
Pep27 Mutant Immunization Inhibits Caspase-14 Expression to Alleviate Inflammatory Bowel Disease via Treg Upregulation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Induction of Colitis and Immunization Protocol
2.3. Macroscopic Assessment of Colitis Severity
2.4. Histopathological Analysis
2.5. Terminal Deoxynucleotidyl Transferase-Mediated dUTP Nick-End Labeling Assay
2.6. Assessment of Myeloperoxidase (MPO) Activity and Malondialdehyde (MDA) Level
2.7. Permeability Assay
2.8. Gut Microbiome Analysis in Fecal Samples
2.9. Treg and Caspase-14 Neutralization
2.10. Protein Extraction and Western Blotting
2.11. cDNA Synthesis and Real-Time PCR
2.12. High-Throughput Sequencing and Transcriptomic Analysis
2.13. Statistical Analysis
2.14. Ethical Considerations
3. Results
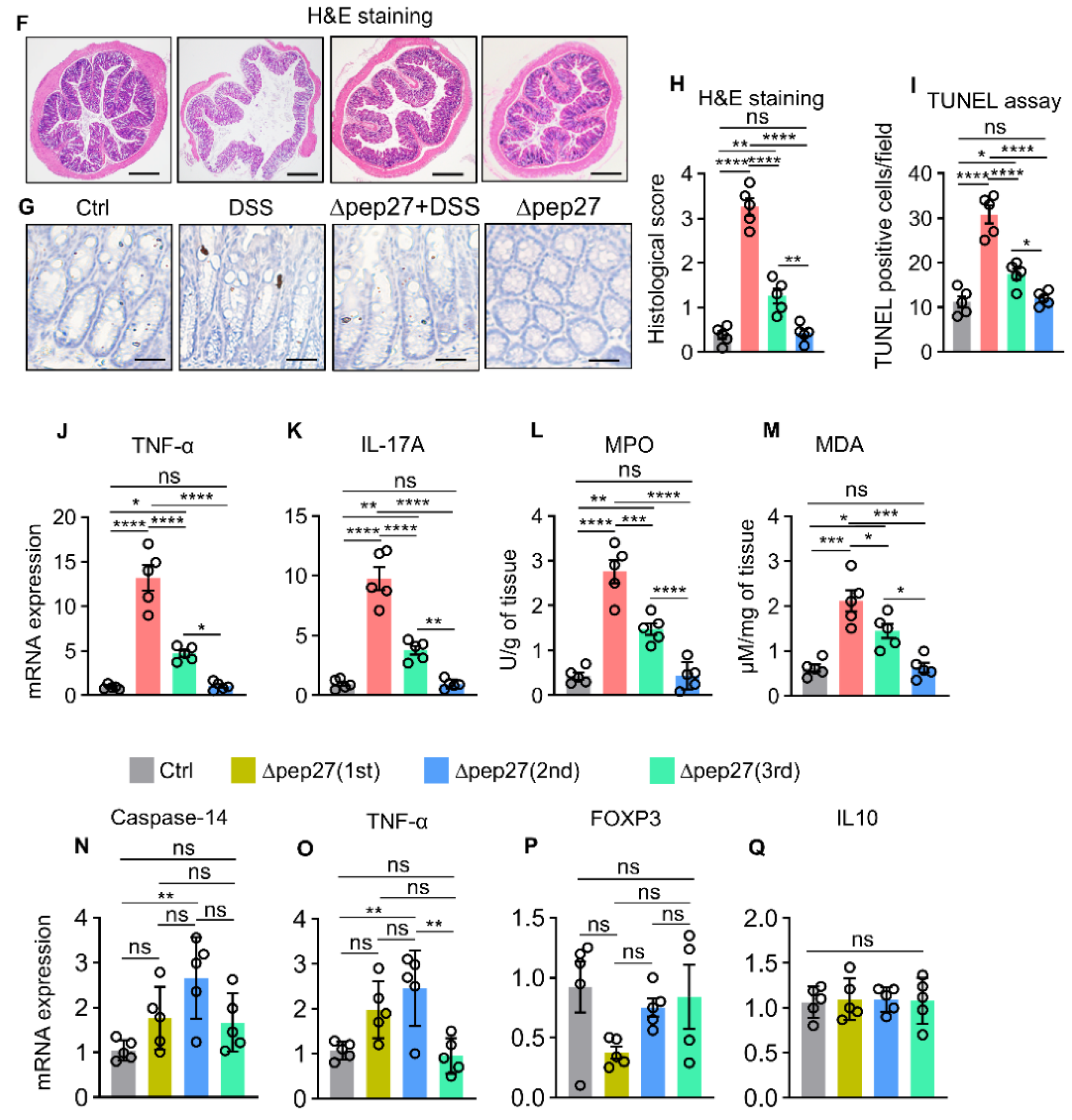
3.1. ∆Pep27 Immunization Attenuated Proinflammatory Cytokines and Oxidative Stress in the DIC
3.2. ∆Pep27 Immunization Diminished Proinflammatory Cytokines, and Caspase-14 Expression via Treg Upregulation to Prevent Barrier Disruption
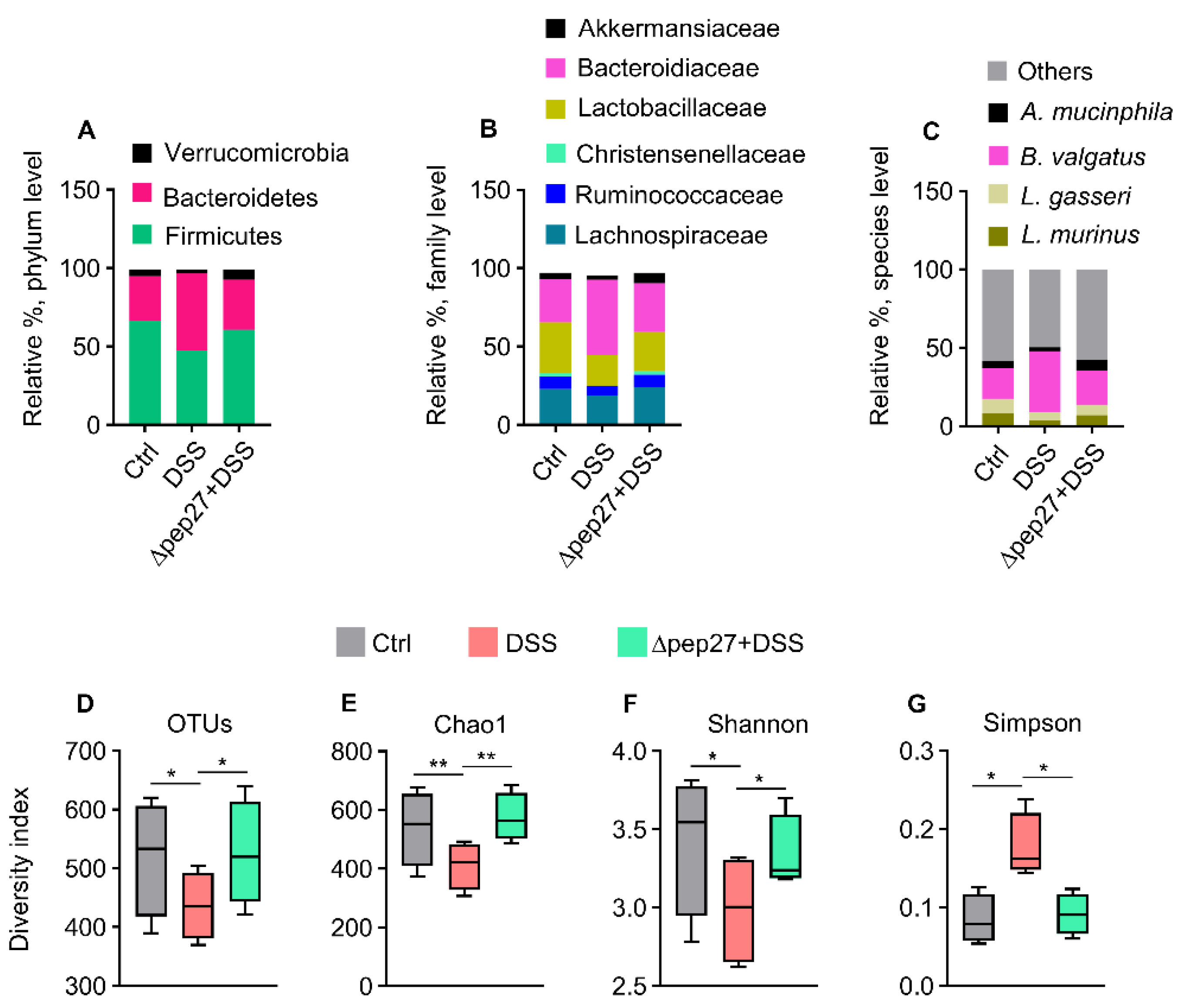
3.3. DIC-perturbed Biodiversity of Gut Microbiota Is Restored by ∆Pep27
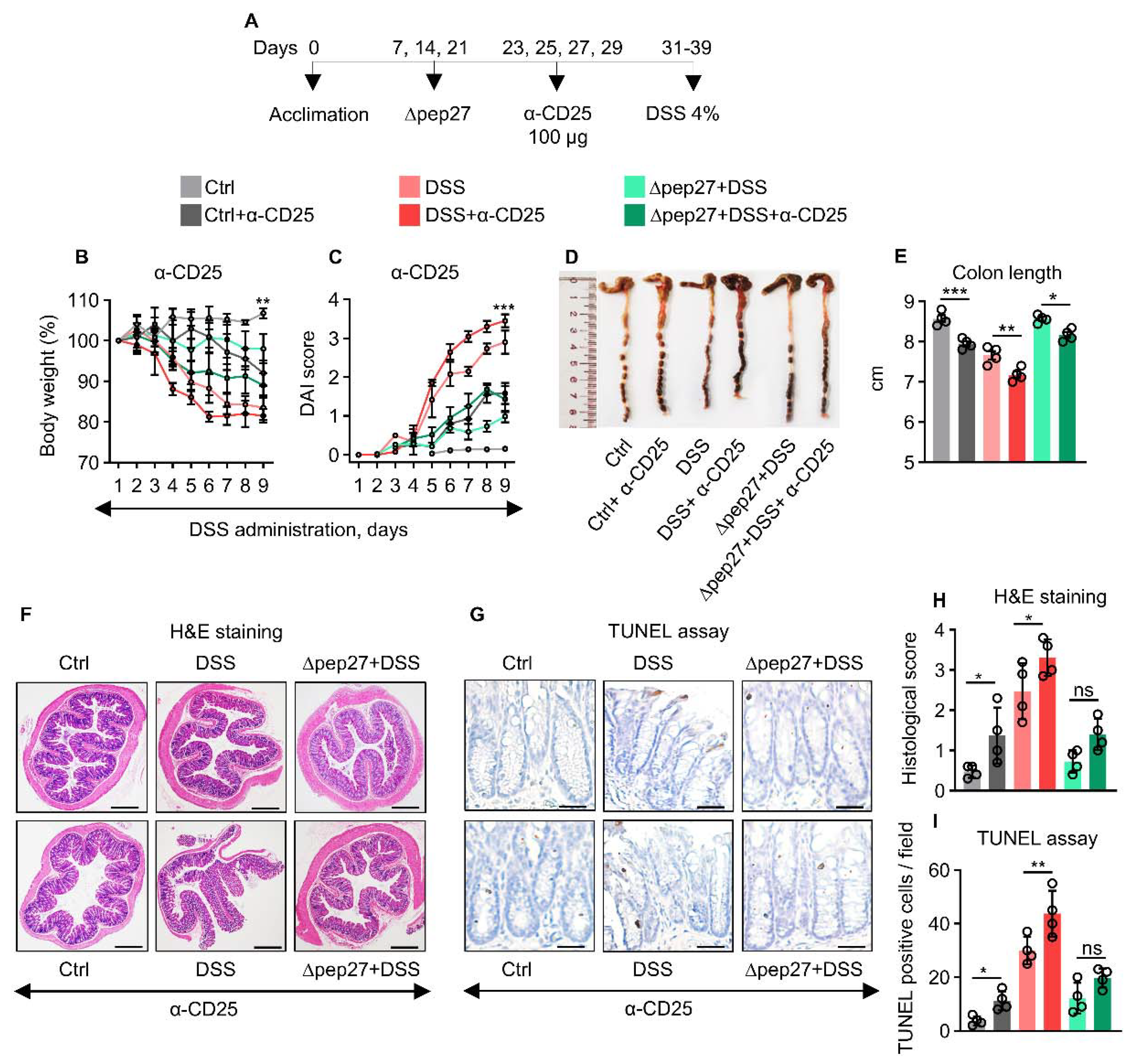
3.4. Treg Depletion Is Restored by ∆Pep27 to Prevent Inflammation and Apoptosis
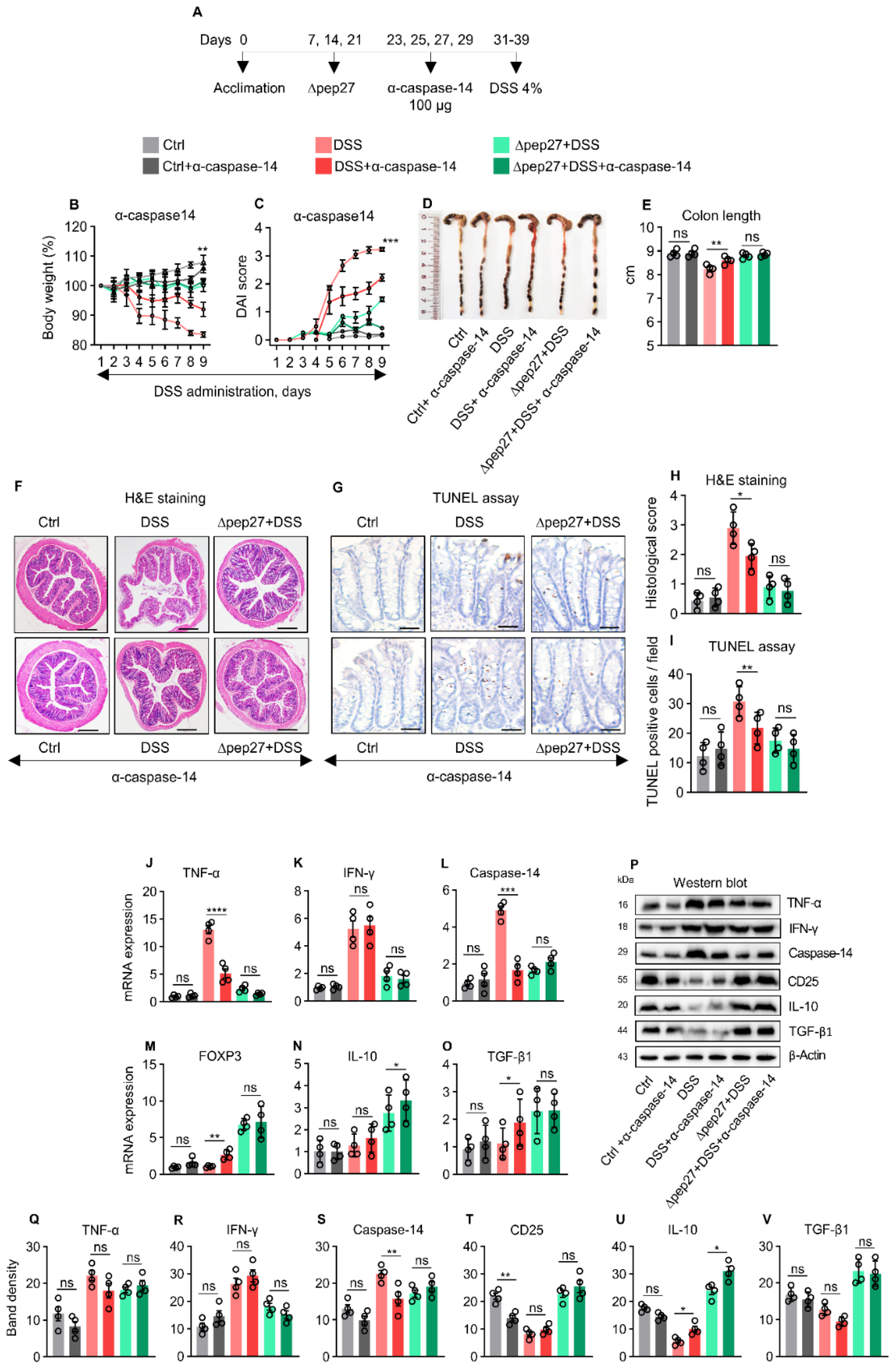
3.5. Caspase-14 Depletion Uncovers Proinflammatory and Apoptotic Nature of Caspase-14
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Neurath, M.F. Cytokines in inflammatory bowel disease. Nat. Rev. Immunol. 2014, 14, 329–342. [Google Scholar] [CrossRef] [PubMed]
