
Biochemical, Catabolic, and PGP Activity of Microbial Communities and Bacterial Strains from the Root Zone of Baccharis linearis in a Mediterranean Mine Tailing

 , , , , and
, , , , and
Abstract
:
1. Introduction
2. Materials and Methods
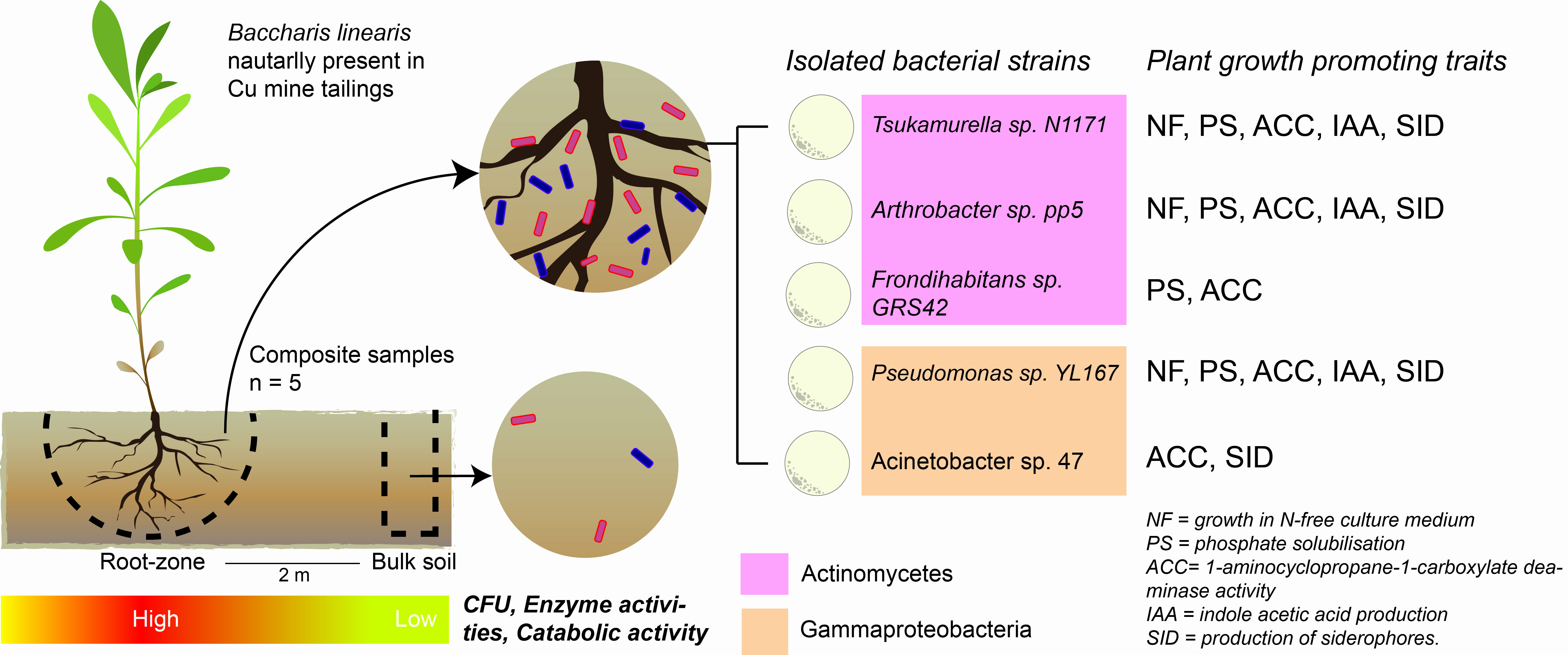
2.1. Study Site
2.2. Physicochemical Characterization of Mine Tailings
2.3. Soil Enzyme Activities
2.4. Community-Level Physiological Profile
2.5. Bacterial Plant Growth-Promoting Traits
2.6. Phylogenetic Analysis
2.7. Statistical Analyses
3. Results
3.1. Physicochemical Properties
3.2. Enzyme Activities, CFUs, and CLPP
3.3. Multivariate Analysis
3.4. Plant Growth-Promoting Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kang, X.; Cui, Y.; Shen, T.; Yan, M.; Tu, W.; Shoaib, M.; Xiang, Q.; Zhao, K.; Gu, Y.; Chen, Q.; et al. Changes of Root Microbial Populations of Natively Grown Plants during Natural Attenuation of V–Ti Magnetite Tailings. Ecotoxicol. Environ. Saf. 2020, 201, 110816. [Google Scholar] [CrossRef]
- Sun, W.; Ji, B.; Khoso, S.A.; Tang, H.; Liu, R.; Wang, L.; Hu, Y. An Extensive Review on Restoration Technologies for Mining Tailings. Environ. Sci. Pollut. Res. 2018, 25, 33911–33925. [Google Scholar] [CrossRef]
- Sun, W.; Xiao, E.; Häggblom, M.; Krumins, V.; Dong, Y.; Sun, X.; Li, F.; Wang, Q.; Li, B.; Yan, B. Bacterial Survival Strategies in an Alkaline Tailing Site and the Physiological Mechanisms of Dominant Phylotypes As Revealed by Metagenomic Analyses. Environ. Sci. Technol. 2018, 52, 13370–13380. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Li, G.; Wei, Z.; Wan, L.; Shui, G.; Jing, X. Stability Analysis of a Copper Tailings Dam via Laboratory Model Tests: A Chinese Case Study. Miner. Eng. 2011, 24, 122–130. [Google Scholar] [CrossRef]
- Aponte, H.; Meli, P.; Butler, B.; Paolini, J.; Matus, F.; Merino, C.; Cornejo, P.; Kuzyakov, Y. Meta-Analysis of Heavy Metal Effects on Soil Enzyme Activities. Sci. Total Environ. 2020, 737, 139744. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, L. Magnetite Recovery from Copper Tailings Increases Arsenic Distribution in Solution Phase and Uptake in Native Grass. J. Environ. Manag. 2017, 186, 175–182. [Google Scholar] [CrossRef]
- Festin, E.S.; Tigabu, M.; Chileshe, M.N.; Syampungani, S.; Odén, P.C. Progresses in Restoration of Post-Mining Landscape in Africa. J. For. Res. 2019, 30, 381–396. [Google Scholar] [CrossRef]
- Wang, L.; Ji, B.; Hu, Y.; Liu, R.; Sun, W. A Review on in Situ Phytoremediation of Mine Tailings. Chemosphere 2017, 184, 594–600. [Google Scholar] [CrossRef]
