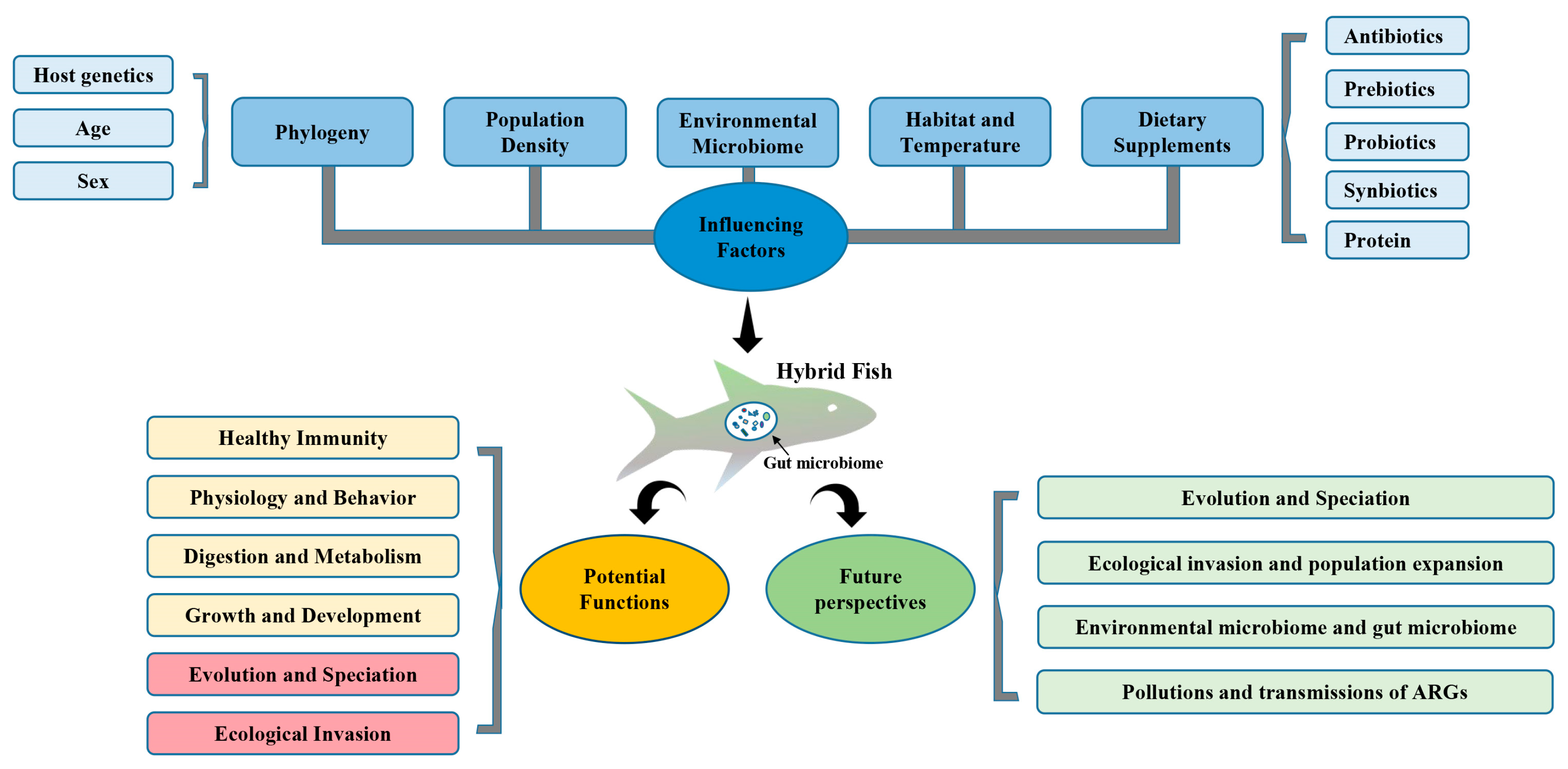
:1. Introduction
2. The Gut Microbiome in Hybrid Fish
2.1. Effects of Diet and Dietary Supplements on the Gut Microbiome and Immune Health of Hybrid Fish
2.1.1. Antibiotics
2.1.2. Probiotics
2.1.3. Prebiotics
2.1.4. Fishmeal Protein Substitutes
2.2. Hybrid Speciation and Gut Microbiome
2.3. The Differences in the Gut Microbiome of the Hybrid Fish and Their Hybrids Origin
2.4. Gut Microbiome Might Promote Ecological Invasion by Hybrid Fish
3. The Potential Impact of Environmental Microbiota
3.1. Habitat Environmental Microbiome Shapes the Early Gut Microbiome of Juvenile Fish
3.2. Do Fish Specifically Select Proteus from the Water Environment?
4. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bordenstein, S.R.; Theis, K.R. Host Biology in Light of the Microbiome: Ten Principles of Holobionts and Hologenomes. PLoS Biol. 2015, 13, e1002226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, E.; Zilber-Rosenberg, I. The hologenome concept of evolution after 10 years. Microbiome 2018, 6, 78. [Google Scholar] [CrossRef] [PubMed]
- McFall-Ngai, M.; Hadfield, M.G.; Bosch, T.C.G.; Carey, H.V.; Domazet-Lošo, T.; Douglas, A.E.; Dubilier, N.; Eberl, G.; Fukami, T.; Gilbert, S.F.; et al. Animals in a bacterial world, a new imperative for the life sciences. Proc. Natl. Acad. Sci. USA 2013, 110, 3229–3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, A.; Xia, K.; Azcarate-Peril, M.A.; Goldman, B.D.; Ahn, M.; Styner, M.A.; Thompson, A.L.; Geng, X.; Gilmore, J.H.; Knickmeyer, R.C. Infant Gut Microbiome Associated With Cognitive Development. Biol. Psychiatry 2018, 83, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Archie, E.A.; Tung, J. Social behavior and the microbiome. Curr. Opin. Behav. Sci. 2015, 6, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Levy, M.; Blacher, E.; Elinav, E. Microbiome, metabolites and host immunity. Curr. Opin. Microbiol. 2017, 35, 8–15. [Google Scholar] [CrossRef]
- Hollister, E.B.; Gao, C.; Versalovic, J. Compositional and functional features of the gastrointestinal microbiome and their effects on human health. Gastroenterology 2014, 146, 1449–1458. [Google Scholar] [CrossRef] [Green Version]
- Kers, J.G.; Velkers, F.; Fischer, E.A.J.; Hermes, G.D.A.; Lamot, D.M.; Stegeman, J.A.; Smidt, H. Take care of the environment: Housing conditions affect the interplay of nutritional interventions and intestinal microbiota in broiler chickens. Anim. Microbiome 2019, 1, 10. [Google Scholar] [CrossRef] [Green Version]
- Youngblut, N.D.; Reischer, G.H.; Walters, W.; Schuster, N.; Walzer, C.; Stalder, G.; Ley, R.E.; Farnleitner, A.H. Host diet and evolutionary history explain different aspects of gut microbiome diversity among vertebrate clades. Nat. Commun. 2019, 10, 2200. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Lozupone, C.A.; Hamady, M.; Knight, R.; Gordon, J.I. Worlds within worlds: Evolution of the vertebrate gut microbiota. Nat. Rev. Microbiol. 2008, 6, 776–788. [Google Scholar] [CrossRef] [Green Version]
- Sharpton, T.J. Role of the Gut Microbiome in Vertebrate Evolution. Msystems 2018, 3, e00174-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Zhang, Y.; Cui, X.; Zhu, Y.; Dai, Q.; Chen, H.; Liu, G.; Yao, R.; Yang, Z. Host Bias in Diet-Source Microbiome Transmission in Wild Cohabitating Herbivores: New Knowledge for the Evolution of Herbivory and Plant Defense. Microbiol. Spectr. 2021, 9, e0075621. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Wu, Q.; Hu, Y.; Huang, G.; Nie, Y.; Yan, L. Conservation metagenomics: A new branch of conservation biology. Sci. China Life Sci. 2019, 62, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johny, T.K.; Puthusseri, R.M.; Bhat, S.G. A primer on metagenomics and next-generation sequencing in fish gut microbiome research. Aquac. Res. 2021, 52, 4574–4600. [Google Scholar] [CrossRef]
- Kim, P.S.; Shin, N.R.; Lee, J.B.; Kim, M.S.; Whon, T.W.; Hyun, D.W.; Yun, J.H.; Jung, M.J.; Kim, J.Y.; Bae, J.W. Host habitat is the major determinant of the gut microbiome of fish. Microbiome 2021, 9, 166. [Google Scholar] [CrossRef]
- Austin, B. The bacterial microflora of fish. Sci. World J. 2002, 2, 558–572. [Google Scholar] [CrossRef] [Green Version]
- Spanggaard, B.; Huber, I.; Nielsen, J.; Nielsen, T.; Appel, K.F.; Gram, L.J.A. The microflora of rainbow trout intestine: A comparison of traditional and molecular identification. Aquaculture 2000, 182, 1–15. [Google Scholar] [CrossRef]
- Pond, M.J.; Stone, D.M.; Alderman, D.J. Comparison of conventional and molecular techniques to investigate the intestinal microflora of rainbow trout (Oncorhynchus mykiss). Aquaculture 2006, 261, 194–203. [Google Scholar] [CrossRef]
- Nayak, S.K. Role of gastrointestinal microbiota in fish. Aquac. Res. 2010, 41, 1553–1573. [Google Scholar] [CrossRef]
- Molinari, L.M.; Scoaris, D.; Pedroso, R.B.; Bittencourt, N.D.L.R.; Filho, B. Bacterial microflora in the gastrointestinal tract of Nile tilapia, Oreochromis niloticus, cultured in a semi-intensive system. Acta Sci. Biol. Sci. 2003, 25, 267–271. [Google Scholar]
- Saha, S.; Roy, R.N.; Sen, S.K.; Ray, A.K. Characterization of cellulase-producing bacteria from the digestive tract of tilapia, Oreochromis mossambica (Peters) and grass carp, Ctenopharyngodon idella (Valenciennes). Aquac. Res. 2010, 37, 380–388. [Google Scholar] [CrossRef]
- Bairagi, A.; Ghosh, K.S.; Sen, S.K.; Ray, A.K. Enzyme producing bacterial flora isolated from fish digestive tracts. Aquac. Int. 2002, 10, 109–121. [Google Scholar] [CrossRef]
- Clements, K.D. Gastrointestinal Microbiology: Volume 1 Gastrointestinal Ecosystems and Fermentations; Mackie, B.A., White, B., Eds.; Springer: Boston, MA, USA, 1997; pp. 156–198. [Google Scholar]
- Carda-Diéguez, M.; Mira, A.; Fouz, B. Pyrosequencing survey of intestinal microbiota diversity in cultured sea bass (Dicentrarchus labrax) fed functional diets. FEMS Microbiol. Ecol. 2014, 87, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Desai, A.R.; Links, M.G.; Collins, S.A.; Mansfield, G.S.; Hill, J. Effects of plant-based diets on the distal gut microbiome of rainbow trout (Oncorhynchus mykiss). Aquaculture 2012, 350–353, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Ghanbari, M.W.; Kneifel, J.A.; Domig, K.J. A new view of the fish gut microbiome: Advances from next-generation sequencing. Aquaculture 2015, 448, 464–475. [Google Scholar] [CrossRef]
- Givens, C.E.; Ransom, B.; Bano, N.; Hollibaugh, J. Comparison of the gut microbiomes of 12 bony fish and 3 shark species. Mar. Ecol. Prog. Ser. 2015, 518, 209–223. [Google Scholar] [CrossRef] [Green Version]
- Ringø, E.; Sperstad, S.; Myklebust, R.; Refstie, S.; Krogdahl, Å. Characterisation of the microbiota associated with intestine of Atlantic cod (Gadus morhua L.): The effect of fish meal, standard soybean meal and a bioprocessed soybean meal. Aquaculture 2006, 261, 829–841. [Google Scholar] [CrossRef]
- Butt, R.L.; Volkoff, H. Gut Microbiota and Energy Homeostasis in Fish. Front. Endocrinol. 2019, 10, 9. [Google Scholar] [CrossRef] [Green Version]
- Handelsman, J.; Rondon, M.R.; Brady, S.F.; Clardy, J.; Goodman, R.M. Molecular biological access to the chemistry of unknown soil microbes: A new frontier for natural products. Chem. Biol. 1998, 5, R245–R249. [Google Scholar] [CrossRef] [Green Version]
- Riesenfeld, C.S.; Schloss, P.D.; Handelsman, J. Metagenomics: Genomic analysis of microbial communities. Annu. Rev. Genet. 2004, 38, 525–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yukgehnaish, K.; Kumar, P.; Sivachandran, P.; Marimuthu, K.; Arshad, A.; Paray, B.A.; Arockiaraj, J. Gut microbiota metagenomics in aquaculture: Factors influencing gut microbiome and its physiological role in fish. Rev. Aquac. 2020, 12, 1903–1927. [Google Scholar] [CrossRef]
- Méndez-Pérez, R.; García-López, R.; Bautista-López, J.S.; Vázquez-Castellanos, J.F.; Peña-Marín, E.S.; Martínez-García, R.; Domínguez-Rodríguez, V.I.; Adams-Schroeder, R.H.; Baltierra-Trejo, E.; Valdés, C.M.; et al. Gut Microbiome Analysis In Adult Tropical Gars (Atractosteus tropicus). bioRxiv 2019. [Google Scholar] [CrossRef]
- Zhang, G.; Li, J.; Zhang, J.; Liang, X.; Zhang, X.; Wang, T.; Yin, S. Integrated Analysis of Transcriptomic, miRNA and Proteomic Changes of a Novel Hybrid Yellow Catfish Uncovers Key Roles for miRNAs in Heterosis. Mol. Cell. Proteom. MCP 2019, 18, 1437–1453. [Google Scholar] [CrossRef]