- de Souza, H.S.; Fiocchi, C. Immunopathogenesis of IBD: Current state of the art. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.T. Pathophysiology of Inflammatory bowel diseases. N. Engl. J. Med. 2020, 383, 2652–2664. [Google Scholar] [CrossRef] [PubMed]
- Quetglas, E.G.; Armuzzi, A.; Wigge, S.; Fiorino, G.; Barnscheid, L.; Froelich, M.; Danese, S. The pharmacokinetics and pharmacodynamics of drugs used in inflammatory bowel disease treatment. Eur. J. Clin. Pharmacol. 2015, 71, 773–799. [Google Scholar] [CrossRef]
- Fujino, S.; Andoh, A.; Bamba, S.; Ogawa, A.; Hata, K.; Araki, Y.; Bamba, T.; Fujiyama, Y. Increased expression of interleukin 17 in inflammatory bowel disease. Gut 2003, 52, 65–70. [Google Scholar] [CrossRef]
- Huber, S.; Gagliani, N.; Esplugues, E.; O’Connor, W., Jr.; Huber, F.J.; Chaudhry, A.; Kamanaka, M.; Kobayashi, Y.; Booth, C.J.; Rudensky, A.Y.; et al. Th17 cells express interleukin-10 receptor and are controlled by Foxp3− and Foxp3+ regulatory CD4+ T cells in an interleukin-10-dependent manner. Immunity 2011, 34, 554–565. [Google Scholar] [CrossRef]
- Ueno, A.; Jeffery, L.; Kobayashi, T.; Hibi, T.; Ghosh, S.; Jijon, H. Th17 plasticity and its relevance to inflammatory bowel disease. J. Autoimmun. 2018, 87, 38–49. [Google Scholar] [CrossRef]
- Himmel, M.E.; Yao, Y.; Orban, P.C.; Steiner, T.S.; Levings, M.K. Regulatory T-cell therapy for inflammatory bowel disease: More questions than answers. Immunology 2012, 136, 115–122. [Google Scholar] [CrossRef]
- Caruso, R.; Lo, B.C.; Nunez, G. Host-microbiota interactions in inflammatory bowel disease. Nat. Rev. Immunol. 2020, 20, 411–426. [Google Scholar] [CrossRef]