- Lee, S.-H.; Park, H.; Kim, J.-G. Current Status of and Challenges for Phytoremediation as a Sustainable Environmental Management Plan for Abandoned Mine Areas in Korea. Sustainability 2023, 15, 2761. [Google Scholar] [CrossRef]
- Shahrokh, V.; Martínez-Martínez, S.; Faz, Á.; Zornoza, R.; Acosta, J.A. Efficiency of Large-Scale Aided Phytostabilization in a Mining Pond. Environ. Geochem. Health 2023, 45, 4665–4677. [Google Scholar] [CrossRef]
- Galvez, G.; Ortega, J.; Fredericksen, F.; Aliaga-Tobar, V.; Parra, V.; Reyes-Jara, A.; Pizarro, L.; Latorre, M. Co-Occurrence Interaction Networks of Extremophile Species Living in a Copper Mining Tailing. Front. Microbiol. 2022, 12, 791127. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sun, Q.; Wang, W.; Liu, X.; Song, L.; Hou, L. Effects of Different Native Plants on Soil Remediation and Microbial Diversity in Jiulong Iron Tailings Area, Jiangxi. Forests 2022, 13, 1106. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, H.; Zhan, J.; Yu, H.; Ye, D.; Zhang, X.; Zheng, Z.; Wang, Y.; Li, T. Indigenous Rhizosphere Microbial Community Characteristics of the Phytostabilizer Athyrium wardii (Hook.) Grown in a Pb/Zn Mine Tailing. Chemosphere 2022, 308, 136552. [Google Scholar] [CrossRef] [PubMed]
- Vidal, C.; González, F.; Santander, C.; Pérez, R.; Gallardo, V.; Santos, C.; Aponte, H.; Ruiz, A.; Cornejo, P. Management of Rhizosphere Microbiota and Plant Production under Drought Stress: A Comprehensive Review. Plants 2022, 11, 2437. [Google Scholar] [CrossRef]
- Pérez, R.; Tapia, Y.; Antilén, M.; Casanova, M.; Vidal, C.; Santander, C.; Aponte, H.; Cornejo, P. Interactive Effect of Compost Application and Inoculation with the Fungus Claroideoglomus Claroideum in Oenothera Picensis Plants Growing in Mine Tailings. Ecotoxicol. Environ. Saf. 2021, 208, 111495. [Google Scholar] [CrossRef]
- Gazitúa, M.C.; Morgante, V.; Poupin, M.J.; Ledger, T.; Rodríguez-Valdecantos, G.; Herrera, C.; del Carmen González-Chávez, M.; Ginocchio, R.; González, B. The Microbial Community from the Early-Plant Colonizer (Baccharis Linearis) Is Required for Plant Establishment on Copper Mine Tailings. Sci. Rep. 2021, 11, 10448. [Google Scholar] [CrossRef]
- Romero-Estonllo, M.; Ramos-Castro, J.; San Miguel del Río, Y.; Rodríguez-Garrido, B.; Prieto-Fernández, Á.; Kidd, P.S.; Monterroso, C. Soil Amendment and Rhizobacterial Inoculation Improved Cu Phytostabilization, Plant Growth and Microbial Activity in a Bench-Scale Experiment. Front. Microbiol. 2023, 14, 1184070. [Google Scholar] [CrossRef]
