
Biodegradation, Decolorization, and Detoxification of Di-Azo Dye Direct Red 81 by Halotolerant, Alkali-Thermo-Tolerant Bacterial Mixed Cultures

 ,
,  ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Dyestuff
2.2. Microorganisms, Culture Conditions and Acclimatization
2.2.1. Sample Collection
2.2.2. Development and Enrichment of Culturable Mixed Microbial Cultures
2.3. Decolorization Studies
2.3.1. Extent of Decolorization in MSM-Y and Real Wastewater
2.3.2. Effect of Initial Dye Concentration on Decolorization in Real Wastewater
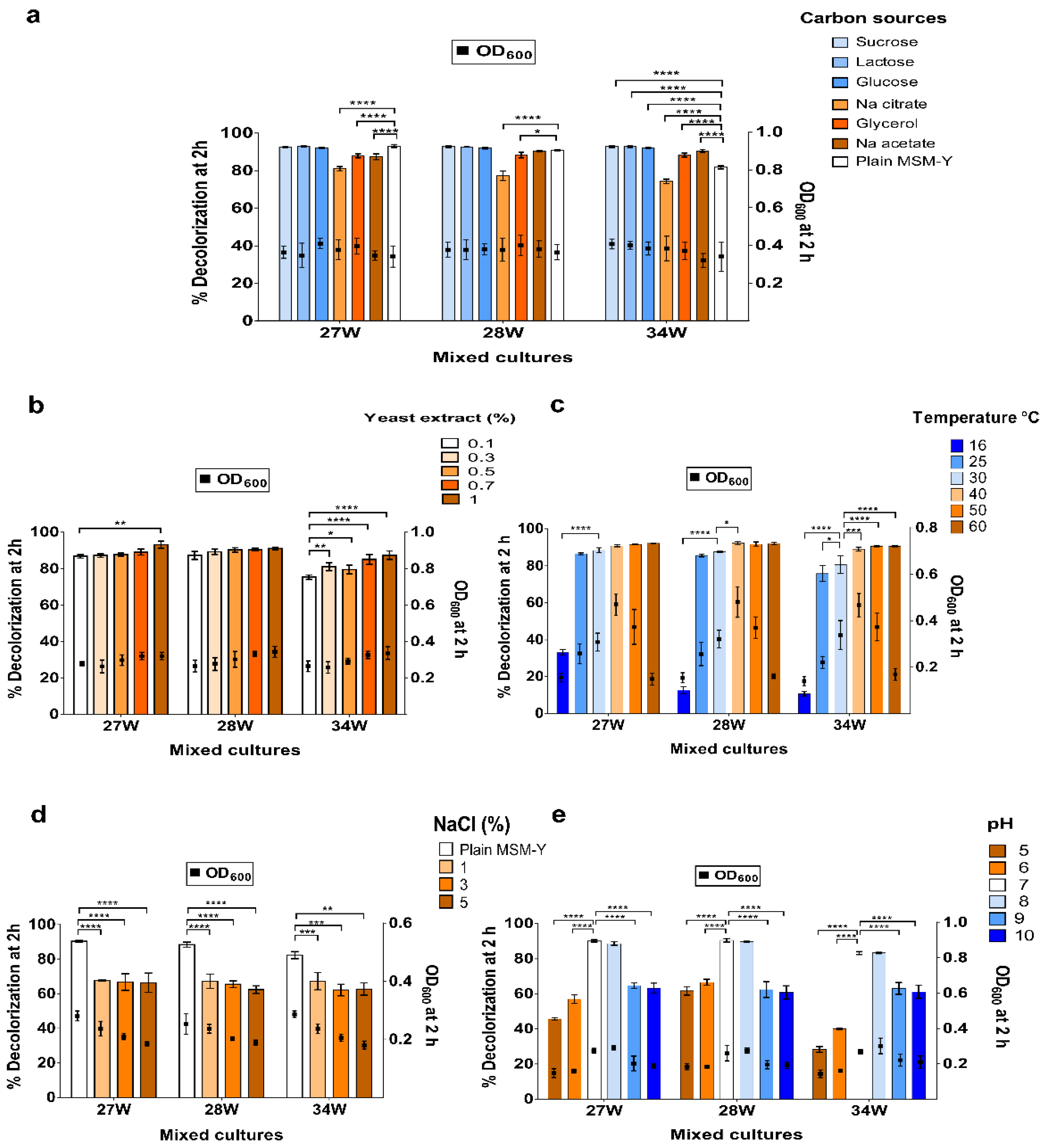
2.4. Factors Affecting DR81 Decolorization by the Selected Mixed Bacterial Cultures
2.5. Molecular Identification of Pure Isolated Members of the Selected Mixed Bacterial Cultures Using Sanger Sequencing of the 16S rRNA Gene Full Length
2.6. Screening for Azo Dye Degrading Enzymes in the Selected Mixed Bacterial Cultures
2.6.1. Preparation of the Cell-Free Supernatant
2.6.2. Screening for Reductive and Oxidative Degrading Enzymes
2.6.3. Azoreductase Optimum Activity and Stability Testing
2.7. Identification of Azoreductase Genes among the Selected Mixed Bacterial Cultures
2.8. Metabolites Extraction and Isolation
2.8.1. Spectroscopic Determination
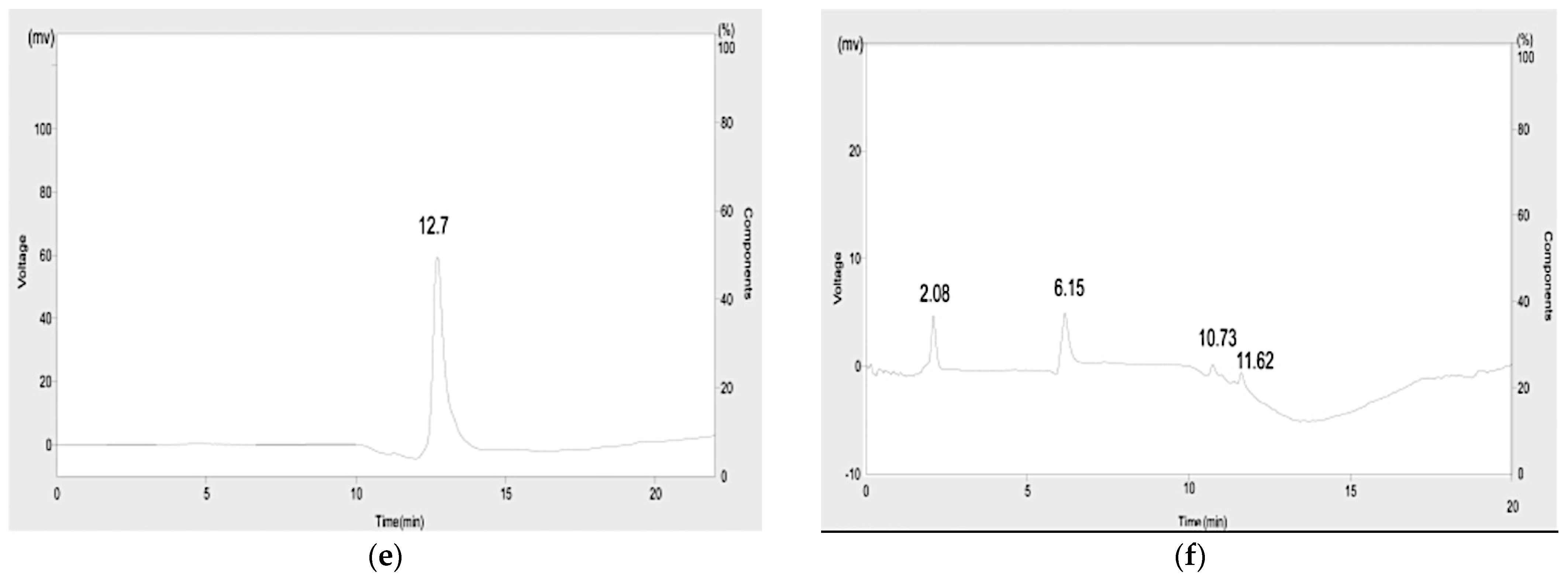
2.8.2. Hyphenated Techniques
2.9. Toxicity Studies
3. Results
3.1. Determination of the Extent of DR81 Decolorization by Mixed Bacterial Cultures
3.2. Effect of Initial Dye Concentration on Decolorization in Real Industrial Wastewater
3.3. Factors Affecting DR81 Decolorization
3.4. Molecular Identification of Members of the Selected Mixed Bacterial Cultures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mixed Cultures | Isolates | Identification | Max Score | Total Score | % Identity 1 | Accession |
|---|---|---|---|---|---|---|
| 27W | A | Stenotrophomonas maltophilia | 1840 | 7361 | 99.22% | CP044092.1 |
| B | Proteus vulgaris | 1812 | 1812 | 99.90% | MN685224.1 | |
| C | Achromobacter xylosoxidans | 1921 | 1921 | 99.90% | MN904889.1 | |
| D | Pseudomonas sp. YP1 | 1652 | 1652 | 99.56% | KF719297.1 | |
| E | Pseudomonas monteilii | 1531 | 1531 | 99.76% | MN889010.1 | |
| F | Paenibacillus agaridevorans | 1886 | 1886 | 99.90% | KU922394.1 | |
| 28W | A | Agromyces mediolanus | 2065 | 2065 | 99.82% | MF459693.1 |
| B | Stenotrophomonas maltophilia | 1927 | 7708 | 100.00% | CP044092.1 | |
| C | Paenibacillus agaridevorans | 1886 | 1886 | 99.90% | KU922394.1 | |
| D | Pseudomonas sp. YP17 | 1565 | 1565 | 100.00% | KF719295.1 | |
| E | Pseudomonas taiwanensis | 1940 | 1940 | 99.81% | MN082103.1 | |
| 34W | A | Alcaligenes faecalis | 1869 | 1869 | 99.90% | MN515060.1 |
| B | Brevundimonas diminuta | 1943 | 1943 | 99.72% | MN923411.1 | |
