
Effects of Coconut Oil and Palm Oil on Growth, Rumen Microbiota, and Fatty Acid Profile of Suckling Calves



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care
2.2. Animals, Treatments, and Management
2.3. Growth Performance and Feed Intake
2.4. Blood Sampling and Analysis
2.5. Ruminal Chyme Fermentation Parameters and Chyme Enzyme Activity
2.6. Analysis of Microbiota
2.7. Sequencing Data Analysis
2.8. Fatty Acid Profile Measurements
2.9. Statistical Analysis
3. Results
3.1. Animal Growth Performance and Health
3.2. Serum Parameters
3.3. Rumen Chyme Fermentation Parameters and Chyme Digestive Enzyme Activities
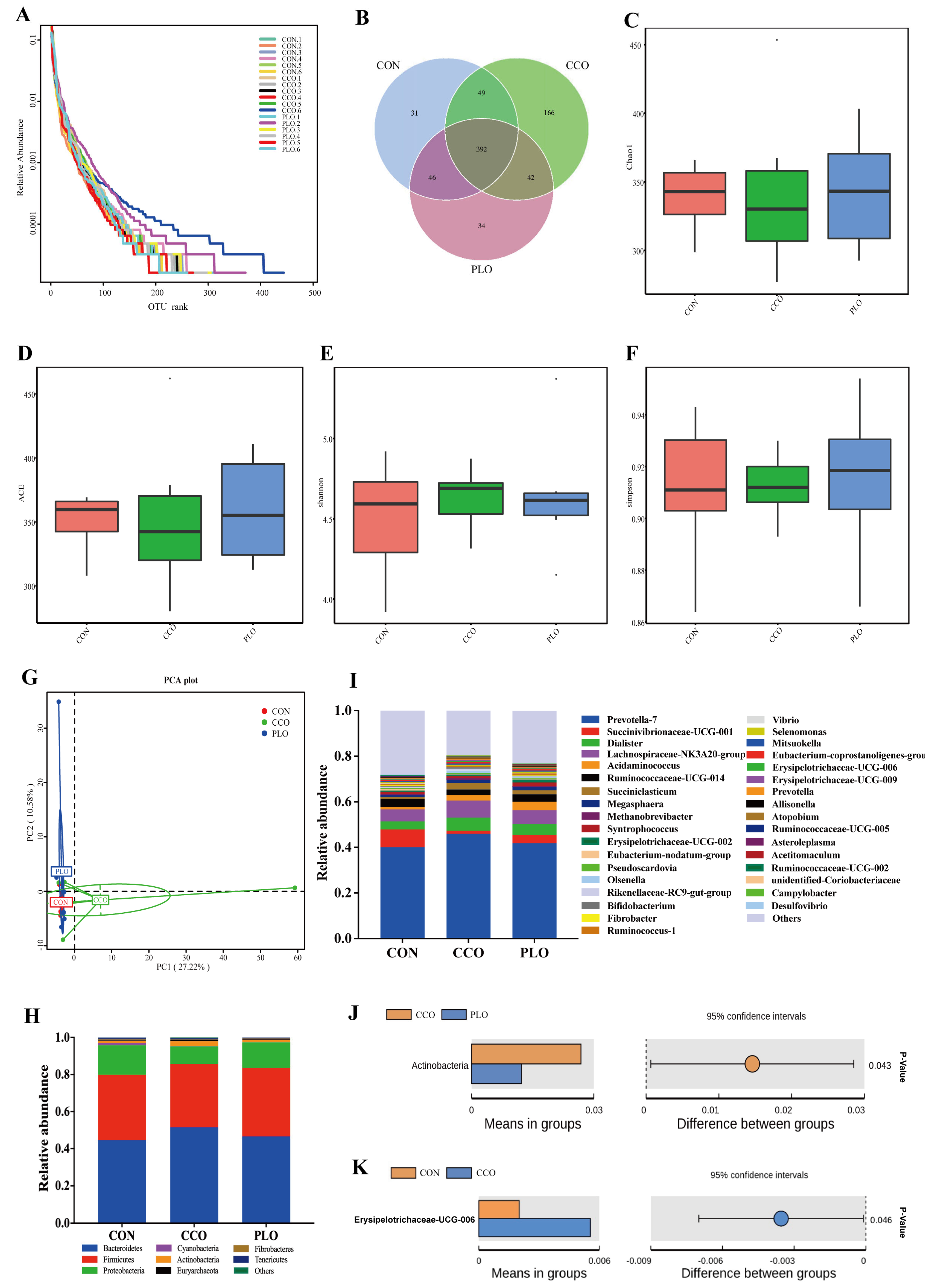
3.4. Community Structure and Composition of Rumen Microbiota
3.5. Fatty Acid Profile of Liver and Muscle Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Santos, H.O.; Howell, S.; Earnest, C.P.; Teixeira, F.J. Coconut oil intake and its effects on the cardiometabolic profile—A structured literature review. Prog. Cardiovasc Dis. 2019, 62, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Soyeurt, H.; Dehareng, F.; Gengler, N.; McParland, S.; Wall, E.; Berry, D.P.; Coffey, M.; Dardenne, P. Mid-infrared prediction of bovine milk fatty acids across multiple breeds, production systems, and countries. J. Dairy Sci. 2011, 94, 1657–1667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghasemi, E.; Azad-Shahraki, M.; Khorvash, M. Effect of different fat supplements on performance of dairy calves during cold season. J. Dairy Sci. 2017, 100, 5319–5328. [Google Scholar] [CrossRef] [PubMed]
- Rajaraman, V.; Nonnecke, B.J.; Horst, R.L. Effects of replacement of native fat in colostrum and milk with coconut oil on fat-soluble vitamins in serum and immune function in calves. J. Dairy Sci. 1997, 80, 2380–2390. [Google Scholar] [CrossRef]
- Huuskonen, A.; Khalili, H.; Kiljala, J.; Joki-Tokola, E.; Nousiainen, J. Effects of vegetable fats versus lard in milk replacers on feed intake, digestibility, and growth in Finnish Ayrshire bull calves. J. Dairy Sci. 2005, 88, 3575–3581. [Google Scholar] [CrossRef] [Green Version]
- Mills, J.K.; Ross, D.A.; Van Amburgh, M.E. The effects of feeding medium-chain triglycerides on the growth, insulin responsiveness, and body composition of Holstein calves from birth to 85 kg of body weight. J. Dairy Sci. 2010, 93, 4262–4273. [Google Scholar] [CrossRef] [Green Version]
- Bowen Yoho, W.S.; Swank, V.A.; Eastridge, M.L.; O’Diam, K.M.; Daniels, K.M. Jersey calf performance in response to high-protein, high-fat liquid feeds with varied fatty acid profiles: Intake and performance. J. Dairy Sci. 2013, 96, 2494–2506. [Google Scholar] [CrossRef] [Green Version]
- Panahiha, P.; Mirzaei-Alamouti, H.; Kazemi-Bonchenari, M.; Aschenbach, J.R. Growth performance, nutrient digestibility, and ruminal fermentation of dairy calves fed starter diets with alfalfa hay versus corn silage as forage and soybean oil versus palm fatty acids as fat source. J. Dairy Sci. 2022, 105, 9597–9609. [Google Scholar] [CrossRef]
