




Effects of the Long-Term Continuous Cropping of Yongfeng Yam on the Bacterial Community and Function in the Rhizospheric Soil

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Study Area and Experimental Design
2.2. Sample Collection and Soil Chemical Properties Detection
2.3. Analysis of Soil Chemical Properties
2.4. DNA Extraction, PCR Amplification, and High-Throughput Sequencing
2.5. Sequence Analysis and Statistical Analysis
3. Results
3.1. Chemical Properties and Enzyme Activities of Rhizospheric Soil
3.2. General Analysis of Sequencing Data
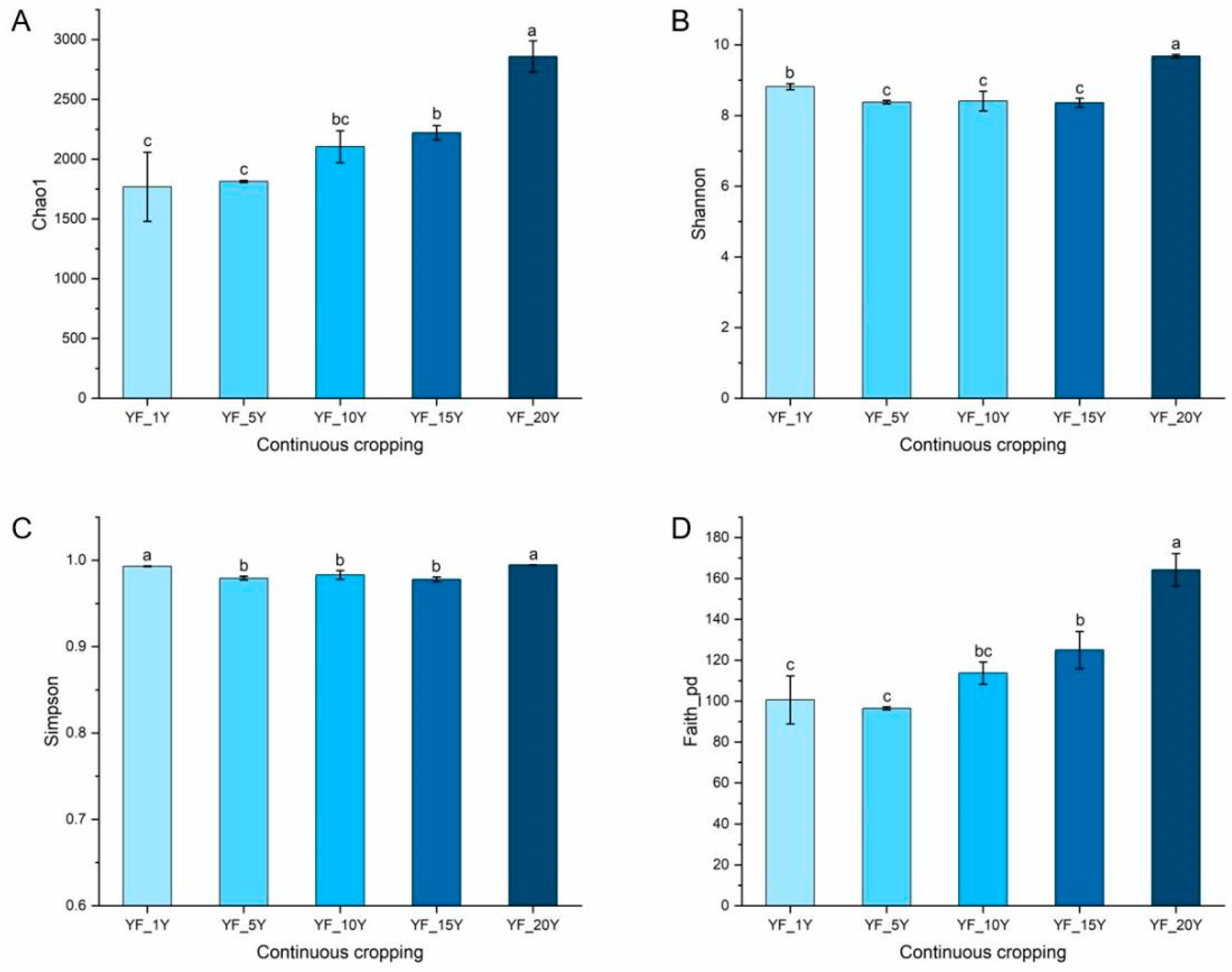
3.3. Alpha Diversity and Beta Diversity Analysis
3.4. Taxonomic Composition Analysis
3.5. Differences in Microbial Community among Different Continuous Cultivation Years
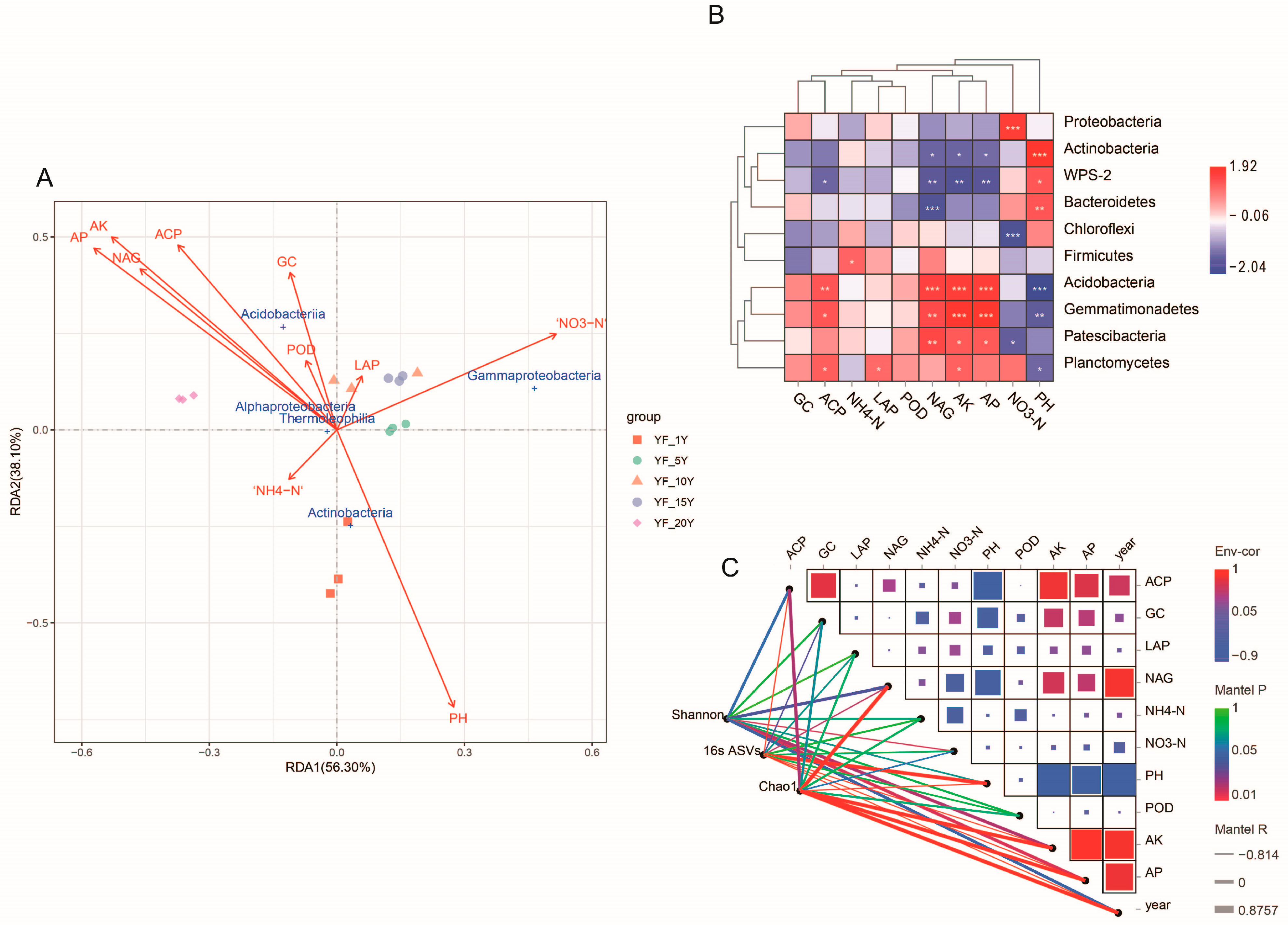
3.6. Relationship between Environmental Factors and Bacterial Community Structure
3.7. Network Analysis of Soil Bacterial Community
3.8. Prediction of Bacterial Community Function
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kolombia, Y.A.; Ogundero, O.; Olajide, E.; Viaene, N.; Kumar, P.L.; Coyne, D.L.; Bert, W. Morphological and molecular characterization of Pratylenchus species from Yam (Dioscorea spp.) in West Africa. J. Nematol. 2021, 52, e2020–e2126. [Google Scholar] [CrossRef] [PubMed]
- Fang, D.Z.; Liu, X.L.; Chen, X.R.; Yan, W.W.; He, Y.L.; Cheng, Y.; Chen, J.; Li, Z.M.; Guo, L.T.; Wang, T.H.; et al. Fusarium species and Fusarium oxysporum species complex genotypes associated with yam wilt in south-central China. Front. Microbiol. 2020, 11, 1964. [Google Scholar]
- Price, E.J.; Bhattacharjee, R.; Lopez-Montes, A.; Fraser, P.D. Carotenoid profiling of yams: Clarity, comparisons and diversity. Food Chem. 2018, 259, 130–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukal, A.; Kidanemariam, D.; Dale, J.; James, A.; Harding, R. Characterization of badnaviruses infecting Dioscorea spp. in the Pacific reveals two putative novel species and the first report of dioscorea bacilliform RT virus 2. Virus Res. 2017, 238, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, B.; Zhang, Y.M.; Sun, X.Q.; Li, M.M.; Xue, J.Y.; Hang, Y.Y. Genetic relationship and identification of Dioscorea polystachya cultivars accessed by ISAP and SCAR markers. Arch. Biol. Sci. 2017, 69, 277–284. [Google Scholar] [CrossRef]