| C | Pseudochrobactrum sp. | 1592 | 1592 | 100.00% | KC337108.1 |
3.5. Screening for Reductive and Oxidative Degrading Enzymes
3.6. Azoreductase Optimum Activity
3.7. Azoreductase Stability Testing
3.8. Identification of Azoreductase Genes in the Selected Mixed Bacterial Cultures
3.9. Extraction of Metabolites and Prediction of Biodegradative Pathway
3.10. Toxicity Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Das, A.; Mishra, S. Removal of textile dye reactive green-19 using bacterial consortium: Process optimization using response surface methodology and kinetics study. J. Environ. Chem. Eng. 2017, 5, 612–627. [Google Scholar] [CrossRef]
- Al-Tohamy, R.; Kenawy, E.-R.; Sun, J.; Ali, S.S. Performance of a newly isolated salt-tolerant yeast strain Sterigmatomyces halophilus SSA-1575 for azo dye decolorization and detoxification. Front. Microbiol. 2020, 11, 1163. [Google Scholar] [CrossRef] [PubMed]
- Rajashekarappa, K.K.; Mahadevan, G.D.; Neelagund, S.E.; Sathynarayana, M.; Vijaya, D.; Mulla, S.I. Decolorization of amaranth RI and fast red E azo dyes by thermophilic Geobacillus thermoleovorans KNG 112. J. Chem. Technol. Biotechnol. 2021, 97, 482–489. [Google Scholar] [CrossRef]
- Yagub, M.T.; Sen, T.K.; Afroze, S.; Ang, H.M. Dye and its removal from aqueous solution by adsorption: A review. Adv. Colloid Interface Sci. 2014, 209, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Yagub, M.T.; Sen, T.K.; Ang, H.M. Equilibrium, kinetics, and thermodynamics of methylene blue adsorption by pine tree leaves. Water Air Soil Pollut. 2012, 223, 5267–5282. [Google Scholar] [CrossRef]
- Shah, M.P. Microbe-mediated degradation of synthetic dyes in wastewater. In Microbial Degradation of Synthetic Dyes in Wastewaters; Springer: Cham, Switzerland, 2015; pp. 205–241. [Google Scholar]
- Kumar, C.G.; Mongolla, P. Microbial Degradation of Basic Dyes in Wastewaters. In Microbial Degradation of Synthetic Dyes in Wastewaters; Singh, S.N., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 85–110. [Google Scholar]
- Tkaczyk, A.; Mitrowska, K.; Posyniak, A. Synthetic organic dyes as contaminants of the aquatic environment and their implications for ecosystems: A review. Sci. Total Environ. 2020, 717, 137222. [Google Scholar] [CrossRef]
- Solís, M.; Solís, A.; Pérez, H.I.; Manjarrez, N.; Flores, M. Microbial decolouration of azo dyes: A review. Process Biochem. 2012, 47, 1723–1748. [Google Scholar] [CrossRef]
- Saxena, G.; Purchase, D.; Mulla, S.I.; Bharagava, R.N. Degradation and detoxification of leather tannery effluent by a newly developed bacterial consortium GS-TE1310 for environmental safety. J. Water Process Eng. 2020, 38, 101592. [Google Scholar] [CrossRef]