- Ragionieri, L.; Cacchioli, A.; Ravanetti, F.; Botti, M.; Ivanovska, A.; Panu, R.; Righi, F.; Quarantelli, A.; Gazza, F. Effect of the supplementation with a blend containing short and medium chain fatty acid monoglycerides in milk replacer on rumen papillae development in weaning calves. Ann. Anat. 2016, 207, 97–108. [Google Scholar] [CrossRef]
- Gorka, P.; Kowalski, Z.M.; Pietrzak, P.; Kotunia, A.; Jagusiak, W.; Holst, J.J.; Guilloteau, P.; Zabielski, R. Effect of method of delivery of sodium butyrate on rumen development in newborn calves. J. Dairy Sci. 2011, 94, 5578–5588. [Google Scholar] [CrossRef] [Green Version]
- Kadkhoday, A.; Riasi, A.; Alikhani, M.; Dehghan-Banadaky, M.; Kowsar, R. Effects of fat sources and dietary C-18:2 to C-18:3 fatty acids ratio on growth performance, ruminal fermentation and some blood components of Holstein calves. Livest. Sci. 2017, 204, 71–77. [Google Scholar] [CrossRef]
- Tadeu Dos Santos, G.; Toullec, R.; Roger, R.; de la Grange, H.; Guilloteau, P. Caractéristiques digestives des veaux de boucherie s’adaptant mal en atelier d’engraissement. Reprod. Nutr. Développement 1986, 26, 1217. [Google Scholar] [CrossRef]
- Abe, M.; Iriki, T.; Kondoh, K.; Shibui, H. Effects of nipple or bucket feeding of milk-substitute on rumen by-pass and on rate of passage in calves. Br. J. Nutr. 1979, 41, 175–181. [Google Scholar] [CrossRef]
- Guilhermet, R.; Mathieu, C.M.; Toullec, R.; Beaufils, M.; Mansion, M.; Manis, Y. Transit Des Aliments Liquides Au Niveau De La GouttiÈre Œsophagienne Chez Le Veau PrÉruminant Et Ruminant. Ann. De Zootech. 1975, 24, 69–79. [Google Scholar] [CrossRef]
- Berends, H.; van den Borne, J.J.; Stockhofe-Zurwieden, N.; Gilbert, M.S.; Zandstra, T.; Pellikaan, W.F.; van Reenen, C.G.; Bokkers, E.A.; Gerrits, W.J. Effects of solid feed level and roughage-to-concentrate ratio on ruminal drinking and passage kinetics of milk replacer, concentrates, and roughage in veal calves. J. Dairy Sci. 2015, 98, 5621–5629. [Google Scholar] [CrossRef] [Green Version]
- Fattore, E.; Fanelli, R. Palm oil and palmitic acid: A review on cardiovascular effects and carcinogenicity. Int J Food Sci Nutr 2013, 64, 648–659. [Google Scholar] [CrossRef]
- Marten, B.; Pfeuffer, M.; Schrezenmeir, J. Medium-chain triglycerides. Int. Dairy J. 2006, 16, 1374–1382. [Google Scholar] [CrossRef]
- Guillot, E.; Vaugelade, P.; Lemarchal, P.; Rerat, A. Intestinal absorption and liver uptake of medium-chain fatty acids in non-anaesthetized pigs. Br. J. Nutr. 1993, 69, 431–442. [Google Scholar] [CrossRef]
- Erwin, E.S.; Sterner, W. Dietary Lipids and Fatty Acid Deposition in Various Calf Tissues. Am. J. Physiol. 1963, 205, 1151–1153. [Google Scholar] [CrossRef]
- Jenkins, K.J.; Kramer, J.K.G. Differential effects of dietary fatty acids on fatty acid composition of phosphotidylinositol in calf tissues. Nutr. Res. 1991, 11, 177–183. [Google Scholar] [CrossRef]
