
Distribution and Oxidation Rates of Ammonia-Oxidizing Archaea Influenced by the Coastal Upwelling off Eastern Hainan Island

Abstract
:1. Introduction
2. Materials and Methods
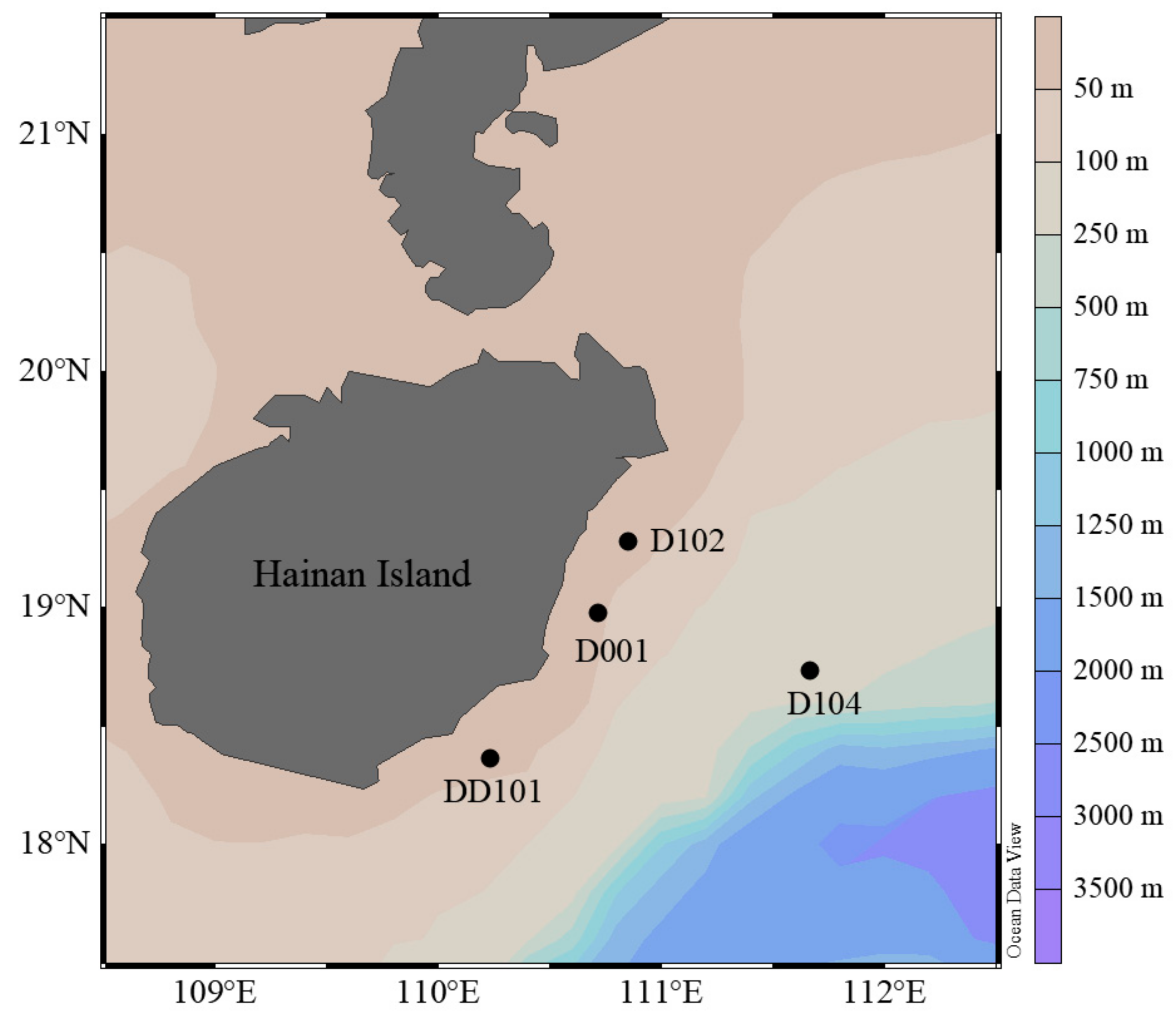
2.1. Sample Collection and Environmental Factor Measurement
2.2. DNA and RNA Extraction and cDNA Synthesis
2.3. Quantitative PCR
2.4. 454 Pyrosequencing and Bioinformatics Analysis
2.5. Nitrification Rate Measurements
2.6. Statistical Analysis and Accession Number
3. Results
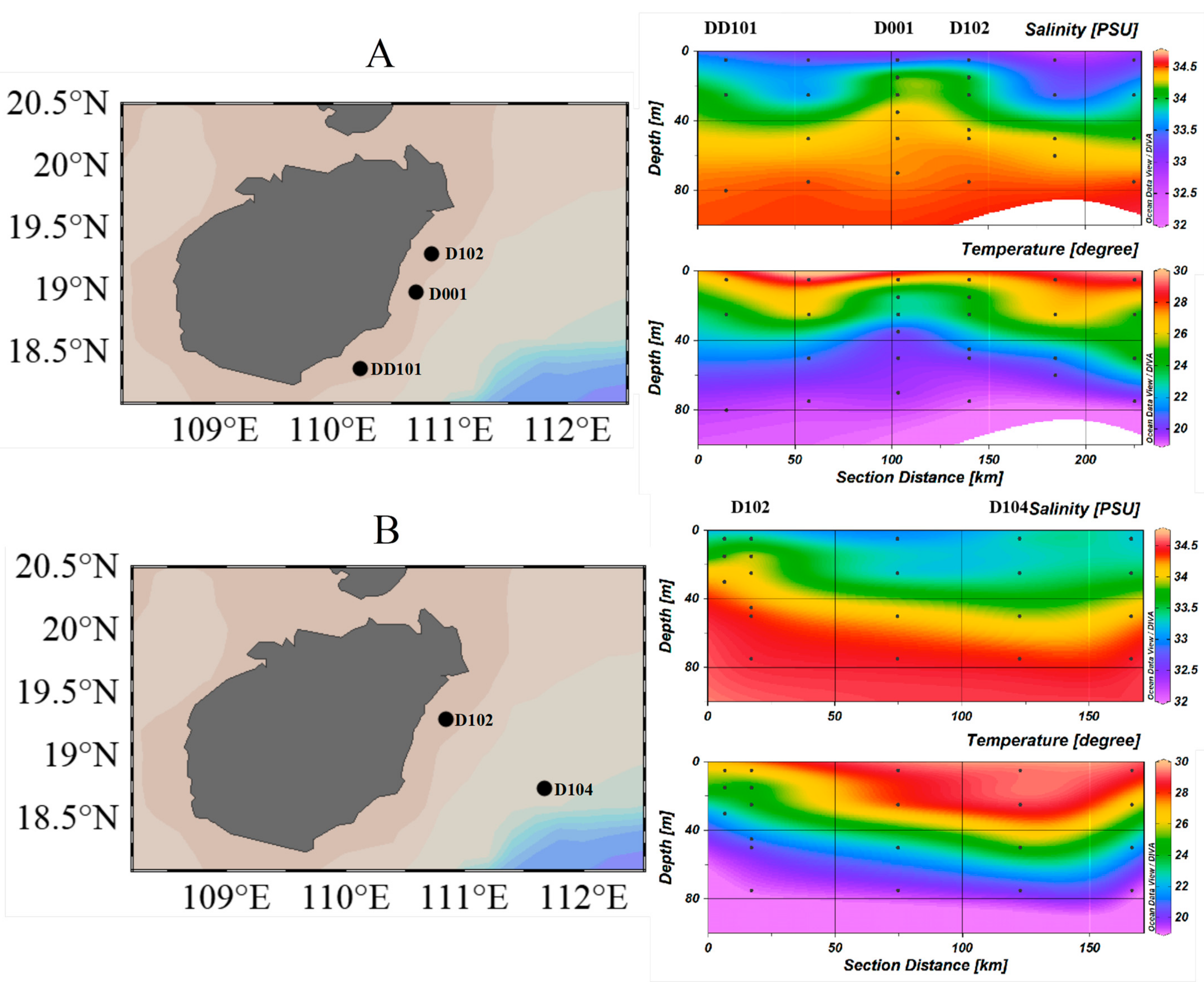
3.1. Hydrographic Conditions of Sampling Stations
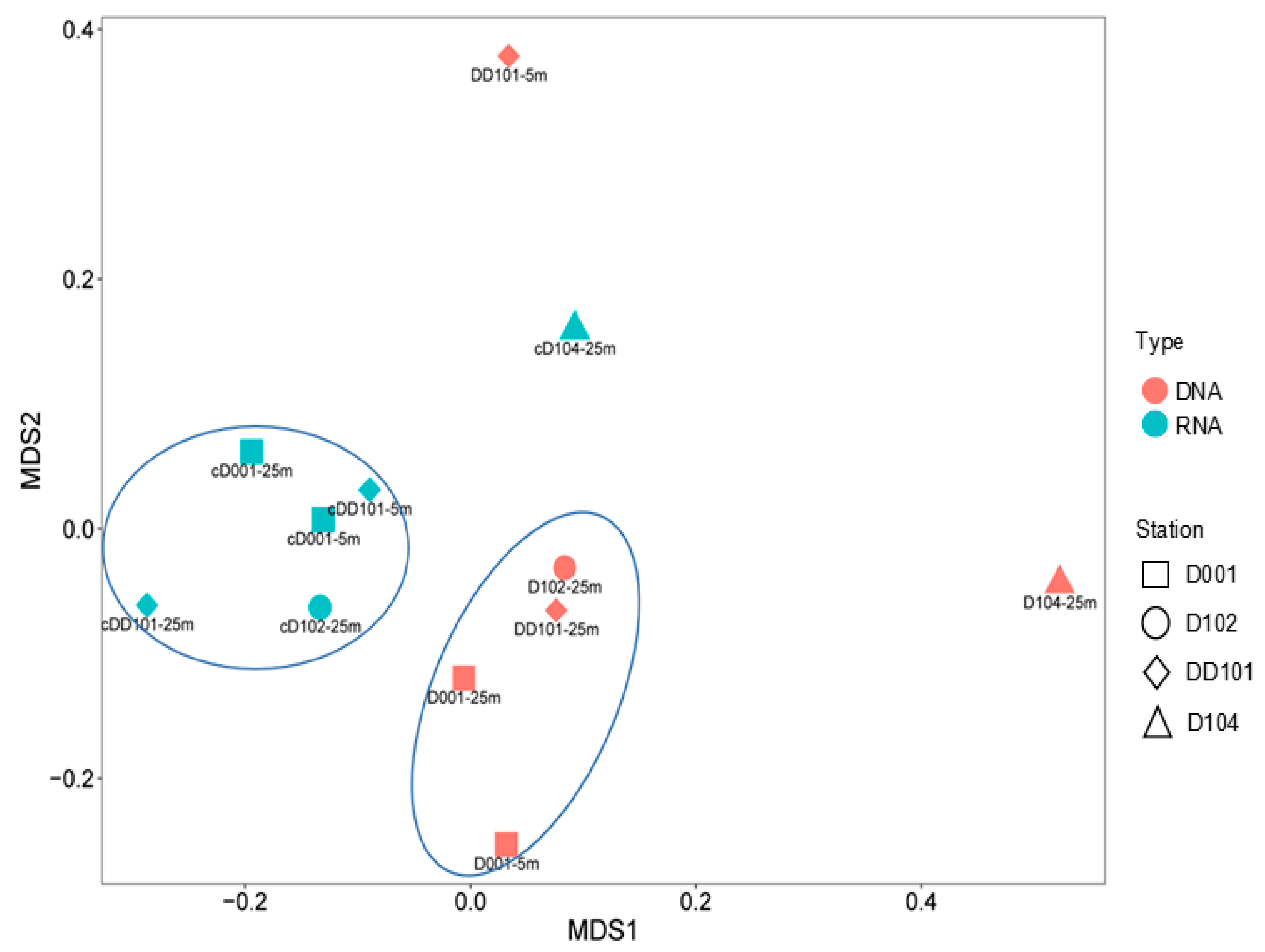
3.2. Diversity of AOA
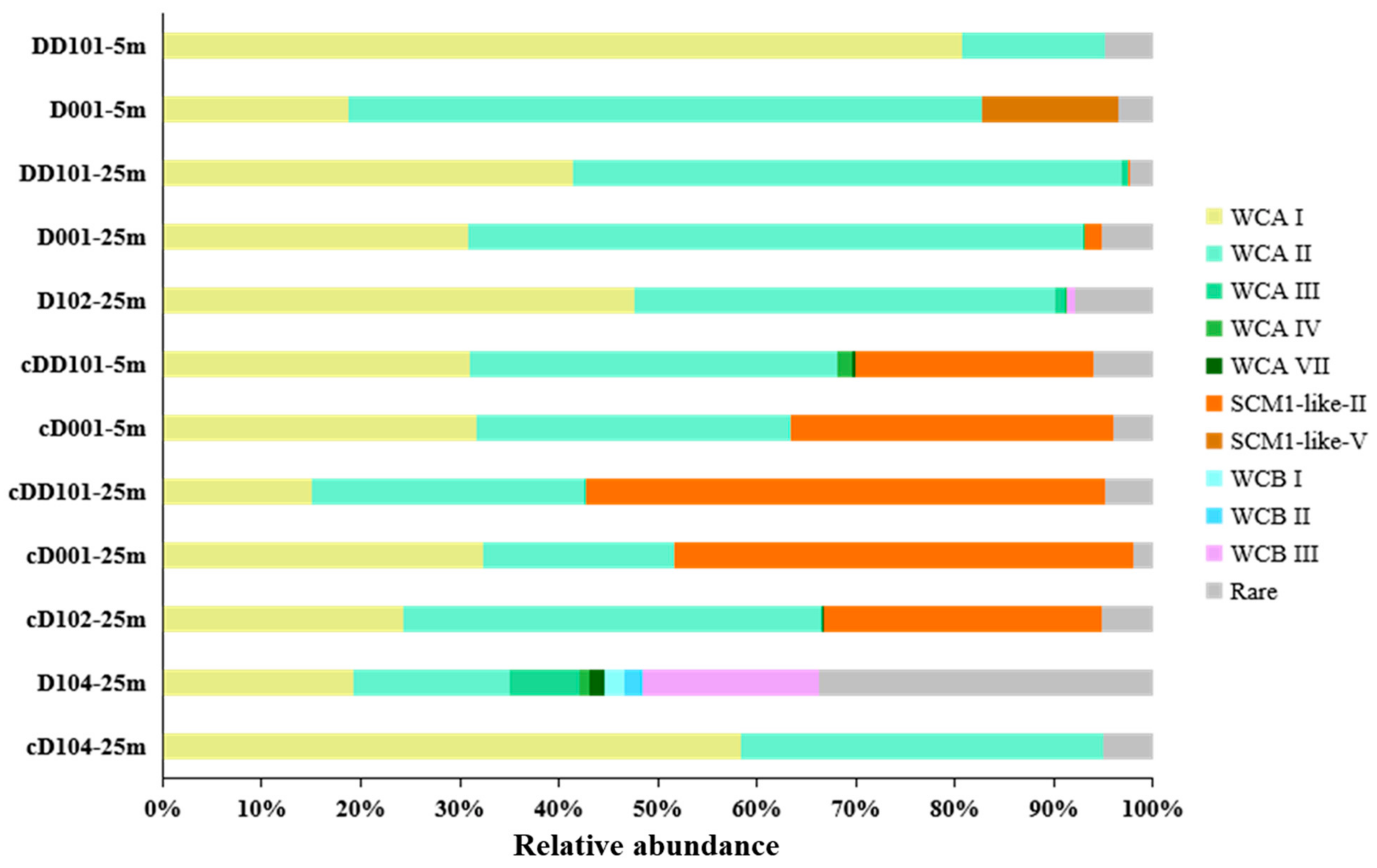
3.3. Phylogeny and Community Composition of AOA
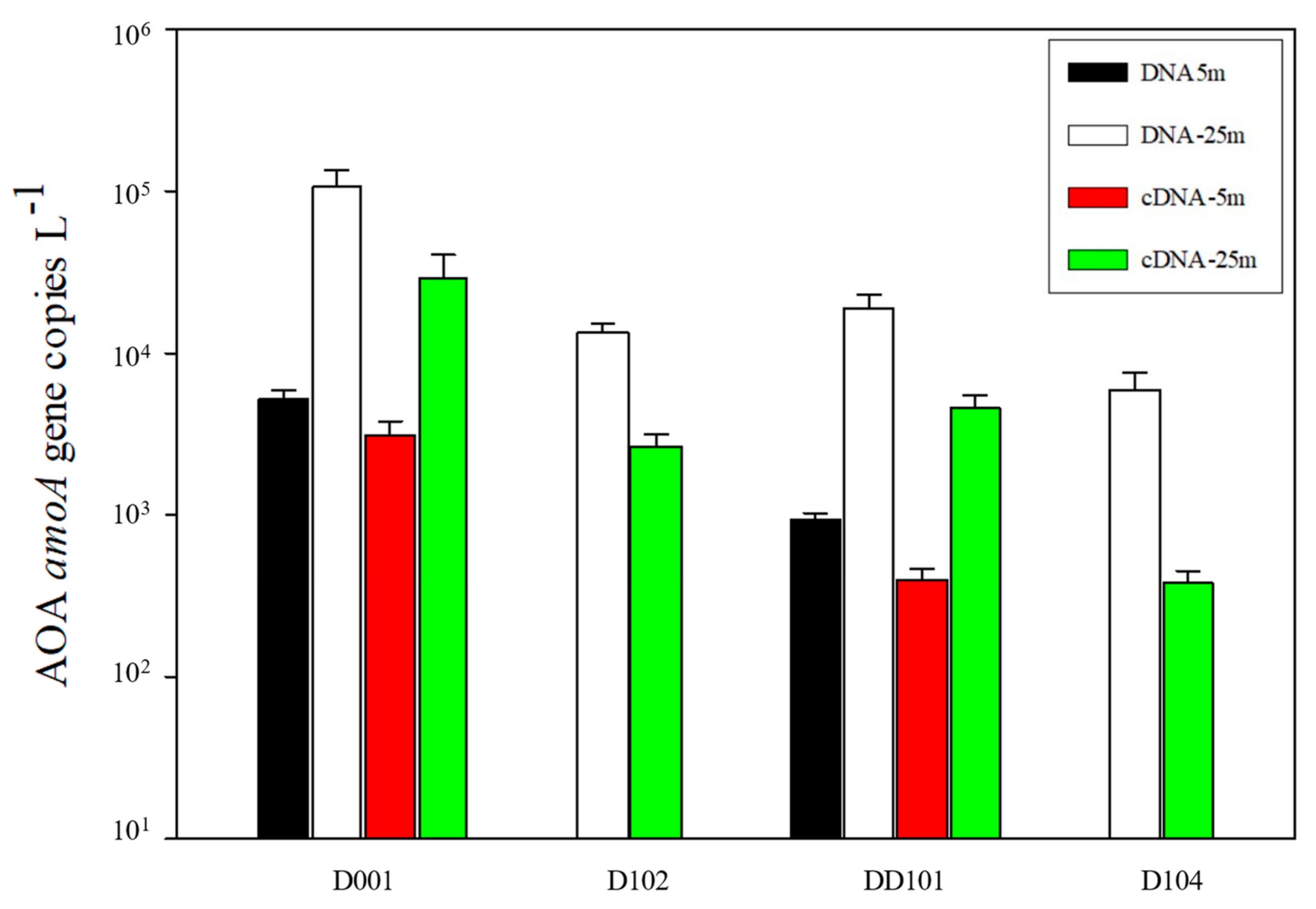
3.4. Abundance of AOA
3.5. Ammonia Oxidation Rates
4. Discussion
4.1. Upwelling Effects on AOA Abundance
4.2. Upwelling Effects on AOA Community Structure
4.3. Ammonia Oxidation Rates during Coastal Upwelling
