
Lactiplantibacillus plantarum from Unexplored Tunisian Ecological Niches: Antimicrobial Potential, Probiotic and Food Applications

 , ,
, ,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of LAB and Preliminary Screening for Antimicrobial Activity
2.2. Molecular Identification of Selected Isolates
2.3. Antimicrobial Activity and Partial Characterization of the Antimicrobial Agent
2.4. Safety Features: Antibiotic Resistance and Hemolytic Activity
2.5. Autoaggregation and Co-Aggregation Assays
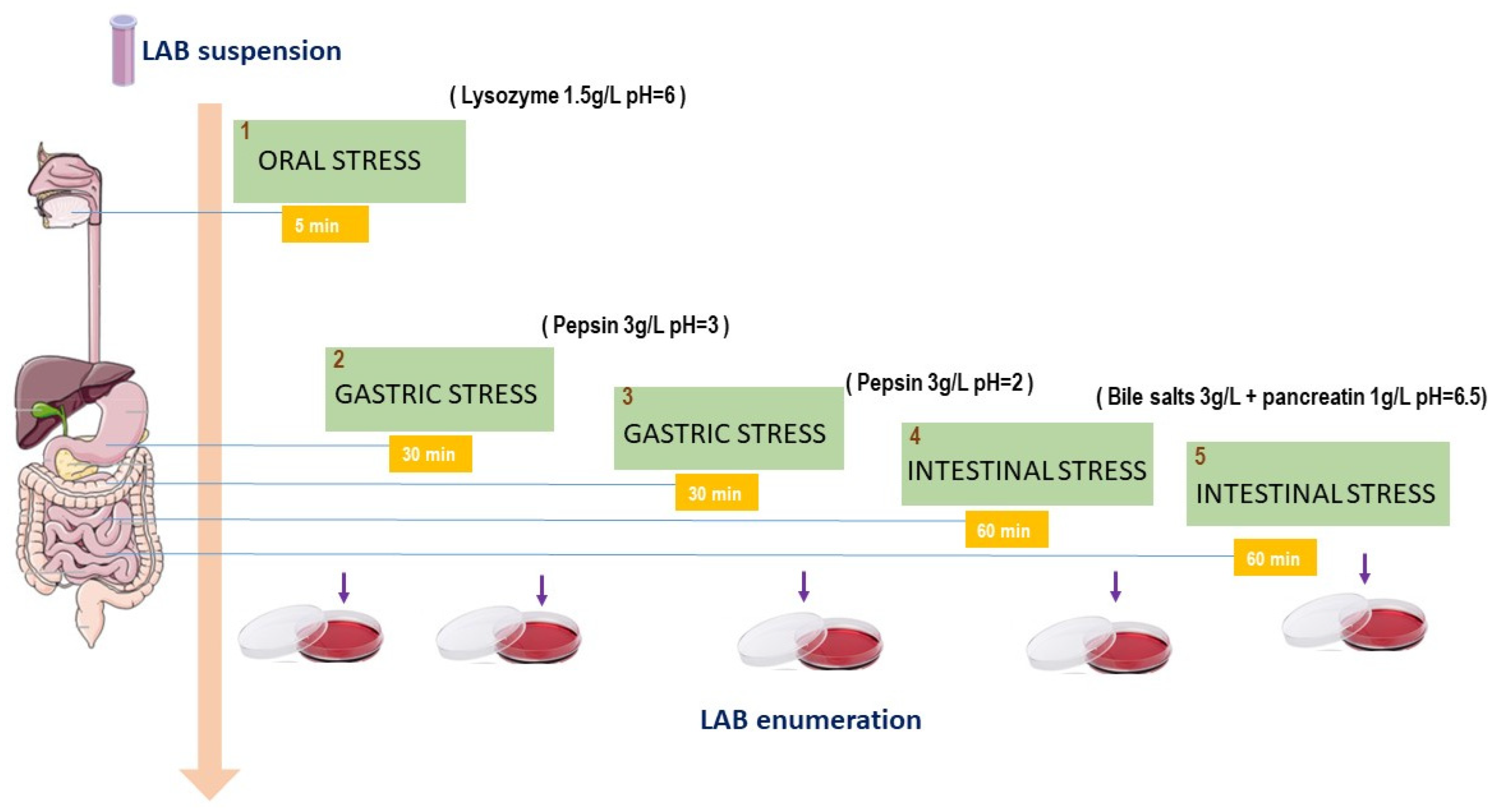
2.6. In Vitro Oro-Gastrointestinal Transit Tolerance Assay
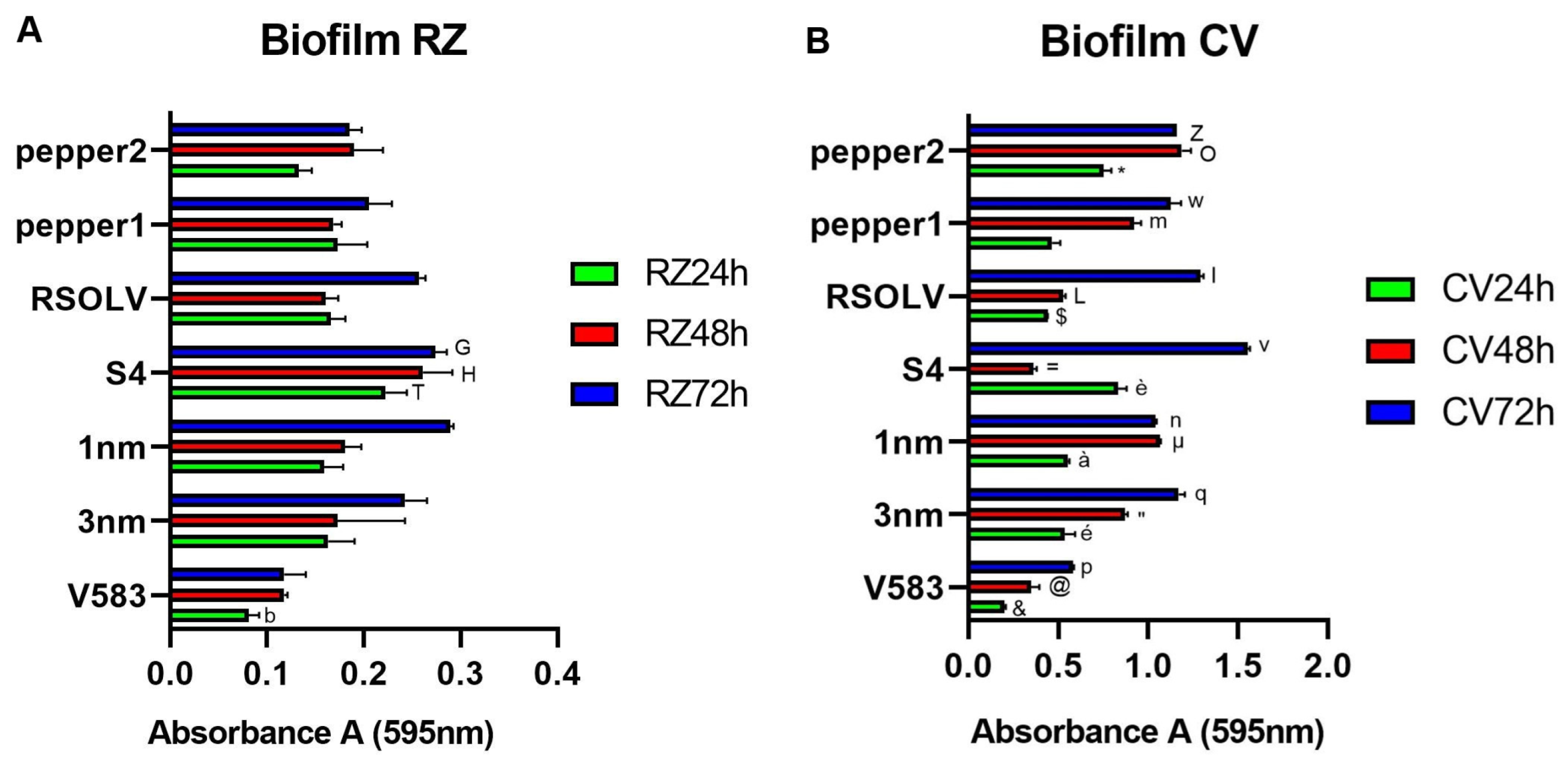
2.7. Biofilm Formation Assay
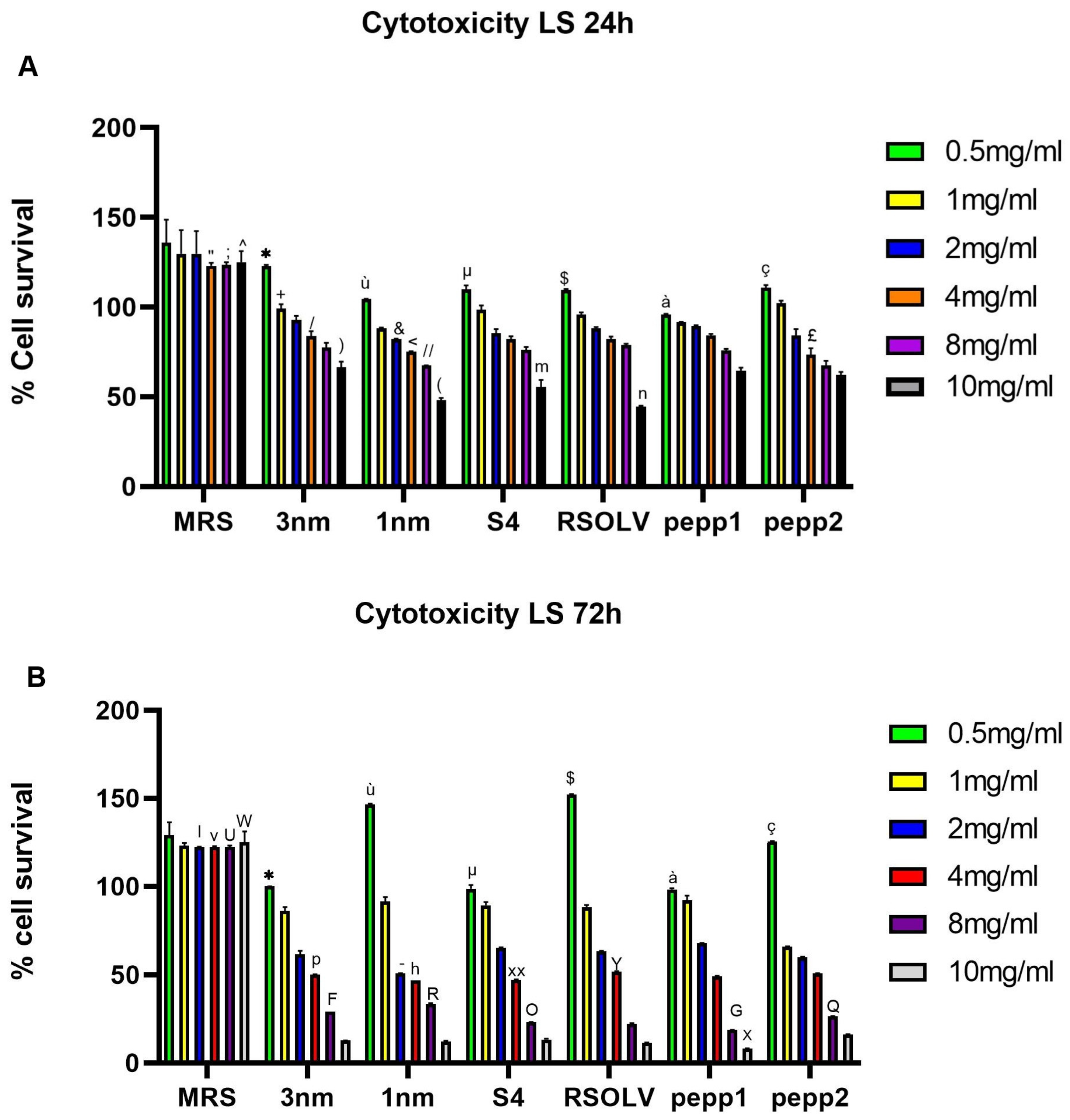
2.8. Cytotoxic Effects of Lactiplantibacillus Plantarum Strains
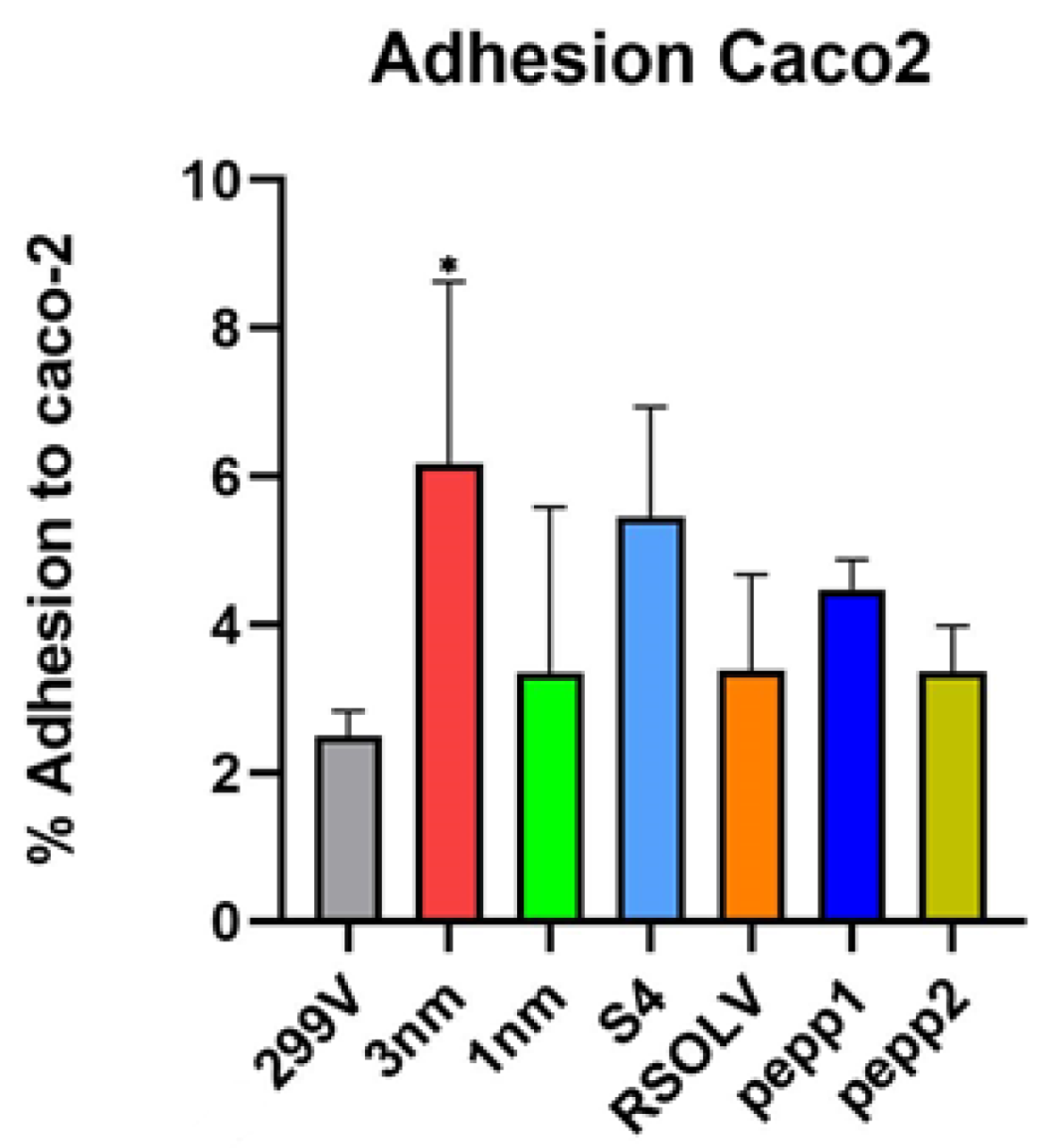
2.9. Adhesion to Human Enterocyte-Like Cells
2.10. Application in the Food Model
2.10.1. Adhesion on the Tomato Surface
2.10.2. Biocontrol/Pathogen Antagonism on Tomatoes
2.11. Statistical Analysis
3. Results
3.1. Isolation and Characterization of LAB

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolated LAB Strain | Source/Matrix | E. coli (mm) | L. monocytogenes (mm) |
|---|---|---|---|
| 3 nm | Intestines of dead locusts | 14.17 ± 0.29 | 13.17 ± 0.57 |
| 1 nm | Intestines of dead locusts | 15.00 ± 0.00 | 14.57 ± 2.32 |
| S4 | Fermented green olive brine | 15.00 ± 1.00 | 11.23 ± 0.68 |
| RSOLV | Fermented olives | 14.17 ± 0.76 | 13.43 ± 0.93 |
| pepp1 | Fermented pepper brine | 15.00 ± 0.00 | 15.10 ± 0.26 |
| pepp2 | Fermented green pepper brine | 15.00 ± 0.00 | 14.50 ± 0.50 |
| S5 | Horse sausage | 10.00 ± 1.00 | 12.07 ± 0.40 |
| S6 | Horse sausage | 0.00 ± 0.00 | 0.00 ± 0.00 |
| N8 | Dried anchovy | 11.00 ± 1.00 | 0.00 ± 0.00 |
| N4c | Dried fermented anchovy | 12.00 ± 1.00 | 8.03 ± 0.25 |
| F1c | Infant feces | 9.13 ± 0.71 | 0.00 ± 0.00 |
| F5a | Infant feces | 12.90 ± 0.17 | 9.97 ± 0.15 |
| LM | Breast milk | 9.83 ± 0.29 | 9.92 ± 0.14 |
| F1 | Infant feces | 10.17 ± 0.29 | 11.00 ± 0.00 |
| Rg4a | Artisanal Tunisian ricotta cheese | 10.17 ± 0.29 | 11.17 ± 1.26 |
| RL4 | Tunisian fermented milk Leben | 8.83 ± 0.29 | 12.93 ± 0.12 |
| K10 | Tunisian artisanal Gueddid | 0.00 ± 0.00 | 10.60 ± 0.36 |
| LC4 | Goat milk | 0.00 ± 0.00 | 10.50 ± 0.50 |
| S1 | Fermented olive brine | 10.80 ± 0.36 | 10.47 ± 0.50 |
| AIB | Rabbit intestines | 0.00 ± 0.00 | 11.00 ± 0.00 |
| S5 | Horse sausage | 11.00 ± 1.00 | 11.00 ± 0.87 |
| N8 | Dried anchovy | 9.90 ± 0.10 | 9.00 ± 0.00 |
| K10 | Tunisian Gueddid | 0.00 ± 0.00 | 0.00 ± 0.00 |
| O3 | Fermented olive brine | 0.00 ± 0.00 | 0.00 ± 0.00 |
3.2. Antimicrobial Activity and Some Insights into the Antibacterial Agents
3.3. Auto-Aggregation and Co-Aggregation Assays
3.4. Safety Features: Antibiotic Resistance and Hemolytic Activity
3.5. In Vitro Oro-Gastrointestinal Transit Tolerance Assay
3.6. Biofilm Formation
3.7. Cytotoxic Effects
3.8. Adhesion to Human Enterocyte-Like Cells
3.9. Application in Food Model
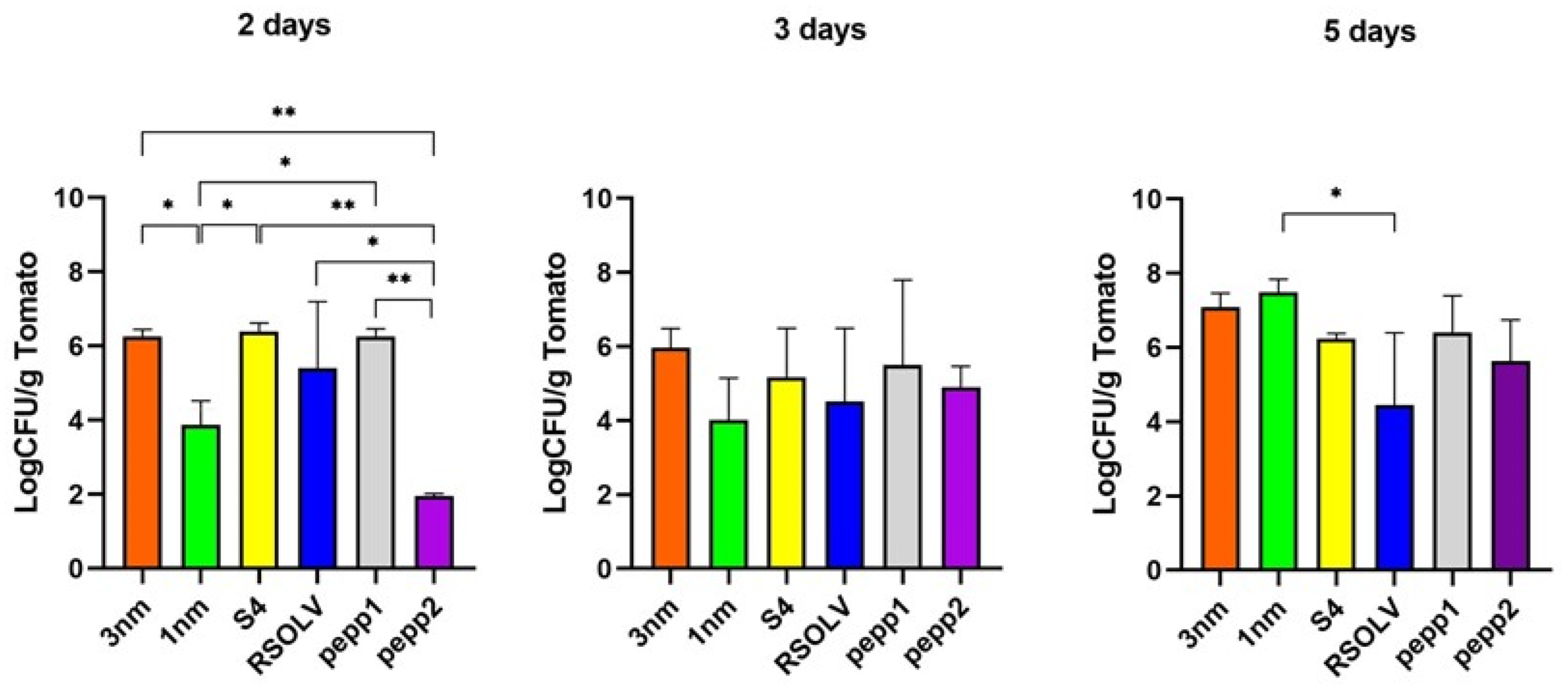
3.9.1. Adhesion on the Tomato Surface
3.9.2. Biocontrol/Pathogen Antagonism on Tomatoes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mokoena, M.P. Lactic Acid Bacteria and Their Bacteriocins: Classification, Biosynthesis and Applications against Uropathogens: A Mini-Review. Molecules 2017, 22, 1255. [Google Scholar] [CrossRef] [PubMed]
- Hazards, E.P.O.B.; Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F. Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 11: Suitability of taxonomic units notified to EFSA until September 2019. EFSA J. 2020, 18, e05965. [Google Scholar]
- Huang, W.; Dong, A.; Pham, H.T.; Zhou, C.; Huo, Z.; Wätjen, A.P.; Prakash, S.; Bang-Berthelsen, C.H.; Turner, M.S. Evaluation of the fermentation potential of lactic acid bacteria isolated from herbs, fruits and vegetables as starter cultures in nut-based milk alternatives. Food Microbiol. 2023, 112, 104243. [Google Scholar] [CrossRef]
- Terefe, N.S. Food Fermentation. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Mokoena, M.P.; Omatola, C.A.; Olaniran, A.O. Applications of Lactic Acid Bacteria and Their Bacteriocins against Food Spoilage Microorganisms and Foodborne Pathogens. Molecules 2021, 26, 7055. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, P.K.; Nehra, K. Bacteriocin-nanoconjugates as emerging compounds for enhancing antimicrobial activity of bacteriocins. J. King Saud. Univ. Sci. 2019, 31, 758–767. [Google Scholar] [CrossRef]
- Touret, T.; Oliveira, M.; Semedo-Lemsaddek, T. Putative probiotic lactic acid bacteria isolated from sauerkraut fermentations. PLoS ONE 2018, 13, e0203501. [Google Scholar] [CrossRef]