- Jenkins, K.J.; Kramer, J.K. Effects of dietary corn oil and fish oil concentrate on lipid composition of calf tissues. J. Dairy Sci. 1990, 73, 2940–2951. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.; Greco, L.F.; Lock, A.L.; Block, E.; Santos, J.E.P.; Thatcher, W.W.; Staples, C.R. Supplementation of essential fatty acids to Holstein calves during late uterine life and first month of life alters hepatic fatty acid profile and gene expression. J. Dairy Sci. 2016, 99, 7085–7101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Zhang, W.-B.; Bi, Y.-l.; Tu, Y.; Beckers, Y.; Du, H.-C.; Diao, Q.-Y. Early Feeding Regime of Waste Milk, Milk, and Milk Replacer for Calves Has Different Effects on Rumen Fermentation and the Bacterial Community. Animals 2019, 9, 443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, B.A. Cold Stress as it Affects Animal Production. J. Anim. Sci. 1981, 52, 154–163. [Google Scholar] [CrossRef]
- Litherland, N.B.; Da Silva, D.N.; LaBerge, R.J.; Schefers, J.; Kertz, A. Supplemental fat for dairy calves during mild cold stress. J. Dairy Sci. 2014, 97, 2980–2989. [Google Scholar] [CrossRef] [Green Version]
- 26. A.I. Official Methods of Analysis, 18th ed.; AOAC Int.: Washington, DC, USA, 2006.
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Braundmeier-Fleming, A.G.; Skenandore, C.S.; Gil, L.; Jacobsen, V.; Cregger, M.; Badger, T.; Karr, M.; Wu, G.; Smith, S.B.; Newell-Fugate, A.E. Dietary substitution of soybean oil with coconut oil in the absence of dietary antibiotics supports growth performance and immune function in nursery and grower pigs. J. Anim. Sci. Biotechnol. 2020, 11, 27. [Google Scholar] [CrossRef]
- Hill, T.M.; Vandehaar, M.J.; Sordillo, L.M.; Catherman, D.R.; Bateman, H.G., 2nd; Schlotterbeck, R.L. Fatty acid intake alters growth and immunity in milk-fed calves. J. Dairy Sci. 2011, 94, 3936–3948. [Google Scholar] [CrossRef] [Green Version]
- Paz, H.A.; Anderson, C.L.; Muller, M.J.; Kononoff, P.J.; Fernando, S.C. Rumen Bacterial Community Composition in Holstein and Jersey Cows Is Different under Same Dietary Condition and Is Not Affected by Sampling Method. Front Microbiol. 2016, 7, 1206. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Ma, T.; Zhao, G.; Zhang, N.; Tu, Y.; Li, F.; Cui, K.; Bi, Y.; Ding, H.; Diao, Q. Effect of Age and Weaning on Growth Performance, Rumen Fermentation, and Serum Parameters in Lambs Fed Starter with Limited Ewe-Lamb Interaction. Animals 2019, 9, 825. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Yang, C.T.; Diao, Q.Y.; Tu, Y. The influence of mulberry leaf flavonoids and Candida tropicalis on antioxidant function and gastrointestinal development of preweaning calves challenged with Escherichia coli O141:K99. J. Dairy Sci. 2018, 101, 6098–6108. [Google Scholar] [CrossRef] [Green Version]
- Folch, J.; Lees, M.; Stanley, G.H.S. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Piao, M.Y.; HU, F.M.; Kong, F.L.; Liu, Y.L.; Wang, S.; Cui, K.; Sun, T.; Diao, Q.Y.; Tu, Y. Effects of dietary amylose to amylopectin ratio on growth performance, carcass quality characteristics and meat fatty acids in Chinese Qinchuan cattle. J. Integr. Agric. 2021, 20, 14. [Google Scholar] [CrossRef]
- Davis, C.L.; Drackley, J.K. The Development, Nutrition, and Management of the Young Calf; Iowa State University Press: Ames, IA, USA, 1998; p. 339. [Google Scholar]
- Jaster, E.H.; McCoy, G.C.; Spanski, N.; Tomkins, T. Effect of Extra Energy as Fat or Milk Replacer Solids in Diets of Young Dairy Calves on Growth During Cold Weather. J. Dairy Sci. 1992, 75, 2524–2531. [Google Scholar] [CrossRef]