- Liu, S.; Luo, J.; Chai, J.; Ren, L.; Zhou, Y.; Huang, F.; Liu, X.; Chen, Y.; Zhang, C.; Tao, M.; et al. Genomic incompatibilities in the diploid and tetraploid offspring of the goldfish × common carp cross. Proc. Natl. Acad. Sci. USA 2016, 113, 1327–1332. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Tao, P.; Chen, J.; Wang, R.; Zang, X.; Yin, S. The complete mitochondrial genome of the hybrid of Pelteobagrus fulvidraco (♀) × Pelteobagrus vachelli (♂). Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2016, 27, 4191–4192. [Google Scholar] [CrossRef]
- Gui, J. Scientific frontiers and hot issues in hydrobiology. Chin. Sci. Bull. 2015, 60, 2051–2057. [Google Scholar] [CrossRef]
- Li, W.; Liu, J.; Tan, H.; Yang, C.; Ren, L.; Liu, Q.; Wang, S.; Hu, F.; Xiao, J.; Zhao, R.; et al. Genetic Effects on the Gut Microbiota Assemblages of Hybrid Fish From Parents With Different Feeding Habits. Front. Microbiol. 2018, 9, 2972. [Google Scholar] [CrossRef]
- Li, X.; Yu, Y.; Li, C.; Yan, Q. Comparative study on the gut microbiotas of four economically important Asian carp species. Sci. China Life Sci. 2018, 61, 696–705. [Google Scholar] [CrossRef]
- Zhong, X.; Wang, X.; Zhou, T.; Jin, Y.; Tan, S.; Jiang, C.; Geng, X.; Li, N.; Shi, H.; Zeng, Q.; et al. Genome-Wide Association Study Reveals Multiple Novel QTL Associated with Low Oxygen Tolerance in Hybrid Catfish. Mar. Biotechnol. 2017, 19, 379–390. [Google Scholar] [CrossRef]
- McGinnity, P.; Prodöhl, P.; Ferguson, A.; Hynes, R.; Maoiléidigh, N.O.; Baker, N.; Cotter, D.; O’Hea, B.; Cooke, D.; Rogan, G.; et al. Fitness reduction and potential extinction of wild populations of Atlantic salmon, Salmo salar, as a result of interactions with escaped farm salmon. Proc. Biol. Sci. 2003, 270, 2443–2450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solberg, M.F.; Kvamme, B.O.; Nilsen, F.; Glover, K.A. Effects of environmental stress on mRNA expression levels of seven genes related to oxidative stress and growth in Atlantic salmon Salmo salar L. of farmed, hybrid and wild origin. BMC Res. Notes 2012, 5, 672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Gaughan, S.; Lamer, J.T.; Deng, C.; Hu, W.; Wachholtz, M.; Qin, S.; Nie, H.; Liao, X.; Ling, Q.; et al. Resolving the genetic paradox of invasions: Preadapted genomes and postintroduction hybridization of bigheaded carps in the Mississippi River Basin. Evol. Appl. 2020, 13, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Brucker, R.M.; Bordenstein, S.R. Speciation by symbiosis. Trends Ecol. Evol. 2012, 27, 443–451. [Google Scholar] [CrossRef]
- Wang, J.; Kalyan, S.; Steck, N.; Turner, L.M.; Harr, B.; Künzel, S.; Vallier, M.; Häsler, R.; Franke, A.; Oberg, H.H.; et al. Analysis of intestinal microbiota in hybrid house mice reveals evolutionary divergence in a vertebrate hologenome. Nat. Commun. 2015, 6, 6440. [Google Scholar] [CrossRef] [Green Version]
- Janson, E.M.; Stireman, J.O., 3rd; Singer, M.S.; Abbot, P. Phytophagous insect-microbe mutualisms and adaptive evolutionary diversification. Evol. Int. J. Org. Evol. 2008, 62, 997–1012. [Google Scholar] [CrossRef] [Green Version]
- Guivier, E.; Martin, J.-F.; Pech, N.; Ungaro, A.; Chappaz, R.; Gilles, A. Microbiota Diversity within and between the Tissues of Two Wild Interbreeding Species. Microb. Ecol. 2018, 75, 799–810. [Google Scholar] [CrossRef]
- Talwar, C.; Nagar, S.; Lal, R.; Negi, R.K. Fish Gut Microbiome: Current Approaches and Future Perspectives. Indian J. Microbiol. 2018, 58, 397–414. [Google Scholar] [CrossRef]
- Tran, N.T.; Wang, G.-T.; Wu, S.-G. A review of intestinal microbes in grass carp Ctenopharyngodon idellus (Valenciennes). Aquac. Res. 2017, 48, 3287–3297. [Google Scholar] [CrossRef]
- Tan, H.Y.; Chen, S.W.; Hu, S.Y. Improvements in the growth performance, immunity, disease resistance, and gut microbiota by the probiotic Rummeliibacillus stabekisii in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2019, 92, 265–275. [Google Scholar] [CrossRef]
- Giatsis, C.; Sipkema, D.; Ramiro-Garcia, J.; Bacanu, G.M.; Abernathy, J.; Verreth, J.; Smidt, H.; Verdegem, M. Probiotic legacy effects on gut microbial assembly in tilapia larvae. Sci. Rep. 2016, 6, 33965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geraylou, Z.; Souffreau, C.; Rurangwa, E.; De Meester, L.; Courtin, C.M.; Delcour, J.A.; Buyse, J.; Ollevier, F. Effects of dietary arabinoxylan-oligosaccharides (AXOS) and endogenous probiotics on the growth performance, non-specific immunity and gut microbiota of juvenile Siberian sturgeon (Acipenser baerii). Fish Shellfish Immunol. 2013, 35, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Wu, Z.; Zhang, P.; Qu, Y.; Fu, S.; Liu, J.; Chen, X. Effects of dietary xylo-oligosaccharide on intestinal microflora of grass carp(Ctenopharyngodon idellus). Microbiology 2011, 38, 317–321. [Google Scholar]
- Xia, Y.; Lu, M.; Chen, G.; Cao, J.; Gao, F.; Wang, M.; Liu, Z.; Zhang, D.; Zhu, H.; Yi, M. Effects of dietary Lactobacillus rhamnosus JCM1136 and Lactococcus lactis subsp. lactis JCM5805 on the growth, intestinal microbiota, morphology, immune response and disease resistance of juvenile Nile tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 2018, 76, 368–379. [Google Scholar] [CrossRef]
- Anderson, D.P.; Moritomo, T.; de Grooth, R. Neutrophil, glass-adherent, nitroblue tetrazolium assay gives early indication of immunization effectiveness in rainbow trout. Veter Immunol. Immunopathol. 1992, 30, 419–429. [Google Scholar] [CrossRef]
- Li, W.; Shen, T.; Chen, N.; Deng, B.; Fu, L.; Zhou, X. Effects of dietary Bacillus subtilis on digestive enzyme activity and intestinal microflora in grass carp Ctenopharyngodon idellus. J. Dalian Ocean Univ. 2012, 27, 221–225. [Google Scholar]
- Wang, Y. Use of probiotics Bacillus coagulans, Rhodopseudomonas palustris and Lactobacillus acidophilus as growth promoters in grass carp (Ctenopharyngodon idella) fingerlings. Aquac. Nutr. 2011, 17, E372–E378. [Google Scholar] [CrossRef]
- Wu, Z.X.; Feng, X.; Xie, L.L.; Peng, X.Y.; Yuan, J.; Chen, X.X. Effect of probiotic Bacillus subtilis Ch9 for grass carp, Ctenopharyngodon idella (Valenciennes, 1844), on growth performance, digestive enzyme activities and intestinal microflora. J. Appl. Ichthyol. 2012, 28, 721–727. [Google Scholar] [CrossRef]
- Zhou, Y.; Yuan, X.C.; Liang, X.F.; Fang, L.; Li, J.; Guo, X.Z.; Bai, X.L.; He, S. Enhancement of growth and intestinal flora in grass carp: The effect of exogenous cellulase. Aquaculture 2013, 416, 1–7. [Google Scholar] [CrossRef]
- Yuan, X.; Zhou, Y.; Liang, X.F.; Guo, X.; Fang, L.; Li, J.; Liu, L.; Li, B. Effect of dietary glutathione supplementation on the biological value of rapeseed meal to juvenile grass carp, Ctenopharyngodon idellus. Aquac. Nutr. 2015, 21, 73–84. [Google Scholar] [CrossRef]
- Yao, D.; Zou, Q.; Liu, W.; Xie, S.; Zhou, A.; Chen, J.; Zou, J. Effects of Bacillus licheniformis and xylo-oligosaccharide on growth performance, intestinal microflora and enzyme activities in grass carp Ctenopharyngodon idella. J. Dalian Ocean Univ. 2014, 29, 136–140. [Google Scholar]
- Koo, H.; Hakim, J.A.; Powell, M.L.; Kumar, R.; Eipers, P.G.; Morrow, C.D.; Crowley, M.; Lefkowitz, E.J.; Watts, S.A.; Bej, A.K. Metagenomics approach to the study of the gut microbiome structure and function in zebrafish Danio rerio fed with gluten formulated diet. J. Microbiol. Methods 2017, 135, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rurangwa, E.; Sipkema, D.; Kals, J.; Ter Veld, M.; Forlenza, M.; Bacanu, G.M.; Smidt, H.; Palstra, A.P. Impact of a novel protein meal on the gastrointestinal microbiota and the host transcriptome of larval zebrafish Danio rerio. Front. Physiol. 2015, 6, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udayangani, R.M.C.; Dananjaya, S.H.S.; Nikapitiya, C.; Heo, G.-J.; Lee, J.; De Zoysa, M. Metagenomics analysis of gut microbiota and immune modulation in zebrafish (Danio rerio) fed chitosan silver nanocomposites. Fish Shellfish Immunol. 2017, 66, 173–184. [Google Scholar] [CrossRef]
- Carlson, J.M.; Leonard, A.B.; Hyde, E.R.; Petrosino, J.F.; Primm, T.P. Microbiome disruption and recovery in the fish Gambusia affinis following exposure to broad-spectrum antibiotic. Infect. Drug Resist. 2017, 10, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Michl, S.C.; Ratten, J.M.; Beyer, M.; Hasler, M.; LaRoche, J.; Schulz, C. The malleable gut microbiome of juvenile rainbow trout (Oncorhynchus mykiss): Diet-dependent shifts of bacterial community structures. PLoS ONE 2017, 12, e0177735. [Google Scholar] [CrossRef] [Green Version]
