
The Retrospective on Atypical Brucella Species Leads to Novel Definitions

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Genomics and Phylogeny
2.1. Classification Criteria for Brucella spp.
2.2. Novel Brucella spp. as Part of the Core Clade
2.3. Novel Brucella spp. as Ancient Ancestors
3. Metabolism
3.1. General Physiological Characteristics of Brucella spp.
3.2. Growth Properties of Novel versus Classical Brucella
3.3. Atypical Brucella spp. Are Characterized by a Diversified Metabolism and Expanded Nutrient Utilization When Compared to Classical Species
3.3.1. Intra- and Interspecies Variation in the Central Carbon Catabolism of Brucella
3.3.2. Macro- and Microdiversity Shape the Nutrient Utilization of Brucella
Microdiversity
Macrodiversity
4. Adaptation to Acid Stress
4.1. Responses to Extreme Acid Stress
4.1.1. The Urease System
4.1.2. The Amino Acid-Dependent Acid Resistance Systems
4.2. Responses to Intermediate Acid Stress
5. In Cellulo and In Vivo Models of Brucella Infection
5.1. General Aspects of Infection Models
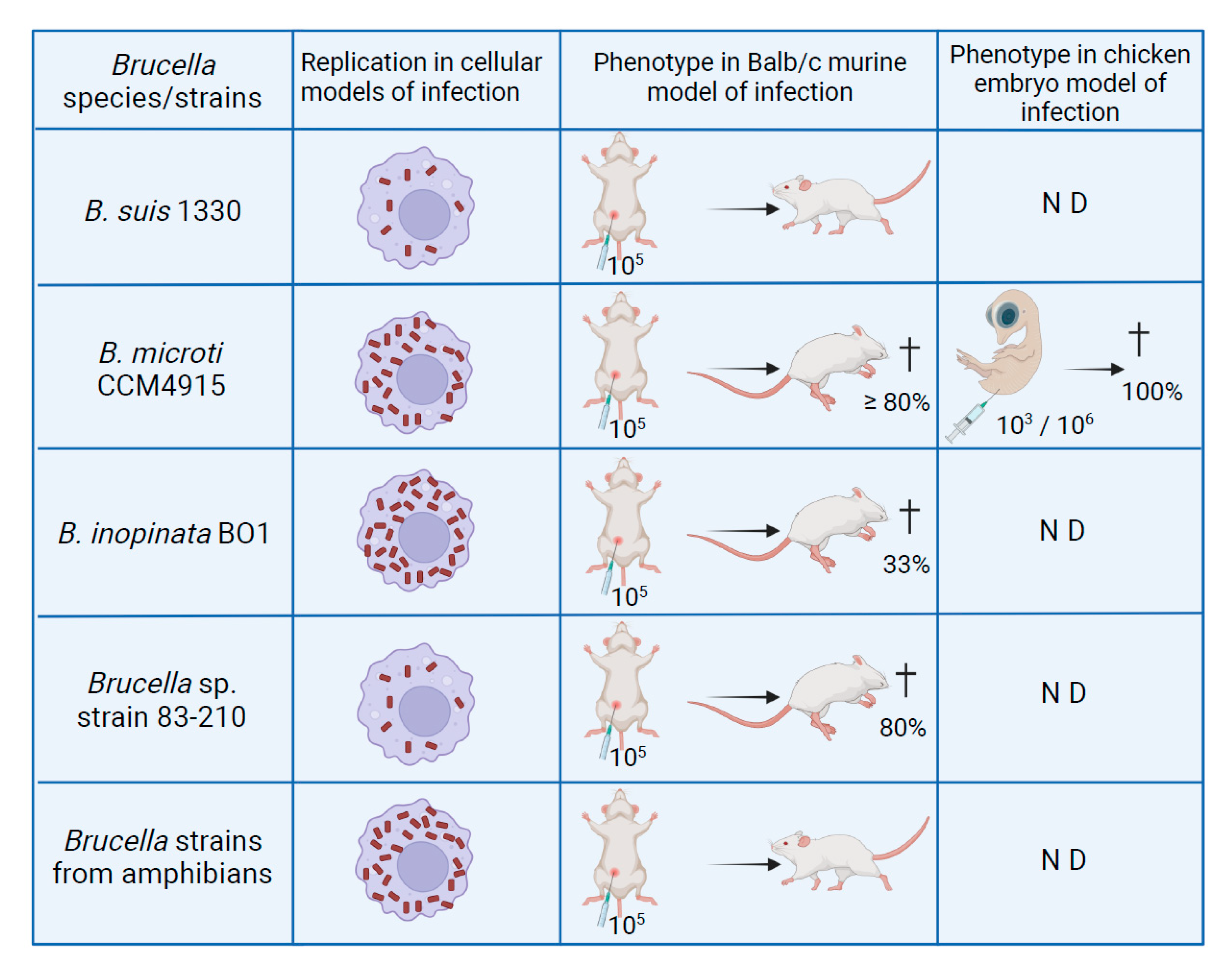
5.2. B. microti and B. inopinata BO1 in Macrophages and Murine Models of Infection
5.3. Brucella Frog Isolates in Established Infection Models
5.4. The Chicken Embryo Model
6. Conclusions Demand a Harmonized Classification of Brucella
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al Dahouk, S.; Neubauer, H.; Hensel, A.; Schöneberg, I.; Nöckler, K.; Alpers, K.; Merzenich, H.; Stark, K.; Jansen, A. Changing epidemiology of human brucellosis, Germany, 1962–2005. Emerg. Infect. Dis. 2007, 13, 1895–1900. [Google Scholar] [CrossRef] [PubMed]
- Traxler, R.M.; Lehman, M.W.; Bosserman, E.A.; Guerra, M.A.; Smith, T.L. A literature review of laboratory-acquired brucellosis. J. Clin. Microbiol. 2013, 51, 3055–3062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappas, G.; Papadimitriou, P.; Akritidis, N.; Christou, L.; Tsianos, E.V. The new global map of human brucellosis. Lancet Infect. Dis. 2006, 6, 91–99. [Google Scholar] [CrossRef]
- Hördt, A.; Lopez, M.G.; Meier-Kolthoff, J.P.; Schleuning, M.; Weinhold, L.M.; Tindall, B.J.; Gronow, S.; Kyrpides, N.C.; Woyke, T.; Göker, M. Analysis of 1000+ Type-Strain Genomes Substantially Improves Taxonomic Classification of Alphaproteobacteria. Front. Microbiol. 2020, 11, 468. [Google Scholar] [CrossRef]
- Jaÿ, M.; Girault, G.; Perrot, L.; Taunay, B.; Vuilmet, T.; Rossignol, F.; Pitel, P.H.; Picard, E.; Ponsart, C.; Mick, V. Phenotypic and Molecular Characterization of Brucella microti-Like Bacteria from a Domestic Marsh Frog (Pelophylax ridibundus). Front. Vet. Sci. 2018, 5, 283. [Google Scholar] [CrossRef]
- Santos, R.L.; Souza, T.D.; Mol, J.P.S.; Eckstein, C.; Paixao, T.A. Canine Brucellosis: An Update. Front. Vet. Sci. 2021, 8, 594291. [Google Scholar] [CrossRef]
- Suarez-Esquivel, M.; Ruiz-Villalobos, N.; Jiménez-Rojas, C.; Barquero-Calvo, E.; Chacon-Diaz, C.; Viquez-Ruiz, E.; Rojas-Campos, N.; Baker, K.S.; Oviedo-Sanchez, G.; Amuy, E.; et al. Brucella neotomae Infection in Humans, Costa Rica. Emerg. Infect. Dis. 2017, 23, 997–1000. [Google Scholar] [CrossRef] [Green Version]
- Foster, G.; Osterman, B.S.; Godfroid, J.; Jacques, I.; Cloeckaert, A. Brucella ceti sp. nov. and Brucella pinnipedialis sp. nov. for Brucella strains with cetaceans and seals as their preferred hosts. Int. J. Syst. Evol. Microbiol. 2007, 57, 2688–2693. [Google Scholar] [CrossRef] [Green Version]
- Whatmore, A.M.; Dawson, C.E.; Groussaud, P.; Koylass, M.S.; King, A.C.; Shankster, S.J.; Sohn, A.H.; Probert, W.S.; McDonald, W.L. Marine mammal Brucella genotype associated with zoonotic infection. Emerg. Infect. Dis. 2008, 14, 517–518. [Google Scholar] [CrossRef]
- Whatmore, A.M.; Foster, J.T. Emerging diversity and ongoing expansion of the genus Brucella. Infect. Genet. Evol. 2021, 92, 104865. [Google Scholar] [CrossRef]
- Alton, G.G.; Jones, L.M.; Angus, R.D.; Verger, J.M. Techniques for the Brucellosis Laboratory; Institut National de la Recherche Agronomique: Paris, France, 1988. [Google Scholar]
- Wattam, A.R.; Inzana, T.J.; Williams, K.P.; Mane, S.P.; Shukla, M.; Almeida, N.F.; Dickerman, A.W.; Mason, S.; Moriyon, I.; O’Callaghan, D.; et al. Comparative genomics of early-diverging Brucella strains reveals a novel lipopolysaccharide biosynthesis pathway. mBio 2012, 3, e00246-00212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zygmunt, M.S.; Jacques, I.; Bernardet, N.; Cloeckaert, A. Lipopolysaccharide heterogeneity in the atypical group of novel emerging Brucella species. Clin. Vaccine Immunol. 2012, 19, 1370–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Dahouk, S.; Köhler, S.; Occhialini, A.; Jimenez de Bagues, M.P.; Hammerl, J.A.; Eisenberg, T.; Vergnaud, G.; Cloeckaert, A.; Zygmunt, M.S.; Whatmore, A.M.; et al. Brucella spp. of amphibians comprise genomically diverse motile strains competent for replication in macrophages and survival in mammalian hosts. Sci. Rep. 2017, 7, 44420. [Google Scholar] [CrossRef] [PubMed]