- Bascom, S.A.; James, R.E.; McGilliard, M.L.; Van Amburgh, M. Influence of dietary fat and protein on body composition of Jersey bull calves. J. Dairy Sci. 2007, 90, 5600–5609. [Google Scholar] [CrossRef] [Green Version]
- Hill, S.R.; Knowlton, K.F.; Daniels, K.M.; James, R.E.; Pearson, R.E.; Capuco, A.V.; Akers, R.M. Effects of milk replacer composition on growth, body composition, and nutrient excretion in preweaned Holstein heifers. J. Dairy Sci. 2008, 91, 3145–3155. [Google Scholar] [CrossRef]
- Hill, T.M.; Bateman, H.G., 2nd; Aldrich, J.M.; Schlotterbeck, R.L. Effects of fat concentration of a high-protein milk replacer on calf performance. J. Dairy Sci. 2009, 92, 5147–5153. [Google Scholar] [CrossRef] [Green Version]
- Gaudreau, J.M.; Brisson, G.J. Abomasum Emptying in Dairy Calves Fed Milk Replacers with Varying Fat and Sources of Protein. J. Dairy Sci. 1980, 63, 426–440. [Google Scholar] [CrossRef]
- Jasem, M.K.; Fattahnia, F.; Mohammadi, Y.; Shokri, A.; Khalilvandi-Behroozyar, H.; Kazemi-Bonchenari, M. Effects of n-3 fatty acid supplementation from flax oil on growth performance, ruminal fermentation, and immune response in Holstein dairy calves fed either coarsely ground or steam-flaked corn grain. Anim. Feed. Sci. Technol. 2022, 290, 115372. [Google Scholar] [CrossRef]
- Garcia, M.; Shin, J.H.; Schlaefli, A.; Greco, L.F.; Maunsell, F.P.; Thatcher, W.W.; Santos, J.E.; Staples, C.R. Increasing intake of essential fatty acids from milk replacer benefits performance, immune responses, and health of preweaned Holstein calves. J. Dairy Sci. 2015, 98, 458–477. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.; Greco, L.F.; Favoreto, M.G.; Marsola, R.S.; Wang, D.; Shin, J.H.; Block, E.; Thatcher, W.W.; Santos, J.E.; Staples, C.R. Effect of supplementing essential fatty acids to pregnant nonlactating Holstein cows and their preweaned calves on calf performance, immune response, and health. J. Dairy Sci. 2014, 97, 5045–5064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klopp, R.N.; Hernandez Franco, J.F.; Hogenesch, H.; Dennis, T.S.; Cowles, K.E.; Boerman, J.P. Effect of medium-chain fatty acids on growth, health, and immune response of dairy calves. J. Dairy Sci. 2022, 105, 7738–7749. [Google Scholar] [CrossRef] [PubMed]
- Verschuren, W.M.M.; Jacobs, D.R.; Bloemberg, B.P.M.; Kromhout, D.; Menotti, A.; Aravanis, C.; Blackburn, H.; Buzina, R.; Dontas, A.S.; Fidanza, F.; et al. Serum Total Cholesterol and Long-term Coronary Heart Disease Mortality in Different Cultures: Twenty-five—Year Follow-up of the Seven Countries Study. JAMA 1995, 274, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Durand, D.; Bauchart, D.; Picherit, C.; Gruffat, D.; Graulet, B. Effects of dietary coconut oil on the density distribution and the chemical composition of plasma lipoproteins in the preruminant calf. Reprod. Nutr. Dev. 1998, 38, 205. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Zhao, D.; Yang, Y.; Zhang, L. Effect of palm oil consumption on plasma lipid concentrations related to cardiovascular disease: A systematic review and meta-analysis. Asia Pac. J. Clin. Nutr. 2019, 28, 495–506. [Google Scholar] [CrossRef]