- Medina-Pradas, E.; Pérez-Díaz, I.M.; Garrido-Fernández, A.; Arroyo-López, F.N. Chapter 9—Review of Vegetable Fermentations With Particular Emphasis on Processing Modifications, Microbial Ecology, and Spoilage. In The Microbiological Quality of Food; Bevilacqua, A., Corbo, M.R., Sinigaglia, M., Eds.; Woodhead Publishing: Cambridge, UK, 2017; pp. 211–236. [Google Scholar]
- Saidi, Y.; Del Rio, B.; Senouci, D.E.; Redruello, B.; Martinez, B.; Ladero, V.; Kihal, M.; Alvarez, M.A. Polyphasic Characterisation of Non-Starter Lactic Acid Bacteria from Algerian Raw Camel’s Milk and Their Technological Aptitudes. Food Technol. Biotechnol. 2020, 58, 260–272. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Seddik, H.A.; Bendali, F.; Gancel, F.; Fliss, I.; Spano, G.; Drider, D. Lactobacillus plantarum and Its Probiotic and Food Potentialities. Probiotics Antimicrob. Proteins 2017, 9, 111–122. [Google Scholar] [CrossRef]
- Karnwal, A.; Malik, T. Characterization and selection of probiotic lactic acid bacteria from different dietary sources for development of functional foods. Front. Microbiol. 2023, 14, 1170725. [Google Scholar]
- Cruz, P.O.D.; Matos, C.J.; Nascimento, Y.M.; Tavares, J.F.; Souza, E.L.; Magalhães, H.I.F. Efficacy of Potentially Probiotic Fruit-Derived Lactobacillus fermentum, L. paracasei and L. plantarum to Remove Aflatoxin M(1) In Vitro. Toxins 2020, 13, 4. [Google Scholar] [CrossRef] [PubMed]
- Čanak, I.; Markov, K.; Melvan, E.; Starčević, A.; Živković, M.; Zadravec, M.; Pleadin, J.; Jakopović, Ž.; Kostelac, D.; Frece, J. Isolation and Characterisation of L. plantarum O1 Producer of Plantaricin as Potential Starter Culture for the Biopreservation of Aquatic Food Products. Food Technol. Biotechnol. 2018, 56, 581–589. [Google Scholar] [CrossRef]
- Mao, B.; Yin, R.; Li, X.; Cui, S.; Zhang, H.; Zhao, J.; Chen, W. Comparative Genomic Analysis of Lactiplantibacillus plantarum Isolated from Different Niches. Genes 2021, 12, 241. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gonzalez, N.; Battista, N.; Prete, R.; Corsetti, A. Health-promoting role of Lactiplantibacillus plantarum isolated from fermented foods. Microorganisms 2021, 9, 349. [Google Scholar] [CrossRef]
- Rocchetti, M.T.; Russo, P.; Capozzi, V.; Drider, D.; Spano, G.; Fiocco, D. Bioprospecting Antimicrobials from Lactiplantibacillus plantarum: Key Factors Underlying Its Probiotic Action. Int. J. Mol. Sci. 2021, 22, 12076. [Google Scholar] [CrossRef]
- Kwon, Y.J.; Chun, B.H.; Jung, H.S.; Chu, J.; Joung, H.; Park, S.Y.; Kim, B.K.; Jeon, C.O. Safety Assessment of Lactiplantibacillus (formerly Lactobacillus) plantarum Q180. J. Microbiol. Biotechnol. 2021, 31, 1420–1429. [Google Scholar] [CrossRef] [PubMed]
- Ismael, M.; Gu, Y.; Cui, Y.; Wang, T.; Yue, F.; Qin, Y.; Lü, X. Probiotic of Lactiplantibacillus plantarum NWAFU-BIO-BS29 Isolated from Chinese Traditional Fermented Milk and Its Potential Therapeutic Applications Based on Gut Microbiota Regulation. Foods 2022, 11, 3766. [Google Scholar] [CrossRef]
- Jeong, S.; Kim, Y.; Park, S.; Lee, D.; Lee, J.; Hlaing, S.P.; Yoo, J.W.; Rhee, S.H.; Im, E. Lactobacillus plantarum Metabolites Elicit Anticancer Effects by Inhibiting Autophagy-Related Responses. Molecules 2023, 28, 1890. [Google Scholar] [CrossRef]
- Du, G.; Guo, Q.; Qiang, L.; Chang, S.; Yan, X.; Chen, H.; Yuan, Y.; Yue, T. Influence of encapsulated Lactobacillus plantarum and eugenol on the physicochemical properties and microbial community of fresh-cut apples. Food Chem. X 2023, 17, 100563. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; Roohi, R.; Akbari, M.; Di Natale, A.; Conte, F. Inactivation of Foodborne Pathogens by Lactiplantibacillus Strains during Meat Fermentation: Kinetics and Mathematical Modelling. Foods 2023, 12, 3150. [Google Scholar] [CrossRef]
- Bvenura, C.; Sivakumar, D. The role of wild fruits and vegetables in delivering a balanced and healthy diet. Food Res. Int. 2017, 99, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Martinez, J.-P.; Lutts, S. Tomato fruit development and metabolism. Front. Plant Sci. 2019, 10, 1554. [Google Scholar] [CrossRef] [PubMed]
- Production of Tomatoes in Tunisia from 2016 to 2021. 2023. Available online: www.statista.com (accessed on 26 April 2023).
- Hwang, D.; Koh, J.H.; Lee, J.; Kim, Y.; Lim, Y.H. Cherry tomato supplementation increases the area of the intestinal mucosa and the number of muscle layers in rats. Food Res. Int. 2014, 64, 298–304. [Google Scholar] [CrossRef]
- Swain, M.R.; Anandharaj, M.; Ray, R.C.; Parveen Rani, R. Fermented fruits and vegetables of Asia: A potential source of probiotics. Biotechnol. Res. Int. 2014, 2014, 250424. [Google Scholar] [CrossRef]
- Jin, T.Z.; Fan, X.; Mukhopadhyay, S. Antimicrobial coating with organic acids and essential oil for the enhancement of safety and shelf life of grape tomatoes. Int. J. Food Microbiol. 2022, 378, 109827. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.A.; Chandry, P.S.; Kaur, M.; Kocharunchitt, C.; Bowman, J.P.; Fox, E.M. Novel Biocontrol Methods for Listeria monocytogenes Biofilms in Food Production Facilities. Front. Microbiol. 2018, 9, 605. [Google Scholar] [CrossRef]