- Scholz, H.C.; Hubalek, Z.; Sedlacek, I.; Vergnaud, G.; Tomaso, H.; Al Dahouk, S.; Melzer, F.; Kampfer, P.; Neubauer, H.; Cloeckaert, A.; et al. Brucella microti sp. nov., isolated from the common vole Microtus arvalis. Int. J. Syst. Evol. Microbiol. 2008, 58, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Hofer, E.; Revilla-Fernandez, S.; Al Dahouk, S.; Riehm, J.M.; Nöckler, K.; Zygmunt, M.S.; Cloeckaert, A.; Tomaso, H.; Scholz, H.C. A potential novel Brucella species isolated from mandibular lymph nodes of red foxes in Austria. Vet. Microbiol. 2012, 155, 93–99. [Google Scholar] [CrossRef]
- Whatmore, A.M.; Davison, N.; Cloeckaert, A.; Al Dahouk, S.; Zygmunt, M.S.; Brew, S.D.; Perrett, L.L.; Koylass, M.S.; Vergnaud, G.; Quance, C.; et al. Brucella papionis sp. nov., isolated from baboons (Papio spp.). Int. J. Syst. Evol. Microbiol. 2014, 64, 4120–4128. [Google Scholar] [CrossRef] [Green Version]
- de Bagues, M.P.J.; Ouahrani-Bettache, S.; Quintana, J.F.; Mitjana, O.; Hanna, N.; Bessoles, S.; Sanchez, F.; Scholz, H.C.; Lafont, V.; Köhler, S.; et al. The new species Brucella microti replicates in macrophages and causes death in murine models of infection. J. Infect. Dis. 2010, 202, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Al Dahouk, S.; Hofer, E.; Tomaso, H.; Vergnaud, G.; Le Flèche, P.; Cloeckaert, A.; Koylass, M.S.; Whatmore, A.M.; Nöckler, K.; Scholz, H.C. Intraspecies biodiversity of the genetically homologous species Brucella microti. Appl. Environ. Microbiol. 2012, 78, 1534–1543. [Google Scholar] [CrossRef] [Green Version]
- Hubalek, Z.; Scholz, H.C.; Sedlacek, I.; Melzer, F.; Sanogo, Y.O.; Nesvadbova, J. Brucellosis of the common vole (Microtus arvalis). Vector Borne Zoonotic Dis. 2007, 7, 679–687. [Google Scholar] [CrossRef]
- Scholz, H.C.; Nöckler, K.; Göllner, C.; Bahn, P.; Vergnaud, G.; Tomaso, H.; Al Dahouk, S.; Kämpfer, P.; Cloeckaert, A.; Maquart, M.; et al. Brucella inopinata sp. nov., isolated from a breast implant infection. Int. J. Syst. Evol. Microbiol. 2010, 60, 801–808. [Google Scholar] [CrossRef] [Green Version]
- Ronai, Z.; Kreizinger, Z.; Dan, A.; Drees, K.; Foster, J.T.; Banyai, K.; Marton, S.; Szeredi, L.; Janosi, S.; Gyuranecz, M. First isolation and characterization of Brucella microti from wild boar. BMC Vet. Res. 2015, 11, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mühldorfer, K.; Wibbelt, G.; Szentiks, C.A.; Fischer, D.; Scholz, H.C.; Zschock, M.; Eisenberg, T. The role of ‘atypical’ Brucella in amphibians: Are we facing novel emerging pathogens? J. Appl. Microbiol. 2017, 122, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.P.; Pembroke, J.T. The Genus Ochrobactrum as Major Opportunistic Pathogens. Microorganisms 2020, 8, 1797. [Google Scholar] [CrossRef] [PubMed]
- Scholz, H.C.; Hofer, E.; Vergnaud, G.; Le Flèche, P.; Whatmore, A.M.; Al Dahouk, S.; Pfeffer, M.; Krüger, M.; Cloeckaert, A.; Tomaso, H. Isolation of Brucella microti from mandibular lymph nodes of red foxes, Vulpes vulpes, in lower Austria. Vector Borne Zoonotic Dis. 2009, 9, 153–156. [Google Scholar] [CrossRef]
- De, B.K.; Stauffer, L.; Koylass, M.S.; Sharp, S.E.; Gee, J.E.; Helsel, L.O.; Steigerwalt, A.G.; Vega, R.; Clark, T.A.; Daneshvar, M.I.; et al. Novel Brucella strain (BO1) associated with a prosthetic breast implant infection. J. Clin. Microbiol. 2008, 46, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Scholz, H.C.; Hubalek, Z.; Nesvadbova, J.; Tomaso, H.; Vergnaud, G.; Le Flèche, P.; Whatmore, A.M.; Al Dahouk, S.; Krüger, M.; Lodri, C.; et al. Isolation of Brucella microti from soil. Emerg. Infect. Dis. 2008, 14, 1316–1317. [Google Scholar] [CrossRef]
- Whatmore, A.M. Current understanding of the genetic diversity of Brucella, an expanding genus of zoonotic pathogens. Infect. Genet. Evol. 2009, 9, 1168–1184. [Google Scholar] [CrossRef]
- Audic, S.; Lescot, M.; Claverie, J.M.; Scholz, H.C. Brucella microti: The genome sequence of an emerging pathogen. BMC Genomics 2009, 10, 352. [Google Scholar] [CrossRef] [Green Version]
- de la Garza-Garcia, J.A.; Ouahrani-Bettache, S.; Lyonnais, S.; Ornelas-Eusebio, E.; Freddi, L.; Al Dahouk, S.; Occhialini, A.; Köhler, S. Comparative Genome-Wide Transcriptome Analysis of Brucella suis and Brucella microti Under Acid Stress at pH 4.5: Cold Shock Protein CspA and Dps Are Associated with Acid Resistance of B. microti. Front. Microbiol. 2021, 12, 794535. [Google Scholar] [CrossRef]
- Tiller, R.V.; Gee, J.E.; Lonsway, D.R.; Gribble, S.; Bell, S.C.; Jennison, A.V.; Bates, J.; Coulter, C.; Hoffmaster, A.R.; De, B.K. Identification of an unusual Brucella strain (BO2) from a lung biopsy in a 52 year-old patient with chronic destructive pneumonia. BMC Microbiol. 2010, 10, 23. [Google Scholar] [CrossRef] [Green Version]
- Scholz, H.C.; Revilla-Fernandez, S.; Dahouk, S.A.; Hammerl, J.A.; Zygmunt, M.S.; Cloeckaert, A.; Koylass, M.; Whatmore, A.M.; Blom, J.; Vergnaud, G.; et al. Brucella vulpis sp. nov., isolated from mandibular lymph nodes of red foxes (Vulpes vulpes). Int. J. Syst. Evol. Microbiol. 2016, 66, 2090–2098. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, T.; Hamann, H.P.; Kaim, U.; Schlez, K.; Seeger, H.; Schauerte, N.; Melzer, F.; Tomaso, H.; Scholz, H.C.; Koylass, M.S.; et al. Isolation of potentially novel Brucella spp. from frogs. Appl. Environ. Microbiol. 2012, 78, 3753–3755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, D.; Lorenz, N.; Heuser, W.; Kämpfer, P.; Scholz, H.C.; Lierz, M. Abscesses associated with a Brucella inopinata-like bacterium in a big-eyed tree frog (Leptopelis vermiculatus). J. Zoo Wildl. Med. 2012, 43, 625–628. [Google Scholar] [CrossRef]