- Ng, T.K.; Hassan, K.; Lim, J.B.; Lye, M.S.; Ishak, R. Nonhypercholesterolemic effects of a palm-oil diet in Malaysian volunteers. Am. J. Clin. Nutr. 1991, 53, 1015S–1020S. [Google Scholar] [CrossRef]
- Tholstrup, T.; Marckmann, P.; Jespersen, J.; Sandstrom, B. Fat high in stearic acid favorably affects blood lipids and factor VII coagulant activity in comparison with fats high in palmitic acid or high in myristic and lauric acids. Am. J. Clin. Nutr. 1994, 59, 371–377. [Google Scholar] [CrossRef]
- Temme, E.H.; Mensink, R.P.; Hornstra, G. Comparison of the effects of diets enriched in lauric, palmitic, or oleic acids on serum lipids and lipoproteins in healthy women and men. Am. J. Clin. Nutr. 1996, 63, 897–903. [Google Scholar] [CrossRef] [Green Version]
- Castro, J.J.; Gomez, A.; White, B.; Loften, J.R.; Drackley, J.K. Changes in the intestinal bacterial community, short-chain fatty acid profile, and intestinal development of preweaned Holstein calves. 2. Effects of gastrointestinal site and age. J. Dairy Sci. 2016, 99, 9703–9715. [Google Scholar] [CrossRef] [Green Version]
- Jolazadeh, A.R.; Mohammadabadi, T.; Dehghan-Banadaky, M.; Chaji, M.; Garcia, M. Effect of supplementation fat during the last 3 weeks of uterine life and the preweaning period on performance, ruminal fermentation, blood metabolites, passive immunity and health of the newborn calf. Br. J. Nutr. 2019, 122, 1346–1358. [Google Scholar] [CrossRef]
- Yabuuchi, Y.; Tani, M.; Matsushita, Y.; Otsuka, H.; Kobayashi, Y. Effects of lauric acid on physical, chemical and microbial characteristics in the rumen of steers on a high grain diet. Anim. Sci. J. 2007, 78, 387–394. [Google Scholar] [CrossRef]
- Jenkins, T.C. Lipid metabolism in the rumen. J. Dairy Sci. 1993, 76, 3851–3863. [Google Scholar] [CrossRef]
- Liu, S.J.; Bu, D.P.; Wang, J.Q.; Liu, L.; Liang, S.; Wei, H.Y.; Zhou, L.Y.; Li, D.; Loor, J.J. Effect of incremental levels of fish oil supplementation on specific bacterial populations in bovine ruminal fluid. J. Anim. Physiol. Anim. Nutr. 2012, 96, 9–16. [Google Scholar] [CrossRef]
- Yang, Z.T.; Liu, S.Y.; Xie, T.; Wang, Q.Q.; Wang, Z.H.; Yang, H.J.; Li, S.L.; Wang, W. Effect of Unsaturated Fatty Acid Ratio In Vitro on Rumen Fermentation, Methane Concentration, and Microbial Profile. Fermentation 2022, 8, 540. [Google Scholar] [CrossRef]
- Van Gastelen, S.; Visker, M.; Edwards, J.E.; Antunes-Fernandes, E.C.; Hettinga, K.A.; Alferink, S.J.J.; Hendriks, W.H.; Bovenhuis, H.; Smidt, H.; Dijkstra, J. Linseed oil and DGAT1 K232A polymorphism: Effects on methane emission, energy and nitrogen metabolism, lactation performance, ruminal fermentation, and rumen microbial composition of Holstein-Friesian cows. J. Dairy Sci. 2017, 100, 8939–8957. [Google Scholar] [CrossRef]
- Karimi, A.; Alijoo, Y.A.; Kazemi-Bonchenari, M.; Mirzaei, M.; Sadri, H. Effects of supplemental fat sources and forage feeding levels on growth performance, nutrient digestibility, ruminal fermentation, and nitrogen utilization in dairy calves. Animal 2021, 15, 100179. [Google Scholar] [CrossRef]
