
Effects of Lactobacillus fermentum Administration on Intestinal Morphometry and Antibody Serum Levels in Salmonella-Infantis-Challenged Chickens

 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Design, Housing Conditions, and Animal Management
2.3. Histological Analyses
2.4. RNA Extraction, Reverse Transcription, and Quantitative Polymerase Chain Reaction Assays
2.5. Antibody Determination
2.6. Statistical Analysis
3. Results
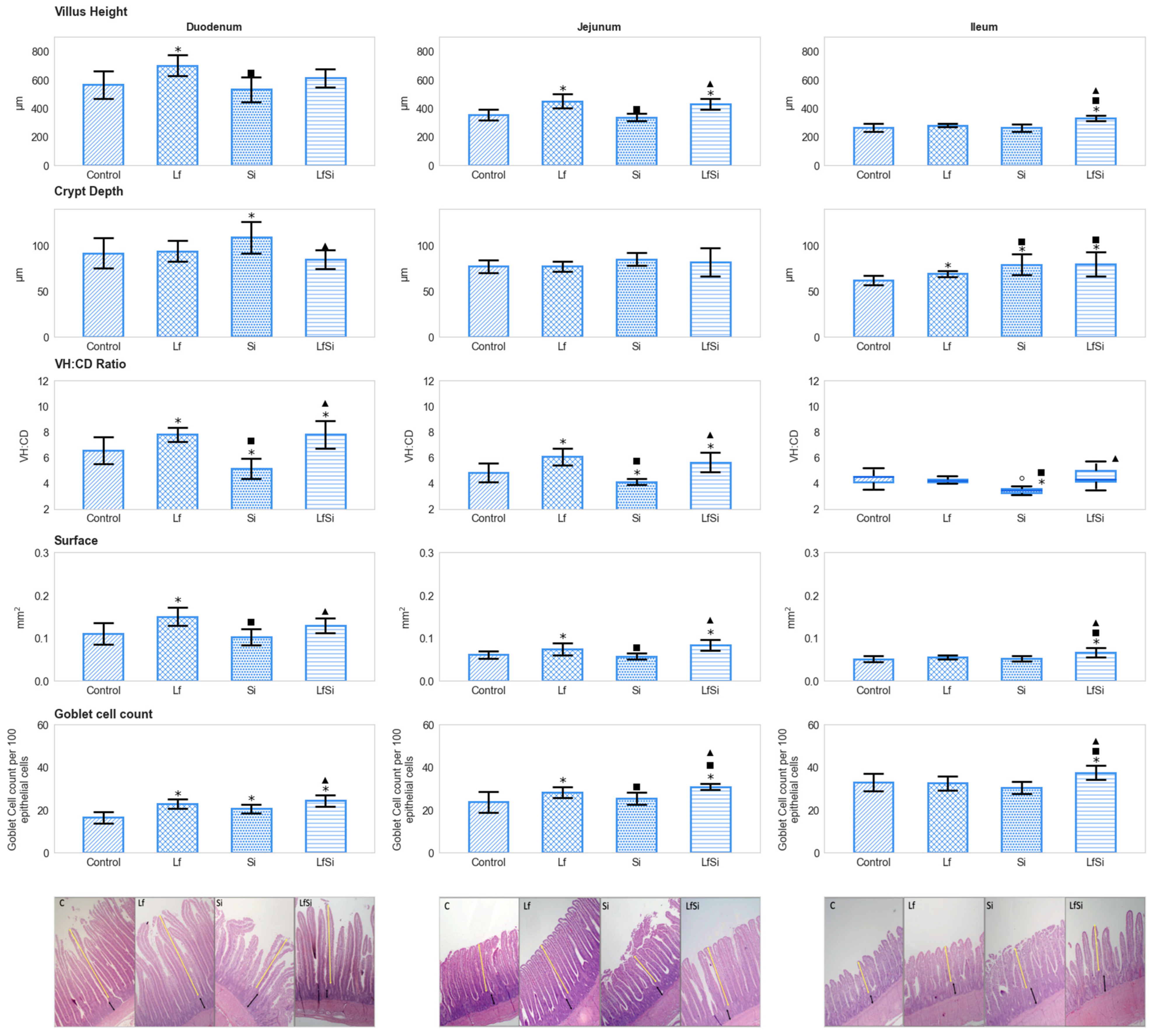
3.1. Intestinal Parameters
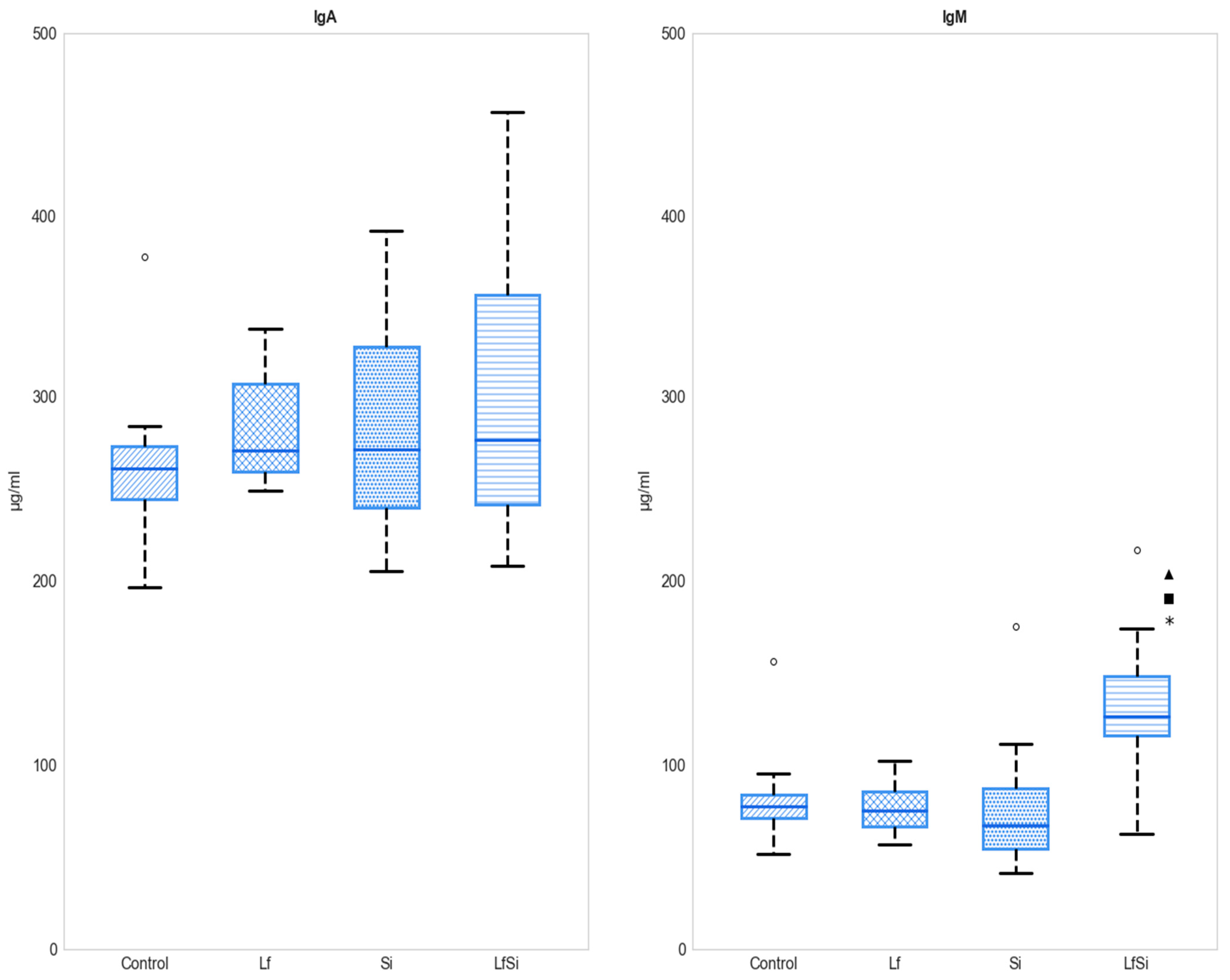
3.2. Antibody Serum Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Spencer, J.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar]
- Smith, S.I.; Seriki, A.; Ajayi, A. Typhoidal and non-typhoidal Salmonella infections in Africa. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1913–1922. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Estimates of the Global Burden of Foodborne Diseases: Foodborne Disease Burden Epidemiology Reference Group 2007–2015; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Schultz, B.M.; Melo-Gonzalez, F.; Salazar, G.A.; Porto, B.N.; Riedel, C.A.; Kalergis, A.M.; Bueno, S.M. New insights on the early interaction between typhoid and non-typhoid Salmonella serovars and the host cells. Front. Microbiol. 2021, 12, 647044. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority [EFSA]; European Centre for Disease Prevention [ECDP]. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2014. EFSA J. 2016, 14, 4380. [Google Scholar]
- Gal-Mor, O.; Valinsky, L.; Weinberger, M.; Guy, S.; Jaffe, J.; Schorr, Y.I.; Raisfeld, A.; Agmon, V.; Nissan, I. Multidrug-resistant Salmonella enterica serovar Infantis, Israel. Emerg. Infect. Dis. 2010, 16, 1754. [Google Scholar] [CrossRef] [PubMed]
- Gili, A.; Tsyba, K.; Steck, N.; Salmon-Divon, M.; Cornelius, A.; Rahav, G.; Grassl, G.A.; Gal-Mor, O. A unique megaplasmid contributes to stress tolerance and pathogenicity of an emergent Salmonella enterica serovar Infantis strain. Environ. Microbiol. 2014, 16, 977–994. [Google Scholar]