- Whatmore, A.M.; Dale, E.J.; Stubberfield, E.; Muchowski, J.; Koylass, M.; Dawson, C.; Gopaul, K.K.; Perrett, L.L.; Jones, M.; Lawrie, A. Isolation of Brucella from a White’s tree frog (Litoria caerulea). JMM Case Rep. 2015, 2, e000017. [Google Scholar] [CrossRef] [Green Version]
- Soler-Llorens, P.F.; Quance, C.R.; Lawhon, S.D.; Stuber, T.P.; Edwards, J.F.; Ficht, T.A.; Robbe-Austerman, S.; O’Callaghan, D.; Keriel, A. A Brucella spp. Isolate from a Pac-Man Frog (Ceratophrys ornata) Reveals Characteristics Departing from Classical Brucellae. Front. Cell Infect. Microbiol. 2016, 6, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M.; Une, Y.; Suzuki, M.; Park, E.S.; Imaoka, K.; Morikawa, S. Isolation of Brucella inopinata-Like Bacteria from White’s and Denny’s Tree Frogs. Vector Borne Zoonotic Dis. 2017, 17, 297–302. [Google Scholar] [CrossRef]
- Latheef, S.; Keyburn, A.; Broz, I.; Bagnara, A.; Bayley, C.; Frith, S.; Dobson, E.C. Atypical Brucella sp in captive Australian green tree frogs (Litoria caerulea): Clinical features, pathology, culture and molecular characterization. Aust Vet. J. 2020, 98, 216–221. [Google Scholar] [CrossRef]
- Scholz, H.C.; Mühldorfer, K.; Shilton, C.; Benedict, S.; Whatmore, A.M.; Blom, J.; Eisenberg, T. The Change of a Medically Important Genus: Worldwide Occurrence of Genetically Diverse Novel Brucella Species in Exotic Frogs. PLoS ONE 2016, 11, e0168872. [Google Scholar] [CrossRef] [Green Version]
- Helmick, K.E.; Garner, M.M.; Rhyan, J.; Bradway, D. Clinicopathologic Features of Infection with Novel Brucella Organisms in Captive Waxy Tree Frogs (Phyllomedusa sauvagii) and Colorado River Toads (Incilius alvarius). J. Zoo Wildl. Med. 2018, 49, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Glabman, R.A.; Thompson, K.A.; Mani, R.; Colburn, R.; Agnew, D.W. Atypical Brucella inopinata-Like Species in 2 Marine Toads. Emerg. Infect. Dis. 2021, 27, 1748–1750. [Google Scholar] [CrossRef]
- Eisenberg, T.; Schlez, K.; Fawzy, A.; Volker, I.; Hechinger, S.; Curic, M.; Schauerte, N.; Geiger, C.; Blom, J.; Scholz, H.C. Expanding the host range: Infection of a reptilian host (Furcifer pardalis) by an atypical Brucella strain. Antonie Van Leeuwenhoek 2020, 113, 1531–1537. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, T.; Risse, K.; Schauerte, N.; Geiger, C.; Blom, J.; Scholz, H.C. Isolation of a novel ‘atypical’ Brucella strain from a bluespotted ribbontail ray (Taeniura lymma). Antonie Van Leeuwenhoek 2017, 110, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Rouzic, N.; Desmier, L.; Cariou, M.E.; Gay, E.; Foster, J.T.; Williamson, C.H.D.; Schmitt, F.; Le Henaff, M.; Le Coz, A.; Lorleac’h, A.; et al. First Case of Brucellosis Caused by an Amphibian-type Brucella. Clin. Infect. Dis. 2021, 72, e404–e407. [Google Scholar] [CrossRef] [PubMed]
- Jaÿ, M.; Freddi, L.; Mick, V.; Durand, B.; Girault, G.; Perrot, L.; Taunay, B.; Vuilmet, T.; Azam, D.; Ponsart, C.; et al. Brucella microti-like prevalence in French farms producing frogs. Transbound. Emerg. Dis. 2020, 67, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Rajendhran, J. Genomic insights into Brucella. Infect. Genet. Evol. 2021, 87, 104635. [Google Scholar] [CrossRef] [PubMed]
- Halling, S.M.; Tatum, F.M.; Bricker, B.J. Sequence and characterization of an insertion sequence, IS711, from Brucella ovis. Gene 1993, 133, 123–127. [Google Scholar] [CrossRef]
- Ouahrani, S.; Michaux, S.; Sri Widada, J.; Bourg, G.; Tournebize, R.; Ramuz, M.; Liautard, J.P. Identification and sequence analysis of IS6501, an insertion sequence in Brucella spp.: Relationship between genomic structure and the number of IS6501 copies. J. Gen. Microbiol. 1993, 139, 3265–3273. [Google Scholar] [CrossRef] [Green Version]
- Bricker, B.J.; Ewalt, D.R.; MacMillan, A.P.; Foster, G.; Brew, S. Molecular characterization of Brucella strains isolated from marine mammals. J. Clin. Microbiol. 2000, 38, 1258–1262. [Google Scholar] [CrossRef] [Green Version]
- Dawson, C.E.; Stubberfield, E.J.; Perrett, L.L.; King, A.C.; Whatmore, A.M.; Bashiruddin, J.B.; Stack, J.A.; Macmillan, A.P. Phenotypic and molecular characterisation of Brucella isolates from marine mammals. BMC Microbiol. 2008, 8, 224. [Google Scholar] [CrossRef] [Green Version]
- Bricker, B.J.; Halling, S.M. Differentiation of Brucella abortus bv. 1, 2, and 4, Brucella melitensis, Brucella ovis, and Brucella suis bv. 1 by PCR. J. Clin. Microbiol. 1994, 32, 2660–2666. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Yoldi, D.; Marin, C.M.; de Miguel, M.J.; Munoz, P.M.; Vizmanos, J.L.; Lopez-Goni, I. Multiplex PCR assay for the identification and differentiation of all Brucella species and the vaccine strains Brucella abortus S19 and RB51 and Brucella melitensis Rev1. Clin. Chem. 2006, 52, 779–781. [Google Scholar] [CrossRef] [PubMed]
- Wayne, L.G.; Brenner, D.J.; Colwell, R.R.; Grimont, P.A.D.; Kandler, O.; Krichevsky, M.I.; Moore, L.H.; Moore, W.E.C.; Murray, R.G.E.; Stackebrandt, E.; et al. Report of the Ad-Hoc-Committee on Reconciliation of Approaches to Bacterial Systematics. Int. J. Syst. Bacteriol. 1987, 37, 463–464. [Google Scholar] [CrossRef] [Green Version]