- Basso, P.J.; Câmara, N.O.S.; Sales-Campos, H. Microbial-Based Therapies in the Treatment of Inflammatory Bowel Disease—An Overview of Human Studies. Front. Pharmacol. 2019, 9, 1571. [Google Scholar] [CrossRef] [Green Version]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Ooijevaar, R.E.; Terveer, E.M.; Verspaget, H.W.; Kuijper, E.J.; Keller, J.J. Clinical Application and Potential of Fecal Microbiota Transplantation. Annu. Rev. Med. 2019, 70, 335–351. [Google Scholar] [CrossRef] [PubMed]
- Kurashima, Y.; Kiyono, H. Mucosal ecological network of epithelium and immune cells for gut homeostasis and tissue healing. Annu. Rev. Immunol. 2017, 35, 119–147. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Tran, T.D.; Briles, D.E.; Rhee, D.K. Inactivated pep27 mutant as an effective mucosal vaccine against a secondary lethal pneumococcal challenge in mice. Clin. Exp. Vaccine Res. 2013, 2, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.; Choi, S.Y.; Kwon, M.K.; Tran, T.D.; Park, S.S.; Lee, K.J.; Bae, S.M.; Briles, D.E.; Rhee, D.K. Streptococcus pneumoniae pep27 mutant as a live vaccine for serotype-independent protection in mice. Vaccine 2012, 30, 2008–2019. [Google Scholar] [CrossRef]
- Kim, G.L.; Choi, S.Y.; Seon, S.H.; Lee, S.; Park, S.S.; Song, J.Y.; Briles, D.E.; Rhee, D.K. Pneumococcal pep27 mutant immunization stimulates cytokine secretion and confers long-term immunity with a wide range of protection, including against non-typeable strains. Vaccine 2016, 34, 6481–6492. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.L.; Lee, S.; Kim, S.J.; Lee, S.O.; Pyo, S.; Rhee, D.K. Pulmonary Colonization Resistance to Pathogens via Noncanonical Wnt and Interleukin-17A by Intranasal pep27 Mutant Immunization. J. Infect. Dis. 2018, 217, 1977–1986. [Google Scholar] [CrossRef]