- Gao, G.D.; Zhao, B. Discussion on the status of Chinese yam industry. Crop Res. 2007, 21, 179–181. [Google Scholar]
- Ju, Y.; Xue, Y.; Huang, J.L.; Zhai, Q.Z.; Wang, X.H. Antioxidant Chinese yam polysaccharides and its pro-proliferative effect on endometrial epithelial cells. Int. J. Biol. Macromol. 2014, 66, 81–85. [Google Scholar] [CrossRef]
- Chen, Y.F.; Zhu, Q.; Wu, S.J. Preparation of oligosaccharides from Chinese yam and their antioxidant activity. Food Chem. 2015, 173, 1107–1110. [Google Scholar] [CrossRef]
- Epping, J.; Laibach, N. An underutilized orphan tuber crop—Chinese yam: A review. Planta 2020, 252, 58. [Google Scholar] [CrossRef]
- Gao, Z.Y.; Hu, Y.Y.; Han, M.K.; Xu, J.J.; Wang, X.; Liu, L.F.; Tang, Z.H.; Jiao, W.J.; Jin, R.; Liu, M.; et al. Effects of continuous cropping of sweet potatoes on the bacterial community structure in rhizospheric soil. BMC Microbiol. 2021, 21, 102. [Google Scholar] [CrossRef]
- Ma, W.Y.; Liao, X.L.; Wang, C.; Zhang, Y. Effects of four cropping patterns of Lilium brownie on rhizosphere microbiome structure and replant disease. Plants 2022, 11, 824. [Google Scholar] [CrossRef]
- Liu, X.J.; Zhang, S.T.; Jiang, Q.P.; Bai, Y.N.; Shen, G.H.; Li, S.L.; Ding, W. Using community analysis to explore bacterial indicators for disease suppression of tobacco bacterial wilt. Sci. Rep. 2016, 6, 36773. [Google Scholar] [CrossRef]
- Zheng, X.F.; Wang, Z.R.; Zhu, Y.J.; Wang, J.P.; Liu, B. Effects of a microbial restoration substrate on plant growth and rhizosphere bacterial community in a continuous tomato cropping greenhouse. Sci. Rep. 2020, 10, 13729. [Google Scholar] [CrossRef]
- Pang, Z.Q.; Tayyab, M.; Kong, C.B.; Liu, Q.; Liu, Y.M.; Hu, C.H.; Huang, J.W.; Weng, P.Y.; Islam, W.; Lin, W.X.; et al. Continuous sugarcane planting negatively impacts soil microbial community structure, soil fertility, and sugarcane agronomic parameters. Microorganisms 2021, 9, 2008. [Google Scholar] [CrossRef]
- Wang, S.; Cheng, J.; Li, T.; Liao, Y. Response of soil fungal communities to continuous cropping of flue-cured tobacco. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Chen, P.; Wang, Y.-Z.; Liu, Q.-Z.; Zhang, Y.-T.; Li, X.-Y.; Li, H.-Q.; Li, W.-H. Phase changes of continuous cropping obstacles in strawberry (Fragaria × ananassa Duch.) production. Appl. Soil Ecol. 2020, 155, 103626. [Google Scholar] [CrossRef]