- Verger, J.M.; Grimont, F.; Grimont, P.A.D.; Grayon, M. Brucella, a Monospecific Genus as Shown by Deoxyribonucleic-Acid Hybridization. Int. J. Syst. Bacteriol. 1985, 35, 292–295. [Google Scholar] [CrossRef]
- Richter, M.; Rossello-Mora, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Gee, J.E.; De, B.K.; Levett, P.N.; Whitney, A.M.; Novak, R.T.; Popovic, T. Use of 16S rRNA gene sequencing for rapid confirmatory identification of Brucella isolates. J. Clin. Microbiol. 2004, 42, 3649–3654. [Google Scholar] [CrossRef] [Green Version]
- Zheludkov, M.M.; Tsirelson, L.E. Reservoirs of Brucella infection in nature. Biol. Bull. 2010, 37, 709–715. [Google Scholar] [CrossRef]
- Mayer-Scholl, A.; Draeger, A.; Göllner, C.; Scholz, H.C.; Nöckler, K. Advancement of a multiplex PCR for the differentiation of all currently described Brucella species. J. Microbiol. Methods 2010, 80, 112–114. [Google Scholar] [CrossRef]
- Paquet, J.Y.; Diaz, M.A.; Genevrois, S.; Grayon, M.; Verger, J.M.; de Bolle, X.; Lakey, J.H.; Letesson, J.J.; Cloeckaert, A. Molecular, antigenic, and functional analyses of Omp2b porin size variants of Brucella spp. J. Bacteriol. 2001, 183, 4839–4847. [Google Scholar] [CrossRef] [Green Version]
- Cook, I.; Campbell, R.W.; Barrow, G. Brucellosis in North Queensland rodents. Aust Vet. J. 1966, 42, 5–8. [Google Scholar] [CrossRef]
- Tiller, R.V.; Gee, J.E.; Frace, M.A.; Taylor, T.K.; Setubal, J.C.; Hoffmaster, A.R.; De, B.K. Characterization of novel Brucella strains originating from wild native rodent species in North Queensland, Australia. Appl. Environ. Microbiol. 2010, 76, 5837–5845. [Google Scholar] [CrossRef] [Green Version]
- Rodionova, I.A.; Li, X.; Thiel, V.; Stolyar, S.; Stanton, K.; Fredrickson, J.K.; Bryant, D.A.; Osterman, A.L.; Best, A.A.; Rodionov, D.A. Comparative genomics and functional analysis of rhamnose catabolic pathways and regulons in bacteria. Front. Microbiol. 2013, 4, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.A.; Sha, Z.; Mayfield, J.E. Regulation of Brucella abortus catalase. Infect. Immun. 2000, 68, 3861–3866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, K.H.; Baumgartner, J.E.; Valderas, M.W.; Roop, R.M., 2nd. Comparative study of the roles of AhpC and KatE as respiratory antioxidants in Brucella abortus 2308. J. Bacteriol. 2010, 192, 4912–4922. [Google Scholar] [CrossRef] [Green Version]
- Mironov, A.; Seregina, T.; Nagornykh, M.; Luhachack, L.G.; Korolkova, N.; Lopes, L.E.; Kotova, V.; Zavilgelsky, G.; Shakulov, R.; Shatalin, K.; et al. Mechanism of H2S-mediated protection against oxidative stress in Escherichia coli. Proc. Natl. Acad. Sci. USA 2017, 114, 6022–6027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, V.K.; Bandyopadhyay, P.; Singh, A. Hydrogen sulfide in physiology and pathogenesis of bacteria and viruses. IUBMB Life 2018, 70, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Varesio, L.M.; Fiebig, A.; Crosson, S. Brucella ovis Cysteine Biosynthesis Contributes to Peroxide Stress Survival and Fitness in the Intracellular Niche. Infect. Immun. 2021, 89, 6. [Google Scholar] [CrossRef] [PubMed]
- Carbonero, F.; Benefiel, A.C.; Alizadeh-Ghamsari, A.H.; Gaskins, H.R. Microbial pathways in colonic sulfur metabolism and links with health and disease. Front. Physiol 2012, 3, 448. [Google Scholar] [CrossRef] [Green Version]
- Szabo, C. A timeline of hydrogen sulfide (H2S) research: From environmental toxin to biological mediator. BioChem. Pharmacol 2018, 149, 5–19. [Google Scholar] [CrossRef]
- Walsh, B.J.C.; Giedroc, D.P. H2S and reactive sulfur signaling at the host-bacterial pathogen interface. J. Biol. Chem. 2020, 295, 13150–13168. [Google Scholar] [CrossRef]
- Vazquez-Torres, A.; Baumler, A.J. Nitrate, nitrite and nitric oxide reductases: From the last universal common ancestor to modern bacterial pathogens. Curr. Opin. Microbiol. 2016, 29, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Al Dahouk, S.; Scholz, H.C.; Tomaso, H.; Bahn, P.; Göllner, C.; Karges, W.; Appel, B.; Hensel, A.; Neubauer, H.; Nöckler, K. Differential phenotyping of Brucella species using a newly developed semi-automated metabolic system. BMC Microbiol. 2010, 10, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.E.; Cameron, H.S. Metabolic characterization of the genus Brucella. II. Oxidative metabolic patterns of the described biotypes. J. Bacteriol. 1961, 82, 396–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, P.; Lahiri, A.; Lahiri, A.; Chakravortty, D. Modulation of the arginase pathway in the context of microbial pathogenesis: A metabolic enzyme moonlighting as an immune modulator. PLoS Pathog. 2010, 6, e1000899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haine, V.; Dozot, M.; Dornand, J.; Letesson, J.J.; De Bolle, X. NnrA is required for full virulence and regulates several Brucella melitensis denitrification genes. J. Bacteriol. 2006, 188, 1615–1619. [Google Scholar] [CrossRef] [Green Version]
- Loisel-Meyer, S.; Jimenez de Bagues, M.P.; Basseres, E.; Dornand, J.; Kohler, S.; Liautard, J.P.; Jubier-Maurin, V. Requirement of norD for Brucella suis virulence in a murine model of in vitro and in vivo infection. Infect. Immun. 2006, 74, 1973–1976. [Google Scholar] [CrossRef] [Green Version]