- Heckenroth, A.; Rabier, J.; Dutoit, T.; Torre, F.; Prudent, P.; Laffont-Schwob, I. Selection of Native Plants with Phytoremediation Potential for Highly Contaminated Mediterranean Soil Restoration: Tools for a Non-Destructive and Integrative Approach. J. Environ. Manag. 2016, 183, 850–863. [Google Scholar] [CrossRef]
- Ginocchio, R.; de la Fuente, L.M.; Orrego, F.; Díaz, M.J.; Báez, J.; Ovalle, J.F. A Novel Fast-Vegetative Propagation Technique of the Pioneer Shrub Baccharis Linearis on Mine Tailings by Adding Compost. Int. J. Phytoremediation 2021, 23, 1169–1174. [Google Scholar] [CrossRef]
- Pérez, R.; Tapia, Y.; Antilén, M.; Casanova, M.; Vidal, C.; Silambarasan, S.; Cornejo, P. Rhizosphere Management for Phytoremediation of Copper Mine Tailings. J. Soil Sci. Plant Nutr. 2021, 21, 3091–3109. [Google Scholar] [CrossRef]
- Riedemann, P.; Aldunate, G. Flora Nativa de Valor Ornamental, Identificacion y Propagación, Chile Zona Central. Zona Norte. In Colección General Biblioteca, 2nd ed.; Jardín Botánico Chagual: Santiago, Chile, 2001. [Google Scholar]
- Ginocchio, R.; León-Lobos, P.; Arellano, E.C.; Anic, V.; Ovalle, J.F.; Baker, A.J.M. Soil Physicochemical Factors as Environmental Filters for Spontaneous Plant Colonization of Abandoned Tailing Dumps. Environ. Sci. Pollut. Res. 2017, 24, 13484–13496. [Google Scholar] [CrossRef]
- Pathom-aree, W.; Matako, A.; Rangseekaew, P.; Seesuriyachan, P.; Srinuanpan, S. Performance of Actinobacteria Isolated from Rhizosphere Soils on Plant Growth Promotion under Cadmium Toxicity. Int. J. Phytoremediation 2021, 23, 1497–1505. [Google Scholar] [CrossRef]
- Hou, J.; Wang, Q.; Liu, W.; Zhong, D.; Ge, Y.; Christie, P.; Luo, Y. Soil Microbial Community and Association Network Shift Induced by Several Tall Fescue Cultivars during the Phytoremediation of a Petroleum Hydrocarbon-Contaminated Soil. Sci. Total Environ. 2021, 792, 148411. [Google Scholar] [CrossRef] [PubMed]
- Armesto, J.J.; Steward, T.; Pickett, A. A Mechanistic Approach to the Study of Succession in the Chilean Matorral. Rev. Chil. Hist. Nat. 1985, 58, 9–17. [Google Scholar]
- Castro, P.; Parada, R.; Corrial, C.; Mendoza, L.; Cotoras, M. Endophytic Fungi Isolated from Baccharis Linearis and Echinopsis Chiloensis with Antifungal Activity against Botrytis Cinerea. J. Fungi 2022, 8, 197. [Google Scholar] [CrossRef]
- CR2 Datos de Precipitación. Available online: https://www.cr2.cl/datos-de-precipitacion/?cp_Precipitacion=2 (accessed on 5 February 2023).
- INIA Red Agrometeorológica. Available online: https://agrometeorologia.cl/# (accessed on 5 February 2023).