- Cohen, E.; Rahav, G.; Gal-Mor, O. Genome sequence of an emerging Salmonella enterica serovar Infantis and genomic comparison with other S. Infantis strains. Genome Biol. Evol. 2020, 12, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Torres-Elizalde, L.; Ortega-Paredes, D.; Loaiza, K.; Fernández-Moreira, E.; Larrea-Álvarez, M. In slico detection of antimicrobial resistance integrons in Salmonella enterica isolates from countries of the Andean community. Antibiotics 2021, 10, 1388. [Google Scholar] [CrossRef]
- Nógrády, N.; Király, M.; Davies, R.; Nagy, B. Multidrug resistant clones of Salmonella Infantis of broiler origin in Europe. Int. J. Food Microbiol. 2012, 157, 108–112. [Google Scholar] [CrossRef]
- He, T.; Zhu, Y.H.; Yu, J.; Xia, B.; Liu, X.; Yang, G.Y.; Su, J.H.; Guo, L.; Wang, M.L.; Wang, J.F. Lactobacillus johnsonii L531 reduces pathogen load and helps maintain short-chain fatty acid levels in the intestines of pigs challenged with Salmonella enterica Infantis. Vet. Microbiol. 2019, 230, 187–194. [Google Scholar] [CrossRef]
- Schneitz, C.; Koivunen, E.; Tuunainen, P.; Valaja, J. The effects of a competitive exclusion product and two probiotics on Salmonella colonization and nutrient digestibility in broiler chickens. J. Appl. Poult. Res. 2016, 25, 396–406. [Google Scholar] [CrossRef]
- Torok, V.A.; Ophel-Keller, K.; Loo, M.; Hughes, R.J. Application of methods for identifying broiler chicken gut bacterial species linked with increased energy metabolism. Appl. Environ. Microbiol. 2008, 74, 783–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrales-Martinez, J.; Ortega-Paredes, D.; Šefcová, M.A.; Larrea-Álvarez, C.M.; de Janon, S.; Medina-Santana, J.; Molina-Cuasapaz, G.; Vinueza-Burgos, C.; Revajová, V.; Larrea-Álvarez, M.; et al. A PMAxxTM qPCR assay reveals that dietary administration of the Microalgae Tetraselmis chuii does not affect Salmnonella Infantis caecal content in early treated broiler chickens. Vet. Sci. 2022, 9, 487. [Google Scholar] [CrossRef] [PubMed]