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dang, H.; Zhou, H.; Yang, J.; Ge, H.; Jiao, N.; Luan, X.; Zhang, C.; Klotz, M.G. Thaumarchaeotal Signature Gene Distribution in Sediments of the Northern South China Sea: An Indicator of the Metabolic Intersection of the Marine Carbon, Nitrogen, and Phosphorus Cycles? Appl. Environ. Microbiol. 2013, 79, 2137–2147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mincer, T.J.; Church, M.J.; Taylor, L.T.; Preston, C.; Karl, D.M.; DeLong, E.F. Quantitative distribution of presumptive archaeal and bacterial nitrifiers in Monterey Bay and the North Pacific Subtropical Gyre. Environ. Microbiol. 2007, 9, 1162–1175. [Google Scholar] [CrossRef] [PubMed]
- Santoro, A.E.; Casciotti, K.; Francis, C. Activity, abundance and diversity of nitrifying archaea and bacteria in the central California Current. Environ. Microbiol. 2010, 12, 1989–2006. [Google Scholar] [CrossRef] [PubMed]
- Beman, J.M.; Sachdeva, R.; Fuhrman, J. Population ecology of nitrifying Archaea and Bacteria in the Southern California Bight. Environ. Microbiol. 2010, 12, 1282–1292. [Google Scholar] [CrossRef] [PubMed]
- Newell, S.; Fawcett, S.; Ward, B.B. Depth distribution of ammonia oxidation rates and ammonia-oxidizer community composition in the Sargasso Sea. Limnol. Oceanogr. 2013, 58, 1491–1500. [Google Scholar] [CrossRef] [Green Version]