- Sadzawka, A.; Carrasco, R.; Mora, M.; Flores, H.; Neaman, A. Métodos de Análisis Recomendados Para Los Suelos de Chile, Revisión 2006; Serie Actas INIA N° 34; Instituto de Investigaciones Agropecuarias: Santiago, Chile, 2006. [Google Scholar]
- Bremner, J.M. Nitrogen Total. In Methods of Soil Analysis, Part 3: Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.M., Tabatabai, M.A., Johnston, C.T., Sommers, M.E., Eds.; ASA-SSSA: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Murphy, J.; Riley, J.P. A Modified Single Solution Method for the Determination of Phosphate in Natural Waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. In USDA Circular Nr 939; US Government Printing Office: Washington, DC, USA, 1954. [Google Scholar]
- Kargas, G.; Chatzigiakoumis, I.; Kollias, A.; Spiliotis, D.; Massas, I.; Kerkides, P. Soil Salinity Assessment Using Saturated Paste and Mass Soil:Water 1:1 and 1:5 Ratios Extracts. Water 2018, 10, 1589. [Google Scholar] [CrossRef]
- EPA Field Portable X-ray Fluorescence for the Determination of Elemental Concentrations in Soil and Sediment. Available online: https://www.epa.gov/hw-sw846/sw-846-test-method-6200-field-portable-x-ray-fluorescence-spectrometry-determination#:~:text=SW-846 (accessed on 14 January 2023).
- Prosser, J.A.; Speir, T.W.; Stott, D.E.; Dick, R.P. Soil Oxidoreductases and FDA Hydrolysis. In Methods of Soil Enzymology; Dick, R.P., Ed.; SSSA Book Series 9; Soil Science Society of America: Madison, WI, USA, 2011; pp. 103–124. ISBN 978-0-89118-854-4. [Google Scholar]
- Camiña, F.; Trasar-Cepeda, C.; Gil-sotres, F.; Leirós, C. Measurement of Dehydrogenase Activity in Acid Soils Rich in Organic Matter. Soil Biol. Biochem. 1998, 30, 1005–1011. [Google Scholar] [CrossRef]
- Deng, S.; Popova, I. Carbohydrate Hydrolases. In Methods of Soil Enzymology; Dick, R.P., Ed.; SSSA Book Series; Soil Science Society of America: Madison, WI, USA, 2011; Volume 9, pp. 185–209. [Google Scholar]
- Kandeler, E.; Gerber, H. Short-Term Assay of Soil Urease Activity Using Colorimetric Determination of Ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Kandeler, E.; Poll, C.; Frankenberger, W.T.; Tabatabai, M.A. Nitrogen Cycle Enzymes. In Methods of Soil Enzymology; Dick, R.P., Ed.; Soil Science Society of America: Madison, WI, USA, 2011; pp. 211–245. [Google Scholar]
- Acosta-Martínez, V.; Ali Tabatabai, M. Phosphorus Cycle Enzymes. In Methods of Soil Enzymology; Dick, R.P., Ed.; Soil Science Society of America: Madison, WI, USA, 2011; Volume 9, pp. 161–183. [Google Scholar]
