
Specific Detection and Enumeration of Burkholderia cepacia Complex by Flow Cytometry Using a Fluorescence-Labeled Oligonucleotide Probe


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Preparation
2.2. Design of the Kef Probe
2.3. RAPID-B Total Plate Count (TPC)
2.4. Optimization of Kef Probe Assay for BCC
2.5. True-Negative Rate (Specificity) in Kef Probe Assay
2.6. Limit of Detection (LOD) in Kef Probe Assay
2.7. Comparison of TPC and Kef Probe Assay
2.7.1. BCC-Spiked 10 μg/mL CHX and 50 μg/mL BZK Samples
2.7.2. Surface Water
3. Results
3.1. Kef Probe Designed for Flow Cytometry
3.2. Optimal Concentration and Temperature of Hybridization Kef Probe
3.3. True-Positive Rate (Sensitivity) and True-Negative Rate (Specificity)
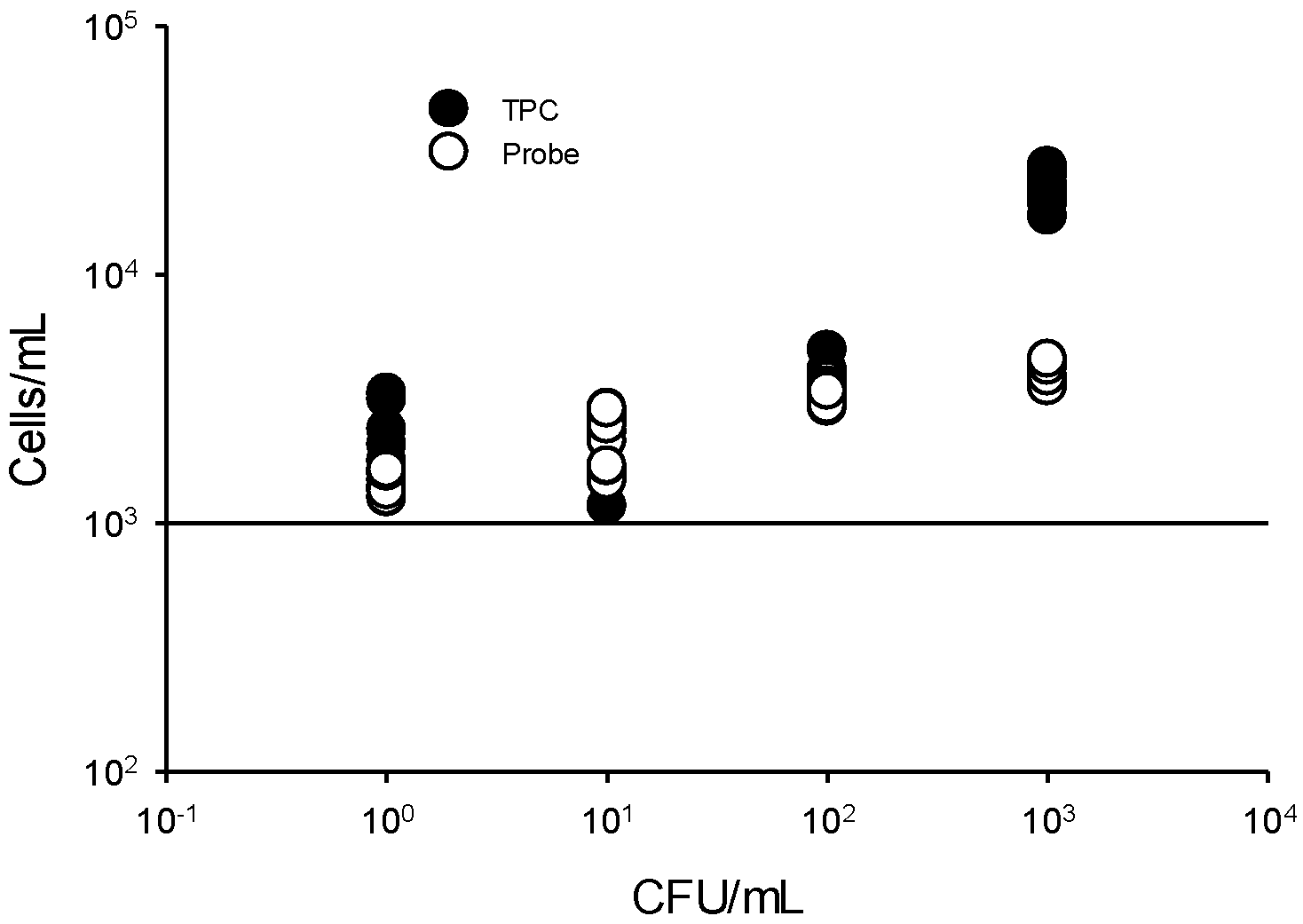
3.4. Limit of Detection (LOD) of the Flow Cytometry Assay
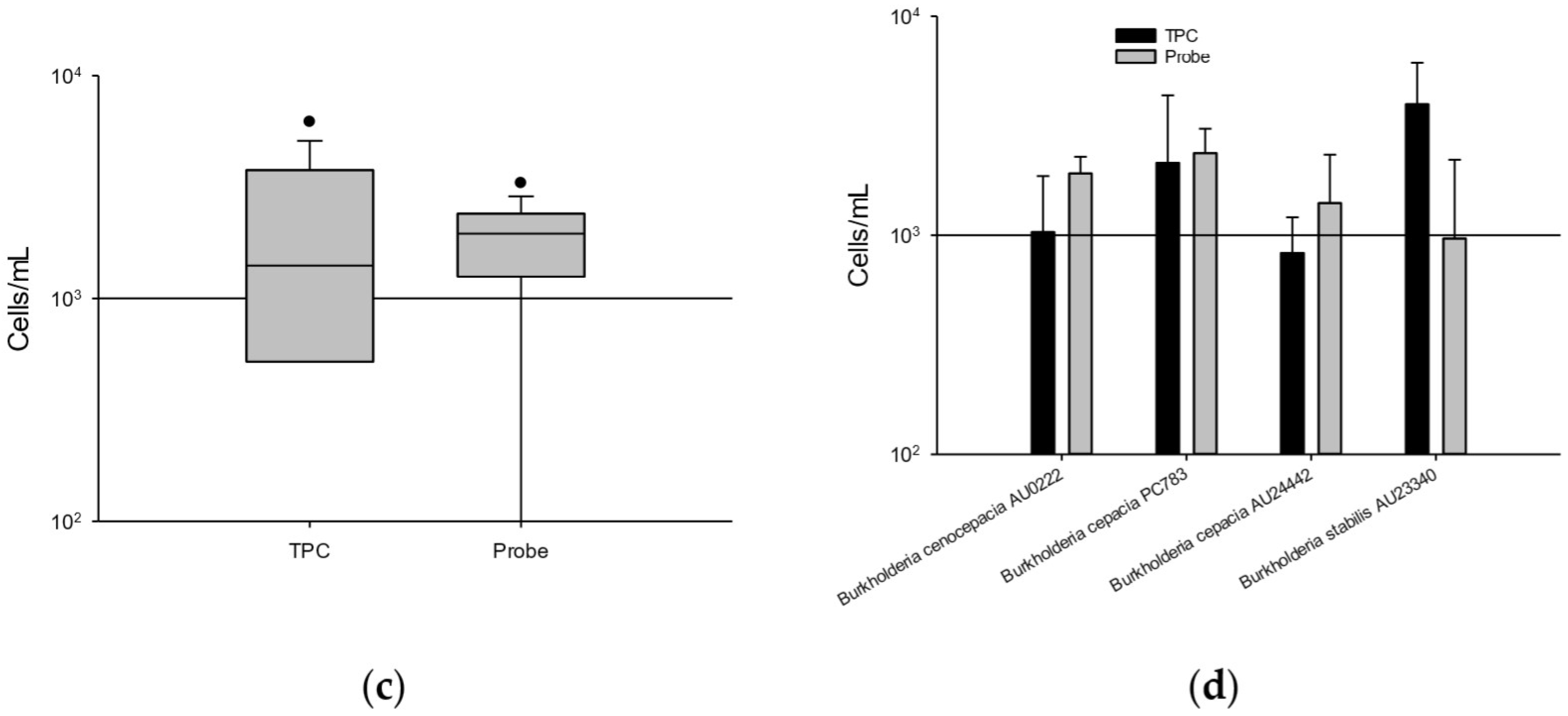
3.5. Counting BCC from Antiseptic Solutions and Surface Water with the Kef Probe Assay
3.5.1. Long-Term Storage in 10 μg/mL CHX and 50 μg/mL BZK
3.5.2. Surface Water
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Depoorter, E.; De Canck, E.; Peeters, C.; Wieme, A.D.; Cnockaert, M.; Zlosnik, J.E.A.; LiPuma, J.J.; Coenye, T.; Vandamme, P. Burkholderia cepacia complex Taxon K: Where to split? Front. Microbiol. 2020, 11, 1594. [Google Scholar] [CrossRef]