- Li, S.; Xu, C.; Wang, J.; Guo, B.; Yang, L.; Chen, J.; Ding, W. Cinnamic, myristic and fumaric acids in tobacco root exudates induce the infection of plants by Ralstonia solanacearum. Plant Soil 2017, 412, 381–395. [Google Scholar] [CrossRef]
- Xie, X.G.; Zhao, Y.Y.; Yang, Y.; Lu, F.; Da, C.C. Endophytic Fungus Alleviates Soil Sickness in Peanut Crops by Improving the Carbon Metabolism and Rhizosphere Bacterial Diversity. Microb. Ecol. 2021, 82, 49–61. [Google Scholar] [CrossRef]
- Zhu, Y.B.; Wu, X.G.; Xin, H.W.; Wang, C.Y.; Xiong, L.; Zhang, D.H. Occurrence characteristics of main diseases and insect pests of organic yam in Wanzai county and key technology of comprehensive control. North. Hort. 2016, 21, 206–208. [Google Scholar]
- Ali, A.; Ghani, M.I.; Elrys, A.S.E.; Ding, H.Y.; Iqbal, M.; Cheng, Z.H.; Cai, Z.C. Different cropping systems regulate the metabolic capabilities and potential ecological functions altered by soil microbiome structure in the plastic shed mono-cropped cucumber rhizosphere. Agr. Ecosyst. Environ. 2021, 318, 107486. [Google Scholar] [CrossRef]
- Watanabe, F.S.; Olsen, S.R. Test of an ascorbic acid method for determining phosphorus in water and NaHCO3 extracts from soil. Soil Sci. Soc. Am. J. 1965, 22, 677–678. [Google Scholar] [CrossRef]
- Knudsen, D.; Peterson, G.; Pratt, P. Lithium, sodium, and potassium. In Methods of Soil Analysis, Part 2, Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Kenney, D.R., Eds.; American Society of Agronomy, Soil Science Society of American: Madison, WI, USA, 1982; pp. 225–246. [Google Scholar]
- Zhou, X.; Liu, J.; Wu, F. Soil microbial communities in cucumber monoculture and rotation systems and their feedback effects on cucumber seedling growth. Plant Soil 2017, 415, 507–520. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. Dada2: High-resolution sample inference from illumina amplicon data. Nat. Methods. 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. Mafft: A novel method for rapid multiple sequence alignment based on fast fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [Green Version]