- Francis, C.A.; Roberts, K.J.; Beman, J.M.; Santoro, A.E.; Oakley, B.B. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. USA 2005, 102, 14683–14688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beman, J.M.; Popp, B.; Francis, C. Molecular and biogeochemical evidence for ammonia oxidation by marine Crenarchaeota in the Gulf of California. ISME J. 2008, 2, 429–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoro, A.E.; Saito, M.A.; Goepfert, T.J.; Lamborg, C.; Dupont, C.L.; DiTullio, G.R. Thaumarchaeal ecotype distributions across the equatorial Pacific Ocean and their potential roles in nitrification and sinking flux attenuation. Limnol. Oceanogr. 2017, 62, 1984–2003. [Google Scholar] [CrossRef]
- Cheung, S.; Mak, W.; Xia, X.; Lu, Y.; Cheung, Y.; Liu, H. Overlooked Genetic Diversity of Ammonia Oxidizing Archaea Lineages in the Global Oceans. J. Geophys. Res. Biogeosci. 2019, 124, 1799–1811. [Google Scholar] [CrossRef]
- Könneke, M.; Bernhard, A.E.; José, R.; Walker, C.B.; Waterbury, J.B.; Stahl, D.A. Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature 2005, 437, 543–546. [Google Scholar] [CrossRef]