- Seon, S.H.; Choi, J.A.; Yang, E.; Pyo, S.; Song, M.K.; Rhee, D.K. Intranasal immunization with an attenuated pep27 mutant provides protection from Influenza virus and secondary pneumococcal infections. J. Infect. Dis. 2018, 217, 637–640. [Google Scholar] [CrossRef]
- Kim, B.G.; Ghosh, P.; Ahn, S.; Rhee, D.K. Pneumococcal pep27 mutant immunization suppresses allergic asthma in mice. Biochem. Biophys. Res. Commun. 2019, 514, 210–216. [Google Scholar] [CrossRef]
- Iqbal, H.; Kim, G.L.; Kim, J.H.; Ghosh, P.; Shah, M.; Lee, W.; Rhee, D.K. Pneumococcal pep27-mutant inhibits Wnt5a expression via the regulation of T helper cells to attenuate colitis. Int. Immunopharmacol. 2022, 109, 108927. [Google Scholar] [CrossRef]
- Geboes, K.; Riddell, R.; Ost, A.; Jensfelt, B.; Persson, T.; Lofberg, R. A reproducible grading scale for histological assessment of inflammation in ulcerative colitis. Gut 2000, 47, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Yi, T.G.; Park, J.M.; Han, Y.M.; Kim, J.H.; Shin, D.H.; Tak, S.J.; Lee, K.; Lee, Y.S.; Jeon, M.S.; et al. Therapeutic effects of mouse bone marrow-derived clonal mesenchymal stem cells in a mouse model of inflammatory bowel disease. J. Clin. Biochem. Nutr. 2015, 57, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, H.; Kim, S.K.; Cha, K.M.; Jeong, M.S.; Ghosh, P.; Rhee, D.K. Korean Red Ginseng alleviates neuroinflammation and promotes cell survival in the intermittent heat stress-induced rat brain by suppressing oxidative stress via estrogen receptor beta and brain-derived neurotrophic factor upregulation. J. Ginseng Res. 2020, 44, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Radu, M.; Chernoff, J. An in vivo assay to test blood vessel permeability. J. Vis. Exp. 2013, 73, e50062. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Kim, K.Y.; Lee, J.H.; Choi, Y. The analysis of oral microbial communities of wild-type and toll-like receptor 2-deficient mice using a 454 GS FLX Titanium pyrosequencer. BMC Microbiol. 2010, 10, 101. [Google Scholar] [CrossRef]
- Ishikawa, D.; Okazawa, A.; Corridoni, D.; Jia, L.G.; Wang, X.M.; Guanzon, M.; Xin, W.; Arseneau, K.O.; Pizarro, T.T.; Cominelli, F. Tregs are dysfunctional in vivo in a spontaneous murine model of Crohn’s disease. Mucosal Immunol. 2013, 6, 267–275. [Google Scholar] [CrossRef]
- Lee, K.H.; Park, M.; Ji, K.Y.; Lee, H.Y.; Jang, J.H.; Yoon, I.J.; Oh, S.S.; Kim, S.M.; Jeong, Y.H.; Yun, C.H.; et al. Bacterial β-(1,3)-glucan prevents DSS-induced IBD by restoring the reduced population of regulatory T cells. Immunobiology 2014, 219, 802–812. [Google Scholar] [CrossRef]
- Ni, J.; Wu, G.D.; Albenberg, L.; Tomov, V.T. Gut microbiota and IBD: Causation or correlation? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 573–584. [Google Scholar] [CrossRef]
- Green, D.R.; Llambi, F. Cell death signalling. Cold Spring Harb. Perspect Biol. 2015, 7, a006080. [Google Scholar] [CrossRef]
- Sahin, A.; Ilhan, A.; Derin, M.E.; Dogan, H.O.; Sahin, M. Low levels of pannexin-1 in Behcet’s syndrome. Int. J. Rheum. Dis. 2019, 22, 1474–1478. [Google Scholar] [CrossRef]
- Solomon, I.; Amann, M.; Goubier, A.; Vargas, F.A.; Zervas, D.; Qing, C.; Henry, J.Y.; Ghorani, E.; Akarca, A.U.; Marafioti, T.; et al. CD25-Treg-depleting antibodies preserving IL-2 signaling on effector T cells enhance effector activation and antitumor immunity. Nat. Cancer 2020, 1, 1153–1166. [Google Scholar] [CrossRef] [PubMed]