- Zaura, E.; Keijser, B.J.F.; Huse, S.M.; Crielaard, W. Defining the healthy “core microbiome” of oral microbial communities. BMC Microbiol. 2009, 9, 259. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Chen, W.J.; Guo, X.D.; Guo, Q.N.; Tan, X.L.; Wang, Z.G. Long-term chili monoculture alters environmental variables affecting the dominant microbial community in rhizosphere soil. Front. Microbiol. 2021, 12, 681953. [Google Scholar]
- Li, C.C.; Gan, Y.D.; Zhang, C.; He, H.; Fang, J.H.; Wang, L.F.; Wang, Y.; Liu, J. “Microplastic communities” in different environments: Differences, links, and role of diversity index in source analysis. Water Res. 2021, 188, 116574. [Google Scholar] [CrossRef]
- Qin, S.; Yeboah, S.; Cao, L.; Zhang, J.; Shi, S.; Liu, Y. Breaking continuous potato cropping with legumes improves soil microbial communities, enzyme activities and tuber yield. PLoS ONE 2017, 12, e0175934. [Google Scholar] [CrossRef] [Green Version]
- Li, H.Y.; Li, C.H.; Song, X.; Liu, Y.; Gao, Q.X.; Zheng, R.; Li, J.T.; Zhang, P.C.; Liu, L. Impacts of continuous and rotational cropping practices on soil chemical properties and microbial communities during peanut cultivation. Sci. Rep. 2022, 12, 2758. [Google Scholar] [CrossRef]
- Li, J.R.; Chen, X.Z.; Li, S.M.; Zuo, Z.M.; Zhan, R.T.; He, R. Variations of rhizospheric soil microbial communities in response to continuous Andrographis paniculata cropping practices. Bot. Stud. 2020, 61, 18. [Google Scholar] [CrossRef]
- Wu, L.K.; Chen, J.; Xiao, Z.G.; Zhu, X.C.; Wang, J.Y.; Wu, H.M.; Wu, Y.H.; Zhang, Z.Y.; Lin, W.X. Barcoded pyrosequencing reveals a shift in the bacterial community in the rhizosphere and rhizoplane of Rehmannia glutinosa under consecutive monoculture. Int. J. Mol. Sci. 2018, 19, 850. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, J.Q.; Liu, Q.; Zhou, Z.F.; Chen, F.L.; Xiang, D. Effects of consecutive monoculture of sweet potato on soil bacterial community as determined by pyrosequencing. J. Basic. Microbiol. 2019, 59, 181–191. [Google Scholar] [CrossRef]
- Dong, L.; Xu, J.; Feng, G.; Li, X.; Chen, S. Soil bacterial and fungal community dynamics in relation to Panax notoginseng death rate in a continuous cropping system. Sci. Rep. 2016, 6, 31802. [Google Scholar] [CrossRef] [Green Version]