- Mahenthiralingam, E.; Urban, T.A.; Goldberg, J.B. The multifarious, multireplicon Burkholderia cepacia complex. Nat. Rev. Microbiol. 2005, 3, 144–156. [Google Scholar] [CrossRef]
- Tavares, M.; Kozak, M.; Balola, A.; Coutinho, C.P.; Godinho, C.P.; Hassan, A.A.; Cooper, V.S.; Sa-Correia, I. Adaptation and survival of Burkholderia cepacia and B. contaminans during long-term incubation in saline solutions containing benzalkonium chloride. Front. Bioeng. Biotechnol. 2020, 8, 630. [Google Scholar] [CrossRef] [PubMed]
- Tavares, M.; Kozak, M.; Balola, A.; Sa-Correia, I. Burkholderia cepacia complex bacteria: A feared contamination risk in water-based pharmaceutical products. Clin. Microbiol. Rev. 2020, 33, e00139-19. [Google Scholar] [CrossRef]
- Ahn, Y.; Kim, J.M.; Kweon, O.; Kim, S.J.; Jones, R.C.; Woodling, K.; Gamboa da Costa, G.; LiPuma, J.J.; Hussong, D.; Marasa, B.S.; et al. Intrinsic resistance of Burkholderia cepacia complex to benzalkonium chloride. MBio 2016, 7, e01716-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos Saalfeld, S.M.; Shinohara, D.R.; Dos Anjos Szczerepa, M.M.; Martinez, H.V.; Vieira de Campos, E.; Mitsugui, C.S.; Oliveira, A.R.; Granzotto, D.C.T.; Nishiyama, S.A.B.; Tognim, M.C.B. Consecutive outbreaks of Burkholderia cepacia complex caused by intrinsically contaminated chlorhexidine mouthwashes. Am. J. Infect. Control 2020, 48, 1348–1353. [Google Scholar] [CrossRef]