- Philippot, L. Denitrification in pathogenic bacteria: For better or worst? Trends Microbiol. 2005, 13, 191–192. [Google Scholar] [CrossRef]
- Baek, S.H.; Rajashekara, G.; Splitter, G.A.; Shapleigh, J.P. Denitrification genes regulate Brucella virulence in mice. J. Bacteriol. 2004, 186, 6025–6031. [Google Scholar] [CrossRef] [Green Version]
- Breitschwerdt, E.B.; Kordick, D.L. Bartonella infection in animals: Carriership, reservoir potential, pathogenicity, and zoonotic potential for human infection. Clin. Microbiol. Rev. 2000, 13, 428–438. [Google Scholar] [CrossRef]
- Ronneau, S.; Moussa, S.; Barbier, T.; Conde-Alvarez, R.; Zuniga-Ripa, A.; Moriyon, I.; Letesson, J.J. Brucella, nitrogen and virulence. Crit Rev. Microbiol. 2016, 42, 507–525. [Google Scholar] [CrossRef]
- Cameron, H.S.; Holm, L.W.; Meyer, M. Comparative metabolic studies on the genus Brucella. I. Evidence of a urea cycle from glutamic acid metabolism. J. Bacteriol. 1952, 64, 709–712. [Google Scholar] [CrossRef] [Green Version]
- Doern, G.V. Detection of selected fastidious bacteria. Clin. Infect. Dis. 2000, 30, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, P. The nutrition of Brucellae. Bacteriol. Rev. 1958, 22, 81–98. [Google Scholar] [CrossRef] [PubMed]
- Plommet, M. Minimal requirements for growth of Brucella suis and other Brucella species. Zent. Bakteriol. 1991, 275, 436–450. [Google Scholar] [CrossRef]
- Zuniga-Ripa, A.; Barbier, T.; Lazaro-Anton, L.; de Miguel, M.J.; Conde-Alvarez, R.; Munoz, P.M.; Letesson, J.J.; Iriarte, M.; Moriyon, I. The Fast-Growing Brucella suis Biovar 5 Depends on Phosphoenolpyruvate Carboxykinase and Pyruvate Phosphate Dikinase but Not on Fbp and GlpX Fructose-1,6-Bisphosphatases or Isocitrate Lyase for Full Virulence in Laboratory Models. Front. Microbiol. 2018, 9, 641. [Google Scholar] [CrossRef] [PubMed]
- Gopalsamy, S.N.; Ramakrishnan, A.; Shariff, M.M.; Gabel, J.; Brennan, S.; Drenzek, C.; Farley, M.M.; Gaynes, R.P.; Cartwright, E.J. Brucellosis Initially Misidentified as Ochrobactrum anthropi Bacteremia: A Case Report and Review of the Literature. Open Forum Infect. Dis. 2021, 8, ofab473. [Google Scholar] [CrossRef]
- Corbel, M.J.; Brinley Morgan, W.J. Classification of the genus Brucella: The current position. Rev. Sci. Tech. 1982, 1, 281–310. [Google Scholar] [CrossRef]
- Jimenez de Bagues, M.P.; Iturralde, M.; Arias, M.A.; Pardo, J.; Cloeckaert, A.; Zygmunt, M.S. The new strains Brucella inopinata BO1 and Brucella species 83-210 behave biologically like classic infectious Brucella species and cause death in murine models of infection. J. Infect. Dis. 2014, 210, 467–472. [Google Scholar] [CrossRef] [Green Version]
- Suarez-Esquivel, M.; Chaves-Olarte, E.; Moreno, E.; Guzman-Verri, C. Brucella Genomics: Macro and Micro Evolution. Int. J. Mol. Sci. 2020, 21, 7749. [Google Scholar] [CrossRef]
- Colin, R.; Ni, B.; Laganenka, L.; Sourjik, V. Multiple functions of flagellar motility and chemotaxis in bacterial physiology. FEMS Microbiol. Rev. 2021, 45, fuab038. [Google Scholar] [CrossRef]
- O’Callaghan, D.; Whatmore, A.M. Brucella genomics as we enter the multi-genome era. Brief. Funct. Genom. 2011, 10, 334–341. [Google Scholar] [CrossRef] [Green Version]
- Moreno, E.; Cloeckaert, A.; Moriyon, I. Brucella evolution and taxonomy. Vet. Microbiol. 2002, 90, 209–227. [Google Scholar] [CrossRef]
- Chain, P.S.; Comerci, D.J.; Tolmasky, M.E.; Larimer, F.W.; Malfatti, S.A.; Vergez, L.M.; Aguero, F.; Land, M.L.; Ugalde, R.A.; Garcia, E. Whole-genome analyses of speciation events in pathogenic Brucellae. Infect. Immun. 2005, 73, 8353–8361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajashekara, G.; Glasner, J.D.; Glover, D.A.; Splitter, G.A. Comparative whole-genome hybridization reveals genomic islands in Brucella species. J. Bacteriol. 2004, 186, 5040–5051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsolis, R.M.; Seshadri, R.; Santos, R.L.; Sangari, F.J.; Lobo, J.M.; de Jong, M.F.; Ren, Q.; Myers, G.; Brinkac, L.M.; Nelson, W.C.; et al. Genome degradation in Brucella ovis corresponds with narrowing of its host range and tissue tropism. PLoS ONE 2009, 4, e5519. [Google Scholar] [CrossRef] [PubMed]
- Wattam, A.R.; Foster, J.T.; Mane, S.P.; Beckstrom-Sternberg, S.M.; Beckstrom-Sternberg, J.M.; Dickerman, A.W.; Keim, P.; Pearson, T.; Shukla, M.; Ward, D.V.; et al. Comparative phylogenomics and evolution of the Brucellae reveal a path to virulence. J. Bacteriol. 2014, 196, 920–930. [Google Scholar] [CrossRef] [Green Version]
- Wattam, A.R.; Williams, K.P.; Snyder, E.E.; Almeida, N.F., Jr.; Shukla, M.; Dickerman, A.W.; Crasta, O.R.; Kenyon, R.; Lu, J.; Shallom, J.M.; et al. Analysis of ten Brucella genomes reveals evidence for horizontal gene transfer despite a preferred intracellular lifestyle. J. Bacteriol. 2009, 191, 3569–3579. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Wang, Y.; Xu, J.; Chen, Y.; Ke, Y.; Zhou, X.; Yuan, X.; Zhou, D.; Yang, Y.; Yang, R.; et al. Parallel gene loss and acquisition among strains of different Brucella species and biovars. J. Microbiol. 2012, 50, 567–574. [Google Scholar] [CrossRef]