- Nava, P.; Koch, S.; Laukoetter, M.G.; Lee, W.Y.; Kolegraff, K.; Capaldo, C.T.; Beeman, N.; Addis, C.; Gerner-Smidt, K.; Neumaier, I.; et al. Interferon-gamma regulates intestinal epithelial homeostasis through converging beta-catenin signaling pathways. Immunity 2010, 32, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Ghosh, P.; Kwon, H.; Park, S.S.; Kim, G.L.; Choi, S.Y.; Kim, E.H.; Tran, T.D.; Seon, S.H.; Le, N.T.; et al. Induction of the pneumococcal vncRS operon by lactoferrin is essential for pneumonia. Virulence 2018, 9, 1562–1575. [Google Scholar] [CrossRef]
- Vermeij, W.P.; Florea, B.I.; Isenia, S.; Alia, A.; Brouwer, J.; Backendorf, C. Proteomic identification of in vivo interactors reveals novel function of skin cornification proteins. J. Proteome Res. 2012, 11, 3068–3076. [Google Scholar] [CrossRef] [PubMed]
- Vermeij, W.P.; Backendorf, C. Skin cornification proteins provide global link between ROS detoxification and cell migration during wound healing. PLoS ONE 2010, 5, e11957. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.O.; Lee, S.; Kim, S.J.; Rhee, D.K. Korean Red Ginseng enhances pneumococcal Δpep27 vaccine efficacy by inhibiting reactive oxygen species production. J. Ginseng Res. 2019, 43, 218–225. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y.; et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef]
- Lindenberg, F.; Krych, L.; Fielden, J.; Kot, W.; Frøkiær, H.; van Galen, G.; Nielsen, D.S.; Hansen, A.K. Expression of immune regulatory genes correlate with the abundance of specific Clostridiales and Verrucomicrobia species in the equine ileum and cecum. Sci. Rep. 2019, 9, 12674. [Google Scholar] [CrossRef]