- Maia, M.R.G.; Chaudhary, L.C.; Figueres, L.; Wallace, R.J. Metabolism of polyunsaturated fatty acids and their toxicity to the microflora of the rumen. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2007, 91, 303–314. [Google Scholar] [CrossRef]
- Yanza, Y.R.; Szumacher-Strabel, M.; Jayanegara, A.; Kasenta, A.M.; Gao, M.; Huang, H.; Patra, A.K.; Warzych, E.; Cieslak, A. The effects of dietary medium-chain fatty acids on ruminal methanogenesis and fermentation in vitro and in vivo: A meta-analysis. J. Anim. Physiol. Anim. Nutr. 2021, 105, 874–889. [Google Scholar] [CrossRef] [Green Version]
- Parra, A.K. The effect of dietary fats on methane emissions, and its other effects on digestibility, rumen fermentation and lactation performance in cattle: A meta-analysis. Livest. Sci. 2013, 155, 244–254. [Google Scholar] [CrossRef]
- Klop, G.; van Laar-van Schuppen, S.; Pellikaan, W.F.; Hendriks, W.H.; Bannink, A.; Dijkstra, J. Changes in in vitro gas and methane production from rumen fluid from dairy cows during adaptation to feed additives in vivo. Animal 2017, 11, 591–599. [Google Scholar] [CrossRef] [Green Version]
- Burdick, M.; Zhou, M.; Guan, L.L.; Oba, M. Effects of medium-chain fatty acid supplementation on performance and rumen fermentation of lactating Holstein dairy cows. Animal 2022, 16, 100491. [Google Scholar] [CrossRef] [PubMed]
- Arriola, K.G.; Oliveira, A.S.; Ma, Z.X.; Lean, I.J.; Giurcanu, M.C.; Adesogan, A.T. A meta-analysis on the effect of dietary application of exogenous fibrolytic enzymes on the performance of dairy cows. J. Dairy Sci. 2017, 100, 4513–4527. [Google Scholar] [CrossRef] [PubMed]
- Prive, F.; Newbold, C.J.; Kaderbhai, N.N.; Girdwood, S.G.; Golyshina, O.V.; Golyshin, P.N.; Scollan, N.D.; Huws, S.A. Isolation and characterization of novel lipases/esterases from a bovine rumen metagenome. Appl. Microbiol. Biotechnol. 2015, 99, 5475–5485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yabuuchi, Y.; Matsushita, Y.; Otsuka, H.; Fukamachi, K.; Kobayashi, Y. Effects of supplemental lauric acid-rich oils in high-grain diet on in vitro rumen fermentation. Anim. Sci. J. 2006, 77, 300–307. [Google Scholar] [CrossRef]
- Hristov, A.N.; Grandeen, K.L.; Ropp, J.K.; McGuire, M.A. Effect of sodium laurate on ruminal fermentation and utilization of ruminal ammonia nitrogen for milk protein synthesis in dairy cows. J. Dairy Sci. 2004, 87, 1820–1831. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, H.Z.; Zhang, Y.L.; Li, X.L.; Jiang, X.R.; Ding, H.B. Effects of Dietary Supplementation with Glycerol Monolaurate (GML) or the Combination of GML and Tributyrin on Growth Performance and Rumen Microbiome of Weaned Lambs. Animals 2022, 12, 1309. [Google Scholar] [CrossRef]
- Granja-Salcedo, Y.T.; Fernandes, R.M.; de Araujo, R.C.; Kishi, L.T.; Berchielli, T.T.; de Resende, F.D.; Berndt, A.; Siqueira, G.R. Long-Term Encapsulated Nitrate Supplementation Modulates Rumen Microbial Diversity and Rumen Fermentation to Reduce Methane Emission in Grazing Steers. Front Microbiol. 2019, 10, 614. [Google Scholar] [CrossRef]