- Sheppard, S.K.; Didelot, X.; Meric, G.; Torralbo, A.; Jolley, K.A.; Kelly, D.J.; Bentley, S.D.; Maiden, M.C.; Parkhill, J.; Falush, D. Genome-wide association study identifies vitamin B5 biosynthesis as a host specificity factor in Campylobacter. Proc. Natl. Acad. Sci. USA 2013, 110, 11923–11927. [Google Scholar] [CrossRef] [Green Version]
- Barbier, T.; Zuniga-Ripa, A.; Moussa, S.; Plovier, H.; Sternon, J.F.; Lazaro-Anton, L.; Conde-Alvarez, R.; De Bolle, X.; Iriarte, M.; Moriyon, I.; et al. Brucella central carbon metabolism: An update. Crit. Rev. Microbiol. 2018, 44, 182–211. [Google Scholar] [CrossRef]
- Barbier, T.; Collard, F.; Zuniga-Ripa, A.; Moriyon, I.; Godard, T.; Becker, J.; Wittmann, C.; Van Schaftingen, E.; Letesson, J.J. Erythritol feeds the pentose phosphate pathway via three new isomerases leading to D-erythrose-4-phosphate in Brucella. Proc. Natl. Acad. Sci. USA 2014, 111, 17815–17820. [Google Scholar] [CrossRef] [Green Version]
- Essenberg, R.C.; Seshadri, R.; Nelson, K.; Paulsen, I. Sugar metabolism by Brucellae. Vet. Microbiol. 2002, 90, 249–261. [Google Scholar] [CrossRef]
- Spaans, S.K.; Weusthuis, R.A.; van der Oost, J.; Kengen, S.W. NADPH-generating systems in bacteria and archaea. Front. Microbiol. 2015, 6, 742. [Google Scholar] [CrossRef] [PubMed]
- Rytter, H.; Jamet, A.; Ziveri, J.; Ramond, E.; Coureuil, M.; Lagouge-Roussey, P.; Euphrasie, D.; Tros, F.; Goudin, N.; Chhuon, C.; et al. The pentose phosphate pathway constitutes a major metabolic hub in pathogenic Francisella. PLoS Pathog. 2021, 17, e1009326. [Google Scholar] [CrossRef] [PubMed]
- Marais, A.; Mendz, G.L.; Hazell, S.L.; Megraud, F. Metabolism and genetics of Helicobacter pylori: The genome era. Microbiol. Mol. Biol. Rev. 1999, 63, 642–674. [Google Scholar] [CrossRef] [Green Version]
- Hofreuter, D. Defining the metabolic requirements for the growth and colonization capacity of Campylobacter jejuni. Front. Cell Infect. Microbiol. 2014, 4, 137. [Google Scholar] [CrossRef] [Green Version]
- Tettelin, H.; Saunders, N.J.; Heidelberg, J.; Jeffries, A.C.; Nelson, K.E.; Eisen, J.A.; Ketchum, K.A.; Hood, D.W.; Peden, J.F.; Dodson, R.J.; et al. Complete genome sequence of Neisseria meningitidis serogroup B strain MC58. Science 2000, 287, 1809–1815. [Google Scholar] [CrossRef] [Green Version]
- Canback, B.; Andersson, S.G.; Kurland, C.G. The global phylogeny of glycolytic enzymes. Proc. Natl. Acad. Sci. USA 2002, 99, 6097–6102. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.S.; Chong, H.; Park, H.S.; Yoon, K.O.; Jung, C.; Kim, J.J.; Hong, J.H.; Kim, H.; Kim, J.H.; Kil, J.I.; et al. The genome sequence of the ethanologenic bacterium Zymomonas mobilis ZM4. Nat. Biotechnol. 2005, 23, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Barbier, T.; Nicolas, C.; Letesson, J.J. Brucella adaptation and survival at the crossroad of metabolism and virulence. FEBS Lett. 2011, 585, 2929–2934. [Google Scholar] [CrossRef] [Green Version]
- Brissac, T.; Ziveri, J.; Ramond, E.; Tros, F.; Kock, S.; Dupuis, M.; Brillet, M.; Barel, M.; Peyriga, L.; Cahoreau, E.; et al. Gluconeogenesis, an essential metabolic pathway for pathogenic Francisella. Mol. Microbiol. 2015, 98, 518–534. [Google Scholar] [CrossRef] [Green Version]
- Blume, M.; Nitzsche, R.; Sternberg, U.; Gerlic, M.; Masters, S.L.; Gupta, N.; McConville, M.J. A Toxoplasma gondii Gluconeogenic Enzyme Contributes to Robust Central Carbon Metabolism and Is Essential for Replication and Virulence. Cell Host Microbe. 2015, 18, 210–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machelart, A.; Willemart, K.; Zuniga-Ripa, A.; Godard, T.; Plovier, H.; Wittmann, C.; Moriyon, I.; De Bolle, X.; Van Schaftingen, E.; Letesson, J.J.; et al. Convergent evolution of zoonotic Brucella species toward the selective use of the pentose phosphate pathway. Proc. Natl. Acad. Sci. USA 2020, 117, 26374–26381. [Google Scholar] [CrossRef] [PubMed]
- Lazaro-Anton, L.; de Miguel, M.J.; Barbier, T.; Conde-Alvarez, R.; Munoz, P.M.; Letesson, J.J.; Iriarte, M.; Moriyon, I.; Zuniga-Ripa, A. Glucose Oxidation to Pyruvate Is Not Essential for Brucella suis Biovar 5 Virulence in the Mouse Model. Front. Microbiol. 2020, 11, 620049. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.E.; Morgan, W.J. Metabolic characterization of Brucella strains that show conflicting identity by biochemical and serological methods. Bull. World Health Organ. 1962, 26, 823–827. [Google Scholar]
- Varesio, L.M.; Willett, J.W.; Fiebig, A.; Crosson, S. A Carbonic Anhydrase Pseudogene Sensitizes Select Brucella Lineages to Low CO2 Tension. J. Bacteriol. 2019, 201, 22. [Google Scholar] [CrossRef] [Green Version]
- Finan, T.M.; Weidner, S.; Wong, K.; Buhrmester, J.; Chain, P.; Vorholter, F.J.; Hernandez-Lucas, I.; Becker, A.; Cowie, A.; Gouzy, J.; et al. The complete sequence of the 1683-kb pSymB megaplasmid from the N2-fixing endosymbiont Sinorhizobium meliloti. Proc. Natl. Acad. Sci. USA 2001, 98, 9889–9894. [Google Scholar] [CrossRef] [Green Version]
- Hermann, L.; Mais, C.N.; Czech, L.; Smits, S.H.J.; Bange, G.; Bremer, E. The ups and downs of ectoine: Structural enzymology of a major microbial stress protectant and versatile nutrient. Biol. Chem. 2020, 401, 1443–1468. [Google Scholar] [CrossRef]