- Na, X.F.; Zheng, G.Q.; Xing, Z.C.; Ma, J.P.; Li, Z.H.; Lu, J.H.; Ma, F. Effects of monocropping on diversity and structure of bacterial community in rhizosphere of replanted Lycium barbarum L. Acta Pedol. Sin. 2017, 54, 1280–1292. [Google Scholar]
- Zhao, J.; Zhang, D.; Yang, Y.Q.; Pan, Y.; Zhao, D.M.; Zhu, J.; Zhang, L.K.; Yang, Z.H. Dissecting the effect of continuous cropping of potato on soil bacterial communities as revealed by high-throughput sequencing. PLoS ONE 2020, 15, e0233356. [Google Scholar] [CrossRef]
- Xuan, D.; Guong, V.; Rosling, A.; Alström, S. Different crop rotation systems as drivers of change in soil bacterial community structure and yield of rice, Oryza sativa. Biol. Fert. Soils 2012, 48, 217–225. [Google Scholar] [CrossRef]
- Acosta-Martínez, V.; Dowd, S.; Sun, Y.; Allen, V. Tag-encoded pyrosequencing analysis of bacterial diversity in a single soil type as affected by management and land use. Soil Biol. Biochem. 2008, 40, 2762–2770. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Fu, W.J.; Hu, C.W.; Chen, G.G.; Xiao, Z.W.; Chen, Y.R.; Wang, Z.J.; Cheng, H.Y. Variation of rhizosphere microbial community in continuous mono-maize seed production. Sci. Rep. 2021, 11, 1544. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Xu, Y.X.; Li, H.; Li, S.S.; Wang, X.L.; Chai, H. Difference of bacterial community structure in the Meadow, Maize, and continuous cropped Alfalfa in Northeast China. Front. Microbiol. 2022, 13, 794848. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.Z.; Li, L.; Jie, W.G.; Cai, B.Y. Effect of continuous cropping on bacterial flora structure in soybean rhizosphere soil. Biotechnol. Bull. 2018, 34, 230–238. [Google Scholar]
- Xiong, W.; Li, Z.G.; Liu, H.J.; Xue, C.; Zhang, R.F.; Wu, H.S.; Li, R.; Shen, Q.R. The effect of long-term continuous cropping of black pepper on soil bacterial communities as determined by 454 pyrosequencing. PLoS ONE 2015, 10, e0136946. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classifcation of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Arafat, Y.; Wei, X.; Jiang, Y.; Chen, T.; Saqib, H.A.S.; Lin, S.; Lin, W. Spatial distribution patterns of root-associated bacterial communities mediated by root exudates in different aged ratooning tea monoculture systems. Int. J. Mol. Sci. 2017, 18, 1727. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.Y.; Zhang, Y.; Xiao, J.L.; Wei, L.; Liang, Z.H. Regulation of soil microbial community structures and watermelon Fusarium wilt by using bio-organic fertilizer. Acta Microbiol. Sin. 2019, 59, 2323–2333. [Google Scholar]
- Liu, Y.L.; Cai, L.Q.; Zhao, R.; Li, H.L.; Wu, J.; Ma, F.J.; Lei, L.J.; Meng, Q.X. Response of soil nutrient and enzyme activity to continuous cropping years of Turpan Hami melon. Soils Fertil. Sci. China 2021, 1, 273–281. [Google Scholar]
- Yang, M.; Pei, W.H.; Dong, J.H.; Pan, K.H.; Wu, D.X.; Yu, L. Effects of konjac continuous cropping on soil microbial quantity and enzyme activity. North. Hortic. 2018, 23, 110–115. [Google Scholar]
- Shao, S.; Chen, M.; Liu, W.; Hu, X.; Li, Y. Long-term monoculture reduces the symbiotic rhizobial biodiversity of peanut. Syst. Appl. Microbiol. 2020, 43, 126101. [Google Scholar] [CrossRef]
- Rao, D.; Meng, F.F.; Yan, X.Y.; Zhang, M.H.; Yao, X.D.; Kim, K.S.; Zhao, J.; Qiu, Q.; Xie, F.; Zhang, W. Changes in Soil Microbial Activity, Bacterial Community Composition and Function in a Long-Term Continuous Soybean Cropping System After Corn Insertion and Fertilization. Front. Microbiol. 2021, 12, 638236. [Google Scholar] [CrossRef]
- Feng, K.; Zhang, Z.J.; Cai, W.W.; Liu, W.Z.; Xu, M.Y.; Yin, H.Q.; Wang, A.J.; He, Z.L.; Deng, Y. Biodiversity and species competition regulate the resilience of microbial biofilm community. Mol. Ecol. 2017, 26, 6170–6182. [Google Scholar] [CrossRef]
- Chen, T.; Hu, R.W.; Zheng, Z.Y.; Yang, J.Y.; Fan, H.; Deng, X.Q.; Yao, W.; Wang, Q.M.; Peng, S.G.; Li, J. Soil bacterial community in the multiple cropping system increased grain yield within 40 cultivation years. Front. Plant Sci. 2021, 12, 804527. [Google Scholar] [CrossRef]
- Coyte, K.Z.; Schluter, J.; Foster, K.R. The ecology of the microbiome: Networks, competition, and stability. Science 2015, 350, 663–666. [Google Scholar] [CrossRef]
- Ghoul, M.; Mitri, S. The ecology and evolution of microbial competition. Trends Microbiol. 2016, 24, 833–845. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| YF_1Y | YF_5Y | YF_10Y | YF_15Y | YF_20Y | |
|---|---|---|---|---|---|