- Graulet, B.; Gruffat-Mouty, D.; Durand, D.; Bauchart, D. Effects of milk diets containing beef tallow or coconut oil on the fatty acid metabolism of liver slices from preruminant calves. Br. J. Nutr. 2007, 84, 309–318. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Items | Treatments | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| CON | CCO | PLO | T | D | T × D | ||
| BW at day 14, kg | 43.6 | 44.9 | 44.3 | 0.85 | 0.830 | - | - |
| BW at day 56, kg | 74.8 | 75.8 | 71.4 | 1.72 | 0.460 | - | - |
| ADG, kg/d | 0.74 | 0.73 | 0.66 | 0.047 | 0.326 | <0.01 | 0.177 |
| DMI 1, kg/d | 1.18 | 1.20 | 1.11 | 0.043 | 0.288 | <0.01 | 0.038 |
| MR intake, kg/d | 0.61 | 0.63 | 0.61 | 0.030 | 0.786 | <0.01 | 0.494 |
| Starter intake, kg/d | 0.57 | 0.57 | 0.52 | 0.028 | 0.085 | <0.01 | 0.623 |
| F:G ratio 2 | 1.78 | 1.79 | 1.92 | 0.146 | 0.609 | <0.01 | 0.282 |
| Fecal score 3 | 2.07 | 2.01 | 2.03 | 0.091 | 0.815 | <0.01 | 0.987 |
| Abnormal, d/week | 0.98 | 0.94 | 0.88 | 0.396 | 0.966 | <0.01 | 0.473 |
| Fatty acid intake, g/d | |||||||
| Caproic acid, C6:0 | 0.98 a | 0.38 b | 0.06 c | 0.076 | <0.01 | - | - |
| Caprylic acid, C8:0 | 0.67 b | 4.26 a | 0.00 c | 0.358 | <0.01 | - | - |
| Capric acid, C10:0 | 1.95 b | 3.76 a | 0.06 c | 0.297 | <0.01 | - | - |
| Lauric acid, C12:0 | 3.35 b | 30.71 a | 0.24 c | 2.616 | <0.01 | - | - |
| Myristic acid, C14:0 | 9.32 b | 12.53 a | 1.03 c | 0.969 | <0.01 | - | - |
| Myristolic acid, C14:1 | 0.85 a | 0.00 b | 0.00 b | 0.079 | <0.01 | ||
| Pentadecanoic acid, C15:0 | 1.04 a | 0.00 b | 0.06 b | 0.093 | <0.01 | ||
| Palmitic acid, C16:0 | 25.29 b | 7.65 c | 40.86 a | 2.720 | <0.01 | - | - |
| Palmitoleic acid, C16:1 | 1.22 a | 0.00 c | 0.12 b | 0.108 | <0.01 | ||
| Stearic acid, C18:0 | 8.11 a | 2.57 b | 4.50 c | 0.453 | <0.01 | - | - |
| Oleic acid, C18:1 | 14.20 b | 5.95 c | 25.47 a | 1.620 | <0.01 | - | - |
| Linoleic acid, C18:2 n-6 | 0.61 c | 1.44 b | 5.96 a | 0.482 | <0.01 | - | - |
| α-linolenic acid, C18:3 n-3 | 0.55 a | 0.00 c | 0.12 b | 0.046 | <0.01 | - | - |
| Arachidic acid, C20:0 | 0.12 b | 0.06 c | 0.37 a | 0.027 | <0.01 | - | - |
| Eicosadienoic acid, C20:1 | 0.06 b | 0.06 b | 0.12 a | 0.006 | <0.01 | - | - |
| Behenic acid, C22:0 | 0.12 a | 0.00 c | 0.06 b | 0.010 | <0.01 | - | - |
| Lignoceric acid, C24:0 | 0.18 a | 0.06 c | 0.12 b | 0.010 | <0.01 | - | - |
| Items 1 | Treatments | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| CON | CCO | PLO | T | D | T × D | ||
| GLU, mmol/L | 4.04 a | 4.06 a | 3.44 b | 0.178 | 0.007 | 0.001 | 0.589 |
| BHBA, mmol/L | 0.74 | 0.89 | 0.72 | 0.106 | 0.131 | 0.013 | 0.008 |
| TC, mmol/L | 1.78 b | 2.43 a | 1.62 b | 0.179 | 0.002 | 0.304 | 0.267 |
| TG, mmol/L | 0.15 ab | 0.18 a | 0.12 b | 0.018 | 0.019 | 0.006 | 0.179 |
| HDL-C, mmol/L | 0.61 b | 0.79 a | 0.55 b | 0.054 | 0.003 | 0.822 | 0.823 |
| LDL-C, mmol/L | 1.10 b | 1.55 a | 1.08 b | 0.101 | 0.007 | 0.493 | 0.529 |
| VLDL-C, mmol/L | 0.38 | 0.40 | 0.35 | 0.058 | 0.278 | 0.319 | 0.281 |
| NEFA, μmol/L | 169.3 | 171.9 | 158.8 | 16.53 | 0.278 | 0.015 | 0.013 |
| Items | Tissue | Treatment | SEM | p-Value | ||
|---|---|---|---|---|---|---|
| CON | CCO | PLO | ||||
| Total fatty acid, % | Liver | 13.98 | 14.21 | 14.13 | 0.987 | 0.406 |
| Longissimus dorsi | 1.12 | 1.09 | 1.25 | 0.075 | 0.436 | |
| Fatty acid profile, % of total fatty acid | ||||||