- Sardari, R.R.; Karlsson, N.E. Marine poly- and oligosaccharides as prebiotics. J. Agric. Food Chem. 2018, 66, 11544–11549. [Google Scholar] [CrossRef] [PubMed]
- Kulshreshtha, G.; Rathgeber, B.; MacIsaac, J.; Boulianne, M.; Brigitte, L.; Stratton, G.; Prithiviraj, B. Feed supplementation with red seaweeds, Chondrus crispus and Sarcodiotheca gaudichaudii, reduce Salmonella Enteritidis in laying hens. Front. Microbiol. 2017, 8, 567. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yan, X.; Han, D.; Liu, Y.; Song, W.; Tong, T.; Ma, Y. Lactobacillus casei DBN023 protects against jejunal mucosal injury in chicks infected with Salmonella pullorum CMCC-533. Res. Vet. Sci. 2019, 127, 33–41. [Google Scholar] [CrossRef]
- Shao, Y.; Guo, Y.; Wang, Z. β-1, 3/1, 6-Glucan alleviated intestinal mucosal barrier impairment of broiler chickens challenged with Salmonella enterica serovar Typhimurium. Poult. Sci. 2013, 92, 1764–1773. [Google Scholar] [CrossRef]
- Berndt, A.; Wilhelm, A.; Jugert, C.; Pieper, J.; Sachse, K.; Methner, U. Chicken cecum immune response to Salmonella enterica serovars of different levels of invasiveness. Infect. Immun. 2007, 75, 5993–6007. [Google Scholar] [CrossRef] [Green Version]
- Deng, P.; Zhongtang, Y. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar]
- Bao, Y.M.; Choct, M. Dietary NSP nutrition and intestinal immune system for broiler chickens. World’s Poult. Sci. J. 2010, 66, 511–518. [Google Scholar] [CrossRef]
- Deng, Z.; Han, D.; Wang, Y.; Wang, Q.; Yan, X.; Wang, S.; Liu, X.; Song, W.; Ma, Y. Lactobacillus casei protects intestinal mucosa from damage in chicks caused by Salmonella pullorum via regulating immunity and the Wnt signaling pathway and maintaining the abundance of gut microbiota. Poult. Sci. 2021, 100, 101283. [Google Scholar] [CrossRef] [PubMed]
- Abudabos, A.M.; Aljumaah, M.R.; Alkhulaifi, M.M.; Alabdullatif, A.; Suliman, G.M.; AL Sulaiman, A.R. Comparative effects of Bacillus subtilis and Bacillus licheniformis on live performance, blood metabolites and intestinal features in broiler inoculated with Salmonella infection during the finisher phase. Microb. Pathog. 2020, 139, 103870. [Google Scholar] [CrossRef] [PubMed]