- Chapman, S.J.; Campbell, C.D.; Artz, R.R.E. Assessing CLPPs Using MicroRespTM. J. Soils Sediments 2007, 7, 406–410. [Google Scholar] [CrossRef]
- Garland, J.L. Analytical Approaches to the Characterization of Samples of Microbial Communities Using Patterns of Potential C Source Utilization. Soil Biol. Biochem. 1996, 28, 213–221. [Google Scholar] [CrossRef]
- Maier, R.M.; Pepper, I.L.; Gerba, C.P. Environmental Microbiology, 2nd ed.; Academic Press: London, UK, 2009. [Google Scholar]
- Leontidou, K.; Genitsaris, S.; Papadopoulou, A.; Kamou, N.; Bosmali, I.; Matsi, T.; Madesis, P.; Vokou, D.; Karamanoli, K.; Mellidou, I. Plant Growth Promoting Rhizobacteria Isolated from Halophytes and Drought-Tolerant Plants: Genomic Characterisation and Exploration of Phyto-Beneficial Traits. Sci. Rep. 2020, 10, 14857. [Google Scholar] [CrossRef] [PubMed]
- Majeed, A.; Abbasi, M.K.; Hameed, S.; Imran, A.; Rahim, N. Isolation and Characterization of Plant Growth-Promoting Rhizobacteria from Wheat Rhizosphere and Their Effect on Plant Growth Promotion. Front. Microbiol. 2015, 6, 198. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Jinal, H.N.; Amaresan, N. Isolation and Characterization of Drought Resistance Bacteria for Plant Growth Promoting Properties and Their Effect on Chilli (Capsicum annuum) Seedling under Salt Stress. Biocatal. Agric. Biotechnol. 2017, 12, 85–89. [Google Scholar] [CrossRef]
- Döbereiner, J. Isolation and Identification of Aerobic Nitrogen-Fixing Bacteria from Soil and Plants. In Methods in Applied Soil Microbiology and Biochemistry; Alef, K., Nannipieri, P., Eds.; Elsevier: San Diego, CA, USA, 1995; pp. 134–141. [Google Scholar]
- Dworkin, M.; Foster, J.W. Experiments with Some Microorganisms Which Utilize Ethane and Hydrogen. J. Bacteriol. 1958, 75, 592–603. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal Chemical Assay for the Detection and Determination of Siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- DeLong, E.F. Archaea in Coastal Marine Environments. Proc. Natl. Acad. Sci. USA 1992, 89, 5685–5689. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix (Version 0.90). 2021. Available online: https//github.com/taiyun/corrplot (accessed on 6 March 2022).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package 2020. R Package Version 2.6-4. Available online: https://github.com/vegandevs/vegan (accessed on 5 March 2022).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses 2020. R Package Version 1.0.7. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 10 March 2022).