- Lavilla, M.; Domingo-Calap, P.; Sevilla-Navarro, S.; Lasagabaster, A. Natural Killers: Opportunities and Challenges for the Use of Bacteriophages in Microbial Food Safety from the One Health Perspective. Foods 2023, 12, 552. [Google Scholar] [CrossRef]
- Iosca, G.; De Vero, L.; Di Rocco, G.; Perrone, G.; Gullo, M.; Pulvirenti, A. Anti-Spoilage Activity and Exopolysaccharides Production by Selected Lactic Acid Bacteria. Foods 2022, 11, 1914. [Google Scholar] [CrossRef]
- Yin, H.B.; Chen, C.H.; Gu, G.; Nou, X.; Patel, J. Pre-harvest biocontrol of Listeria and Escherichia coli O157 on lettuce and spinach by lactic acid bacteria. Int. J. Food Microbiol. 2023, 387, 110051. [Google Scholar] [CrossRef] [PubMed]
- González-Pérez, C.; Vargas-Arispuro, I.; Aispuro-Hernández, E.; Aguilar-Gil, C.; Aguirre-Guzmán, Y.; Castillo, A.; Hernández-Mendoza, A.; Ayala-Zavala, J.; Martínez-Téllez, M. Potential control of foodborne pathogenic bacteria by Pediococcus pentosaceus and Lactobacillus graminis isolated from fresh vegetables. Microbiol. Biotechnol. Lett. 2019, 47, 183–194. [Google Scholar] [CrossRef]
- Özer, F.; Karakaya, Ş.; Ünlü, N.; Erganiş, O.; Kav, K.; Imazato, S. Comparison of antibacterial activity of two dentin bonding systems using agar well technique and tooth cavity model. J. Dent. 2003, 31, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Angmo, K.; Kumari, A.; Bhalla, T.C. Probiotic characterization of lactic acid bacteria isolated from fermented foods and beverage of Ladakh. LWT Food Sci. Technol. 2016, 66, 428–435. [Google Scholar] [CrossRef]
- Semedo, T.; Almeida Santos, M.; Martins, P.; Silva Lopes, M.F.T.; Figueiredo Marques, J.J.; Tenreiro, R.; Barreto Crespo, M.T. Comparative study using type strains and clinical and food isolates to examine hemolytic activity and occurrence of the cyl operon in enterococci. J. Clin. Microbiol. 2003, 41, 2569–2576. [Google Scholar] [CrossRef]
- Dommels, Y.E.; Kemperman, R.A.; Zebregs, Y.E.; Draaisma, R.B.; Jol, A.; Wolvers, D.A.; Vaughan, E.E.; Albers, R. Survival of Lactobacillus reuteri DSM 17938 and Lactobacillus rhamnosus GG in the human gastrointestinal tract with daily consumption of a low-fat probiotic spread. Appl. Environ. Microbiol. 2009, 75, 6198–6204. [Google Scholar] [CrossRef]
- Hancock, L.E.; Perego, M. The Enterococcus faecalis fsr two-component system controls biofilm development through production of gelatinase. J. Bacteriol. 2004, 186, 5629–5639. [Google Scholar] [CrossRef]
- Hancock, L.; Perego, M. Two-component signal transduction in Enterococcus faecalis. J. Bacteriol. 2002, 184, 5819–5825. [Google Scholar] [CrossRef]
- Ahrne, S.; Hagslatt, M.-L.J. Effect of lactobacilli on paracellular permeability in the gut. Nutrients 2011, 3, 104–117. [Google Scholar] [CrossRef]
- Russo, P.; Arena, M.P.; Fiocco, D.; Capozzi, V.; Drider, D.; Spano, G. Lactobacillus plantarum with broad antifungal activity: A promising approach to increase safety and shelf-life of cereal-based products. Int. J. Food Microbiol. 2017, 247, 48–54. [Google Scholar] [CrossRef]
- Pei, J.; Li, X.; Han, H.; Tao, Y. Purification and characterization of plantaricin SLG1, a novel bacteriocin produced by Lb. plantarum isolated from yak cheese. Food Control 2018, 84, 111–117. [Google Scholar] [CrossRef]
- De Simone, N.; Rocchetti, M.T.; la Gatta, B.; Spano, G.; Drider, D.; Capozzi, V.; Russo, P.; Fiocco, D. Antimicrobial Properties, Functional Characterisation and Application of Fructobacillus fructosus and Lactiplantibacillus plantarum Isolated from Artisanal Honey. Probiotics Antimicrob. Proteins 2023, 15, 1406–1423. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 31st ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2021. [Google Scholar]
- Charteris, W.P.; Kelly, P.M.; Morelli, L.; Collins, J.K. Antibiotic susceptibility of potentially probiotic Lactobacillus species. J. Food Prot. 1998, 61, 1636–1643. [Google Scholar] [CrossRef]
- Reuben, R.; Roy, P.; Sarkar, S.; Alam, A.R.U.; Jahid, I. Characterization and evaluation of lactic acid bacteria from indigenous raw milk for potential probiotic properties. J. Dairy. Sci. 2020, 103, 1223–1237. [Google Scholar] [CrossRef]
- Armas, F.; Camperio, C.; Marianelli, C. In vitro assessment of the probiotic potential of Lactococcus lactis LMG 7930 against ruminant mastitis-causing pathogens. PLoS ONE 2017, 12, e0169543. [Google Scholar] [CrossRef]