- Huyben, D.; Nyman, A.; Vidaković, A.; Passoth, V.; Moccia, R.; Kiessling, A.; Dicksved, J.; Lundh, T. Effects of dietary inclusion of the yeasts Saccharomyces cerevisiae and Wickerhamomyces anomalus on gut microbiota of rainbow trout. Aquaculture 2017, 473, 528–537. [Google Scholar] [CrossRef]
- Lyons, P.P.; Turnbull, J.F.; Dawson, K.A.; Crumlish, M. Effects of low-level dietary microalgae supplementation on the distal intestinal microbiome of farmed rainbow trout Oncorhynchus mykiss (Walbaum). Aquac. Res. 2017, 48, 2438–2452. [Google Scholar] [CrossRef]
- Zhai, Q.; Yu, L.; Li, T.; Zhu, J.; Zhang, C.; Zhao, J.; Zhang, H.; Chen, W. Effect of dietary probiotic supplementation on intestinal microbiota and physiological conditions of Nile tilapia (Oreochromis niloticus) under waterborne cadmium exposure. Antonie Van Leeuwenhoek 2017, 110, 501–513. [Google Scholar] [CrossRef]
- Yu, L.; Qiao, N.; Li, T.; Yu, R.; Chen, W.J.P. Dietary supplementation with probiotics regulates gut microbiota structure and function in Nile tilapia exposed to aluminum. PeerJ 2019, 7, e6963. [Google Scholar] [CrossRef]
- Hortillosa, E.; Amar, M.; Nuñal, S.; Pedroso, F.; Ferriols, V.M.E. Effects of putative dietary probiotics from the gut of milkfish (Chanos chanos) on the growth performance and intestinal enzymatic activities of juvenile Nile tilapia (Oreochromis niloticus). Aquac. Res. 2021, 53, 98–108. [Google Scholar] [CrossRef]
- Zhang, M.; Dong, B.; Lai, X.; Chen, Z.; Hou, L.; Shu, R.; Huang, Y.; Shu, H. Effects of Clostridium butyricum on growth, digestive enzyme activity, antioxidant capacity and gut microbiota in farmed tilapia (Oreochromis niloticus). Aquac. Res. 2021, 52, 1573–1584. [Google Scholar] [CrossRef]
- Foysal, J.; Alam, M.; Momtaz, F.; Chaklader, M.R.; Siddik, M.A.B.; Rahman, M. Dietary supplementation of garlic (Allium sativum) modulates gut microbiota and health status of tilapia (Oreochromis niloticus) against Streptococcus iniae infection. Aquac. Res. 2019, 50, 2107–2116. [Google Scholar] [CrossRef]
- Tachibana, L.; Telli, G.S.; Dias, D.d.C.; Gonçalves, G.S.; Guimarães, M.C.; Ishikawa, C.M.; Cavalcante, R.B.; Natori, M.M.; Fernandez Alarcon, M.F.; Tapia-Paniagua, S.; et al. Bacillus subtilis and Bacillus licheniformis in diets for Nile tilapia (Oreochromis niloticus): Effects on growth performance, gut microbiota modulation and innate immunology. Aquac. Res. 2021, 52, 1630–1642. [Google Scholar] [CrossRef]
- Liao, Q.; Zhen, Y.; Qin, Y.; Jiang, Q.; Lan, T.; Huang, L.; and Shen, P. Effects of dietary Metschnikowia sp. GXUS03 on growth, immunity, gut microbiota and Streptococcus agalactiae resistance of Nile tilapia (Oreochromis niloticus). Aquac. Res. 2022, 53, 1918–1927. [Google Scholar] [CrossRef]
- Piazzon, M.C.; Calduch-Giner, J.A.; Fouz, B.; Estensoro, I.; Simó-Mirabet, P.; Puyalto, M.; Karalazos, V.; Palenzuela, O.; Sitjà-Bobadilla, A.; Pérez-Sánchez, J. Under control: How a dietary additive can restore the gut microbiome and proteomic profile, and improve disease resilience in a marine teleostean fish fed vegetable diets. Microbiome 2017, 5, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legrand, T.; Catalano, S.R.; Wos-Oxley, M.L.; Wynne, J.W.; Weyrich, L.S.; Oxley, A.P.A. Antibiotic-induced alterations and repopulation dynamics of yellowtail kingfish microbiota. Anim. Microbiome 2020, 2, 26. [Google Scholar] [CrossRef] [PubMed]
- Sáenz, J.S.; Marques, T.V.; Barone, R.S.C.; Cyrino, J.E.P.; Kublik, S.; Nesme, J.; Schloter, M.; Rath, S.; Vestergaard, G. Oral administration of antibiotics increased the potential mobility of bacterial resistance genes in the gut of the fish Piaractus mesopotamicus. Microbiome 2019, 7, 24. [Google Scholar] [CrossRef]
- Munir, M.B.; Marsh, T.L.; Blaud, A.; Hashim, R.; Janti Anak Joshua, W.; Mohd Nor, S.A. Analysing the effect of dietary prebiotics and probiotics on gut bacterial richness and diversity of Asian snakehead fingerlings using T-RFLP method. Aquac. Res. 2018, 49, 3350–3361. [Google Scholar] [CrossRef]
- Meng, X.; Hu, W.; Wu, S.; Zhu, Z.; Lu, R.; Yang, G.; Qin, C.; Yang, L.; Nie, G. Chinese yam peel enhances the immunity of the common carp (Cyprinus carpio L.) by improving the gut defence barrier and modulating the intestinal microflora. Fish Shellfish Immunol. 2019, 95, 528–537. [Google Scholar] [CrossRef]
- Reyshari, A.; Mohammadiazarm, H.; Mohammadian, T.; Mozanzadeh, M.T. Effects of sodium diformate on growth performance, gut microflora, digestive enzymes and innate immunological parameters of Asian sea bass (Lates calcarifer) juveniles. Aquac. Nutr. 2019, 25, 1135–1144. [Google Scholar] [CrossRef]
- Guimarães, M.C.; Moriñigo, M.Á.; Moyano, F.J.; Tachibana, L. Oral administration of Bacillus subtilis and Lactobacillus plantarum modulates the gut microbiota and increases the amylase activity of Nile tilapia (Oreochromis niloticus). Aquac. Int. 2021, 29, 91–104. [Google Scholar] [CrossRef]
- Psofakis, P.; Meziti, A.; Berillis, P.; Mente, E.; Kormas, K.A.; Karapanagiotidis, I.T. Effects of Dietary Fishmeal Replacement by Poultry By-Product Meal and Hydrolyzed Feather Meal on Liver and Intestinal Histomorphology and on Intestinal Microbiota of Gilthead Seabream (Sparus aurata). Appl. Sci. 2021, 11, 8806. [Google Scholar] [CrossRef]
- Rimoldi, S.; Gini, E.; Koch, J.F.A.; Iannini, F.; Brambilla, F.; Terova, G. Effects of hydrolyzed fish protein and autolyzed yeast as substitutes of fishmeal in the gilthead sea bream (Sparus aurata) diet, on fish intestinal microbiome. BMC Vet. Res. 2020, 16, 118. [Google Scholar]
- Rimoldi, S.; Torrecillas, S.; Montero, D.; Gini, E.; Makol, A.; Valdenegro, V.V.; Izquierdo, M.; Terova, G. Assessment of dietary supplementation with galactomannan oligosaccharides and phytogenics on gut microbiota of European sea bass (Dicentrarchus Labrax) fed low fishmeal and fish oil based diet. PLoS ONE 2020, 15, e0231494. [Google Scholar] [CrossRef] [Green Version]
- Abid, A.; Davies, S.J.; Waines, P.; Emery, M.; Castex, M.; Gioacchini, G.; Carnevali, O.; Bickerdike, R.; Romero, J.; Merrifield, D.L. Dietary synbiotic application modulates Atlantic salmon (Salmo salar) intestinal microbial communities and intestinal immunity. Fish Shellfish Immunol. 2013, 35, 1948–1956. [Google Scholar] [CrossRef]
- Pereira, G.d.V.; Pereira, S.A.; Soares, A.; Mouriño, J.L.P.; Merrifield, D. Autochthonous probiotic bacteria modulate intestinal microbiota of Pirarucu, Arapaima gigas. J. World Aquac. Soc. 2019, 50, 1152–1167. [Google Scholar] [CrossRef]
- Xie, M.; Zhou, W.; Xie, Y.; Li, Y.; Zhang, Z.; Yang, Y.; Olsen, R.E.; Ran, C.; Zhou, Z. Effects of Cetobacterium somerae fermentation product on gut and liver health of common carp (Cyprinus carpio) fed diet supplemented with ultra-micro ground mixed plant proteins. Aquaculture 2021, 543, 736943. [Google Scholar] [CrossRef]
- Xu, Q.; Yang, Z.; Chen, S.; Zhu, W.; Xiao, S.; Liu, J.; Wang, H.; Lan, S. Effects of Replacing Dietary Fish Meal by Soybean Meal Co-Fermented Using Bacillus subtilis and Enterococcus faecium on Serum Antioxidant Indices and Gut Microbiota of Crucian Carp Carassius auratus. Fishes 2022, 7, 54. [Google Scholar] [CrossRef]
- González-Félix, M.L.; Gatlin, D.M.; Urquidez-Bejarano, P.; de la Reé-Rodríguez, C.; Duarte-Rodríguez, L.; Sánchez, F.; Casas-Reyes, A.; Yamamoto, F.Y.; Ochoa-Leyva, A.; Perez-Velazquez, M. Effects of commercial dietary prebiotic and probiotic supplements on growth, innate immune responses, and intestinal microbiota and histology of Totoaba macdonaldi. Aquaculture 2018, 491, 239–251. [Google Scholar] [CrossRef]
- Larios-Soriano, E.; Zavala, R.C.; López, L.M.; Gómez-Gil, B.; Ramírez, D.T.; Sanchez, S.; Canales, K.; Galaviz, M.A. Soy protein concentrate effects on gut microbiota structure and digestive physiology of Totoaba macdonaldi. J. Appl. Microbiol. 2022, 132, 1384–1396. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xie, S.; Chi, S.; Zhang, S.; Cao, J.; Tan, B. Short-term dietary antibiotics altered the intestinal microbiota and improved the lipid metabolism in hybrid grouper fed medium and high-lipid diets. Aquaculture 2022, 547, 737453. [Google Scholar] [CrossRef]