- Šefcová, M.A.; Larrea-Álvarez, M.; Larrea-Álvarez, C.M.; Karaffová, V.; Ortega-Paredes, D.; Vinueza-Burgos, C.; Ševčíková, Z.; Levkut, M.; Herich, R.; Revajová, V. The probiotic Lactobacillus fermentum Biocenol CCM 7514 moderates Campylobacter jejuni-induced body weight impairment by improving gut morphometry and regulating cecal cytokine abundance in broiler chickens. Animals 2021, 11, 235. [Google Scholar] [CrossRef] [PubMed]
- Šefcová, M.; Larrea-Álvarez, M.; Larrea-Álvarez, C.; Revajová, V.; Karaffová, V.; Koščová, J.; Nemcová, R.; Ortega-Paredes, D.; Vinueza-Burgos, C.; Levkut, M.; et al. Effects of Lactobacillus fermentum supplementation on body weight and pro-inflammatory cytokine expression in Campylobacter jejuni-challenged chickens. Vet. Sci. 2020, 7, 121. [Google Scholar] [CrossRef] [PubMed]
- Šefcová, M.; Larrea-Álvarez, M.; Larrea-Álvarez, C.; Karaffová, V.; Revajová, V.; Gancarčíková, S.; Ševčíková, Z.; Herich, R. Lactobacillus fermentum administration modulates cytokine expression and lymphocyte subpopulation levels in broiler chickens challenged with Campylobacter coli. Foodborne Pathog. Dis. 2020, 17, 485–493. [Google Scholar] [CrossRef]
- Duangnumsawang, Y.; Zentek, J.; Goodarzi Boroojeni, F. Development and functional properties of intestinal mucus layer in poultry. Front. Immunol. 2021, 12, 745849. [Google Scholar] [CrossRef]
- Bron, P.A.; Van Baarlen, P.; Kleerebezem, M. Emerging molecular insights into the interaction between probiotics and the host intestinal mucosa. Nat. Rev. Microbiol. 2012, 10, 66–78. [Google Scholar] [CrossRef]
- Lazic, S.E.; Clarke-Williams, C.J.; Munaf, M.R. What exactly is ‘N’ in cell culture and animal experiments? PLoS Biol. 2018, 16, e2005282. [Google Scholar] [CrossRef] [Green Version]
- Cobb500 Broiler Performance and Nutrition Supplement. Available online: https://www.cobb-vantress.com/resource/featured?q=nutrition (accessed on 3 September 2021).