- Gheziel, C.; Russo, P.; Arena, M.P.; Spano, G.; Ouzari, H.I.; Kheroua, O.; Saidi, D.; Fiocco, D.; Kaddouri, H.; Capozzi, V. Evaluating the Probiotic Potential of Lactobacillus plantarum Strains from Algerian Infant Feces: Towards the Design of Probiotic Starter Cultures Tailored for Developing Countries. Probiotics Antimicrob. Proteins 2019, 11, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Senatore, G.; Mastroleo, F.; Leys, N.; Mauriello, G. Growth of Lactobacillus reuteri DSM17938 under two simulated microgravity systems: Changes in reuterin production, gastrointestinal passage resistance, and stress genes expression response. Astrobiology 2020, 20, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gómez, N.C.; Ramiro, J.M.; Quecan, B.X.; de Melo Franco, B.D. Use of Potential Probiotic Lactic Acid Bacteria (LAB) Biofilms for the Control of Listeria monocytogenes, Salmonella typhimurium, and Escherichia coli O157:H7 Biofilms Formation. Front. Microbiol. 2016, 7, 863. [Google Scholar] [CrossRef]
- Cruz, C.D.; Shah, S.; Tammela, P. Defining conditions for biofilm inhibition and eradication assays for Gram-positive clinical reference strains. BMC Microbiol. 2018, 18, 173. [Google Scholar] [CrossRef]
- Bøhle, L.A.; Riaz, T.; Egge-Jacobsen, W.; Skaugen, M.; Busk, Ø.L.; Eijsink, V.G.; Mathiesen, G. Identification of surface proteins in Enterococcus faecalis V583. BMC Genom. 2011, 12, 135. [Google Scholar] [CrossRef]
- Patti, J.M.; Höök, M. Microbial adhesins recognizing extracellular matrix macromolecules. Curr. Opin. Cell Biol. 1994, 6, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Borges, S.; Silva, J.; Teixeira, P. Survival and biofilm formation by Group B streptococci in simulated vaginal fluid at different pHs. Antonie Leeuwenhoek 2012, 101, 677–682. [Google Scholar] [CrossRef]
- Chuah, L.O.; Foo, H.L.; Loh, T.C.; Mohammed Alitheen, N.B.; Yeap, S.K.; Abdul Mutalib, N.E.; Abdul Rahim, R.; Yusoff, K. Postbiotic metabolites produced by Lactobacillus plantarum strains exert selective cytotoxicity effects on cancer cells. BMC Complement. Altern. Med. 2019, 19, 114. [Google Scholar] [CrossRef] [PubMed]
- Fernández de Palencia, P.; López, P.; Corbí, A.L.; Peláez, C.; Requena, T. Probiotic strains: Survival under simulated gastrointestinal conditions, in vitro adhesion to Caco-2 cells and effect on cytokine secretion. Eur. Food Res. Technol. 2008, 227, 1475–1484. [Google Scholar] [CrossRef]
- Klarin, B.; Johansson, M.L.; Molin, G.; Larsson, A.; Jeppsson, B. Adhesion of the probiotic bacterium Lactobacillus plantarum 299v onto the gut mucosa in critically ill patients: A randomised open trial. Crit. Care 2005, 9, R285–R293. [Google Scholar] [CrossRef]
- Adlerberth, I.; Ahrne, S.; Johansson, M.L.; Molin, G.; Hanson, L.A.; Wold, A.E. A mannose-specific adherence mechanism in Lactobacillus plantarum conferring binding to the human colonic cell line HT-29. Appl. Environ. Microbiol. 1996, 62, 2244–2251. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Sokorai, K.; Ukuku, D.O.; Jin, T.; Fan, X.; Olanya, M.; Juneja, V. Inactivation of Salmonella in grape tomato stem scars by organic acid wash and chitosan-allyl isothiocyanate coating. Int. J. Food Microbiol. 2018, 266, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Arellano-Ayala, K.; Ascencio-Valle, F.J.; Gutiérrez-González, P.; Estrada-Girón, Y.; Torres-Vitela, M.R.; Macías-Rodríguez, M.E. Hydrophobic and adhesive patterns of lactic acid bacteria and their antagonism against foodborne pathogens on tomato surface (Solanum lycopersicum L.). J. Appl. Microbiol. 2020, 129, 876–891. [Google Scholar] [CrossRef] [PubMed]
- Sornplang, P.; Piyadeatsoontorn, S. Probiotic isolates from unconventional sources: A review. J. Anim. Sci. Technol. 2016, 58, 26. [Google Scholar] [CrossRef]
- Meradji, M.; Bachtarzi, N.; Mora, D.; Kharroub, K. Characterization of Lactic Acid Bacteria Strains Isolated from Algerian Honeybee and Honey and Exploration of Their Potential Probiotic and Functional Features for Human Use. Foods 2023, 12, 2312. [Google Scholar] [CrossRef]
- Linares-Morales, J.R.; Cuellar-Nevárez, G.E.; Rivera-Chavira, B.E.; Gutiérrez-Méndez, N.; Pérez-Vega, S.B.; Nevárez-Moorillón, G.V. Selection of Lactic Acid Bacteria Isolated from Fresh Fruits and Vegetables Based on Their Antimicrobial and Enzymatic Activities. Foods 2020, 9, 1399. [Google Scholar] [CrossRef]
- Siregar, D.J.S.; Julianti, E.; Tafsin, M.R.; Suryanto, D. Selection of probiotic candidate of lactic acid bacteria from Hermetia illucens larvae fed with different feeding substrates. Biodiversitas J. Biol. Divers. 2022, 23, d231228. [Google Scholar] [CrossRef]
- Siddiqui, R.; Elmashak, Y.; Khan, N.A. Cockroaches: A potential source of novel bioactive molecule(s) for the benefit of human health. Appl. Entomol. Zool. 2023, 58, 1–11. [Google Scholar] [CrossRef]
- Alameri, F.; Tarique, M.; Osaili, T.; Obaid, R.; Abdalla, A.; Masad, R.; Al-Sbiei, A.; Fernandez-Cabezudo, M.; Liu, S.Q.; Al-Ramadi, B.; et al. Lactic Acid Bacteria Isolated from Fresh Vegetable Products: Potential Probiotic and Postbiotic Characteristics Including Immunomodulatory Effects. Microorganisms 2022, 10, 389. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, A.; Reguant, C.; Bordons, A.; Rozès, N. Lactic acid bacteria from fermented table olives. Food Microbiol. 2012, 31, 1–8. [Google Scholar] [CrossRef]