- Zhou, S.; Song, D.; Zhou, X.; Mao, X.; Zhou, X.; Wang, S.; Wei, J.; Huang, Y.; Wang, W.; Xiao, S.-M.; et al. Characterization of Bacillus subtilis from gastrointestinal tract of hybrid Hulong grouper (Epinephelus fuscoguttatus × E. lanceolatus) and its effects as probiotic additives. Fish Shellfish Immunol. 2019, 84, 1115–1124. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, X.; Chen, D.; Wang, Y.; Zhang, A.; Zhou, H. Comparison of adhesive gut bacteria composition, immunity, and disease resistance in juvenile hybrid tilapia fed two different Lactobacillus strains. Fish Shellfish Immunol. 2013, 35, 54–62. [Google Scholar] [CrossRef]
- Wu, X.; Teame, T.; Hao, Q.; Ding, Q.; Liu, H.; Ran, C.; Yang, Y.; Zhang, Y.; Zhou, Z.; Duan, M.; et al. Use of a paraprobiotic and postbiotic feed supplement (HWF™) improves the growth performance, composition and function of gut microbiota in hybrid sturgeon (Acipenser baerii x Acipenser schrenckii). Fish Shellfish Immunol. 2020, 104, 36–45. [Google Scholar] [CrossRef]
- Hahor, W.; Thongprajukaew, K.; Suanyuk, N. Effects of dietary supplementation of oligosaccharides on growth performance, gut health and immune response of hybrid catfish (Pangasianodon gigas × Pangasianodon hypophthalmus). Aquaculture 2019, 507, 97–107. [Google Scholar] [CrossRef]
- Wang, Q.; Ayiku, S.; Liu, H.; Tan, B.; Dong, X.; Chi, S.; Yang, Q.; Zhang, S.; Zhou, W. Effects of dietary ESTAQUA® yeast culture supplementation on growth, immunity, intestinal microbiota and disease-resistance against Vibrio harveyi in hybrid grouper (♀ Epinephelus fuscoguttatus × ♂ E. lanceolatus). Aquac. Rep. 2022, 22, 100922. [Google Scholar] [CrossRef]
- Xu, G.; Xing, W.; Li, T.; Ma, Z.; Liu, C.; Jiang, N.; Luo, L. Effects of dietary raffinose on growth, non-specific immunity, intestinal morphology and microbiome of juvenile hybrid sturgeon (Acipenser baeri Brandt ♀ × A. schrenckii Brandt ♂). Fish Shellfish Immunol. 2018, 72, 237–246. [Google Scholar] [CrossRef]
- Chen, G.; Yin, B.; Liu, H.; Tan, B.; Dong, X.; Yang, Q.; Chi, S.; Zhang, S. Supplementing Chitosan Oligosaccharide Positively Affects Hybrid Grouper (Epinephelus fuscoguttatus & FEMALE; x E. lanceolatus & MALE;) Fed Dietary Fish Meal Replacement With Cottonseed Protein Concentrate: Effects on Growth, Gut Microbiota, Antioxidant Function and Immune Response. Front. Mar. Sci. 2021, 8, 707627. [Google Scholar] [CrossRef]
- Chen, G.; Yin, B.; Liu, H.; Tan, B.; Dong, X.; Yang, Q.; Chi, S.; Zhang, S. Effects of fishmeal replacement with cottonseed protein concentrate on growth, digestive proteinase, intestinal morphology and microflora in pearl gentian grouper (♀ Epinephelus fuscoguttatus × ♂ Epinephelus lanceolatu). Aquac. Res. 2020, 51, 2870–2884. [Google Scholar] [CrossRef]
- Ye, G.; Dong, X.; Yang, Q.; Chi, S.; Liu, H.; Zhang, H.; Tan, B.; Zhang, S. Dietary replacement of fish meal with peanut meal in juvenile hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂): Growth performance, immune response and intestinal microbiota. Aquac. Rep. 2020, 17, 100327. [Google Scholar] [CrossRef]
- Ye, G.; Dong, X.; Yang, Q.; Chi, S.; Liu, H.; Zhang, H.; Tan, B.; Zhang, S. Low-gossypol cottonseed protein concentrate used as a replacement of fish meal for juvenile hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂): Effects on growth performance, immune responses and intestinal microbiota. Aquaculture 2020, 524, 735309. [Google Scholar] [CrossRef]
- He, Y.; Guo, X.; Tan, B.; Dong, X.; Yang, Q.; Liu, H.; Zhang, S.; Chi, S. Partial fishmeal protein replacement with peptides from swine blood modulates the nutritional status, immune response, and intestinal microbiota of hybrid groupers (female Epinephelus fuscoguttatus× male E. lanceolatus). Aquaculture 2021, 533, 736154. [Google Scholar] [CrossRef]
- Fowler, E.C.; Poudel, P.; White, B.; St-Pierre, B.; Brown, M. Effects of a Bioprocessed Soybean Meal Ingredient on the Intestinal Microbiota of Hybrid Striped Bass, Morone chrysops × M. saxatilis. Microorganisms 2021, 9, 1032. [Google Scholar] [CrossRef]
- Sevellec, M.; Laporte, M.; Bernatchez, A.; Derome, N.; Bernatchez, L. Evidence for host effect on the intestinal microbiota of whitefish (Coregonus sp.) species pairs and their hybrids. Ecol. Evol. 2019, 9, 11762–11774. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Zhang, Z.; Chen, H.; Lamer, J.T.; Wang, J.; Wei, W.; Fu, L.; Tang, M.; Wang, C.; Lu, G. Gut microbiomes of bigheaded carps and hybrids provide insights into invasion: A hologenome perspective. Evol. Appl. 2021, 14, 735–745. [Google Scholar] [CrossRef]
- Deng, Y.; Zhang, Y.; Chen, H.; Xu, L.; Wang, Q.; Feng, J. Gut-Liver Immune Response and Gut Microbiota Profiling Reveal the Pathogenic Mechanisms of Vibrio harveyi in Pearl Gentian Grouper (Epinephelus lanceolatus ♂ × E. fuscoguttatus ♀). Front. Immunol. 2020, 11, 607754. [Google Scholar] [CrossRef]
- He, Y.; Liang, J.; Dong, X.; Yang, Q.; Liu, H.; Zhang, S.; Chi, S.; Tan, B. Glutamine alleviates β-conglycinin-induced enteritis in juvenile hybrid groupers Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂ by suppressing the MyD88/NF-κB pathway. Aquaculture 2022, 549, 737735. [Google Scholar] [CrossRef]
- Liu, X.; Shi, H.; He, Q.; Lin, F.; Wang, Q.; Xiao, S.; Dai, Y.; Zhang, Y.; Yang, H.; Zhao, H. Effect of starvation and refeeding on growth, gut microbiota and non-specific immunity in hybrid grouper (Epinephelus fuscoguttatus♀ × E. lanceolatus♂). Fish Shellfish Immunol. 2020, 97, 182–193. [Google Scholar] [CrossRef]
- Fowler, E.; St-Pierre, B.; Poudel, P.; White, B.; Brown, M. 183 Effects of Protein Supplementation on the Gut Bacterial Community Composition of Hybrid Striped Bass, Morone Chrysops X M. Saxatilis. J. Anim. Sci. 2021, 99, 88–89. [Google Scholar] [CrossRef]
- Ran, C.; Hu, J.; Liu, W.; Liu, Z.; He, S.; Bui Chau Truc, D.; Nguyen Ngoc, D.; Ooi, E.L.; Zhou, Z. Thymol and Carvacrol Affect Hybrid Tilapia through the Combination of Direct Stimulation and an Intestinal Microbiota-Mediated Effect: Insights from a Germ-Free Zebrafish Model. J. Nutr. 2016, 146, 1132–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, C.; Zhang, Y.; Liu, W.; Xu, L.; Yang, Y.; Zhou, Z. Effects of chito-oligosaccharides supplementation on growth performance, intestinal cytokine expression, autochthonous gut bacteria and disease resistance in hybrid tilapia Oreochromis niloticus ♀ × Oreochromis aureus ♂. Fish Shellfish Immunol. 2014, 40, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Qiang, J.; He, J.; Tao, Y.; Bao, J.; Xu, P. Physiological parameters and gut microbiome associated with different dietary lipid levels in hybrid yellow catfish (Tachysurus fulvidraco♀× Pseudobagrus vachellii♂). Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 37, 100777. [Google Scholar] [CrossRef] [PubMed]
- Ye, G.; Dong, X.; Yang, Q.; Chi, S.; Liu, H.; Zhang, H.; Tan, B.; Zhang, S. A formulated diet improved digestive capacity, immune function and intestinal microbiota structure of juvenile hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂) when compared with chilled trash fish. Aquaculture 2020, 523, 735230. [Google Scholar] [CrossRef]
- Hao, Q.; Teame, T.; Wu, X.; Ding, Q.; Ran, C.; Yang, Y.; Xing, Y.; Zhang, Z.; Zhou, Z. Influence of diet shift from bloodworm to formulated feed on growth performance, gut microbiota structure and function in early juvenile stages of hybrid sturgeon (Acipenser baerii × Acipenser schrenckii). Aquaculture 2021, 533, 736165. [Google Scholar] [CrossRef]
- Wang, L.; Tian, Y.; Li, Z.; Li, Z.; Chen, S.; Li, L.; Li, W.; Wang, Q.; Lin, H.; Li, B. Comparison of the gut microbiota composition between asymptomatic and diseased Epinephelus moara ♀ × Epinephelus lanceolatus ♂ with nervous necrosis virus infection. Aquac. Res. 2022, 53, 633–641. [Google Scholar] [CrossRef]
- Hlordzi, V.; Wang, J.; Li, T.; Cui, Z.; Tan, B.; Liu, H.; Yang, Q.; Dong, X.; Zhang, S.; Chi, S. Effects of Lower Fishmeal with Hydrolyzed Fish Protein Powder on the Growth Performance and Intestinal Development of Juvenile Pearl Gentian Grouper (Epinephelus fuscoguttatus ♀ and Epinephelus lanceolatus ♂). Front. Mar. Sci. 2022, 9, 398. [Google Scholar] [CrossRef]
- Huang, X.; Zhong, L.; Fan, W.; Feng, Y.; Xiong, G.; Liu, S.; Wang, K.; Geng, Y.; Ouyang, P.; Chen, D.; et al. Enteritis in hybrid sturgeon (Acipenser schrenckii♂ × Acipenser baeri♀) caused by intestinal microbiota disorder. Aquac. Rep. 2020, 18, 100456. [Google Scholar] [CrossRef]