- Biller, S.J.; Mosier, A.C.; Wells, G.F.; Francis, C.A. Global Biodiversity of Aquatic Ammonia-Oxidizing Archaea is Partitioned by Habitat. Front. Microbiol. 2012, 3, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoro, A.E.; Richter, R.A.; Dupont, C.L. Planktonic Marine Archaea. Annu. Rev. Mar. Sci. 2019, 11, 131–158. [Google Scholar] [CrossRef] [PubMed]
- Pester, M.; Rattei, T.; Flechl, S.; Grngrft, A.; Wagner, M. Amoa-based consensus phylogeny of ammonia-oxidizing archaea and deep sequencing of amoA genes from soils of four different geographic regions. Environ. Microbiol. 2012, 14, 525–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sintes, E.; Bergauer, K.; De Corte, D.; Yokokawa, T.; Herndl, G.J. Archaeal amoA gene diversity points to distinct biogeography of ammonia-oxidizing Crenarchaeota in the ocean. Environ. Microbiol. 2013, 15, 1647–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina, V.; Belmar, L.; Levipan, H.A.; Ramírez-Flandes, S.; Anguita, C.; Galán, A.; Montes, I.; Ulloa, O. Spatiotemporal Distribution of Key Pelagic Microbes in a Seasonal Oxygen-Deficient Coastal Upwelling System of the Eastern South Pacific Ocean. Front. Mar. Sci. 2020, 7, 561597. [Google Scholar] [CrossRef]
- Lu, Y.; Xia, X.; Cheung, S.; Jing, H.; Liu, H. Differential Distribution and Determinants of Ammonia Oxidizing Archaea Sublineages in the Oxygen Minimum Zone off Costa Rica. Microorganisms 2019, 7, 453. [Google Scholar] [CrossRef] [Green Version]
- Happel, E.; Bartl, I.; Voss, M.; Riemann, L. Extensive nitrification and active ammonia oxidizers in two contrasting coastal systems of the Baltic Sea. Environ. Microbiol. 2018, 20, 2913–2926. [Google Scholar] [CrossRef]
- Ward, B.; Zafiriou, O. Nitrification and nitric oxide in the oxygen minimum of the eastern tropical North Pacific. Deep Sea Res. Part A Oceanogr. Res. Pap. 1988, 35, 1127–1142. [Google Scholar] [CrossRef]
- Dolmatova, L.S.; Eliseykina, M.G.; Timchenko, N.F.; Kovaleva, A.L.; Shitkova, O.A. Generation of reactive oxygen species in different fractions of the coelomocytes of holothurianEupentacta fraudatrix in response to the thermostable toxin ofYersinia pseudotuberculosis in vitro. Chin. J. Oceanol. Limnol. 2003, 21, 293–304. [Google Scholar] [CrossRef]
- Gan, J.; Cheung, A.; Guo, X.; Li, L. Intensified upwelling over a widened shelf in the northeastern South China Sea. J. Geophys. Res. Earth Surf. 2009, 114, 09019. [Google Scholar] [CrossRef]
- Liu, S.; Hong, B.; Wang, G.; Wang, W.; Xie, Q.; Ni, Z.; Yu, L.; Jiang, H.; Long, T.; Xu, H. Physical structure and phytoplankton community off the eastern Hainan coast during summer 2015. Acta Oceanol. Sin. 2020, 39, 103–114. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoro, A.E.; Dupont, C.L.; Richter, R.A.; Craig, M.T.; Carini, P.; McIlvin, M.R.; Yang, Y.; Orsi, W.D.; Moran, D.M.; Saito, M.A. Genomic and proteomic characterization of “Candidatus Nitrosopelagicus brevis: An ammonia-oxidizing archaeon from the open ocean. Proc. Natl. Acad. Sci. USA 2015, 112, 1173–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, M.R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.M.; Stevens, H.H.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6. 2019. Available online: https://CRAN.R-project.org/package (accessed on 1 August 2021).