- Mais, C.N.; Hermann, L.; Altegoer, F.; Seubert, A.; Richter, A.A.; Wernersbach, I.; Czech, L.; Bremer, E.; Bange, G. Degradation of the microbial stress protectants and chemical chaperones ectoine and hydroxyectoine by a bacterial hydrolase-deacetylase complex. J. Biol. Chem. 2020, 295, 9087–9104. [Google Scholar] [CrossRef]
- Thuoc, D.V.; Hien, T.T.; Sudesh, K. Identification and characterization of ectoine-producing bacteria isolated from Can Gio mangrove soil in Vietnam. Ann. Microbiol. 2019, 69, 819–828. [Google Scholar] [CrossRef]
- Decamp, A.; Michelo, O.; Rabbat, C.; Laroche, C.; Grizeau, D.; Pruvost, J.; Goncalves, O. A New, Quick, and Simple Protocol to Evaluate Microalgae Polysaccharide Composition. Mar. Drugs 2021, 19, 101. [Google Scholar] [CrossRef]
- Flamholz, A.; Noor, E.; Bar-Even, A.; Liebermeister, W.; Milo, R. Glycolytic strategy as a tradeoff between energy yield and protein cost. Proc. Natl. Acad. Sci. USA 2013, 110, 10039–10044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lund, P.; Tramonti, A.; De Biase, D. Coping with low pH: Molecular strategies in neutralophilic bacteria. FEMS Microbiol. Rev. 2014, 38, 1091–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangari, F.J.; Cayon, A.M.; Seoane, A.; Garcia-Lobo, J.M. Brucella abortus ure2 region contains an acid-activated urea transporter and a nickel transport system. BMC Microbiol. 2010, 10, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangari, F.J.; Seoane, A.; Rodriguez, M.C.; Aguero, J.; Garcia Lobo, J.M. Characterization of the urease operon of Brucella abortus and assessment of its role in virulence of the bacterium. Infect. Immun. 2007, 75, 774–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandara, A.B.; Contreras, A.; Contreras-Rodriguez, A.; Martins, A.M.; Dobrean, V.; Poff-Reichow, S.; Rajasekaran, P.; Sriranganathan, N.; Schurig, G.G.; Boyle, S.M. Brucella suis urease encoded by ure1 but not ure2 is necessary for intestinal infection of BALB/c mice. BMC Microbiol. 2007, 7, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorvel, J.P.; Moreno, E.; Moriyon, I. Is Brucella an enteric pathogen? Nature Rev. Microbiol. 2009, 7, 250. [Google Scholar] [CrossRef]
- Pennacchietti, E.; D’Alonzo, C.; Freddi, L.; Occhialini, A.; De Biase, D. The Glutaminase-Dependent Acid Resistance System: Qualitative and Quantitative Assays and Analysis of Its Distribution in Enteric Bacteria. Front. Microbiol. 2018, 9, 2869. [Google Scholar] [CrossRef] [Green Version]
- Freddi, L.; Damiano, M.A.; Chaloin, L.; Pennacchietti, E.; Al Dahouk, S.; Köhler, S.; De Biase, D.; Occhialini, A. The Glutaminase-Dependent System Confers Extreme Acid Resistance to New Species and Atypical Strains of Brucella. Front. Microbiol. 2017, 8, 2236. [Google Scholar] [CrossRef] [Green Version]
- Valderas, M.W.; Alcantara, R.B.; Baumgartner, J.E.; Bellaire, B.H.; Robertson, G.T.; Ng, W.L.; Richardson, J.M.; Winkler, M.E.; Roop, R.M., 2nd. Role of HdeA in acid resistance and virulence in Brucella abortus 2308. Vet. Microbiol. 2005, 107, 307–312. [Google Scholar] [CrossRef]
- De Biase, D.; Tramonti, A.; Bossa, F.; Visca, P. The response to stationary-phase stress conditions in Escherichia coli: Role and regulation of the glutamic acid decarboxylase system. Mol. Microbiol. 1999, 32, 1198–1211. [Google Scholar] [CrossRef] [Green Version]
- Castanie-Cornet, M.P.; Penfound, T.A.; Smith, D.; Elliott, J.F.; Foster, J.W. Control of acid resistance in Escherichia coli. J. Bacteriol. 1999, 181, 3525–3535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotter, P.D.; Gahan, C.G.; Hill, C. A glutamate decarboxylase system protects Listeria monocytogenes in gastric fluid. Mol. Microbiol. 2001, 40, 465–475. [Google Scholar] [CrossRef] [PubMed]
- De Biase, D.; Pennacchietti, E. Glutamate decarboxylase-dependent acid resistance in orally acquired bacteria: Function, distribution and biomedical implications of the gadBC operon. Mol. Microbiol. 2012, 86, 770–786. [Google Scholar] [CrossRef] [PubMed]
- De Biase, D.; Tramonti, A.; John, R.A.; Bossa, F. Isolation, overexpression, and biochemical characterization of the two isoforms of glutamic acid decarboxylase from Escherichia coli. Protein Expr. Purif. 1996, 8, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Occhialini, A.; Jimenez de Bagues, M.P.; Saadeh, B.; Bastianelli, D.; Hanna, N.; De Biase, D.; Köhler, S. The glutamic acid decarboxylase system of the new species Brucella microti contributes to its acid resistance and to oral infection of mice. J. Infect. Dis. 2012, 206, 1424–1432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damiano, M.A.; Bastianelli, D.; Al Dahouk, S.; Köhler, S.; Cloeckaert, A.; De Biase, D.; Occhialini, A. Glutamate decarboxylase-dependent acid resistance in Brucella spp.: Distribution and contribution to fitness under extremely acidic conditions. Appl. Environ. Microbiol. 2015, 81, 578–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grassini, G.; Pennacchietti, E.; Cappadocio, F.; Occhialini, A.; De Biase, D. Biochemical and spectroscopic properties of Brucella microti glutamate decarboxylase, a key component of the glutamate-dependent acid resistance system. FEBS Open Bio 2015, 5, 209–218. [Google Scholar] [CrossRef]