- Ginocchio, R.; Carvallo, G.; Toro, I.; Bustamante, E.; Silva, Y.; Sepúlveda, N. Micro-Spatial Variation of Soil Metal Pollution and Plant Recruitment near a Copper Smelter in Central Chile. Environ. Pollut. 2004, 127, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Alloway, B.J. (Ed.) Heavy Metals in Soils; Environmental Pollution; Springer: Dordrecht, The Netherlands, 2013; Volume 22, ISBN 978-94-007-4469-1. [Google Scholar]
- Harter, R.D.; Naidu, R. Role of Metal-Organic Complexation in Metal Sorption by Soils. Adv. Agron. 1995, 55, 219–263. [Google Scholar] [CrossRef]
- Hossner, L.; Shahandeh, H. Rehabilitation of Minerals Processing Residue (Tailings). In Encyclopedia of Soil Science, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 1450–1455. [Google Scholar]
- Cornejo, P.; Meier, S.; Borie, G.; Rillig, M.C.; Borie, F. Glomalin-Related Soil Protein in a Mediterranean Ecosystem Affected by a Copper Smelter and Its Contribution to Cu and Zn Sequestration. Sci. Total Environ. 2008, 406, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Schoffer, J.T.; Aponte, H.; Neaman, A.; de la Fuente, L.M.; Arellano, E.C.; Gil, P.M.; Ginocchio, R. Copper Content in Soils and Litter from Fruit Orchards in Central Chile and Its Relationship with Soil Microbial Activity. Plant Soil Environ. 2022, 68, 115–128. [Google Scholar] [CrossRef]
- Aponte, H.; Mondaca, P.; Santander, C.; Meier, S.; Paolini, J.; Butler, B.; Rojas, C.; Diez, M.C.; Cornejo, P. Enzyme Activities and Microbial Functional Diversity in Metal(Loid) Contaminated Soils near to a Copper Smelter. Sci. Total Environ. 2021, 779, 146423. [Google Scholar] [CrossRef]
- Azarbad, H.; Niklińska, M.; van Gestel, C.A.M.; van Straalen, N.M.; Röling, W.F.M.; Laskowski, R. Microbial Community Structure and Functioning along Metal Pollution Gradients. Environ. Toxicol. Chem. 2013, 32, 1992–2002. [Google Scholar] [CrossRef]
- Tietjen, T.; Wetzel, R.G. Extracellular Enzyme-Clay Mineral Complexes: Enzyme Adsorption, Alteration of Enzyme Activity, and Protection from Photodegradation. Aquat. Ecol. 2003, 37, 331–339. [Google Scholar] [CrossRef]
- Xian, X.; In Shokohifard, G. Effect of PH on Chemical Forms and Plant Availability of Cadmium, Zinc, and Lead in Polluted Soils. Water Air Soil Pollut. 1989, 45, 265–273. [Google Scholar] [CrossRef]
- Speir, T.W.; Ross, D.J. Soil Phosphatase and Sulphatase. In Soil Enzymes; Academic Press: London, UK, 1978; pp. 197–250. [Google Scholar]
- Stazi, S.R.; Moscatelli, M.C.; Papp, R.; Crognale, S.; Grego, S.; Martin, M.; Marabottini, R. A Multi-Biological Assay Approach to Assess Microbial Diversity in Arsenic (As) Contaminated Soils. Geomicrobiol. J. 2017, 34, 183–192. [Google Scholar] [CrossRef]
- Rodriguez, R.; Durán, P. Natural Holobiome Engineering by Using Native Extreme Microbiome to Counteract the Climate Change Effects. Front. Bioeng. Biotechnol. 2020, 8, 568. [Google Scholar] [CrossRef]
- Li, Y.; Wen, H.; Chen, L.; Yin, T. Succession of Bacterial Community Structure and Diversity in Soil along a Chronosequence of Reclamation and Re-Vegetation on Coal Mine Spoils in China. PLoS ONE 2014, 9, e115024. [Google Scholar] [CrossRef]