- Ben Taheur, F.; Kouidhi, B.; Fdhila, K.; Elabed, H.; Ben Slama, R.; Mahdouani, K.; Bakhrouf, A.; Chaieb, K. Anti-bacterial and anti-biofilm activity of probiotic bacteria against oral pathogens. Microb. Pathog. 2016, 97, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Fhoula, I.; Najjari, A.; Turki, Y.; Jaballah, S.; Boudabous, A.; Ouzari, H. Diversity and antimicrobial properties of lactic acid bacteria isolated from rhizosphere of olive trees and desert truffles of Tunisia. BioMed Res. Int. 2013, 2013, 405708. [Google Scholar] [CrossRef]
- Klibi, N.; Ben Slimen, N.; Fhoula, I.; López, M.; Ben Slama, K.; Daffonchio, D.; Boudabous, A.; Torres, C.; Ouzari, H. Genotypic diversity, antibiotic resistance and bacteriocin production of enterococci isolated from rhizospheres. Microbes Environ. 2012, 27, 533–537. [Google Scholar] [CrossRef] [PubMed]
- Abouloifa, H.; Rokni, Y.; Bellaouchi, R.; Ghabbour, N.; Karboune, S.; Brasca, M.; Ben Salah, R.; Chihib, N.E.; Saalaoui, E.; Asehraou, A. Characterization of probiotic properties of antifungal Lactobacillus strains isolated from traditional fermenting green olives. Probiotics Antimicrob. Proteins 2020, 12, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Kachouri, F.; Hamdi, M. Use Lactobacillus plantarum in olive oil process and improvement of phenolic compounds content. J. Food Eng. 2006, 77, 746–752. [Google Scholar] [CrossRef]
- Garofalo, C.; Osimani, A.; Milanović, V.; Taccari, M.; Cardinali, F.; Aquilanti, L.; Riolo, P.; Ruschioni, S.; Isidoro, N.; Clementi, F. The microbiota of marketed processed edible insects as revealed by high-throughput sequencing. Food Microbiol. 2017, 62, 15–22. [Google Scholar] [CrossRef]
- Chiba, M.; Itabashi, T.; Hirai, K.; Sakamoto, M.; Ohkuma, M.; Ishige, T.; Kawasaki, S. Lactobacillus metriopterae sp. nov., a novel lactic acid bacterium isolated from the gut of grasshopper Metrioptera engelhardti. Int. J. Syst. Evol. Microbiol. 2018, 68, 1484–1489. [Google Scholar] [CrossRef]
- Leska, A.; Nowak, A.; Miśkiewicz, K.; Rosicka-Kaczmarek, J. Binding and Detoxification of Insecticides by Potentially Probiotic Lactic Acid Bacteria Isolated from Honeybee (Apis mellifera L.) Environment-An In Vitro Study. Cells 2022, 11, 3743. [Google Scholar] [CrossRef]
- Vergalito, F.; Testa, B.; Cozzolino, A.; Letizia, F.; Succi, M.; Lombardi, S.J.; Tremonte, P.; Pannella, G.; Di Marco, R.; Sorrentino, E.; et al. Potential Application of Apilactobacillus kunkeei for Human Use: Evaluation of Probiotic and Functional Properties. Foods 2020, 9, 1535. [Google Scholar] [CrossRef]
- Yuan, S.; Wang, Y.; Zhao, F.; Kang, L. Complete Genome Sequence of Weissella confusa LM1 and Comparative Genomic Analysis. Front. Microbiol. 2021, 12, 749218. [Google Scholar] [CrossRef]
- Stoops, J.; Crauwels, S.; Waud, M.; Claes, J.; Lievens, B.; Van Campenhout, L. Microbial community assessment of mealworm larvae (Tenebrio molitor) and grasshoppers (Locusta migratoria migratorioides) sold for human consumption. Food Microbiol. 2016, 53, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and Inputs From Lactic Acid Bacteria and Their Bacteriocins as Alternatives to Antibiotic Growth Promoters During Food-Animal Production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef]
- Dai, J.; Fang, L.; Zhang, M.; Deng, H.; Cheng, X.; Yao, M.; Huang, L. Isolation and identification of new source of bacteriocin-producing Lactobacillus plantarum C010 and growth kinetics of its batch fermentation. World J. Microbiol. Biotechnol. 2022, 38, 67. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Xue, W.J.; Ding, H.; An, C.; Ma, S.J.; Liu, Y. Probiotic Potential of Lactobacillus Strains Isolated From Fermented Vegetables in Shaanxi, China. Front. Microbiol. 2021, 12, 774903. [Google Scholar] [CrossRef] [PubMed]
- Kivanc, M.; Yilmaz, M.; Çakir, E. Isolation and identification of lactic acid bacteria from boza, and their microbial activity against several reporter strains. Turk. J. Biol. 2011, 35, 313–324. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, X.; Xu, Z. Identification of antibacterial substances of Lactobacillus plantarum DY-6 for bacteriostatic action. Food Sci. Nutr. 2020, 8, 2854–2863. [Google Scholar] [CrossRef] [PubMed]
- Spaggiari, L.; Sala, A.; Ardizzoni, A.; De Seta, F.; Singh, D.K.; Gacser, A.; Blasi, E.; Pericolini, E. Lactobacillus acidophilus, L. plantarum, L. rhamnosus, and L. reuteri Cell-Free Supernatants Inhibit Candida parapsilosis Pathogenic Potential upon Infection of Vaginal Epithelial Cells Monolayer and in a Transwell Coculture System In Vitro. Microbiol. Spectr. 2022, 10, e0269621. [Google Scholar] [CrossRef]