- Broiler Management Guide. Available online: https://www.cobb-vantress.com/resource/management-guides (accessed on 3 September 2021).
- Aljumaah, M.R.; Alkhulaifi, M.M.; Abudabos, A.M.; Aljumaah, R.S.; Alsaleh, A.N.; Stanley, D. Bacillus subtilis PB6 based probiotic supplementation plays a role in the recovery after the necrotic enteritis challenge. PLoS ONE 2020, 15, e0232781. [Google Scholar] [CrossRef]
- Liu, T.; Wang, C.; Wu, X.; Ren, M.; Hu, Q.; Jin, E.; Gu, Y. Effect of bron on microstructure, immune function, expression of tight junction protein, cell proliferation and apoptosis of duodenum in rats. Biol. Trace Elem. Res. 2021, 199, 205–215. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Wang, B.; Mei, X.; Jiang, S.; Li, W. Protocatechuic acid improved growth performance, meat quality, and intestinal health of Chinese yellow-feathered broilers. Poult. Sci. 2019, 98, 3138–3149. [Google Scholar] [CrossRef] [PubMed]
- De Boever, S.; Vangestel, C.; De Backer, P.; Croubels, S.; Sys, S.U. Identification and validation of housekeeping genes as internal control for gene expression in an intravenous LPS inflammation model in chickens. Vet. Immunol. Immunopathol. 2008, 122, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Šefcová, M.; Levkut, M.; Bobíková, K.; Karaffová, V.; Revajová, V.; Cingel’ová Maruščáková, I.; Levkutová, M.; Šefčíková, Z.; Herich, R.; Levkut, M. Cytokine response after stimulation of culture cells by zinc and probiotic strain. In Vitro Cell. Dev. Biol. Anim. 2019, 55, 830–837. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kürekci, C.; Sahin, S.; Iwan, E.; Kwit, R.; Bomba, A.; Wasyl, D. Whole-genome sequence analysis of Salmonella Infantis isolated from raw chicken meat samples and insights into pESI-like megaplasmid. Int. J. Food Microbiol. 2021, 337, 108956. [Google Scholar] [CrossRef]
- Martínez-Puchol, S.; Riveros, M.; Ruidias, K.; Granda, A.; Ruiz-Roldán, L.; Zapata-Cachay, C.; Ochoa, T.J.; Pons, M.J.; Ruiz, J. Dissemination of a multidrug resistant CTX-M-65 producer Salmonella enterica serovar Infantis clone between marketed chicken meat and children. Int. J. Food Microbiol. 2021, 344, 109109. [Google Scholar] [CrossRef]
- Antunes, P.; Mourão, J.; Campos, J.; Peixe, L. Salmonellosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Tyson, G.H.; Li, C.; Harrison, L.B.; Martin, G.; Hsu, C.-H.; Tate, H.; Tran, T.; Strain, E.; Zhao, S. A multidrug-resistant Salmonella Infantis clone is spreading and recombining in the United States. Microb. Drug Resist. 2021, 27, 792–799. [Google Scholar] [CrossRef]
- Sabry, M.A.; Abdel-Moein, K.A.; Abdel-Kader, F.; Hamza, E. Extended-spectrum β-lactamase-producing Salmonella serovars among healthy and diseased chickens and their public health implication. J. Glob. Antimicrob. Resist. 2020, 22, 742–748. [Google Scholar] [CrossRef]
- Collett, S.R. Nutrition and wet litter problems in poultry. Anim. Feed Sci. Technol. 2012, 173, 65–75. [Google Scholar] [CrossRef]
- Chen, J.; Tellez, G.; Richards, J.D.; Escobar, J. Identification of potential biomarkers for gut barrier failure in broiler chickens. Front. Vet. Sci. 2015, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Omar, A.E.; Al-Khalaifah, H.S.; Mohamed, W.A.M.; Gharib, H.S.A.; Osman, A.; Al-Gabri, N.A.; Amer, S.A. Effects of phenolic-rich onion (Allium cepa L.) extract on the growth performance, behavior, intestinal histology, amino acid digestibility, antioxidant activity, and the immune status of broiler chickens. Front. Vet. Sci. 2020, 7, 582612. [Google Scholar] [CrossRef]