- Long, S.; You, Y.; Dong, X.; Tan, B.; Zhang, S.; Chi, S.; Yang, Q.; Liu, H.; Xie, S.; Yang, Y.; et al. Effect of dietary oxidized fish oil on growth performance, physiological homeostasis and intestinal microbiome in hybrid grouper (♀ Epi-nephelus fuscoguttatus × ♂ Epinephelus lanceolatus). Aquac. Rep. 2022, 24, 101130. [Google Scholar] [CrossRef]
- Long, S.; Dong, X.; Yan, X.; Liu, H.; Tan, B.; Zhang, S.; Chi, S.; Yang, Q.; Liu, H.; Yang, Y.; et al. The effect of oxidized fish oil on antioxidant ability, histology and transcriptome in intestine of the juvenile hybrid grouper (♀ Epinephelus fuscoguttatus × ♂ Epinephelus lanceolatus). Aquac. Rep. 2022, 22, 100921. [Google Scholar] [CrossRef]
- Yan, X.; Chen, Y.; Dong, X.; Tan, B.; Liu, H.; Zhang, S.; Chi, S.; Yang, Q.; Liu, H.; Yang, Y. Ammonia Toxicity Induces Oxidative Stress, Inflammatory Response and Apoptosis in Hybrid Grouper (♀ Epinephelus fuscoguttatus × ♂ E. lanceolatu). Front. Mar. Sci. 2021, 8, 100880. [Google Scholar] [CrossRef]
- Wei, H.; Li, R.; Yang, Q.; Tan, B.; Ray, G.W.; Dong, X.; Chi, S.; Liu, H.; Zhang, S. Effects of Zn on growth performance, immune enzyme activities, resistance to disease and intestinal flora for juvenile pearl gentian grouper(Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀) under low fishmeal diet. Aquac. Rep. 2021, 21, 100880. [Google Scholar] [CrossRef]
- Wu, X.; Hao, Q.; Teame, T.; Ding, Q.; Liu, H.; Ran, C.; Yang, Y.; Xia, L.; Wei, S.; Zhou, Z.; et al. Gut microbiota induced by dietary GWF® contributes to growth promotion, immune regulation and disease resistance in hybrid sturgeon (Acipenserbaerii x Acipenserschrenckii): Insights from a germ-free zebrafish model. Aquaculture 2020, 520, 734966. [Google Scholar] [CrossRef]
- Limbu, S.M.; Chen, L.-Q.; Zhang, M.-L.; Du, Z.-Y. A global analysis on the systemic effects of antibiotics in cultured fish and their potential human health risk: A review. Rev. Aquac. 2021, 13, 1015–1059. [Google Scholar] [CrossRef]
- Sx, A.; Yin, P.; Tian, L.; Liu, Y.; Tan, B.; Niu, J. Interactions between dietary lipid levels and chronic exposure of legal aquaculture dose of sulfamethoxazole in juvenile largemouth bass Micropterus salmoides. Aquat. Toxicol. 2020, 229, 105670. [Google Scholar]
- Limbu, S.M.; Ma, Q.; Zhang, M.-L.; Du, Z.-Y. High fat diet worsens the adverse effects of antibiotic on intestinal health in juvenile Nile tilapia (Oreochromis niloticus). Sci. Total Environ. 2019, 680, 169–180. [Google Scholar] [CrossRef]
- Zhou, L.; Limbu, S.M.; Qiao, F.; Du, Z.Y.; Zhang, M.J.Z. Influence of Long-Term Feeding Antibiotics on the Gut Health of Zebrafish. Zebrafish 2018, 15, 340–348. [Google Scholar] [CrossRef]
- Nakano, T.; Hayashi, S.; Nagamine, N. Effect of excessive doses of oxytetracycline on stress-related biomarker expression in coho salmon. Environ. Sci. Pollut. Res. Int. 2018, 25, 7121–7128. [Google Scholar] [CrossRef]
- Zhou, Z.C.; Lin, Z.J.; Shuai, X.Y.; Zheng, J.; Meng, L.X.; Zhu, L.; Sun, Y.J.; Shang, W.C.; Chen, H. Temporal variation and sharing of antibiotic resistance genes between water and wild fish gut in a peri-urban river. J. Environ. Sci. 2021, 103, 12–19. [Google Scholar] [CrossRef]
- Carbone, D.; Faggio, C. Importance of prebiotics in aquaculture as immunostimulants. Effects on immune system of Sparus aurata and Dicentrarchus labrax. Fish Shellfish Immunol. 2016, 54, 172–178. [Google Scholar] [CrossRef]
- Song, S.K.; Beck, B.R.; Kim, D.; Park, J.; Kim, J.; Kim, H.D.; Ringø, E. Prebiotics as immunostimulants in aquaculture: A review. Fish Shellfish Immunol. 2014, 40, 40–48. [Google Scholar] [CrossRef]
- Taverniti, V.; Guglielmetti, S. The immunomodulatory properties of probiotic microorganisms beyond their viability (ghost probiotics: Proposal of paraprobiotic concept). Genes Nutr. 2011, 6, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Wang, M.; Gao, F.; Lu, M.; Chen, G. Effects of dietary probiotic supplementation on the growth, gut health and disease resistance of juvenile Nile tilapia (Oreochromis niloticus). Anim. Nutr. 2020, 6, 69–79. [Google Scholar] [CrossRef]
- Beck, B.R.; Kim, D.; Jeon, J.; Lee, S.M.; Kim, H.K.; Kim, O.J.; Lee, J.I.; Suh, B.S.; Do, H.K.; Lee, K.H.J.F.; et al. The effects of combined dietary probiotics Lactococcus lactis BFE920 and Lactobacillus plantarum FGL0001 on innate immunity and disease resistance in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2015, 42, 177–183. [Google Scholar] [CrossRef]
- Liu, L.; Wu, R.; Zhang, J.; Shang, N.; Li, P. D-Ribose Interferes with Quorum Sensing to Inhibit Biofilm Formation of Lactobacillus paraplantarum L-ZS9. Front. Microbiol. 2017, 8, 1860. [Google Scholar] [CrossRef]
- Chai, P.C.; Song, X.L.; Chen, G.F.; Xu, H.; Huang, J. Dietary supplementation of probiotic Bacillus PC465 isolated from the gut of Fenneropenaeus chinensis improves the health status and resistance of Litopenaeus vannamei against white spot syndrome virus. Fish Shellfish Immunol. 2016, 54, 602–611. [Google Scholar] [CrossRef]
- Staykov, Y.; Spring, P.; Denev, S.; Sweetman, J.W. Effect of a mannan oligosaccharide on the growth performance and immune status of rainbow trout (Oncorhynchus mykiss). Aquac. Int. 2007, 15, 153–161. [Google Scholar] [CrossRef]
- Foligné, B.; Dewulf, J.; Breton, J.; Claisse, O.; Lonvaud-Funel, A.; Pot, B. Probiotic properties of non-conventional lactic acid bacteria: Immunomodulation by Oenococcus oeni. Int. J. Food Microbiol. 2010, 140, 136–145. [Google Scholar] [CrossRef]
- Panigrahi, A.; Kiron, V.; Satoh, S.; Hirono, I.; Kobayashi, T.; Sugita, H.; Puangkaew, J.; Aoki, T. Immune modulation and expression of cytokine genes in rainbow trout Oncorhynchus mykiss upon probiotic feeding. Dev. Comp. Immunol. 2007, 31, 372–382. [Google Scholar] [CrossRef]
- Yang, M.; Wang, X.; Chen, D.; Wang, Y.; Zhang, A.; Zhou, H. TGF-β1 exerts opposing effects on grass carp leukocytes: Implication in teleost immunity, receptor signaling and potential self-regulatory mechanisms. PLoS ONE 2012, 7, e35011. [Google Scholar] [CrossRef] [Green Version]
- Kadowaki, T.; Yasui, Y.; Takahashi, Y.; Kohchi, C.; Soma, G.; Inagawa, H. Comparative immunological analysis of innate immunity activation after oral administration of wheat fermented extract to teleost fish. Anticancer Res. 2009, 29, 4871–4877. [Google Scholar] [PubMed]
- He, S.; Liu, W.; Zhou, Z. Evaluation of probiotic strain Bacillus subtilis C-3102 as a feed supplement for koi carp (Cyprinus carpio). J. Aquac. Res. Dev. 2011, S1, 005. [Google Scholar] [CrossRef] [Green Version]
- Gibson, G.R.; Probert, H.M.; Loo, J.V.; Rastall, R.A.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Updating the concept of prebiotics. Nutr. Res. Rev. 2004, 17, 259–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, G.R.; Roberfroid, M.B. Dietary Modulation of the Human Colonic Microbiota: Introducing the Concept of Prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Zoheiri, F.; Dadar, M.; Rufchaei, R.; Ringø, E. Dietary galactooligosaccharide elicits positive effects on non-specific immune parameters and growth performance in Caspian white fish (Rutilus frisii kutum) fry. Fish Shellfish Immunol. 2016, 56, 467–472. [Google Scholar] [CrossRef]
- Watzl, B.; Girrbach, S.; Roller, M. Inulin, oligofructose and immunomodulation. Br. J. Nutr. 2005, 93 (Suppl. 1), S49–S55. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Khalili, M.; Rostami, H.K.; Esteban, M. Immunology, Dietary galactooligosaccharide affects intestinal microbiota, stress resistance, and performance of Caspian roach (Rutilus rutilus) fry. Fish Shellfish Immunol. 2013, 35, 1416–1420. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Sharifian, M.; Vesaghi, M.J.; Khalili, M.; Esteban, M. The effects of dietary xylooligosaccharide on mucosal parameters, intestinal microbiota and morphology and growth performance of Caspian white fish (Rutilus frisii kutum) fry—ScienceDirect. Fish Shellfish Immunol. 2014, 39, 231–236. [Google Scholar] [CrossRef]
- Ali, S.S.R.; Ambasankar, K.; Nandakumar, S.; Praveena, P.E.; Syamadayal, J. Effect of dietary prebiotic inulin on growth, body composition and gut microbiota of Asian seabass (Lates calcarifer). Anim. Feed Sci. Technol. 2016, 217, 87–94. [Google Scholar]