- Guo, G.; Hao, J.; Tian, F.; Liu, C.; Ding, K.; Zhang, C.; Yang, F.; Xu, J. Decolorization of metanil yellow G by a halophilic alkalithermophilic bacterial consortium. Bioresour. Technol. 2020, 316, 123923. [Google Scholar] [CrossRef]
- Donner, E.; Eriksson, E.; Holten-Lützhøft, H.-C.; Scholes, L.; Revitt, M.; Ledin, A. Identifying and classifying the sources and uses of xenobiotics in urban environments. In Xenobiotics in the Urban Water Cycle; Springer: Cham, Switzerland, 2010; pp. 27–50. [Google Scholar]
- Chen, H. Recent advances in azo dye degrading enzyme research. Curr. Protein Pept. Sci. 2006, 7, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Sreedharan, V.; Rao, K.V.B. Biodegradation of textile azo dyes. In Nanoscience and Biotechnology for Environmental Applications; Springer: Cham, Switzerland, 2019; pp. 115–139. [Google Scholar]
- Hsueh, C.-C.; Chen, C.-T.; Hsu, A.-W.; Wu, C.-C.; Chen, B.-Y. Comparative assessment of azo dyes and nitroaromatic compounds reduction using indigenous dye-decolorizing bacteria. J. Taiwan Inst. Chem. Eng. 2017, 79, 134–140. [Google Scholar] [CrossRef]
- Dave, S.R.; Patel, T.L.; Tipre, D.R. Bacterial Degradation of Azo Dye Containing Wastes. In Microbial Degradation of Synthetic Dyes in Wastewaters; Singh, S.N., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 57–83. [Google Scholar]
- Ogola, H.J.O.; Ashida, H.; Ishikawa, T.; Sawa, Y. Explorations and Applications of Enzyme-linked Bioremediation of Synthetic Dyes. In Advances in Bioremediation of Wastewater and Polluted Soil; Shiomi, N., Ed.; Intech: London, UK, 2015; pp. 111–144. [Google Scholar]
- Masarbo, R.S.; Karegoudar, T. Decolourisation of toxic azo dye Fast Red E by three bacterial strains: Process optimisation and toxicity assessment. Int. J. Environ. Anal. Chem. 2020. [Google Scholar] [CrossRef]
- Sahasrabudhe, M.M.; Saratale, R.G.; Saratale, G.D.; Pathade, G.R. Decolorization and detoxification of sulfonated toxic diazo dye CI Direct Red 81 by Enterococcus faecalis YZ 66. J. Environ. Health Sci. Eng. 2014, 12, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elfarash, A.; Mawad, A.M.; Yousef, N.M.; Shoreit, A.A. Azoreductase kinetics and gene expression in the synthetic dyes-degrading Pseudomonas. Egypt J. Basic Appl. Sci. 2017, 4, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Amin, S.; Rastogi, R.P.; Chaubey, M.G.; Jain, K.; Divecha, J.; Desai, C.; Madamwar, D. Degradation and toxicity analysis of a reactive textile diazo dye-Direct Red 81 by newly isolated Bacillus sp. DMS2. Front. Microbiol. 2020, 11, 2280. [Google Scholar] [CrossRef] [PubMed]