- Logares, R.; Audic, S.; Bass, D.; Bittner, L.; Boutte, C.; Christen, R.; Claverie, J.-M.; Decelle, J.; Dolan, J.R.; Dunthorn, M.; et al. Patterns of Rare and Abundant Marine Microbial Eukaryotes. Curr. Biol. 2014, 24, 813–821. [Google Scholar] [CrossRef] [Green Version]
- Jing, H.; Cheung, S.Y.; Xia, X.; Suzuki, K.; Nishioka, J.; Liu, H. Geographic Distribution of Ammonia-Oxidizing Archaea along the Kuril Islands in the Western Subarctic Pacific. Front. Microbiol. 2017, 8, 1247. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Ward, B.B.; Kilpatrick, K.A.; Renger, E.H.; Eppley, R.W. Biological nitrogen cycling in the nitracline. Limnol. Oceanogr. 1989, 34, 493–513. [Google Scholar] [CrossRef]
- Sigman, D.M.; Casciotti, K.L.; Andreani, M.; Barford, C.; Galanter, M.; Böhlke, J.K. A Bacterial Method for the Nitrogen Isotopic Analysis of Nitrate in Seawater and Freshwater. Anal. Chem. 2001, 73, 4145–4153. [Google Scholar] [CrossRef]
- Casciotti, K.L.; Sigman, D.M.; Hastings, M.G.; Böhlke, J.K.; Hilkert, A. Measurement of the Oxygen Isotopic Composition of Nitrate in Seawater and Freshwater Using the Denitrifier Method. Anal. Chem. 2002, 74, 4905–4912. [Google Scholar] [CrossRef]
- Knapp, A.N.; Sigman, D.M.; Lipschultz, F. N isotopic composition of dissolved organic nitrogen and nitrate at the Bermuda Atlantic Time-series Study site. Glob. Biogeochem. Cycles 2005, 19, 1018. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Agogué, H.; Brink, M.; Dinasquet, J.; Herndl, G.J. Erratum: Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic. Nature 2009, 457, 500. [Google Scholar] [CrossRef] [Green Version]
- Tolar, B.B.; King, G.M.; Hollibaugh, J.T. An Analysis of Thaumarchaeota Populations from the Northern Gulf of Mexico. Front. Microbiol. 2013, 4, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.M.; Damashek, J.; Chavez, F.P.; Francis, C.A. Factors influencing nitrification rates and the abundance and transcriptional activity of ammonia-oxidizing microorganisms in the dark northeast Pacific Ocean. Limnol. Oceanogr. 2016, 61, 596–609. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Cheung, S.Y.; Chen, L.; Kao, S.-J.; Xia, X.; Gan, J.; Dai, M.; Liu, H. New insight to niche partitioning and ecological function of ammonia oxidizing archaea in subtropical estuarine ecosystem. Biogeosciences 2020, 17, 6017–6032. [Google Scholar] [CrossRef]
- Merbt, S.N.; Stahl, D.A.; Casamayor, E.O.; Martí, E.; Nicol, G.W.; Prosser, J.I. Differential photoinhibition of bacterial and archaeal ammonia oxidation. FEMS Microbiol. Lett. 2011, 327, 41–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.M.; Chavez, F.P.; Francis, C. Ammonium Uptake by Phytoplankton Regulates Nitrification in the Sunlit Ocean. PLoS ONE 2014, 9, e108173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, W.; Amin, S.A.; Martens-Habbena, W.; Walker, C.B.; Urakawa, H.; Devol, A.H.; Ingalls, A.E.; Moffett, J.W.; Armbrust, E.V.; Stahl, D.A. Marine ammonia-oxidizing archaeal isolates display obligate mixotrophy and wide ecotypic variation. Proc. Natl. Acad. Sci. USA 2014, 111, 12504–12509. [Google Scholar] [CrossRef] [Green Version]
- Martens-Habbena, W.; Berube, P.M.; Urakawa, H.; de la Torre, J.R.; Stahl, D.A. Ammonia oxidation kinetics determine niche separation of nitrifying Archaea and Bacteria. Nature 2009, 461, 976–979. [Google Scholar] [CrossRef]