- Chang, H.M.; Foo, H.L.; Loh, T.C.; Lim, E.T.C.; Abdul Mutalib, N.E. Comparative Studies of Inhibitory and Antioxidant Activities, and Organic Acids Compositions of Postbiotics Produced by Probiotic Lactiplantibacillus plantarum Strains Isolated From Malaysian Foods. Front. Vet. Sci. 2020, 7, 602280. [Google Scholar] [PubMed]
- Khani, N.; Shkouhian, S.M.J.; Kafil, H.S.; Gilani, N.; Abbasi, A.; Rad, A.H. Assessing the growth-inhibitory activity of postbiotics of Lactobacillus spp. against Staphylococcus aureus under in vitro circumstances and food model. Lett. Appl. Microbiol. 2023, 76, ovac056. [Google Scholar]
- Thorakkattu, P.; Khanashyam, A.C.; Shah, K.; Babu, K.S.; Mundanat, A.S.; Deliephan, A.; Deokar, G.S.; Santivarangkna, C.; Nirmal, N.P. Postbiotics: Current Trends in Food and Pharmaceutical Industry. Foods 2022, 11, 3094. [Google Scholar] [PubMed]
- Divyashree, S.; Anjali, P.; Somashekaraiah, R.; Sreenivasa, M. Probiotic properties of Lactobacillus casei–MYSRD 108 and Lactobacillus plantarum-MYSRD 71 with potential antimicrobial activity against Salmonella paratyphi. Biotechnol. Rep. 2021, 32, e00672. [Google Scholar]
- Hu, C.H.; Ren, L.Q.; Zhou, Y.; Ye, B.C. Characterization of antimicrobial activity of three Lactobacillus plantarum strains isolated from Chinese traditional dairy food. Food Sci. Nutr. 2019, 7, 1997–2005. [Google Scholar] [PubMed]
- Mirzaei, E.Z.; Lashani, E.; Davoodabadi, A. Antimicrobial properties of lactic acid bacteria isolated from traditional yogurt and milk against Shigella strains. GMS Hyg. Infect. Control. 2018, 13, Doc01. [Google Scholar]
- Sirisopapong, M.; Shimosato, T.; Okrathok, S.; Khempaka, S. Assessment of lactic acid bacteria isolated from the chicken digestive tract for potential use as poultry probiotics. Asian Australas. J. Anim. Sci. 2023, 36, 1209. [Google Scholar] [CrossRef]
- Abd Ellatif, S.A.; Bouqellah, N.A.; Abu-Serie, M.M.; Razik, E.S.A.; Al-Surhanee, A.A.; Askary, A.E.; Daigham, G.E.; Mahfouz, A.Y. Assessment of probiotic efficacy and anticancer activities of Lactiplantibacillus plantarum ESSG1 (MZ683194.1) and Lactiplantibacillus pentosus ESSG2 (MZ683195.1) isolated from dairy products. Environ. Sci. Pollut. Res. Int. 2022, 29, 39684–39701. [Google Scholar]
- De Simone, N.; Capozzi, V.; de Chiara, M.L.V.; Amodio, M.L.; Brahimi, S.; Colelli, G.; Drider, D.; Spano, G.; Russo, P. Screening of Lactic Acid Bacteria for the Bio-Control of Botrytis cinerea and the Potential of Lactiplantibacillus plantarum for Eco-Friendly Preservation of Fresh-Cut Kiwifruit. Microorganisms 2021, 9, 773. [Google Scholar]
- Arsoy, E.S.; Gül, L.B.; Çon, A.H. Characterization and Selection of Potential Antifungal Lactic Acid Bacteria Isolated From Turkish Spontaneous Sourdough. Curr. Microbiol. 2022, 79, 148. [Google Scholar]
- Simões, L.; Fernandes, N.; Teixeira, J.; Abrunhosa, L.; Dias, D.R. Brazilian Table Olives: A Source of Lactic Acid Bacteria with Antimycotoxigenic and Antifungal Activity. Toxins 2023, 15, 71. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Hao, X.; Yang, F.; Wang, Y.; Fan, X.; Wang, Y. Antifungal Activity of Lactobacillus plantarum ZZUA493 and Its Application to Extend the Shelf Life of Chinese Steamed Buns. Foods 2022, 11, 195. [Google Scholar] [CrossRef] [PubMed]
- Grujović, M.; Mladenović, K.G.; Semedo-Lemsaddek, T.; Laranjo, M.; Stefanović, O.D.; Kocić-Tanackov, S.D. Advantages and disadvantages of non-starter lactic acid bacteria from traditional fermented foods: Potential use as starters or probiotics. Compr. Rev. Food Sci. Food Saf. 2022, 21, 1537–1567. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Ni, L.; Tian, W.; Chi, H. Screening and Identification of Goat-Milk-Derived Lactic Acid Bacteria with Bacteriocin-like Activity and Probiotic Potentials. Microorganisms 2023, 11, 849. [Google Scholar] [CrossRef] [PubMed]
- Rabaoui, G.; Sánchez-Juanes, F.; Tebini, M.; Naghmouchi, K.; Bellido, J.L.M.; Ben-Mahrez, K.; Réjiba, S. Potential Probiotic Lactic Acid Bacteria with Anti-Penicillium expansum Activity from Different Species of Tunisian Edible Snails. Probiotics Antimicrob. Proteins 2022, 15, 82–106. [Google Scholar] [CrossRef] [PubMed]
- Chouikhi, A.; Ktari, N.; Bardaa, S.; Hzami, A.; Ben Slima, S.; Trabelsi, I.; Asehraou, A.; Ben Salah, R. A novel probiotic strain, Lactiplantibacillus plantarum LC38, isolated from Tunisian camel milk promoting wound healing in Wistar diabetic rats. Arch. Microbiol. 2021, 204, 24. [Google Scholar] [CrossRef] [PubMed]
- Ben Salah, R.; Trabelsi, I.; Ben Mansour, R.; Lassoued, S.; Chouayekh, H.; Bejar, S. A new Lactobacillus plantarum strain, TN8, from the gastro intestinal tract of poultry induces high cytokine production. Anaerobe 2012, 18, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, M.T.; Russo, P.; De Simone, N.; Capozzi, V.; Spano, G.; Fiocco, D. Immunomodulatory Activity on Human Macrophages by Cell-Free Supernatants to Explore the Probiotic and Postbiotic Potential of Lactiplantibacillus plantarum Strains of Plant Origin. Probiotics Antimicrob. Proteins 2023. [Google Scholar] [CrossRef]