- Kudlich, M.; Hetheridge, M.J.; Knackmuss, H.-J.; Stolz, A. Autoxidation reactions of different aromatic o-aminohydroxynaphthalenes that are formed during the anaerobic reduction of sulfonated azo dyes. Environ. Sci. Technol. 1999, 33, 896–901. [Google Scholar] [CrossRef]
- Balapure, K.H.; Jain, K.; Chattaraj, S.; Bhatt, N.S.; Madamwar, D. Co-metabolic degradation of diazo dye—Reactive blue 160 by enriched mixed cultures BDN. J. Hazard. Mater. 2014, 279, 85–95. [Google Scholar] [CrossRef]
- Ismail, M.M.; Essam, T.M.; Ragab, Y.M.; Mourad, F.E. Biodegradation of ketoprofen using a microalgal–bacterial consortium. Biotechnol. Lett. 2016, 38, 1493–1502. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Alvarez, J.A.; Marañón-Ruiz, V.-F.; Jáuregui-Rincón, J.; Medina-Ramírez, I.; Frausto-Reyes, C.; Salinas-Gutiérrez, R. Removal of direct dyes with alginic acid. J. Mex. Chem. Soc. 2015, 59, 215–227. [Google Scholar]
- Walger, E.; Marlin, N.; Molton, F.; Mortha, G. Study of the Direct Red 81 Dye/Copper(II)-Phenanthroline System. Molecules 2018, 23, 242. [Google Scholar] [CrossRef] [Green Version]
- Khalid, A.; Arshad, M.; Crowley, D. Bioaugmentation of Azo Dyes. In Biodegradation of Azo Dyes; Atacag Erkurt, H., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1–37. [Google Scholar]
- Hashem, R.A.; Samir, R.; Essam, T.M.; Ali, A.E.; Amin, M.A. Optimization and enhancement of textile reactive Remazol black B decolorization and detoxification by environmentally isolated pH tolerant Pseudomonas aeruginosa KY284155. AMB Expr. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Eraqi, W.A.; ElRakaiby, M.T.; Megahed, S.A.; Yousef, N.H.; Elshahed, M.S.; Yassin, A.S. The Nile River microbiome reveals a remarkably stable community between wet and dry seasons, and sampling sites, in a large urban metropolis (Cairo, Egypt). OMICS A J. Integr. Biol. 2018, 22, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Kalyani, D.; Telke, A.; Dhanve, R.; Jadhav, J. Ecofriendly biodegradation and detoxification of Reactive Red 2 textile dye by newly isolated Pseudomonas sp. SUK1. J. Hazard. Mater. 2009, 163, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Franciscon, E.; Grossman, M.J.; Paschoal, J.A.R.; Reyes, F.G.R.; Durrant, L.R. Decolorization and biodegradation of reactive sulfonated azo dyes by a newly isolated Brevibacterium sp. strain VN-15. Springerplus 2012, 1, 37. [Google Scholar] [CrossRef] [Green Version]
- Saratale, R.G.; Gandhi, S.S.; Purankar, M.V.; Kurade, M.B.; Govindwar, S.P.; Oh, S.E.; Saratale, G.D. Decolorization and detoxification of sulfonated azo dye CI Remazol Red and textile effluent by isolated Lysinibacillus sp. RGS. J. Biosci. Bioeng. 2013, 115, 658–667. [Google Scholar] [CrossRef]
- Kruger, N.J. The Bradford Method for Protein Quantitation. In The Protein Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 17–24. [Google Scholar]