- Ortiz, J.; Soto, J.; Almonacid, L.; Fuentes, A.; Campos-Vargas, R.; Arriagada, C. Alleviation of Metal Stress by Pseudomonas Orientalis and Chaetomium Cupreum Strains and Their Effects on Eucalyptus Globulus Growth Promotion. Plant Soil 2019, 436, 449–461. [Google Scholar] [CrossRef]
- Zengerer, V.; Schmid, M.; Bieri, M.; Müller, D.C.; Remus-Emsermann, M.N.P.; Ahrens, C.H.; Pelludat, C. Pseudomonas Orientalis F9: A Potent Antagonist against Phytopathogens with Phytotoxic Effect in the Apple Flower. Front. Microbiol. 2018, 9, 145. [Google Scholar] [CrossRef] [PubMed]
- Roy, P.; Kumar, A. Arthrobacter. In Beneficial Microbes in Agro-Ecology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 3–11. [Google Scholar]
- Xu, X.; Xu, M.; Zhao, Q.; Xia, Y.; Chen, C.; Shen, Z. Complete Genome Sequence of Cd(II)-Resistant Arthrobacter Sp. PGP41, a Plant Growth-Promoting Bacterium with Potential in Microbe-Assisted Phytoremediation. Curr. Microbiol. 2018, 75, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Nordstedt, N.P.; Jones, M.L. Isolation of Rhizosphere Bacteria That Improve Quality and Water Stress Tolerance in Greenhouse Ornamentals. Front. Plant Sci. 2020, 11, 826. [Google Scholar] [CrossRef]
- Safaei, S.; Fatahi-Bafghi, M.; Pouresmaeil, O. Role of Tsukamurella Species in Human Infections: First Literature Review. New Microbes New Infect. 2018, 22, 6–12. [Google Scholar] [CrossRef]
- Limcharoensuk, T.; Sooksawat, N.; Sumarnrote, A.; Awutpet, T.; Kruatrachue, M.; Pokethitiyook, P.; Auesukaree, C. Bioaccumulation and Biosorption of Cd2+ and Zn2+ by Bacteria Isolated from a Zinc Mine in Thailand. Ecotoxicol. Environ. Saf. 2015, 122, 322–330. [Google Scholar] [CrossRef]
- Zhang, H.; Han, L.; Jiang, B.; Long, C. Identification of a Phosphorus-Solubilizing Tsukamurella Tyrosinosolvens Strain and Its Effect on the Bacterial Diversity of the Rhizosphere Soil of Peanuts Growth-Promoting. World J. Microbiol. Biotechnol. 2021, 37, 109. [Google Scholar] [CrossRef]
- Kügler, J.H.; Muhle-Goll, C.; Kühl, B.; Kraft, A.; Heinzler, R.; Kirschhöfer, F.; Henkel, M.; Wray, V.; Luy, B.; Brenner-Weiss, G.; et al. Trehalose Lipid Biosurfactants Produced by the Actinomycetes Tsukamurella Spumae and T. Pseudospumae. Appl. Microbiol. Biotechnol. 2014, 98, 8905–8915. [Google Scholar] [CrossRef]
- Glick, B.R. Bacterial ACC Deaminase and the Alleviation of Plant Stress. Adv. Appl. Microbiol. 2004, 56, 291–312. [Google Scholar] [PubMed]
- Lyngwi, N.A.; Nongkhlaw, M.; Kalita, D.; Joshi, S.R. Bioprospecting of Plant Growth Promoting Bacilli and Related Genera Prevalent in Soils of Pristine Sacred Groves: Biochemical and Molecular Approach. PLoS ONE 2016, 11, e0152951. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Singh, S.; Singh, J.; Upadhyay, N. Potential of Plant Growth Promoting Traits by Bacteria Isolated from Heavy Metal Contaminated Soils. Bull. Environ. Contam. Toxicol. 2015, 94, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, M.; Sandhya, S.; Prasad, M.N.V.; Freitas, H. Perspectives of Plant-Associated Microbes in Heavy Metal Phytoremediation. Biotechnol. Adv. 2012, 30, 1562–1574. [Google Scholar] [CrossRef] [PubMed]