- Šefcová, M.A.; Santacruz, F.; Larrea-Álvarez, C.M.; Vinueza-Burgos, C.; Ortega-Paredes, D.; Molina-Cuasapaz, G.; Rodríguez, J.; Calero-Cáceres, W.; Revajová, V.; Fernández-Moreira, E.; et al. Administration of dietary microalgae ame-liorates intestinal parameters, improves body weight, and reduces thawing loss of fillets in broiler chickens: A pilot study. Animals 2021, 11, 3601. [Google Scholar] [CrossRef] [PubMed]
- Shini, S.; Zhang, D.; Aland, R.C.; Li, X.; Dart, P.J.; Callaghan, M.J.; Speight, R.E.; Bryden, W.L. Probiotic Bacillus amyloliquefaciens H57 ameliorates subclinical necrotic enteritis in broiler chicks by maintaining intestinal mucosal integrity and improving feed efficiency. Poult. Sci. 2020, 99, 4278–4293. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.W.; El-Nezami, H.; Shah, N.P. The protective effects of enriched citrulline fermented milk with Lactobacillus helveticus on the intestinal epithelium integrity against Escherichia coli infection. Sci. Rep. 2020, 10, 499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Z.; Torres-Fuentes, C.; Heeney, D.D.; Marco, M.L. Synergy between probiotic Lactobacillus casei and milk to maintain barrier integrity of intestinal epithelial cells. J. Agric. Food Chem. 2019, 67, 1955–1962. [Google Scholar] [CrossRef]
- Dagaas, C.T.; Mangubat, K.M.M.; Angeles, A.A.; Abanto, O.D. Dietary effects of commercial probiotics on growth performance, digestibility, and intestinal morphometry of broiler chickens. Trop. Anim. Health Prod. 2019, 51, 1105–1115. [Google Scholar]
- Forte, C.; Manuali, E.; Abbate, Y.; Papa, P.; Vieceli, L.; Tentellini, M.; Trabalza-Marinucci, M.; Moscati, L. Dietary Lactobacillus acidophilus positively influences growth performance, gut morphology, and gut microbiology in rurally reared chickens. Poult. Sci. 2018, 97, 930–936. [Google Scholar] [CrossRef]
- Wang, B.; Gong, L.; Zhou, Y.; Tang, L.; Zeng, Z.; Wang, Q.; Zou, P.; Yu, D.; Li, W. Probiotic Paenibacillus polymyxa 10 and Lactobacillus plantarum 16 enhance growth performance of broilers by improving the intestinal health. Anim. Nutr. 2021, 7, 829–840. [Google Scholar] [CrossRef]
- Jing, L.; Guo, L.; Chen, B.; Hao, K.; Ma, H.; Liu, Y.; Min, Y. Effects of different probiotic fermented feeds on production performance and intestinal health of laying hens. Poult. Sci. 2022, 101, 101570. [Google Scholar]
- Gyawali, I.; Zeng, Y.; Zhou, J.; Li, J.; Wu, T.; Jiang, Q.; Zhu, C. Effect of Novel Lactobacillus paracaesi microcapsule on growth performance, gut health and microbiome community of broiler chickens. Poult. Sci. 2022, 101, 101912. [Google Scholar] [CrossRef] [PubMed]
- Awad, W.A.; Bohm, J.; Razzazi-Fazeli, E.; Ghareeband, K.; Zentek, J. Effect of addition of a probiotic microorganism to broiler diets contaminated with deoxynivalenol on performance and histological alterations of intestinal villi of broiler chickens. Poult. Sci. 2006, 85, 974–979. [Google Scholar] [CrossRef] [PubMed]
- Awad, W.A.; Ghareeb, K.; Abdel-Raheem, S.; Bohm, J. Effects of dietary inclusion of probiotic and synbiotic on growth performance, organ weights, and intestinal histomorphology of broiler chickens. Poult. Sci. 2009, 88, 49–56. [Google Scholar] [CrossRef]
- Bogucka, J.; Ribeiro, D.M.; Bogusławska-Tryk, M.; Dankowiakowska, A.; da Costa, R.P.R.; Bednarczyk, M. Microstructure of the small intestine in broiler chickens fed a diet with probiotic or synbiotic supplementation. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1785–1791. [Google Scholar] [CrossRef] [PubMed]