- Ooi, T.; Shibata, T.; Sato, R.; Ohno, H.; Kinoshita, S.; Thuoc, T.L.; Taguchi, S. An azoreductase, aerobic NADH-dependent flavoprotein discovered from Bacillus sp.: Functional expression and enzymatic characterization. Appl. Microbiol. Biotechnol. 2007, 75, 377–386. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- San Millán, R.M.; Martínez-Ballesteros, I.; Rementeria, A.; Garaizar, J.; Bikandi, J. Online exercise for the design and simulation of PCR and PCR-RFLP experiments. BMC Res. Notes 2013, 6, 513. [Google Scholar] [CrossRef] [Green Version]
- Elbanna, A.H.; Mahrous, E.A.; Khaleel, A.E.; El-Alfy, T.S. Chemical investigation of Bauhinia vahlii Wight and Arnott leaves grown in Egypt. Int. J. Pharm. Pharm. Sci. 2016, 8, 269–272. [Google Scholar]
- Rasheed, T.; Bilal, M.; Iqbal, H.M.; Shah, S.Z.H.; Hu, H.; Zhang, X.; Zhou, Y. TiO2/UV-assisted rhodamine B degradation: Putative pathway and identification of intermediates by UPLC/MS. Environ. Technol. 2018, 39, 1533–1543. [Google Scholar] [CrossRef]
- Imran, M.; Arshad, M.; Negm, F.; Khalid, A.; Shaharoona, B.; Hussain, S.; Nadeem, S.M.; Crowley, D.E. Yeast extract promotes decolorization of azo dyes by stimulating azoreductase activity in Shewanella sp. strain IFN4. Ecotoxicol. Environ. Saf. 2016, 124, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.-S.; Kuo, T.-S.; Chao, Y.-P.; Ho, J.-Y.; Lin, P.-J. Azo dye decolorization with a mutant Escherichia coli strain. Biotechnol. Lett. 2000, 22, 807–812. [Google Scholar] [CrossRef]
- Al-Tohamy, R.; Ali, S.S.; Li, F.; Okasha, K.M.; Mahmoud, Y.A.-G.; Elsamahy, T.; Jiao, H.; Fu, Y.; Sun, J. A critical review on the treatment of dye-containing wastewater: Ecotoxicological and health concerns of textile dyes and possible remediation approaches for environmental safety. Ecotoxicol. Environ. Saf. 2022, 231, 113160. [Google Scholar] [CrossRef] [PubMed]
- Saratale, R.G.; Saratale, G.; Chang, J.; Govindwar, S. Bacterial decolorization and degradation of azo dyes: A review. J. Taiwan Inst. Chem. Eng. 2011, 42, 138–157. [Google Scholar] [CrossRef]
- Ledakowicz, S.; Paździor, K. Recent Achievements in Dyes Removal Focused on Advanced Oxidation Processes Integrated with Biological Methods. Molecules 2021, 26, 870. [Google Scholar] [CrossRef]
- Chen, K.-C.; Wu, J.-Y.; Liou, D.-J.; Hwang, S.-C.J. Decolorization of the textile dyes by newly isolated bacterial strains. J. Biotechnol. 2003, 101, 57–68. [Google Scholar] [CrossRef]
- Theerachat, M.; Guieysse, D.; Morel, S.; Remaud-Siméon, M.; Chulalaksananukul, W. Laccases from marine organisms and their applications in the biodegradation of toxic and environmental pollutants: A review. Appl. Biochem. Biotechnol. 2019, 187, 583–611. [Google Scholar] [CrossRef]