- Wilson-Corral, V.; Anderson, C.W.N.; Rodriguez-Lopez, M. Gold Phytomining. A Review of the Relevance of This Technology to Mineral Extraction in the 21st Century. J. Environ. Manag. 2012, 111, 249–257. [Google Scholar] [CrossRef]
- Li, K.; Ramakrishna, W. Effect of Multiple Metal Resistant Bacteria from Contaminated Lake Sediments on Metal Accumulation and Plant Growth. J. Hazard. Mater. 2011, 189, 531–539. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Bulk Soil | Root Zone | ||||||
|---|---|---|---|---|---|---|---|---|
| pH | 4.77 | ± | 0.13 | a | 5.30 | ± | 0.23 | a |
| CE (S cm−1) | 820.1 | ± | 286.3 | a | 559.6 | ± | 349.2 | a |
| TOC (%) | 0.64 | ± | 0.13 | a | 1.11 | ± | 0.24 | a |
| PTotal | 0.20 | ± | 0.00 | a | 0.20 | ± | 0.09 | a |
| PAvailable | 16.09 | ± | 0.92 | a | 12.86 | ± | 1.82 | a |
| NTotal | 0.06 | ± | 0.02 | a | 0.08 | ± | 0.03 | a |
| KTotal | 1.14 | ± | 0.04 | a | 1.15 | ± | 0.05 | a |
| CaTotal | 2.50 | ± | 0.03 | a | 2.35 | ± | 0.08 | a |
| MgTotal | 0.87 | ± | 0.02 | a | 0.81 | ± | 0.04 | a |
| STotal | 0.65 | ± | 0.10 | a | 0.52 | ± | 0.05 | a |
| FeTotal | 5.06 | ± | 0.06 | a | 4.81 | ± | 0.08 | b |
| CuTotal | 977.04 | ± | 137.02 | a | 510.31 | ± | 39.88 | b |
| ZnTotal | 51.48 | ± | 0.78 | a | 48.03 | ± | 1.52 | a |
| AsTotal | 23.77 | ± | 1.99 | a | 20.80 | ± | 1.20 | a |
| MnTotal | 994.23 | ± | 36.94 | a | 980.00 | ± | 46.79 | a |
| MoTotal | 37.11 | ± | 2.39 | a | 41.80 | ± | 3.42 | a |
| CuH2O | 16.70 | ± | 6.20 | a | 1.40 | ± | 0.34 | b |
| ZnH2O | 1.14 | ± | 0.31 | a | 0.29 | ± | 0.14 | b |
| CaH2O | 58.21 | ± | 19.55 | a | 41.73 | ± | 17.07 | a |
| MgH2O | 37.98 | ± | 13.08 | a | 23.78 | ± | 10.25 | a |
| Bacterial Strain | NF | PS | ACC | IAA | SID |
|---|---|---|---|---|---|
| Pseudomonas sp. YL167 | + | + | + | + | + |
| Arthrobacter sp. pp5 | + | + | + | + | + |
| Frondihabitans sp. GRS42 | − | + | + | − | − |
| Acinetobacter sp. 47 | − | − | + | − | + |
| Tsukamurella sp. N1171 | + | + | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aponte, H.; Sulbaran-Bracho, Y.; Mondaca, P.; Vidal, C.; Pérez, R.; Meier, S.; Cornejo, P.; Rojas, C. Biochemical, Catabolic, and PGP Activity of Microbial Communities and Bacterial Strains from the Root Zone of Baccharis linearis in a Mediterranean Mine Tailing. Microorganisms 2023, 11, 2639. https://doi.org/10.3390/microorganisms11112639
Aponte H, Sulbaran-Bracho Y, Mondaca P, Vidal C, Pérez R, Meier S, Cornejo P, Rojas C. Biochemical, Catabolic, and PGP Activity of Microbial Communities and Bacterial Strains from the Root Zone of Baccharis linearis in a Mediterranean Mine Tailing. Microorganisms. 2023; 11(11):2639. https://doi.org/10.3390/microorganisms11112639
Chicago/Turabian StyleAponte, Humberto, Yoelvis Sulbaran-Bracho, Pedro Mondaca, Catalina Vidal, Rodrigo Pérez, Sebastián Meier, Pablo Cornejo, and Claudia Rojas. 2023. "Biochemical, Catabolic, and PGP Activity of Microbial Communities and Bacterial Strains from the Root Zone of Baccharis linearis in a Mediterranean Mine Tailing" Microorganisms 11, no. 11: 2639. https://doi.org/10.3390/microorganisms11112639