- Dou, X.; Gao, N.; Yan, D.; Shan, A. Sodium Butyrate Alleviates Mouse Colitis by Regulating Gut Microbiota Dysbiosis. Animals 2020, 10, 1154. [Google Scholar] [CrossRef]
- Yan, S.; Yang, B.; Zhao, J.; Zhao, J.; Stanton, C.; Ross, R.P.; Zhang, H.; Chen, W. A ropy exopolysaccharide producing strain Bifidobacterium longum subsp. longum YS108R alleviates DSS-induced colitis by maintenance of the mucosal barrier and gut microbiota modulation. Food Funct. 2019, 10, 1595–1608. [Google Scholar] [CrossRef]
- Schwerk, C.; Schulze-Osthoff, K. Non-apoptotic functions of caspases in cellular proliferation and differentiation. Biochem. Pharmacol. 2003, 66, 1453–1458. [Google Scholar] [CrossRef]
- Denecker, G.; Ovaere, P.; Vandenabeele, P.; Declercq, W. Caspase-14 reveals its secrets. J. Cell Biol. 2008, 180, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Lin, H.; Deng, X.; Li, S.; Zhang, J. MiR-1246 promotes anti-apoptotic effect of mini-αA in oxidative stress-induced apoptosis in retinal pigment epithelial cells. Clin. Exp. Ophthalmol. 2020, 48, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.; Suresh, P.K. Enhanced Induction of Apoptosis in HaCaT Cells by Luteolin Encapsulated in PEGylated Liposomes-Role of Caspase-3/Caspase-14. Appl. Biochem. Biotechnol. 2019, 188, 147–164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; He, A.; Liu, S.; He, Q.; Luo, Y.; He, Z.; Chen, Y.; Tao, A.; Yan, J. Inhibition of HtrA2 alleviated dextran sulfate sodium (DSS)-induced colitis by preventing necroptosis of intestinal epithelial cells. Cell Death Dis. 2019, 10, 344. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence (5′ to 3′) | |