- Liu, G.; Zhou, J.; Wang, J.; Zhang, X.; Dong, B.; Wang, N. Reductive Decolorization of Azo Dye by Bacteria. In Microbial Degradation of Synthetic Dyes in Wastewaters; Singh, S.N., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 111–133. [Google Scholar]
- Uddin, M.S.; Zhou, J.; Qu, Y.; Guo, J.; Wang, P.; Zhao, L. Biodecolorization of azo dye acid red B under high salinity condition. Bull. Environ. Contam. Toxicol. 2007, 79, 440–444. [Google Scholar] [CrossRef]
- Agrawal, S.; Tipre, D.; Patel, B.; Dave, S. Optimization of triazo Acid Black 210 dye degradation by Providencia sp. SRS82 and elucidation of degradation pathway. Process Biochem. 2014, 49, 110–119. [Google Scholar] [CrossRef]
- Ramadan, M.A.; El-Tayeb, O.; Alexander, M. Inoculum size as a factor limiting success of inoculation for biodegradation. Appl. Environ. Microbiol. 1990, 56, 1392–1396. [Google Scholar] [CrossRef] [Green Version]
- Eslami, H.; Shariatifar, A.; Rafiee, E.; Shiranian, M.; Salehi, F.; Hosseini, S.S.; Eslami, G.; Ghanbari, R.; Ebrahimi, A.A. Decolorization and biodegradation of reactive Red 198 Azo dye by a new Enterococcus faecalis–Klebsiella variicola bacterial consortium isolated from textile wastewater sludge. World J. Microbiol. Biotechnol. 2019, 35, 38. [Google Scholar] [CrossRef]
- Khehra, M.S.; Saini, H.S.; Sharma, D.K.; Chadha, B.S.; Chimni, S.S. Comparative studies on potential of consortium and constituent pure bacterial isolates to decolorize azo dyes. Water Res. 2005, 39, 5135–5141. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Bhardwaj, U.; Iqbal, H.M.; Joshi, N. Synergistic role of bacterial consortium to biodegrade toxic dyes containing wastewater and its simultaneous reuse as an added value. Chemosphere 2021, 284, 131273. [Google Scholar] [CrossRef] [PubMed]
- Mendes, S.; Robalo, M.P.; Martins, L.O. Bacterial enzymes and multi-enzymatic systems for cleaning-up dyes from the environment. In Microbial Degradation of Synthetic Dyes in Wastewaters; Springer: Cham, Switzerland, 2015; pp. 27–55. [Google Scholar]
- Shah, M.P.; Patel, K.A.; Nair, S.S.; Darji, A.; Maharaul, S. Microbial degradation of azo dye by Pseudomonas spp. MPS-2 by an application of sequential microaerophilic & aerobic process. Am. J. Microbiol. Res. 2013, 43, 105–112. [Google Scholar]
- Zahran, S.A.; Ali-Tammam, M.; Hashem, A.M.; Aziz, R.K.; Ali, A.E. Azoreductase activity of dye-decolorizing bacteria isolated from the human gut microbiota. Sci. Rep. 2019, 9, 5508. [Google Scholar] [CrossRef] [PubMed]
- Nanjani, S.; Paul, D.; Keharia, H. Genome analysis to decipher syntrophy in the bacterial consortium ‘SCP’for azo dye degradation. BMC Microbiol. 2021, 21, 177. [Google Scholar] [CrossRef]