- Guerreiro, I.; Serra, C.R.; Enes, P.; Couto, A.; Salvador, A.; Costas, B.; Oliva-Teles, A. Effect of short chain fructooligosaccharides (scFOS) on immunological status and gut microbiota of gilthead sea bream (Sparus aurata) reared at two temperatures. Fish Shellfish Immunol. 2016, 49, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Lp, A.; Yúfera, M.; Navarro-Guillén, C.; Moyano, F.J.; Soverini, M.; D’Amico, F.; Candela, M.; Fontanillas, R.; Gatta, P.P.; Bonaldo, A. Effects of calcium carbonate inclusion in low fishmeal diets on growth, gastrointestinal pH, digestive enzyme activity and gut bacterial community of European sea bass (Dicentrarchus labrax L.) juveniles. Aquaculture 2019, 510, 283–292. [Google Scholar]
- Li, P.; Gatlin, D.M. Dietary brewers yeast and the prebiotic Grobiotic™AE influence growth performance, immune responses and resistance of hybrid striped bass (Morone chrysops×M. saxatilis) to Streptococcus iniae infection. Aquaculture 2004, 231, 445–456. [Google Scholar] [CrossRef]
- Cui, L.; Xu, W.; Ai, Q.; Wang, D.; Mai, K. Effects of dietary chitosan oligosaccharide complex with rare earth on growth performance and innate immune response of turbot, Scophthalmus maximus L. Aquac. Res. 2013, 44, 683–690. [Google Scholar] [CrossRef]
- Lin, S.; Mao, S.; Yong, G.; Lin, L.; Li, L.; Yu, P. Effects of dietary chitosan oligosaccharides and Bacillus coagulans on the growth, innate immunity and resistance of koi (Cyprinus carpio koi). Aquaculture 2012, 342–343, 36–41. [Google Scholar] [CrossRef]
- Luo, L.; Cai, X.; He, C.; Xue, M.; Wu, X.; Cao, H. Immune response, stress resistance and bacterial challenge in juvenile rainbow trouts Oncorhynchus mykiss fed diets containing chitosan-oligosaccharides. Curr. Zool. 2009, 55, 416–422. [Google Scholar] [CrossRef]
- Olsen, R.E.; Myklebust, R.; Kryvi, H.; Mayhew, T.M.; Ringø, E. Damaging effect of dietary inulin on intestinal enterocytes in Arctic charr (Salvelinus alpinus L.). Aquac. Res. 2001, 32, 931–934. [Google Scholar] [CrossRef]
- Tacona, A.G.J.; Metianb, M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture 2008, 285, 146–158. [Google Scholar] [CrossRef]
- Hardy, R.W.; Tacon, A.G.J. Fish meal: Historical uses, production trends and future outlook for sustainable supplies. Responsible Mar. Aquac. 2002, 311–325. [Google Scholar]
- Hardy, R.W. Hardy, Utilization of plant proteins in fish diets: Effects of global demand and supplies of fishmeal. Aquac. Res. 2010, 41, 770–776. [Google Scholar] [CrossRef]
- Daniel, A. Studies, A review on replacing fish meal in aqua feeds using plant protein sources. Int. J. Fish. Aquat. Stud. 2018, 6, 164–179. [Google Scholar]
- Robinson, E.H.; Li, M.H. Use of Plant Proteins in Catfish Feeds: Replacement of Soybean Meal with Cottonseed Meal and Replacement of Fish Meal with Soybean Meal and Cottonseed Meal. J. World Aquac. Soc. 1994, 25, 271–276. [Google Scholar] [CrossRef]
- Li, Y.; Ai, Q.; Mai, K.; Xu, W.; Deng, J.; Cheng, Z. Comparison of high-protein soybean meal and commercial soybean meal partly replacing fish meal on the activities of digestive enzymes and aminotransferases in juvenile Japanese seabass, Lateolabrax japonicus (Cuvier, 1828). Aquac. Res. 2014, 45, 1051–1060. [Google Scholar] [CrossRef]
- Laporte, J.; Trushenski, J. Production performance, stress tolerance and intestinal integrity of sunshine bass fed increasing levels of soybean meal. J. Anim. Physiol. Anim. Nutr. 2012, 96, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Baeverfjord, G.T.; Krogdahl, A.J. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L., distal intestine: A comparison with the intestines of fasted fish. J. Fish Dis. 2010, 19, 375–387. [Google Scholar] [CrossRef]
- Theodosopoulos, A.N.; Hund, A.K.; Taylor, S.A. Parasites and Host Species Barriers in Animal Hybrid Zones. Trends Ecol. Evol. 2019, 34, 19–30. [Google Scholar] [CrossRef]
- Dittmer, J.; Bouchon, D. Feminizing Wolbachia influence microbiota composition in the terrestrial isopod Armadillidium vulgare. Sci. Rep. 2018, 8, 6998. [Google Scholar] [CrossRef]
- Zchori-Fein, E.; Faktor, O.; Zeidan, M.; Gottlieb, Y.; Czosnek, H.; Rosen, D. Parthenogenesis-inducing microorganisms in Aphytis (Hymenoptera: Aphelinidae). Insect Mol. Biol. 1995, 4, 173–178. [Google Scholar] [CrossRef]
- Nadal-Jimenez, P.; Griffin, J.S.; Davies, L.; Frost, C.L.; Marcello, M.; Hurst, G.D.D. Genetic manipulation allows in vivo tracking of the life cycle of the son-killer symbiont, Arsenophonus nasoniae, and reveals patterns of host invasion, tropism and pathology. Environ. Microbiol. 2019, 21, 3172–3182. [Google Scholar] [CrossRef] [Green Version]
- Perlmutter, J.I.; Bordenstein, S.R. Microorganisms in the reproductive tissues of arthropods. Nat. Rev. Microbiol. 2020, 18, 97–111. [Google Scholar] [CrossRef]
- Gebiola, M.; Kelly, S.E.; Hammerstein, P.; Giorgini, M.; Hunter, M.S. Darwin′s corollary and cytoplasmic incompatibility induced by Cardinium may contribute to speciation in Encarsia wasps (Hymenoptera: Aphelinidae). Evol. Int. J. Org. Evol. 2016, 70, 2447–2458. [Google Scholar] [CrossRef]
- Gatenby, J.B. Symbionticism and the Origin of Species. Nature 1928, 121, 164–165. [Google Scholar] [CrossRef]
- Miller, A.K.; Westlake, C.S.; Cross, K.L.; Leigh, B.A.; Bordenstein, S.R. The microbiome impacts host hybridization and speciation. PLoS Biol. 2021, 19, e3001417. [Google Scholar] [CrossRef] [PubMed]
- Brooks, A.W.; Kohl, K.D.; Brucker, R.M.; van Opstal, E.J.; Bordenstein, S.R. Phylosymbiosis: Relationships and Functional Effects of Microbial Communities across Host Evolutionary History. PLoS Biol. 2016, 14, e2000225. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, T.; Koga, R.; Fukatsu, T. Host plant specialization governed by facultative symbiont. Science 2004, 303, 1989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nardon, P.; Grenier, A.M. Serial endosymbiosis theory and weevil evolution: The role of symbiosis. In Symbiosis as a Source of Evolutionary Innovation: Speciation and Morphogenesis; Margulis, L., Fester, R., Eds.; MIT Press: Cambridage, MA, USA, 1991; pp. 153–169. [Google Scholar]
- Hosokawa, T.; Kikuchi, Y.; Nikoh, N.; Shimada, M.; Fukatsu, T. Strict host-symbiont cospeciation and reductive genome evolution in insect gut bacteria. PLoS Biol. 2006, 4, e337. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.J.; Ehrman, L.; Schneider, D. Infectious speciation revisited: Impact of symbiont-depletion on female fitness and mating behavior of Drosophila paulistorum. PLoS Pathog. 2010, 6, e1001214. [Google Scholar] [CrossRef] [Green Version]
- Breeuwer, J.A.; Werren, J.H. Microorganisms associated with chromosome destruction and reproductive isolation between two insect species. Nature 1990, 346, 558–560. [Google Scholar] [CrossRef]
- Bordenstein, S.R.; O′Hara, F.P.; Werren, J.H. Wolbachia-induced incompatibility precedes other hybrid incompatibilities in Nasonia. Nature 2001, 409, 707–710. [Google Scholar] [CrossRef]
- Brucker, R.M.; Bordenstein, S.R. The hologenomic basis of speciation: Gut bacteria cause hybrid lethality in the genus Nasonia. Science 2013, 341, 667–669. [Google Scholar] [CrossRef]
- Li, Z.; Wright, A.G.; Si, H.; Wang, X.; Qian, W.; Zhang, Z.; Li, G. Changes in the rumen microbiome and metabolites reveal the effect of host genetics on hybrid crosses. Environ. Microbiol. Rep. 2016, 8, 1016–1023. [Google Scholar] [CrossRef]
- Petri, R.M.; Schwaiger, T.; Penner, G.B.; Beauchemin, K.A.; Forster, R.J.; McKinnon, J.J.; McAllister, T.A. Characterization of the core rumen microbiome in cattle during transition from forage to concentrate as well as during and after an acidotic challenge. PLoS ONE 2013, 8, e83424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, J.E.; Schennink, A.; Burden, F.; Long, S.; van Doorn, D.A.; Pellikaan, W.F.; Dijkstra, J.; Saccenti, E.; Smidt, H. Domesticated equine species and their derived hybrids differ in their fecal microbiota. Anim. Microbiome 2020, 2, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Zhou, Z.; Li, H.; Wang, S.; Ren, L.; Hu, J.; Liu, Q.; Wu, C.; Tang, C.; Hu, F.; et al. Successional Changes of Microbial Communities and Host-Microbiota Interactions Contribute to Dietary Adaptation in Allodiploid Hybrid Fish. Microb. Ecol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Belkova, N.L.; Sidorova, T.V.; Glyzina, O.Y.; Yakchnenko, V.M.; Sapozhnikova, Y.P.; Bukin, Y.S.; Baturina, O.A.; Sukhanova, L.V. Gut microbiome of juvenile coregonid fishes: Comparison of sympatric species and their F1 hybrids. Fundam. Appl. Limnol. 2017, 189, 279–290. [Google Scholar] [CrossRef]