|---|---|---|
| Caspase-14 | F | GATGAGGTTGCTGTGCTCAA |
| R | TTGGTTTTCCTTCCTGCATC | |
| Claudin-1 | F | GATGTGGATGGCTGTCATTG |
| R | CCACTAATGTCGCCAGACCT | |
| FOXP3 | F | CCTGCCTTGGTACATTCGTG |
| R | TGTTGTGGGTGAGTGCTTTG | |
| IFN-γ | F | ACAGCTCTCCGTCCTCGTAT |
| R | CCTTAGAGTAGAAAGACCG | |
| IL-10 | F | AGCCACATGCTCCTAGAGC |
| R | GCCTGGGGCATCACTTCTAC | |
| IL-17A | F | TCCAGAAGGCCCTCAGACTA |
| R | AGCATCTTCTCGACCCTGAA | |
| TGF-β1 | F | ACCATGCCAACTTCTGTCTG |
| R | CGGGTTGTGTTGGTTGTAGA | |
| TNF-α | F | GCC TCTTCTCATTCCTGCTT |
| R | TGGGAACTTCTCATCCCT TTG | |
| GAPDH | F | TCAACAGCAACTCCCACTCTTCCA |
| R | ACCCTGTTGCTGTAGCCGTATTCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iqbal, H.; Kim, G.-L.; Kim, J.-H.; Ghosh, P.; Shah, M.; Lee, W.; Rhee, D.-K. Pep27 Mutant Immunization Inhibits Caspase-14 Expression to Alleviate Inflammatory Bowel Disease via Treg Upregulation. Microorganisms 2022, 10, 1871. https://doi.org/10.3390/microorganisms10091871
Iqbal H, Kim G-L, Kim J-H, Ghosh P, Shah M, Lee W, Rhee D-K. Pep27 Mutant Immunization Inhibits Caspase-14 Expression to Alleviate Inflammatory Bowel Disease via Treg Upregulation. Microorganisms. 2022; 10(9):1871. https://doi.org/10.3390/microorganisms10091871
Chicago/Turabian StyleIqbal, Hamid, Gyu-Lee Kim, Ji-Hoon Kim, Prachetash Ghosh, Masaud Shah, Wonsik Lee, and Dong-Kwon Rhee. 2022. "Pep27 Mutant Immunization Inhibits Caspase-14 Expression to Alleviate Inflammatory Bowel Disease via Treg Upregulation" Microorganisms 10, no. 9: 1871. https://doi.org/10.3390/microorganisms10091871