- Siddique, K.; Shahid, M.; Shahzad, T.; Mahmood, F.; Nadeem, H.; Saif ur Rehman, M.; Hussain, S.; Sadak, O.; Gunasekaran, S.; Kamal, T. Comparative efficacy of biogenic zinc oxide nanoparticles synthesized by Pseudochrobactrum sp. C5 and chemically synthesized zinc oxide nanoparticles for catalytic degradation of dyes and wastewater treatment. Environ. Sci. Pollut. Res. 2021, 28, 28307–28318. [Google Scholar] [CrossRef] [PubMed]
- Al-Shareef, W.A.; Afifi, S.S.; Ramadan, M.A.; Sakr, N.R. Improvement of the Decolorization of Azo Dye (Direct Red 81) by Immobilized Cells of Bacillus pumilus and Free Cells of Aspergillus clavatus in Textile Waste Waters. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1690–1707. [Google Scholar] [CrossRef]
- Rathour, R.; Jain, K.; Madamwar, D.; Desai, C. Microaerophilic biodegradation of raw textile effluent by synergistic activity of bacterial community DR4. J. Environ. Manag. 2019, 250, 109549. [Google Scholar] [CrossRef]
- Ramadan, M.A.; Hashem, A.M.; Al-Shereet, W.A.; Essam, T.M. Decolorization of Reactive black 5 by Micrococcus luteus and Candida albicans in wastewater. World Appl. Sci. 2014, 32, 153–163. [Google Scholar]
- Niessen, W. Thermospray Ionization in Mass Spectrometry. In Encyclopedia of Spectroscopy and Spectrometry, 3rd ed.; Lindon, J.C., Tranter, G.E., Koppenaal, D.W., Eds.; Elsevier Reference Module Chemical, Molecular Sciences and Engineering: Amsterdam, The Netherlands, 2017; Volume 1, pp. 452–457. [Google Scholar]
- Thakur, J.K.; Paul, S.; Dureja, P.; Annapurna, K.; Padaria, J.C.; Gopal, M. Degradation of sulphonated azo dye red HE7B by Bacillus sp. and elucidation of degradative pathways. Curr. Microbiol. 2014, 69, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Paul, J.; Kadam, A.A.; Govindwar, S.P.; Kumar, P.; Varshney, L. An insight into the influence of low dose irradiation pretreatment on the microbial decolouration and degradation of Reactive Red-120 dye. Chemosphere 2013, 90, 1348–1358. [Google Scholar] [CrossRef] [PubMed]
- Palas, B.; Ersöz, G.; Atalay, S. Catalytic wet air oxidation of Reactive Black 5 in the presence of LaNiO3 perovskite catalyst as a green process for azo dye removal. Chemosphere 2018, 209, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Cassano, D.; Zapata, A.; Brunetti, G.; Del Moro, G.; Di Iaconi, C.; Oller, I.; Malato, S.; Mascolo, G. Comparison of several combined/integrated biological-AOPs setups for the treatment of municipal landfill leachate: Minimization of operating costs and effluent toxicity. Chem. Eng. J. 2011, 172, 250–257. [Google Scholar] [CrossRef]