- Del Re, B.; Sgorbati, B.; Miglioli, M.; Palenzona, D. Adhesion, autoaggregation and hydrophobicity of 13 strains of Bifidobacterium longum. Lett. Appl. Microbiol. 2000, 31, 438–442. [Google Scholar] [CrossRef]
- das Neves Selis, N.; de Oliveira, H.B.M.; Leão, H.F.; Dos Anjos, Y.B.; Sampaio, B.A.; Correia, T.M.L.; Almeida, C.F.; Pena, L.S.C.; Reis, M.M.; Brito, T.L.S.; et al. Lactiplantibacillus plantarum strains isolated from spontaneously fermented cocoa exhibit potential probiotic properties against Gardnerella vaginalis and Neisseria gonorrhoeae. BMC Microbiol. 2021, 21, 198. [Google Scholar]
- Gorreja, F.; Walker, W.A. The potential role of adherence factors in probiotic function in the gastrointestinal tract of adults and pediatrics: A narrative review of experimental and human studies. Gut Microbes 2022, 14, 2149214. [Google Scholar] [CrossRef] [PubMed]
- Mekky, A.F.; Hassanein, W.A.; Reda, F.M.; Elsayed, H.M. Anti-biofilm potential of Lactobacillus plantarum Y3 culture and its cell-free supernatant against multidrug-resistant uropathogen Escherichia coli U12. Saudi J. Biol. Sci. 2022, 29, 2989–2997. [Google Scholar] [CrossRef]
- Maunders, E.; Welch, M. Matrix exopolysaccharides; the sticky side of biofilm formation. FEMS Microbiol. Lett. 2017, 364, fnx120. [Google Scholar] [CrossRef]
- Wallis, J.K.; Krömker, V.; Paduch, J.H. Biofilm formation and adhesion to bovine udder epithelium of potentially probiotic lactic acid bacteria. AIMS Microbiol. 2018, 4, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Rasika, D.; Vidanarachchi, J.K.; Luiz, S.F.; Azeredo, D.R.P.; Cruz, A.G.; Ranadheera, C.S. Probiotic delivery through non-dairy plant-based food matrices. Agriculture 2021, 11, 599. [Google Scholar] [CrossRef]
- Martín, I.; Rodríguez, A.; Delgado, J.; Córdoba, J.J. Strategies for Biocontrol of Listeria monocytogenes Using Lactic Acid Bacteria and Their Metabolites in Ready-to-Eat Meat- and Dairy-Ripened Products. Foods 2022, 11, 542. [Google Scholar] [CrossRef]


| Isolate | Source | Closest Species/Strain | Percentage of Identity | Accession Number * |
|---|---|---|---|---|
| 3 nm | Dead locust (intestines) | Lactiplantibacillus plantarum | 100% | OR431596 |
| 1 nm | Dead locust (intestines) | Lactiplantibacillus plantarum | 99.86% | OR431597 |
| S4 | Fermented olive | Lactiplantibacillus plantarum | 99.91% | OR431698 |
| RSOLV | Fermented olive | Lactiplantibacillus plantarum | 99.93% | OR431599 |
| pepp1 | Fermented pepper | Lactiplantibacillus plantarum | 99.82% | OR431600 |
| pepp2 | Fermented pepper | Lactiplantibacillus plantarum | 100% | OR431601 |
| Experimental Assay | 3 nm | 1 nm | S4 | RSOLV | pepp1 | Pepp2 |
|---|---|---|---|---|---|---|
| Mobility | - | - | - | - | - | - |
| 2% NaCl growth | + | + | + | + | + | + |
| 4% NaCl growth | + | + | + | + | + | + |
| 8% NaCl growth | + | + | + | + | + | + |
| MRS.Glc 2% | + | + | + | + | + | + |
| MRS.Glc 4% | + | + | + | + | + | + |
| MRS.Fruc 2% | + | + | + | + | + | + |
| MRS.Fruc 4% | + | + | + | + | + | + |
| MRS.Sucr 2% | + | + | + | + | + | + |
| MRS.Sucr 4% | + | + | + | + | + | + |
| MRS pH 3 | + | + | + | + | + | + |
| MRS pH 4 | + | + | + | + | + | + |
| MRS pH 5 | + | + | + | + | + | + |
| MRS pH 9.2 | + | + | + | + | + | + |
| E. coli | ||||||
|---|---|---|---|---|---|---|
| Strain | LAB Culture | Crude CFS | pH-Neutralized CFS | Temperature-Treated CFS | Crude Cells | Temperature-Treated Cells |
| 3 nm | ++ | ++ | - | ++ | - | - |
| 1 nm | ++ | ++ | - | ++ | - | - |
| RSOLV | ++ | ++ | - | ++ | - | - |
| S4 | ++ | ++ | - | ++ | - | - |
| pepp1 | ++ | ++ | - | ++ | - | - |
| pepp2 | ++ | ++ | - | ++ | - | - |
| L. monocytogenes | ||||||
|---|---|---|---|---|---|---|
| Strain | LAB Culture | Crude CFS | pH-Neutralized CFS | Temperature-Treated CFS | Crude Cells | Temperature-Treated Cells |
| 3 nm | ++ | ++ | - | ++ | - | - |
| 1 nm | ++ | ++ | - | ++ | - | - |
| RSOLV | ++ | ++ | - | ++ | - | - |
| S4 | ++ | ++ | - | ++ | - | - |
| pepp1 | ++ | ++ | - | ++ | - | - |
| pepp2 | ++ | ++ | - | ++ | - | - |
| Tartaric Acid | Malic Acid | Ascorbic Acid | Lactic Acid | Acetic Acid | Succinic Acid | Fumaric Acid | Citric Acid | |
|---|---|---|---|---|---|---|---|---|
| MRS | 503.27 ± 20.85 | 3786.64 ± 95.81 | 51.80 ± 1.30 | 0.00 ± 0.00 | 17,906.53 ± 162.88 | 1066.96 ± 27.55 | 0.00 ± 0.00 | 1206.78 |
| 3 nm | 1202.55 ± 30.22 | 1326.48 ± 176.26 | 100.09 ± 42.11 | 14,931.50 ± 133.44 | 16,421.03 ± 2 236.89 | 5758.93 ± 729.88 | 15.47 ± 6.71 | 2662.19 ± 403.92 |
| 1 nm | 1347.92 ± 25.81 | 1808.24 ± 42.34 | 102.69 ± 5.66 | 14,620 ± 214.70 | 17,033.25 ± 877.62 | 11,693.70 ± 708.58 | 0.00 ± 0.00 | 1833.64 ± 134.04 |