- Criado-Mesas, L.; Abdelli, N.; Noce, A.; Farré, M.; Pérez, J.F.; Solà-Oriol, D.; Martin-Venegas, R.; Forouzandeh, A.; González-Solé, F.; Folch, J.M. Transversal gene expression panel to evaluate intestinal health in broiler chickens in different challenging conditions. Sci. Rep. 2021, 11, 6315. [Google Scholar] [CrossRef] [PubMed]
- Wilson, F.D.; Cummings, T.S.; Barbosa, T.M.; Williams, C.J.; Gerard, P.D.; Peebles, E.D. Comparison of two methods for determination of intestinal villus to crypt ratios and documentation of early age-associated ratio changes in broiler chickens. Poult. Sci. 2018, 97, 1757–1761. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Matur, E.; Eraslan, E. The impact of probiotics on the gastrointestinal physiology. New Adv. Basic Clin. Gastroenterol. 2012, 1, 51–74. [Google Scholar]
- Mitjans, M.; Barniol, G.; Ferrer, R. Mucosal surface area in chicken small intestine during development. Cell Tissue Res. 1997, 290, 71–78. [Google Scholar] [CrossRef]
- Aljumaah, M.R.; Alkhulaifi, M.M.; Abudabos, A.M.; Alabdullatifb, A.; El-Mubarak, A.H.; Al Suliman, A.R.; Stanley, D. Organic acid blend supplementation increases butyrate and acetate production in Salmonella enterica serovar Typhimurium challenged broilers. PLoS ONE 2020, 15, e0232831. [Google Scholar] [CrossRef]
- Thiam, M.; Barreto Sánchez, A.L.; Zhang, J.; Zheng, M.; Wen, J.; Zhao, G.; Wang, Q. Association of heterophil/lymphocyte ratio with intestinal barrier function and immune response to Salmonella enteritidis infection in chicken. Animals 2021, 11, 3498. [Google Scholar] [CrossRef] [PubMed]
- Elmi, V.A.; Moradi, S.; Harsini, S.G.; Rahimi, M. Effects of Lactobacillus acidophilus and natural antibacterials on growth performance and Salmonella colonization in broiler chickens challenged with Salmonella enteritidis. Livest. Sci. 2020, 233, 103948. [Google Scholar] [CrossRef]
- Wu, H.; Ye, L.; Lu, X.; Xie, S.; Yang, Q.; Yu, Q. Lactobacillus acidophilus alleviated Salmonella-induced goblet cells loss and colitis by notch pathway. Mol. Nutr. Food Res. 2018, 62, 1800552. [Google Scholar] [CrossRef]
- Kong, S.; Zhang, Y.H.; Zhang, W. Regulation of intestinal epithelial cells properties and functions by amino acids. Biomed Res Int. 2018, 9, 2819154. [Google Scholar] [CrossRef]
- Johansson, M.E.; Hansson, G.C. Immunological aspects of intestinal mucus and mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar] [CrossRef]
- McGuckin, M.A.; Lindén, S.K.; Sutton, P.; Florin, T.H. Mucin dynamics and enteric pathogens. Nat. Rev. Microbiol. 2011, 9, 265–278. [Google Scholar] [CrossRef]
- Zhen, W.; Shao, Y.; Gong, X.; Wu, Y.; Geng, Y.; Wang, Z.; Guo, Y. Effect of dietary Bacillus coagulans supplementation on growth performance and immune responses of broiler chickens challenged by Salmonella enteritidis. Poult. Sci. 2018, 97, 2654–2666. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Zhao, S.; Jiang, L.; Lu, L.; Yang, Q.; Yu, Q. Lactobacillus reuteri stimulates intestinal epithelial proliferation and induces differentiation into goblet cells in young chickens. J. Agric. Food Chem. 2019, 67, 13758–13766. [Google Scholar] [CrossRef]
- Liu, J.; Gu, Z.; Song, F.; Zhang, H.; Zhao, J.; Chen, W. Lactobacillus plantarum ZS2058 and Lactobacillus rhamnosus GG use different mechanisms to prevent Salmonella infection in vivo. Front. Microbiol. 2019, 10, 299. [Google Scholar] [CrossRef] [Green Version]