- Guo, G.; Tian, F.; Zhao, Y.; Tang, M.; Liu, W.; Liu, C.; Xue, S.; Kong, W.; Sun, Y.; Wang, S. Aerobic decolorization and detoxification of Acid Scarlet GR by a newly isolated salt-tolerant yeast strain Galactomyces geotrichum GG. Int. Biodeterior. Biodegrad. 2019, 145, 104818. [Google Scholar] [CrossRef]
- Santhanarajan, A.E.; Sul, W.-J.; Yoo, K.-J.; Seong, H.-J.; Kim, H.-G.; Koh, S.-C. Metagenomic Insight of a Full Scale Eco-Friendly Treatment System of Textile Dye Wastewater Using Bioaugmentation of the Composite Culture CES-1. Microorganisms 2021, 9, 1503. [Google Scholar] [CrossRef]
- Santhanarajan, A.-E.; Rhee, C.; Sul, W.J.; Yoo, K.; Seong, H.J.; Kim, H.-G.; Koh, S.-C. Transcriptomic Analysis of Degradative Pathways for Azo Dye Acid Blue 113 in Sphingomonas melonis B-2 from the Dye Wastewater Treatment Process. Microorganisms 2022, 10, 438. [Google Scholar] [CrossRef]








| Enzymes | Time (h) | 27W | 28W | 34W |
|---|---|---|---|---|
| Azoreductase 1 | 0 | 0.056 ± 0.027 | 0.026 ± 0.008 | 0.056 ± 0.032 |
| 2 | 1.39 ± 0.30 ****3 | 1.52 ± 0.42 **** | 1.81 ± 0.40 **** | |
| Laccase 2 | 0 | 0.041 ± 0.049 | 0.063 ± 0.046 | 0.066 ± 0.014 |
| 2 | 0.52 ± 0.13 * | 0.55 ± 0.12 * | 0.77 ± 0.21 *** | |
| Lignin peroxidase 2 | 0 | 0.067 ± 0.038 | 0.087 ± 0.003 | 0.055 ± 0.048 |
| 2 | 0.75 ± 0.32 *** | 0.79 ± 0.14 *** | 0.75 ± 0.30 *** | |
| Veratryl alcohol oxidase 2 | 0 | 0.025 ± 0.029 | 0.064 ± 0.083 | 0.047 ± 0.063 |
| 2 | 0.71 ± 0.34 *** | 0.76 ± 0.34 ***4 | 0.31 ± 0.13 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamal, I.M.; Abdeltawab, N.F.; Ragab, Y.M.; Farag, M.A.; Ramadan, M.A. Biodegradation, Decolorization, and Detoxification of Di-Azo Dye Direct Red 81 by Halotolerant, Alkali-Thermo-Tolerant Bacterial Mixed Cultures. Microorganisms 2022, 10, 994. https://doi.org/10.3390/microorganisms10050994
Kamal IM, Abdeltawab NF, Ragab YM, Farag MA, Ramadan MA. Biodegradation, Decolorization, and Detoxification of Di-Azo Dye Direct Red 81 by Halotolerant, Alkali-Thermo-Tolerant Bacterial Mixed Cultures. Microorganisms. 2022; 10(5):994. https://doi.org/10.3390/microorganisms10050994
Chicago/Turabian StyleKamal, Islam M., Nourtan F. Abdeltawab, Yasser M. Ragab, Mohamed A. Farag, and Mohammed A. Ramadan. 2022. "Biodegradation, Decolorization, and Detoxification of Di-Azo Dye Direct Red 81 by Halotolerant, Alkali-Thermo-Tolerant Bacterial Mixed Cultures" Microorganisms 10, no. 5: 994. https://doi.org/10.3390/microorganisms10050994
APA StyleKamal, I. M., Abdeltawab, N. F., Ragab, Y. M., Farag, M. A., & Ramadan, M. A. (2022). Biodegradation, Decolorization, and Detoxification of Di-Azo Dye Direct Red 81 by Halotolerant, Alkali-Thermo-Tolerant Bacterial Mixed Cultures. Microorganisms, 10(5), 994. https://doi.org/10.3390/microorganisms10050994