- Bahrndorff, S.; Alemu, T.; Alemneh, T.; Nielsen, J.L. The Microbiome of Animals: Implications for Conservation Biology. Int. J. Genom. 2016, 2016, 5304028. [Google Scholar] [CrossRef] [Green Version]
- Chown, S.L.; Hodgins, K.A.; Griffin, P.C.; Oakeshott, J.G.; Byrne, M.; Hoffmann, A.A. Biological invasions, climate change and genomics. Evol. Appl. 2015, 8, 23–46. [Google Scholar] [CrossRef]
- Simberloff, D.; RejmÁNek, M. Encyclopedia of Biological Invasions, 1st ed.; Simberloff, D., Rejmánek, M., Eds.; University of California Press: Oakland, CA, USA, 2011. [Google Scholar] [CrossRef]
- Lefort, M.C.; Boyer, S.; De Romans, S.; Glare, T.; Armstrong, K.; Worner, S. Invasion success of a scarab beetle within its native range: Host range expansion versus host-shift. PeerJ 2014, 2, e262. [Google Scholar] [CrossRef] [Green Version]
- Lefort, M.C.; Boyer, S.; Glare, T.R. A response to Pennisi—How do gut microbiomes help herbivores, a hint into next-generation biocontrol solutions. Rethink. Ecol. 2017, 1, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Thairu, M.W.; Hansen, A.K. Novel Insights into Insect-Microbe Interactions-Role of Epigenomics and Small RNAs. Front. Plant Sci. 2016, 7, 1164. [Google Scholar] [CrossRef] [Green Version]
- Dlugosch, K.M.; Parker, I.M. Founding events in species invasions: Genetic variation, adaptive evolution, and the role of multiple introductions. Mol. Ecol. 2008, 17, 431–449. [Google Scholar] [CrossRef]
- Clark, J.S.; Lewis, M.; Horvath, L. Invasion by extremes: Population spread with variation in dispersal and reproduction. Am. Nat. 2001, 157, 537–554. [Google Scholar] [CrossRef] [PubMed]
- Ficetola, G.F.; Siesa, M.E.; de Bernardi, F.; Padoa-Schioppa, E. Complex impact of an invasive crayfish on freshwater food webs. Biodivers. Conserv. 2012, 21, 2641–2651. [Google Scholar] [CrossRef]
- Figueroa, M.E.; Castillo, J.M.; Redondo, S.; Luque, T.; Castellanos, E.M.; Nieva, F.J.; Luque, C.J.; Rubio-Casal, A.E.; Davy, A.J. Facilitated invasion by hybridization of Sarcocornia species in a salt-marsh succession. J. Ecol. 2003, 91, 616–626. [Google Scholar] [CrossRef]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost microbiomes: The state of the art in their characterization, manipulation and importance in aquaculture and fisheries. Front. Microbiol. 2014, 5, 207. [Google Scholar] [CrossRef] [Green Version]
- Burns, A.R.; Stephens, W.Z.; Stagaman, K.; Wong, S.; Rawls, J.F.; Guillemin, K.; Bohannan, B.J. Contribution of neutral processes to the assembly of gut microbial communities in the zebrafish over host development. ISME J. 2016, 10, 655–664. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.C.; Snowberg, L.K.; Caporaso, J.G.; Knight, R.; Bolnick, D.I. Dietary input of microbes and host genetic variation shape among-population differences in stickleback gut microbiota. ISME J. 2015, 9, 2515–2526. [Google Scholar] [CrossRef]
- Sylvain, F.E.; Derome, N. Vertically and horizontally transmitted microbial symbionts shape the gut microbiota ontogenesis of a skin-mucus feeding discus fish progeny. Sci. Rep. 2017, 7, 5263. [Google Scholar] [CrossRef] [Green Version]
- Bates, J.M.; Mittge, E.; Kuhlman, J.; Baden, K.N.; Cheesman, S.E.; Guillemin, K. Distinct signals from the microbiota promote different aspects of zebrafish gut differentiation. Dev. Biol. 2006, 297, 374–386. [Google Scholar] [CrossRef]
- Yan, Q.; Li, J.; Yu, Y.; Wang, J.; He, Z.; Van Nostrand, J.D.; Kempher, M.L.; Wu, L.; Wang, Y.; Liao, L.; et al. Environmental filtering decreases with fish development for the assembly of gut microbiota. Environ. Microbiol. 2016, 18, 4739–4754. [Google Scholar] [CrossRef]
- Stephens, W.Z.; Burns, A.R.; Stagaman, K.; Wong, S.; Rawls, J.F.; Guillemin, K.; Bohannan, B.J. The composition of the zebrafish intestinal microbial community varies across development. ISME J. 2016, 10, 644–654. [Google Scholar] [CrossRef] [Green Version]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Roeselers, G.; Mittge, E.K.; Stephens, W.Z.; Parichy, D.M.; Cavanaugh, C.M.; Guillemin, K.; Rawls, J.F. Evidence for a core gut microbiota in the zebrafish. ISME J. 2011, 5, 1595–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullam, K.E.; Essinger, S.D.; Lozupone, C.A.; O’Connor, M.P.; Rosen, G.L.; Knight, R.; Kilham, S.S.; Russell, J.A. Environmental and ecological factors that shape the gut bacterial communities of fish: A meta-analysis. Mol. Ecol. 2012, 21, 3363–3378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Wang, G.; Angert, E.R.; Wang, W.; Li, W.; Zou, H. Composition, diversity, and origin of the bacterial community in grass carp intestine. PLoS ONE 2012, 7, e30440. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.G.; Tian, J.Y.; Gatesoupe, F.J.; Li, W.X.; Zou, H.; Yang, B.J.; Wang, G.T. Intestinal microbiota of gibel carp (Carassius auratus gibelio) and its origin as revealed by 454 pyrosequencing. World J. Microbiol. Biotechnol. 2013, 29, 1585–1595. [Google Scholar] [CrossRef]
- Wong, S.; Stephens, W.Z.; Burns, A.R.; Stagaman, K.; David, L.A.; Bohannan, B.J.; Guillemin, K.; Rawls, J.F. Ontogenetic Differences in Dietary Fat Influence Microbiota Assembly in the Zebrafish Gut. Mbio 2015, 6, e00687-15. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Amberg, J.; Chapman, D.; Gaikowski, M.; Liu, W.T. Fish gut microbiota analysis differentiates physiology and behavior of invasive Asian carp and indigenous American fish. ISME J. 2014, 8, 541–551. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Song, W.; Lin, H.; Wang, W.; Du, L.; Xing, W. Antibiotics and antibiotic resistance genes in global lakes: A review and meta-analysis. Environ. Int. 2018, 116, 60–73. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Species | Class | Order | Family | Ingredients | Intervention Type | 16s rRNA Sequencing | References |
|---|---|---|---|---|---|---|---|
| Oreochromis niloticus | Actinopteri | Cichliformes | Cichlidae | Rummeliibacillus stabekisii | Probiotic | Illumina MiSeq, Amplicon: V3–V4 | [51] |
| Oreochromis niloticus | Actinopteri | Cichliformes | Cichlidae | Bacillus subtilis | Probiotic | Illumina HiSeq, Amplicon: V4 | [52] |
| Acipenser baerii | Actinopteri | Acipenseriformes | Acipenseridae | Arabinoxylan-oligosaccharides (A.X.O.S.) + Lactococcus lactis spp. lactis or Bacillus circulans | Synbiotic | 454 GS FLX Titanium, Amplicon | [53] |
| Ctenopharyngodon idellus | Actinopteri | Cypriniformes | Xenocyprididae | Xylo-oligosaccharide | Prebiotic | Illumina MiSeq | [54] |
| Oreochromis niloticus | Actinopteri | Cichliformes | Cichlidae | Lactobacillus rhamnosus JCM1136 and Lactococcus lactis subsp. lactis JCM5805 | Probiotic | Illumina MiSeq, Amplicon: V3–V4 | [55] |
| Dicentrarchus labrax | Actinopteri | Perciformes | Moronidae | Calcium carbonate | Prebiotic | Illumina MiSeq, Amplicon: V3–V4 | [56] |
| Ctenopharyngodon idellus | Actinopteri | Cypriniformes | Xenocyprididae | Bacillus subtilis | Probiotic | No | [57] |
| Ctenopharyngodon idellus | Actinopteri | Cypriniformes | Xenocyprididae | Bacillus coagulans, Rhodopseudomonas palustris and Lactobacillus acidophilus | Probiotic | No | [58] |
| Ctenopharyngodon idellus | Actinopteri | Cypriniformes | Xenocyprididae | Bacillus subtilis Ch9 | Probiotic | No | [59] |
| Ctenopharyngodon idellus | Actinopteri | Cypriniformes | Xenocyprididae | Exogenous cellulase | Prebiotic | Amplicon: V3 | [60] |
| Ctenopharyngodon idellus | Actinopteri | Cypriniformes | Xenocyprididae | Glutathione | Prebiotic | No | [61] |
| Ctenopharyngodon idellus | Actinopteri | Cypriniformes | Xenocyprididae | B. licheniformis + xylo-oligosaccharide | Synbiotic | No | [62] |
| Danio rerio | Actinopteri | Cypriniformes | Danionidae | Gluten formulated diet | Protein | Illumina Miseq, Amplicon: V4 | [63] |
| Danio rerio | Actinopteri | Cypriniformes | Danionidae | Protein meal of animal origin (ragworm Nereis virens) | Protein | 454 GS FLX Titanium, Amplicon | [64] |
| Danio rerio | Actinopteri | Cypriniformes | Danionidae | Chitosan silver nanocomposites (CAgNCs) | Composites | 454 GS FLX Titanium, Amplicon | [65] |
| Gambusia affinis | Actinopteri | Cyprinodontiformes | Poeciliidae | Rifampicin | Antibiotic | Illumina HiSeq, Amplicon: V4 | [66] |
| Oncorhynchus mykiss | Actinopteri | Salmoniformes | Salmonidae | Dietary plant proteins | Protein | Illumina Miseq, Amplicon: V6–V8 | [67] |