- Sutton, S.; Jimenez, L. A review of reported recalls involving microbiological control 2004–2011 with emphasis on FDA considerations of “objectionable organisms”. Am. Pharm. Rev. 2012, 15, 42–57. [Google Scholar]
- Jimenez, L. Microbial diversity in pharmaceutical product recalls and environments. PDA J. Pharm. Sci. Technol. 2007, 61, 383–399. [Google Scholar]
- David, J.; Bell, R.E.; Clark, G.C. Mechanisms of disease: Host-pathogen interactions between Burkholderia species and lung epithelial cells. Front. Cell. Infect. Microbiol. 2015, 5, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- USP. <61> Microbiological Examination of Nonsterile Products—Microbial Enumeration Tests. 2016. Available online: https://www.usp.org/sites/default/files/usp/document/harmonization/gen-method/q05b_pf_ira_34_6_2008.pdf (accessed on 1 March 2022).
- USP. <1111> Microbiological Examination of Non-Sterile Products: Acceptance Criteria for Pharmaceutical Preparations and Substances for Pharmaceutical Use. 2016. Available online: https://www.usp.org/sites/default/files/usp/document/harmonization/gen-method/q05c_pf_ira_33_2_2007.pdf (accessed on 1 March 2022).
- USP. <60> Microbiological Examination of Nonsterile Products—Tests for Burkholderia cepacia Complex. 2018. Available online: http://www.usppf.com/pf/pub/data/v445/CHA_IPR_445_c60.xml (accessed on 1 March 2022).