| S4 | 1218.73 ± 42.21 | 1294.83 ± 58.28 | 117.81 ± 1.86 | 15,419.91 ± 89.09 | 16,312.59 ± 1 242.27 | 9009.26 ± 1 374.46 | 0.00 ± 0.00 | 2224.48 ± 19.15 |
| RSOLV | 677.69 ± 128.32 | 1735.10 ± 171.48 | 0.00 ± 0.00 | 14,985.98 ± 195.53 | 19,853.59 ± 541.76 | 10,193.35 ± 562.11 | 0.00 ± 0.00 | 1050.29 ± 200.52 |
| pepp1 | 1201.36 ± 89.76 | 1309.51 ± 9.18 | 135.44 ± 4.85 | 13,829.46 ± 132.96 | 19,888.39 ± 809.60 | 6897.52 ± 781.84 | 0.00 ± 0.00 | 446.71 ± 45.63 |
| pepp2 | 1092.72 ± 116.94 | 1335.78 ± 99.67 | 0.00 ± 0.00 | 15,439.67 ± 140.01 | 42,409.27 ± 4 762.60 | 7336.16 ± 473.52 | 0.00 ± 0.00 | 894.15 ± 25.05 |
| Strain | P. expansum | A. niger | B. cinerea CECT 20973 | F. culmorum | S. cerevisiae |
|---|---|---|---|---|---|
| 3 nm | + | ++ | + | ++ | - |
| 1 nm | + | ++ | + | + | - |
| RSOLV | ++ | + | ++ | ++ | - |
| S4 | ++ | ++ | + | + | - |
| pepp1 | ++ | ++ | + | + | - |
| pepp2 | ++ | ++ | ++ | ++ | - |
| T4 | T24 | |
|---|---|---|
| 3 nm | 24.31% ± 6.84% | 53.46% ± 2.01% |
| 1 nm | 16.82% ± 2.33% | 53.59% ± 12.61% |
| S4 | 18.19% ± 2.54% | 70.30% ± 4.73% |
| RSLOV | 28.87% ± 3.68% | 60.97% ± 5.67% |
| pepp1 | 21.50% ± 1.75% | 34.67% ± 0.99% |
| pepp2 | 16.70% ± 1.12% | 69.82% ± 2.20% |
| L. monocytogenes | E. coli | |||
|---|---|---|---|---|
| Strain | T4 | T24 | T4 | T24 |
| 3 nm | 7.94% ± 0.94% | 7.28% ± 0.47% | 17.41% ± 1.54% | 43.42% ± 6.08% |
| 1 nm | 6.38% ± 1.21% | 5.52% ± 0.18% | 17.73% ± 7.75% | 54.88% ± 6.98% |
| S4 | 7.47% ± 0.14% | 7.37% ± 0.46% | 15.05% ± 0.89% | 36.96% ± 5.72% |
| RSOLV | 7.169% ± 2.25% | 8.76% ± 1.16% | 18.27% ± 4.32% | 51.15% ± 4.69% |
| pepp1 | 5.38% ± 0.00% | 5.38% ± 1.34% | 20.72% ± 2.67% | 43.53% ± 0.00% |
| pepp2 | 4.99% ± 0.55% | 4.61% ± 0.79% | 28.30% ± 10.70% | 43.16% ± 1.65% |
| ATB/Strain | 3 nm | 1 nm | S4 | RSOLV | pepp1 | pepp2 |
|---|---|---|---|---|---|---|
| Ampicillin | I | S | I | I | S | I |
| Vancomycin | R | R | S | R | R | R |
| Gentamycin | I | I | R | S | S | I |
| Kanamycin | R | R | R | R | R | R |
| Streptomycin | R | S | R | R | R | R |
| Tetracyclin | R | R | R | R | R | R |
| Erythromycin | I | R | R | I | R | I |
| Clindamycin | I | R | R | I | I | I |
| Claromycin | I | R | R | R | S | I |
| Strain | IC50 (after 24 h) mg/mL | IC50 (after 72 h) mg/mL |
|---|---|---|
| 3 nm | 11.58 | 5.06 |
| 1 nm | 7.48 | 2.74 |
| S4 | 12.06 | 4.96 |
| RSOLV | 10.80 | 2.25 |
| pepp1 | 9.12 | 2.82 |
| pepp2 | 11.67 | 2.18 |
| Number of Tomato-Attached Bacteria (Log CFU/g Tomato) | ||||
|---|---|---|---|---|
| Medium Agar | 1 Day | 3 Days | 5 Days | |
| Listeria (control) | PALCAM | 1.68 ± 1.39 | 1.67 ± 1.10 | 1.74 ± 0.70 $ |
| 3 nm–Listeria | PALCAM | 1.73 ± 0.87 | 1.44 ± 0.74 | 1.52 ± 1.48 |
| 1 nm–Listeria | PALCAM | 1.70 ± 1.23 | 1.02 ± 0.62 | 0.00 |
| S4–Listeria | PALCAM | 1.46 ± 0.22 | 1.00 ± 0.67 | 0.00 |
| RSOLV–Listeria | PALCAM | 1.77 ± 0.80 | 1.614 ± 1.38 | 1.18 |
| pepp1–Listeria | PALCAM | 1.49 ± 0.997 | 1.54 ± 1.41 | 1.15 ± 0.45 |
| pepp2–Listeria | PALCAM | 1.70 ± 1.14 | 1.16 ± 0.84 | 1.63 ± 1.16 |
| E. coli (control) | SMAC | 1.92 ± 1.61 & | 1.72 ± 0.99 | 1.63 ± 1.54 |
| 3 nm–E. coli | SMAC | 1.82 ± 1.13 | 1.55 ± 1.43 | 1.34 ± 0.94 |
| 1 nm–E. coli | SMAC | 1.87 ± 1.24 | 1.70 ± 0.97 | 1.38 ± 0.31 |
| S4–E. coli | SMAC | 1.88 ± 1.05 | 1.73 ± 1.15 | 1.13 ± 0.45 |
| RSOLV–E. coli | SMAC | 1.87 ± 1.04 | 1.51 ± 1.33 | 1.56 ± 1.24 |
| pepper1–E. coli | SMAC | 1.81 ± 1.22 | 1.45 ± 1.11 | 1.24 ± 0.45 |
| pepper2–E. coli | SMAC | 1.84 ± 1.15 | 1.53 ± 0.96 | 1.22 ± 0.41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selmi, H.; Rocchetti, M.T.; Capozzi, V.; Semedo-Lemsaddek, T.; Fiocco, D.; Spano, G.; Abidi, F. Lactiplantibacillus plantarum from Unexplored Tunisian Ecological Niches: Antimicrobial Potential, Probiotic and Food Applications. Microorganisms 2023, 11, 2679. https://doi.org/10.3390/microorganisms11112679
Selmi H, Rocchetti MT, Capozzi V, Semedo-Lemsaddek T, Fiocco D, Spano G, Abidi F. Lactiplantibacillus plantarum from Unexplored Tunisian Ecological Niches: Antimicrobial Potential, Probiotic and Food Applications. Microorganisms. 2023; 11(11):2679. https://doi.org/10.3390/microorganisms11112679
Chicago/Turabian StyleSelmi, Hiba, Maria Teresa Rocchetti, Vittorio Capozzi, Teresa Semedo-Lemsaddek, Daniela Fiocco, Giuseppe Spano, and Ferid Abidi. 2023. "Lactiplantibacillus plantarum from Unexplored Tunisian Ecological Niches: Antimicrobial Potential, Probiotic and Food Applications" Microorganisms 11, no. 11: 2679. https://doi.org/10.3390/microorganisms11112679
APA StyleSelmi, H., Rocchetti, M. T., Capozzi, V., Semedo-Lemsaddek, T., Fiocco, D., Spano, G., & Abidi, F. (2023). Lactiplantibacillus plantarum from Unexplored Tunisian Ecological Niches: Antimicrobial Potential, Probiotic and Food Applications. Microorganisms, 11(11), 2679. https://doi.org/10.3390/microorganisms11112679