- Xu, M.N.; Zhang, W.; Zhu, Y.; Liu, L.; Zheng, Z.; Wan, X.S.; Qian, W.; Dai, M.; Gan, J.; Hutchins, D.A.; et al. Enhanced Ammonia Oxidation Caused by Lateral Kuroshio Intrusion in the Boundary Zone of the Northern South China Sea. Geophys. Res. Lett. 2018, 45, 6585–6593. [Google Scholar] [CrossRef]
- Ward, B.B. Nitrification in the marine environment. In Nitrogen in the Marine Environment; Capone, D.G., Bronk, D.A., Mulholland, M.R., Carpenter, E.J., Eds.; Elsevier: San Diego, CA, USA, 2008; pp. 199–262. [Google Scholar]
- Wetz, M.; Wheeler, P. Response of bacteria to simulated upwelling phytoplankton blooms. Mar. Ecol. Prog. Ser. 2004, 272, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Fernández, C.; Farias, L.; Alcaman, M. Primary production and nitrogen regeneration processes in surface waters of the Peruvian upwelling system. Prog. Oceanogr. 2009, 83, 159–168. [Google Scholar] [CrossRef]
- Church, M.J.; Wai, B.; Karl, D.M.; DeLong, E.F. Abundances of crenarchaeal amoA genes and transcripts in the Pacific Ocean. Environ. Microbiol. 2010, 12, 679–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | No. of Seqs * | No. of OTUs | J (Pielous’ Evenness) | Shannon | Simpson | Coverage (%) |
|---|---|---|---|---|---|---|
| D001—5m | 1200 | 47 | 0.41 | 1.59 | 0.62 | 97.82 |
| cD001—5m | 1200 | 50 | 0.49 | 1.91 | 0.78 | 97.92 |
| D001—25m | 1200 | 68 | 0.42 | 1.79 | 0.67 | 96.81 |
| cD001—25m | 1200 | 30 | 0.49 | 1.68 | 0.74 | 98.81 |
| DD101—5m | 1200 | 43 | 0.33 | 1.23 | 0.51 | 98.18 |
| cDD101—5m | 1200 | 56 | 0.58 | 2.33 | 0.84 | 98.07 |
| DD101—25m | 1200 | 38 | 0.44 | 1.60 | 0.68 | 98.38 |
| cDD101—25m | 1200 | 61 | 0.42 | 1.73 | 0.67 | 97.29 |
| D102—25m | 1200 | 85 | 0.52 | 2.31 | 0.79 | 96.08 |
| cD102—25m | 1200 | 65 | 0.46 | 1.90 | 0.74 | 96.91 |
| D104—25m | 1200 | 138 | 0.83 | 4.09 | 0.97 | 97.64 |
| cD104—25m | 1200 | 54 | 0.53 | 2.12 | 0.79 | 97.78 |
| Station | Depth (m) | Ambient NH4+ (nM) | Ambient NOx− (μM) | AO rate (nmol L−1 d−1) | |
|---|---|---|---|---|---|
| Upwelling | D001 | 25 | 72.75 ± 2.58 | 0.20 ± 0.08 | 2.64 ± 0.29 |
| 75 | 97.13 ± 8.40 | 7.01 ± 1.61 | 112.00 ± 14.27 | ||
| D102 | 5 | 55.93 ± 10.33 | 0.13 ± 0.08 | 1.53 ± 1.11 | |
| 25 | 57.56 ± 5.22 | 0.48 ± 0.40 | 5.63 ± 5.84 | ||
| 50 | 86.57 ± 6.84 | 0.87 ± 0.26 | 11.97 ± 5.37 | ||
| DD101 | 25 | 19.98 ± 0.68 | 0.23 ± 0.12 | 0.77 ± 0.47 | |
| 75 | 51.28 ± 3.99 | 3.93 ± 2.61 | 52.00 ± 13.25 | ||
| Non-Upwelling | 5 | 13.62 ± 0.69 | 0.19 ± 0.03 | 0.13 ± 0.01 | |
| D104 | 25 | 14.26 ± 0.29 | 3.83 ± 1.54 | 4.11 ± 5.93 | |
| 50 | 9.98 ± 0.15 | 8.51 ± 2.41 | 3.31 ± 1.67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Zhou, P.; Cheung, S.; Lu, Y.; Liu, H.; Jing, H. Distribution and Oxidation Rates of Ammonia-Oxidizing Archaea Influenced by the Coastal Upwelling off Eastern Hainan Island. Microorganisms 2022, 10, 952. https://doi.org/10.3390/microorganisms10050952
Liu H, Zhou P, Cheung S, Lu Y, Liu H, Jing H. Distribution and Oxidation Rates of Ammonia-Oxidizing Archaea Influenced by the Coastal Upwelling off Eastern Hainan Island. Microorganisms. 2022; 10(5):952. https://doi.org/10.3390/microorganisms10050952
Chicago/Turabian StyleLiu, Hao, Peng Zhou, Shunyan Cheung, Yanhong Lu, Hongbin Liu, and Hongmei Jing. 2022. "Distribution and Oxidation Rates of Ammonia-Oxidizing Archaea Influenced by the Coastal Upwelling off Eastern Hainan Island" Microorganisms 10, no. 5: 952. https://doi.org/10.3390/microorganisms10050952