- Attia, M.A.; Ali, A.E.; Essam, T.M.; Amin, M.A. Direct detection of Burkholderia cepacia in susceptible pharmaceutical products using semi-nested PCR. PDA J. Pharm. Sci. Technol. 2016, 70, 99–108. [Google Scholar] [CrossRef]
- Ahn, Y.; Gibson, B.; Williams, A.; Alusta, P.; Buzatu, D.A.; Lee, Y.J.; LiPuma, J.J.; Hussong, D.; Marasa, B.; Cerniglia, C.E. A comparison of culture-based, real-time PCR, droplet digital PCR and flow cytometric methods for the detection of Burkholderia cepacia complex in nuclease-free water and antiseptics. J. Ind. Microbiol. Biotechnol. 2020, 47, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Mahenthiralingam, E.; Bischof, J.; Byrne, S.K.; Radomski, C.; Davies, J.E.; Av-Gay, Y.; Vandamme, P. DNA-Based diagnostic approaches for identification of Burkholderia cepacia complex, Burkholderia vietnamiensis, Burkholderia multivorans, Burkholderia stabilis, and Burkholderia cepacia genomovars I and III. J. Clin. Microbiol. 2000, 38, 3165–3173. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, L.; Jashari, T.; Vasquez, J.; Zapata, S.; Bochis, J.; Kulko, M.; Ellman, V.; Gardner, M.; Choe, T. Real-time PCR detection of Burkholderia cepacia in pharmaceutical products contaminated with low levels of bacterial contamination. PDA J. Pharm. Sci. Technol. 2018, 72, 73–80. [Google Scholar] [CrossRef]
- Furlan, J.P.R.; Pitondo-Silva, A.; Braz, V.S.; Gallo, I.F.L.; Stehling, E.G. Evaluation of different molecular and phenotypic methods for identification of environmental Burkholderia cepacia complex. World J. Microbiol. Biotechnol. 2019, 35, 39. [Google Scholar] [CrossRef]
- Daddy Gaoh, S.; Kweon, O.; Lee, Y.-J.; LiPuma, J.J.; Hussong, D.; Marasa, B.; Ahn, Y. Loop-mediated isothermal amplification (LAMP) assay for detecting Burkholderia cepacia complex in non-sterile pharmaceutical products. Pathogens 2021, 10, 1071. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhong, Q.; Shang, M.Y.; Li, M.; Jiang, Y.S.; Zou, J.J.; Ma, S.S.; Huang, Q.; Lu, W.P. Preliminary evaluation of rapid visual identification of Burkholderia pseudomallei using a newly developed lateral flow strip-based recombinase polymerase amplification (LF-RPA) system. Front. Cell. Infect. Microbiol. 2021, 11, 804737. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Zheng, X.; Kan, B.; Li, W.; Zhang, W.; Jiang, T.; Lu, J.; Qin, A. Rapid detection of Burkholderia pseudomallei with a lateral flow recombinase polymerase amplification assay. PLoS ONE 2019, 14, e0213416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buzatu, D.A.; Cooper, W.M.; Summage-West, C.; Sutherland, J.B.; Williams, A.J.; Bass, D.A.; Smith, L.L.; Woodruff, R.S.; Christman, J.M.; Reid, S.; et al. Photobleaching with phloxine B sensitizer to reduce food matrix interference for detection of Escherichia coli serotype O157:H7 in fresh spinach by flow cytometry. Food Microbiol. 2013, 36, 416–425. [Google Scholar] [CrossRef] [Green Version]
- Wilkes, J.G.; Tucker, R.K.; Montgomery, J.A.; Cooper, W.M.; Sutherland, J.B.; Buzatu, D.A. Reduction of food matrix interference by a combination of sample preparation and multi-dimensional gating techniques to facilitate rapid, high sensitivity analysis for Escherichia coli serotype O157 by flow cytometry. Food Microbiol. 2012, 30, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.J.; Cooper, W.M.; Ramsaroop, S.; Alusta, P.; Buzatu, D.A.; Wilkes, J.G. Rapid flow cytometry detection of a single viable Escherichia coli O157:H7 cell in raw spinach using a simplified sample preparation technique. Front. Microbiol. 2017, 8, 1493. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.J.; Cooper, W.M.; Summage-West, C.V.; Sims, L.M.; Woodruff, R.; Christman, J.; Moskal, T.J.; Ramsaroop, S.; Sutherland, J.B.; Alusta, P.; et al. Level 2 validation of a flow cytometric method for detection of Escherichia coli O157:H7 in raw spinach. Int. J. Food Microbiol. 2015, 215, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Y.; Wilkes, J.G.; Moskal, T.J.; Williams, A.J.; Cooper, W.M.; Nayak, R.; Rafii, F.; Buzatu, D.A. Development of a flow cytometry-based method for rapid detection of Escherichia coli and Shigella spp. using an oligonucleotide probe. PLoS ONE 2016, 11, e0150038. [Google Scholar] [CrossRef]
- Amann, R.I.; Binder, B.J.; Olson, R.J.; Chisholm, S.W.; Devereux, R.; Stahl, D.A. Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl. Env. Microbiol. 1990, 56, 1919–1925. [Google Scholar] [CrossRef] [Green Version]
- Joachimsthal, E.L.; Ivanov, V.; Tay, S.T.L.; Tay, J.H. Quantification of whole-cell in situ hybridization with oligonucleotide probes by flow cytometry of Escherichia coli cells. World J. Microbiol. Biotechnol. 2003, 19, 527–533. [Google Scholar] [CrossRef]
- Panchuk-Voloshina, N.; Haugland, R.P.; Bishop-Stewart, J.; Bhalgat, M.K.; Millard, P.J.; Mao, F.; Leung, W.Y.; Haugland, R.P. Alexa dyes, a series of new fluorescent dyes that yield exceptionally bright, photostable conjugates. J. Histochem. Cytochem. 1999, 47, 1179–1188. [Google Scholar] [CrossRef]
- FDA. Statistical Guidance on Reporting Results from Studies Evaluating Diagnostic Tests—Guidance for Industry and FDA Staff. 2007. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/statistical-guidance-reporting-results-studies-evaluating-diagnostic-tests-guidance-industry-and-fda (accessed on 1 March 2022).