| Oncorhynchus mykiss | Actinopteri | Salmoniformes | Salmonidae | Wickerhamomyces anomalus + Saccharomyces cerevisiae | Synbiotic | Illumina HiSeq, Amplicon | [68] |
| Oncorhynchus mykiss | Actinopteri | Salmoniformes | Salmonidae | Microalgae meal (Schizochytrium limacinum) | Prebiotic | Illumina HiSeq, Amplicon | [69] |
| Oreochromis niloticus | Actinopteri | Cichliformes | Cichlidae | Lactobacillus plantarum CCFM8610 | Probiotic | Illumina MiSeq, Amplicon: V4–V5 | [70] |
| Oreochromis niloticus | Actinopteri | Cichliformes | Cichlidae | Lactobacillus plantarum CCFM639 | Probiotic | Illumina MiSeq, Amplicon | [71] |
| Oreochromis niloticus | Actinopteri | Cichliformes | Cichlidae | Vibrio sp. CC8 and Bacillus cereus CC27, | Probiotic | No | [72] |
| Oreochromis niloticus | Actinopteri | Cichliformes | Cichlidae | Clostridium butyricum | Probiotic | Illumina HiSeq, Amplicon- | [73] |
| Oreochromis niloticus | Actinopteri | Cichliformes | Cichlidae | Allium sativum | Plant | Illumina MiSeq, Amplicon: V4–V5 | [74] |
| Oreochromis niloticus | Actinopteri | Cichliformes | Cichlidae | Bacillus subtilis and Bacillus licheniformis | Probiotic | No | [75] |
| Oreochromis niloticus | Actinopteri | Cichliformes | Cichlidae | Metschnikowia sp. GXUS03 | Probiotic | No | [76] |
| Sparus aurata | Actinopteri | Spariformes | Sparidae | Sodium butyrate | Butyrate | 455 GS FLX Titanium, Amplicon: V1–V3 | [77] |
| Seriola lalandi | Actinopteri | Carangiformes | Carangidae | Oxytetracycline, erythromycin and metronidazole | Antibiotic | Illumina MiSeq, Amplicon: V1–V2 | [78] |
| Piaractus mesopotamicus | Actinopteri | Characiformes | Serrasalmidae | Florfenicol | Antibiotic | Illumina MiSeq, Shotgun metagenome | [79] |
| Channa striata | Actinopteri | Anabantiformes | Channidae | β-glucan, galactooligosaccharides, mannan-oligosaccharide | Prebiotic | T-RFLP fragment sequencing, Amplicon | [80] |
| Channa striata | Actinopteri | Anabantiformes | Channidae | Saccharomyces cerevisiae and Lactobacillus acidophilus | Probiotic | T-RFLP fragment sequencing, Amplicon | [80] |
| Cyprinus carpio | Actinopteri | Cypriniformes | Cyprinidae | Chinese yam peel | Plant | Illumina MiSeq, Amplicon: V3–V4 | [81] |
| Lates calcarifer | Actinopteri | Perciformes | Centropomidae | Sodium diformate | Formate | No | [82] |
| Oreochromis niloticus | Actinopteri | Cichliformes | Cichlidae | Bacillus subtilis and Lactobacillus plantarum | Probiotic | ABI PRISM 377 sequencer (Perkin-Elmer), Amplicon: V6–V8 | [83] |
| Sparus aurata | Actinopteri | Spariformes | Sparidae | Poultry by-product meal and Hydrolyzed feather meal | Protein | 455 GS FLX Titanium, Amplicon: V3–V4 | [84] |
| Sparus aurata | Actinopteri | Spariformes | Sparidae | Fish protein hydrolysate or Autolysed dried yeast | Protein | Illumina MiSeq, Amplicon: V3–V4 | [85] |
| Dicentrarchus labrax | Actinopteri | Perciformes | Moronidae | Galactomannan oligosaccharides and A mixture of garlic and labiatae-plants oils | Prebiotic | Illumina MiSeq, Amplicon: V3–V4 | [86] |
| Salmo salar | Actinopteri | Salmoniformes | Salmonidae | Pediococcus acidilactici MA18/5M and Short chain fructooligosaccharides | Synbiotic | Amplicon: V3 | [87] |
| Arapaima gigas | Actinopteri | Osteoglossiformes | Osteoglossidae | Lactococcus lactis subsp. lactis and Enterococcus faecium | Probiotic | Amplicon: V1–V2 | [88] |
| Cyprinus carpio | Actinopteri | Cypriniformes | Cyprinidae | Dietary plant proteins | Protein | Illumina HiSeq, Amplicon: V3–V4 | [89] |
| Carassius auratus | Actinopteri | Cypriniformes | Cyprinidae | Bacillus subtilis and Enterococcus faecium | Probiotic | Amplicon: V3–V4 | [90] |
| Totoaba macdonaldi | Actinopteri | Perciformes | Sciaenidae | Commercial dietary prebiotic and probiotic | Synbiotic | Illumina MiSeq, Amplicon: V3–V4 | [91] |
| Totoaba macdonaldi | Actinopteri | Perciformes | Sciaenidae | Soy protein concentrate | Protein | Illumina MiSeq, Amplicon: V3–V4 | [92] |
| Host/Parents | Class | Order | Family | NGS Platform | Amplicon Sequencing | Reference |
|---|---|---|---|---|---|---|
| Culter alburnus ♀ × Megalobrama amblycephala ♂ | Actinopteri | Cypriniformes | Xenocyprididae | Illumina MiSeq | Amplicon: V3–V4 | [39] |
| Parachondrostoma toxostoma/Chondrostoma nasus | Actinopteri | Cypriniformes | Leuciscidae | Illumina MiSeq | Amplicon: V4 | [48] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Illumina NovaSeq | Amplicon: V3–V4 | [93] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Unknown | Unknown | [94] |
| Oreochromis niloticus ♀ × O. aureus ♂ | Actinopteri | Cichliformes | Cichlidae | Unknown | Amplicon: V3 | [95] |
| Acipenser baerii × A. schrenckii | Actinopteri | Acipenseriformes | Acipenseridae | Illumina HiSeq | Amplicon: V3–V4 | [96] |
| Pangasianodon gigas × Pangasianodon hypophthalmus | Actinopteri | Siluriformes | Pangasiidae | Unknown | Unknown | [97] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Illumina HiSeq | Amplicon: V3–V4 | [98] |
| Acipenser baeri Brandt ♀ × A. schrenckii Brandt ♂ | Actinopteri | Acipenseriformes | Acipenseridae | Illumina MiSeq | Amplicon: V3–V4 | [99] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Illumina HiSeq | Amplicon: V3–V4 | [100] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Lon GeneStudio S5™ | Amplicon: V4 | [101] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Illumina | Amplicon: V3–V4 | [102] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Illumina | Amplicon: V3–V4 | [103] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Illumina MiSeq | Amplicon: V3–V4 | [104] |
| Morone chrysops × M. saxatilis | Actinopteri | Perciformes | Moronidae | Illumina MiSeq | Amplicon: V1–V3 | [105] |
| Coregonus | Actinopteri | Salmoniformes | Salmonidae | Illumina MiSeq | Amplicon: V3–V4 | [106] |
| Hypophthalmichthys nobilis × H. molitr | Actinopteri | Cypriniformes | Xenocyprididae | Illumina MiSeq | Amplicon: V4 | [107] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Illumina MiSeq | Amplicon: V3–V4 | [108] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Unknown | Amplicon | [109] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Illumina MiSeq | Amplicon | [110] |
| Morone Chrysops × M. Saxatilis | Actinopteri | Perciformes | Moronidae | Illumina MiSeq | Amplicon: V1–V3 | [111] |
| Oreochromis niloticus ♀ × O. aureus ♂ | Actinopteri | Cichliformes | Cichlidae | 454 Sequencer F.L.X. | Amplicon: V6–V8 | [112] |
| Oreochromis niloticus ♀ × O. aureus ♂ | Actinopteri | Cichliformes | Cichlidae | Unknown | Amplicon: V4 | [113] |
| Tachysurus fulvidraco ♀ × Pseudobagrus vachellii ♂ | Actinopteri | Siluriformes | Bagridae | Illumina MiSeq | Amplicon | [114] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Illumina | Amplicon: V3–V4 | [115] |
| Acipenser baerii × A. schrenckii | Actinopteri | Acipenseriformes | Acipenseridae | Illumina HiSeq | Amplicon: V3–V4 | [116] |
| Epinephelus moara ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Illumina HiSeq | Amplicon: V3–V4 | [117] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Illumina MiSeq | Amplicon: V3–V4 | [118] |
| Acipenser baerii × A. schrenckii | Actinopteri | Acipenseriformes | Acipenseridae | Illumina MiSeq | Amplicon: V3–V4 | [119] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Illumina HiSeq | Amplicon: V3–V4 | [120] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Illumina HiSeq | Amplicon | [121] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Unknown | Amplicon | [122] |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | Actinopteri | Perciformes | Serranidae | Illumina HiSeq | Amplicon | [123] |
| Acipenser baerii × A. schrenckii | Actinopteri | Acipenseriformes | Acipenseridae | Illumina HiSeq | Amplicon: V3–V4 | [124] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, X.; Zhang, Q.; Zhang, Q.; Zhang, Y.; Chen, H.; Liu, G.; Zhu, L. Research Progress of the Gut Microbiome in Hybrid Fish. Microorganisms 2022, 10, 891. https://doi.org/10.3390/microorganisms10050891
Cui X, Zhang Q, Zhang Q, Zhang Y, Chen H, Liu G, Zhu L. Research Progress of the Gut Microbiome in Hybrid Fish. Microorganisms. 2022; 10(5):891. https://doi.org/10.3390/microorganisms10050891
Chicago/Turabian StyleCui, Xinyuan, Qinrong Zhang, Qunde Zhang, Yongyong Zhang, Hua Chen, Guoqi Liu, and Lifeng Zhu. 2022. "Research Progress of the Gut Microbiome in Hybrid Fish" Microorganisms 10, no. 5: 891. https://doi.org/10.3390/microorganisms10050891