- Kim, H.S.; Caswell, C.C.; Foreman, R.; Roop, R.M., 2nd; Crosson, S. The Brucella abortus general stress response system regulates chronic mammalian infection and is controlled by phosphorylation and proteolysis. J. Biol. Chem. 2013, 288, 13906–13916. [Google Scholar] [CrossRef] [Green Version]
- Raivio, T.L.; Silhavy, T.J. Periplasmic stress and ECF sigma factors. Ann. Rev. Microbiol. 2001, 55, 591–624. [Google Scholar] [CrossRef]
- Silva, T.M.; Costa, E.A.; Paixao, T.A.; Tsolis, R.M.; Santos, R.L. Laboratory animal models for brucellosis research. J. Biomed. Biotechnol. 2011, 2011, 518323. [Google Scholar] [CrossRef] [Green Version]
- Rossetti, C.A.; Drake, K.L.; Siddavatam, P.; Lawhon, S.D.; Nunes, J.E.; Gull, T.; Khare, S.; Everts, R.E.; Lewin, H.A.; Adams, L.G. Systems biology analysis of Brucella infected Peyer’s patch reveals rapid invasion with modest transient perturbations of the host transcriptome. PLoS ONE 2013, 8, e81719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elzer, P.H.; Hagius, S.D.; Davis, D.S.; DelVecchio, V.G.; Enright, F.M. Characterization of the caprine model for ruminant brucellosis. Vet. Microbiol. 2002, 90, 425–431. [Google Scholar] [CrossRef]
- Hensel, M.E.; Garcia-Gonzalez, D.G.; Chaki, S.P.; Hartwig, A.; Gordy, P.W.; Bowen, R.; Ficht, T.A.; Arenas-Gamboa, A.M. Vaccine Candidate Brucella melitensis 16MDeltavjbR Is Safe in a Pregnant Sheep Model and Confers Protection. mSphere 2020, 5, e00120-20. [Google Scholar] [CrossRef] [PubMed]
- Montaraz, J.A.; Winter, A.J. Comparison of living and nonliving vaccines for Brucella abortus in BALB/c mice. Infect. Immun. 1986, 53, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Young, E.J.; Gomez, C.I.; Yawn, D.H.; Musher, D.M. Comparison of Brucella abortus and Brucella melitensis infections of mice and their effect on acquired cellular resistance. Infect. Immun. 1979, 26, 680–685. [Google Scholar] [CrossRef] [Green Version]
- Rasool, O.; Freer, E.; Moreno, E.; Jarstrand, C. Effect of Brucella abortus lipopolysaccharide on oxidative metabolism and lysozyme release by human neutrophils. Infect. Immun. 1992, 60, 1699–1702. [Google Scholar] [CrossRef] [Green Version]
- Ouahrani-Bettache, S.; Jimenez De Bagues, M.P.; De La Garza, J.; Freddi, L.; Bueso, J.P.; Lyonnais, S.; Al Dahouk, S.; De Biase, D.; Köhler, S.; Occhialini, A. Lethality of Brucella microti in a murine model of infection depends on the wbkE gene involved in O-polysaccharide synthesis. Virulence 2019, 10, 868–878. [Google Scholar] [CrossRef]
- O’Callaghan, D.; Cazevieille, C.; Allardet-Servent, A.; Boschiroli, M.L.; Bourg, G.; Foulongne, V.; Frutos, P.; Kulakov, Y.; Ramuz, M. A homologue of the Agrobacterium tumefaciens VirB and Bordetella pertussis Ptl type IV secretion systems is essential for intracellular survival of Brucella suis. Mol. Microbiol. 1999, 33, 1210–1220. [Google Scholar] [CrossRef]
- Hanna, N.; Jimenez de Bagues, M.P.; Ouahrani-Bettache, S.; El Yakhlifi, Z.; Köhler, S.; Occhialini, A. The virB operon is essential for lethality of Brucella microti in the Balb/c murine model of infection. J. Infect. Dis. 2011, 203, 1129–1135. [Google Scholar] [CrossRef]
- Pappas, G.; Akritidis, N.; Bosilkovski, M.; Tsianos, E. Brucellosis. N. Engl. J. Med. 2005, 352, 2325–2336. [Google Scholar] [CrossRef]
- de Bagues, M.P.J.; de Martino, A.; Quintana, J.F.; Alcaraz, A.; Pardo, J. Course of infection with the emergent pathogen Brucella microti in immunocompromised mice. Infect. Immun. 2011, 79, 3934–3939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wareth, G.; Böttcher, D.; Melzer, F.; Shehata, A.A.; Roesler, U.; Neubauer, H.; Schoon, H.A. Experimental infection of chicken embryos with recently described Brucella microti: Pathogenicity and pathological findings. Comp. Immunol. Microbiol. Infect. Dis. 2015, 41, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Detilleux, P.G.; Cheville, N.F.; Deyoe, B.L. Pathogenesis of Brucella abortus in chicken embryos. Vet. Pathol. 1988, 25, 138–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gay, K.; Damon, S.R. A yolk sac technique for the routine isolation of Brucella; injection of clotted blood specimens into embryonating eggs with recovery of all three species. Public Health Rep. 1951, 66, 1204–1211. [Google Scholar] [CrossRef]
- Garcia-Mendez, K.B.; Hielpos, S.M.; Soler-Llorens, P.F.; Arce-Gorvel, V.; Hale, C.; Gorvel, J.P.; O’Callaghan, D.; Keriel, A. Infection by Brucella melitensis or Brucella papionis modifies essential physiological functions of human trophoblasts. Cell Microbiol. 2019, 21, e13019. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Occhialini, A.; Hofreuter, D.; Ufermann, C.-M.; Al Dahouk, S.; Köhler, S. The Retrospective on Atypical Brucella Species Leads to Novel Definitions. Microorganisms 2022, 10, 813. https://doi.org/10.3390/microorganisms10040813
Occhialini A, Hofreuter D, Ufermann C-M, Al Dahouk S, Köhler S. The Retrospective on Atypical Brucella Species Leads to Novel Definitions. Microorganisms. 2022; 10(4):813. https://doi.org/10.3390/microorganisms10040813
Chicago/Turabian StyleOcchialini, Alessandra, Dirk Hofreuter, Christoph-Martin Ufermann, Sascha Al Dahouk, and Stephan Köhler. 2022. "The Retrospective on Atypical Brucella Species Leads to Novel Definitions" Microorganisms 10, no. 4: 813. https://doi.org/10.3390/microorganisms10040813