| pH | 5.1 ± 0.04 a | 4.5 ± 0.09 b | 4.4 ± 0.04 bc | 4.3 ± 0.04 c | 4.1 ± 0.04 d |
| EC (μS/cm) | 64.03 ± 2.15 d | 109.00 ± 1.63 a | 71.50 ± 0.60 c | 94.46 ± 0.99 b | 54.501.25 e |
| AP (mg/g) | 1.74 ± 1.01 c | 4.68 ± 0.78 bc | 3.73 ± 0.65 bc | 5.15 ± 1.25 b | 9.31 ± 0.78 a |
| AK (mg/kg) | 82.16 ± 8.57 d | 189.59 ± 10.07 b | 153.09 ± 10.14 c | 198.49 ± 14.93 b | 316.32 ± 7.85 a |
| NH4-N(μg/g) | 15.75 ± 4.96 a | 12.77 ± 0.88 a | 11.54 ± 0.61 a | 17.34 ± 1.38 a | 15.45 ± 0.46 a |
| NO3-N(μg/g) | 13.86 ± 4.48 b | 39.71 ± 0.82 a | 23.61 ± 1.73 b | 23.81 ± 7.39 b | 12.65 ± 1.48 b |
| GC (U/g) | 0.18 ± 0.12 c | 2.00 ± 0.31 a | 1.29 ± 0.65 ab | 0.57 ± 0.05 bc | 1.76 ± 0.09 a |
| POD (U/g) | 1.58 ± 0.93 a | 1.40 ± 0.82 a | 2.84 ± 2.20 a | 1.91 ± 0.46 a | 2.00 ± 0.26 a |
| LAP (U/g) | 1.31 ± 0.69 a | 2.40 ± 1.25 a | 0.95 ± 0.40 a | 2.59 ± 1.00 a | 1.90 ± 0.89 a |
| ACP (U/g) | 5.09 ± 0.09 d | 6.10 ± 0.29 ab | 5.60 ± 0.09 c | 5.71 ± 0.06 bc | 6.36 ± 0.02 a |
| NAG (U/g) | 0.40 ± 0.19 c | 0.17 ± 0.10 c | 0.93 ± 0.11 bc | 1.55 ± 0.34 ab | 1.90 ± 0.52 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, J.; Wu, C.; Fan, L.; Kang, M.; Liu, Z.; Huang, Y.; Xu, X.; Yao, Y. Effects of the Long-Term Continuous Cropping of Yongfeng Yam on the Bacterial Community and Function in the Rhizospheric Soil. Microorganisms 2023, 11, 274. https://doi.org/10.3390/microorganisms11020274
Yao J, Wu C, Fan L, Kang M, Liu Z, Huang Y, Xu X, Yao Y. Effects of the Long-Term Continuous Cropping of Yongfeng Yam on the Bacterial Community and Function in the Rhizospheric Soil. Microorganisms. 2023; 11(2):274. https://doi.org/10.3390/microorganisms11020274
Chicago/Turabian StyleYao, Jian, Caiyun Wu, Linjuan Fan, Meihua Kang, Zirong Liu, Yuhui Huang, Xueliang Xu, and Yingjuan Yao. 2023. "Effects of the Long-Term Continuous Cropping of Yongfeng Yam on the Bacterial Community and Function in the Rhizospheric Soil" Microorganisms 11, no. 2: 274. https://doi.org/10.3390/microorganisms11020274
APA StyleYao, J., Wu, C., Fan, L., Kang, M., Liu, Z., Huang, Y., Xu, X., & Yao, Y. (2023). Effects of the Long-Term Continuous Cropping of Yongfeng Yam on the Bacterial Community and Function in the Rhizospheric Soil. Microorganisms, 11(2), 274. https://doi.org/10.3390/microorganisms11020274