- Salim, H.M.; Kim, W.K.; Kim, D.W.; Jong, H.B.; Akter, N.; Kang, H.K. Supplementation of direct-fed microbials as an alternative to antibiotic on growth performance, immune response, cecal microbial population, and ileal morphology of broiler chickens. Poult. Sci. 2013, 92, 2084–2090. [Google Scholar] [CrossRef]
- Liu, J.; Hu, D.; Chen, Y.; Huang, H.; Zhang, H.; Zhao, J.; Gu, Z.; Chen, W. Strain-specific properties of Lactobacillus plantarum for prevention of Salmonella infection. Food Funct. 2018, 9, 3673–3682. [Google Scholar] [CrossRef] [PubMed]
- Havelaar, A.H.; Van Pelt, W.; Ang, C.W.; Wagenaar, J.A.; Van Putten, J.P.M.; Gross, U.; Newell, D.G. Immunity to Campylobacter: Its role in risk assessment and epidemiology. Crit. Rev. Microbiol. 2009, 35, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Mountzouris, K.C.; Balaskas, C.; Xanthakos, I.; Tzivinikou, A.; Fegeros, K. Effects of a multi-species probiotic on biomarkers of competitive exclusion efficacy in broilers challenged with Salmonella enteritidis. Br. Poult. Sci. 2009, 50, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Mountzouris, K.C.; Tsitrsikos, P.; Palamidi, I.; Arvaniti, A.; Mohnl, M.; Schatzmayr, G.; Fegeros, K. Effects of probiotic inclusion levels in broiler nutrition on growth performance, nutrient digestibility, plasma immunoglobulins, and cecal microflora composition. Poult. Sci. 2010, 89, 58–67. [Google Scholar] [CrossRef]
- European Food Safety Authority [EFSA]. Opinion of the scientific panel on biological hazards on a request from the Commission related to the use of vaccines for the control of salmonella in poultry. EFSA J. 2004, 114, 1–74. [Google Scholar]



{kind=link}
{kind=link}
| Expression Levels (2−ΔCt) | ||||
|---|---|---|---|---|
| Gene symbol | Control | L. fermentum | S. Infantis | L. fermentum + S. Infantis |
| muc-2 | 0.107 ± 0.216 | 0.256 ± 0.613 | 0.159 ± 0.348 | 0.455 ± 1.262 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šefcová, M.A.; Ortega-Paredes, D.; Larrea-Álvarez, C.M.; Mina, I.; Guapás, V.; Ayala-Velasteguí, D.; Leoro-Garzón, P.; Molina-Cuasapaz, G.; Vinueza-Burgos, C.; Revajová, V.; et al. Effects of Lactobacillus fermentum Administration on Intestinal Morphometry and Antibody Serum Levels in Salmonella-Infantis-Challenged Chickens. Microorganisms 2023, 11, 256. https://doi.org/10.3390/microorganisms11020256
Šefcová MA, Ortega-Paredes D, Larrea-Álvarez CM, Mina I, Guapás V, Ayala-Velasteguí D, Leoro-Garzón P, Molina-Cuasapaz G, Vinueza-Burgos C, Revajová V, et al. Effects of Lactobacillus fermentum Administration on Intestinal Morphometry and Antibody Serum Levels in Salmonella-Infantis-Challenged Chickens. Microorganisms. 2023; 11(2):256. https://doi.org/10.3390/microorganisms11020256
Chicago/Turabian StyleŠefcová, Miroslava Anna, David Ortega-Paredes, César Marcelo Larrea-Álvarez, Iván Mina, Victoria Guapás, David Ayala-Velasteguí, Paula Leoro-Garzón, Gabriel Molina-Cuasapaz, Christian Vinueza-Burgos, Viera Revajová, and et al. 2023. "Effects of Lactobacillus fermentum Administration on Intestinal Morphometry and Antibody Serum Levels in Salmonella-Infantis-Challenged Chickens" Microorganisms 11, no. 2: 256. https://doi.org/10.3390/microorganisms11020256
APA StyleŠefcová, M. A., Ortega-Paredes, D., Larrea-Álvarez, C. M., Mina, I., Guapás, V., Ayala-Velasteguí, D., Leoro-Garzón, P., Molina-Cuasapaz, G., Vinueza-Burgos, C., Revajová, V., & Larrea-Álvarez, M. (2023). Effects of Lactobacillus fermentum Administration on Intestinal Morphometry and Antibody Serum Levels in Salmonella-Infantis-Challenged Chickens. Microorganisms, 11(2), 256. https://doi.org/10.3390/microorganisms11020256