- Ahn, Y.; Kim, J.M.; Ahn, H.; Lee, Y.J.; LiPuma, J.J.; Hussong, D.; Cerniglia, C.E. Evaluation of liquid and solid culture media for the recovery and enrichment of Burkholderia cenocepacia from distilled water. J. Ind. Microbiol. Biotechnol. 2014, 41, 1109–1118. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, S.J.; DeLong, E.F.; Olsen, G.J.; Pace, N.R. Phylogenetic group-specific oligodeoxynucleotide probes for identification of single microbial cells. J. Bacteriol. 1988, 170, 720–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLong, E.F.; Wickham, G.S.; Pace, N.R. Phylogenetic stains: Ribosomal RNA-based probes for the identification of single cells. Science 1989, 243, 1360–1363. [Google Scholar] [CrossRef] [PubMed]
- Czechowska, K.; Johnson, D.R.; van der Meer, J.R. Use of flow cytometric methods for single-cell analysis in environmental microbiology. Curr. Opin. Microbiol. 2008, 11, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Frossard, A.; Hammes, F.; Gessner, M.O. Flow cytometric assessment of bacterial abundance in soils, sediments and sludge. Front. Microbiol. 2016, 7, 903. [Google Scholar] [CrossRef]
- Joux, F.; Lebaron, P. Use of fluorescent probes to assess physiological functions of bacteria at single-cell level. Microbes Infect. 2000, 2, 1523–1535. [Google Scholar] [CrossRef]
- Ahn, Y.; Lee, U.J.; Lee, Y.J.; LiPuma, J.J.; Hussong, D.; Marasa, B.; Cerniglia, C.E. Oligotrophic media compared with a tryptic soy agar or broth for the recovery of Burkholderia cepacia complex from different storage temperatures and culture conditions. J. Microbiol. Biotechnol. 2019, 29, 1495–1505. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Ahn, Y.; LiPuma, J.J.; Hussong, D.; Cerniglia, C.E. Survival and susceptibility of Burkholderia cepacia complex in chlorhexidine gluconate and benzalkonium chloride. J. Ind. Microbiol. Biotechnol. 2015, 42, 905–913. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daddy Gaoh, S.; Williams, A.; Le, D.; Kweon, O.; Alusta, P.; Buzatu, D.A.; Ahn, Y. Specific Detection and Enumeration of Burkholderia cepacia Complex by Flow Cytometry Using a Fluorescence-Labeled Oligonucleotide Probe. Microorganisms 2022, 10, 1170. https://doi.org/10.3390/microorganisms10061170
Daddy Gaoh S, Williams A, Le D, Kweon O, Alusta P, Buzatu DA, Ahn Y. Specific Detection and Enumeration of Burkholderia cepacia Complex by Flow Cytometry Using a Fluorescence-Labeled Oligonucleotide Probe. Microorganisms. 2022; 10(6):1170. https://doi.org/10.3390/microorganisms10061170
Chicago/Turabian StyleDaddy Gaoh, Soumana, Anna Williams, David Le, Ohgew Kweon, Pierre Alusta, Dan A. Buzatu, and Youngbeom Ahn. 2022. "Specific Detection and Enumeration of Burkholderia cepacia Complex by Flow Cytometry Using a Fluorescence-Labeled Oligonucleotide Probe" Microorganisms 10, no. 6: 1170. https://doi.org/10.3390/microorganisms10061170
APA StyleDaddy Gaoh, S., Williams, A., Le, D., Kweon, O., Alusta, P., Buzatu, D. A., & Ahn, Y. (2022). Specific Detection and Enumeration of Burkholderia cepacia Complex by Flow Cytometry Using a Fluorescence-Labeled Oligonucleotide Probe. Microorganisms, 10(6), 1170. https://doi.org/10.3390/microorganisms10061170