| Butyric acid, C4:0 | Liver | 0.22 | 0.23 | 0.25 | 0.015 | 0.186 |
| Longissimus dorsi | 0.26 | 0.23 | 0.31 | 0.016 | 0.126 | |
| Caproic acid, C6:0 | Liver | 0.37 | 0.38 | 0.42 | 0.131 | 0.218 |
| Longissimus dorsi | 0.85 | 0.77 | 0.93 | 0.037 | 0.202 | |
| Myristic acid, C14:0 | Liver | 0.82 b | 1.62 a | 0.25 c | 0.155 | <0.01 |
| Longissimus dorsi | 0.71 b | 2.37 a | 0.34 b | 0.271 | <0.01 | |
| Palmitic acid, C16:0 | Liver | 13.06 | 13.30 | 12.70 | 0.364 | 0.364 |
| Longissimus dorsi | 16.48 | 16.29 | 16.93 | 0.373 | 0.848 | |
| Palmitoleic acid, C16:1 | Liver | 0.60 a | 0.42 ab | 0.22 b | 0.057 | 0.010 |
| Longissimus dorsi | 1.19 a | 1.26 a | 0.75 b | 0.076 | 0.004 | |
| Heptadecanoic acid, C17:0 | Liver | 1.59 | 1.49 | 1.50 | 0.016 | 0.129 |
| Longissimus dorsi | 0.85 a | 0.75 ab | 0.68 b | 0.067 | 0.043 | |
| Stearic acid, C18:0 | Liver | 22.58 | 21.40 | 22.52 | 0.341 | 0.297 |
| Longissimus dorsi | 13.84 a | 12.32 b | 12.79 b | 0.222 | 0.007 | |
| Oleic acid, C18:1 | Liver | 15.27 a | 13.39 b | 14.32 ab | 0.359 | 0.021 |
| Longissimus dorsi | 24.89 | 23.32 | 22.49 | 0.509 | 0.154 | |
| Linoleic acid, C18:2 n-6 | Liver | 15.08 | 15.62 | 15.50 | 0.332 | 0.345 |
| Longissimus dorsi | 20.21 | 20.59 | 22.62 | 0.689 | 0.203 | |
| α-linolenic acid, C18:3 n-3 | Liver | 0.62 a | 0.18 b | 0.20 b | 0.070 | <0.01 |
| Longissimus dorsi | 0.53 a | 0.09 b | 0.07 b | 0.059 | <0.01 | |
| Eicosenoic acid, C20:1 | Liver | 1.04 a | 0.20 b | 0.01 b | 0.126 | <0.01 |
| Longissimus dorsi | 1.71 a | 1.31 b | 0.66 c | 0.117 | <0.01 | |
| Eicosadienoic acid, C20:2 n-6 | Liver | 0.65 | 0.64 | 0.65 | 0.054 | 0.516 |
| Longissimus dorsi | - | - | - | - | - | |
| Arachidonic acid, C20:4 n-6 | Liver | 9.76 b | 10.89 a | 11.40 a | 0.450 | <0.01 |
| Longissimus dorsi | 9.01 b | 9.95 b | 11.33 a | 0.354 | 0.003 | |
| Eicosapentaenoic acid, C20:5 n-3(EPA) | Liver | 1.35 a | 0.66 b | 0.09 c | 0.146 | <0.01 |
| Longissimus dorsi | 1.21 a | 0.65 b | 0.42 b | 0.095 | <0.01 | |
| Docosahexaenoic acid, C22:6 n-3(DHA) | Liver | 2.24 a | 1.62 b | 1.70 b | 0.119 | 0.041 |
| Longissimus dorsi | - | - | - | - | - | |
| Behenic acid, C22:0 | Liver | 13.06 | 14.25 | 14.35 | 0.618 | 0.186 |
| Longissimus dorsi | 5.70 | 6.85 | 6.12 | 0.215 | 0.080 | |
| Nervonic acid, C24:1 | Liver | 1.59 c | 2.75 b | 3.73 a | 0.247 | <0.01 |
| Longissimus dorsi | 1.72 b | 1.81 b | 2.45 a | 0.102 | <0.01 | |
| Others 1 | Liver | 0.10 | 0.96 | 0.19 | - | - |
| Longissimus dorsi | 0.84 | 1.44 | 1.11 | - | - | |
| MCFAs | Liver | 0.37 b | 0.56 a | 0.42 b | 0.029 | 0.010 |
| Longissimus dorsi | 0.90 b | 1.46 a | 0.93 b | 0.085 | 0.001 | |
| SFAs | Liver | 51.80 | 52.72 | 52.11 | 0.603 | 0.156 |
| Longissimus dorsi | 38.78 | 39.92 | 38.30 | 0.586 | 0.171 | |
| UFAs | Liver | 48.20 a | 46.37 b | 47.83 ab | 0.621 | 0.034 |
| Longissimus dorsi | 60.48 a | 58.98 b | 60.79 a | 0.435 | 0.046 | |
| MUFAs | Liver | 18.52 a | 16.76 b | 18.28 a | 0.246 | 0.049 |
| Longissimus dorsi | 29.50 a | 27.73 ab | 26.36 b | 0.528 | 0.041 | |
| PUFAs | Liver | 29.70 | 29.61 | 29.54 | 0.448 | 0.128 |
| Longissimus dorsi | 30.96 b | 31.28 b | 34.44 a | 0.679 | 0.046 | |
| n-3 PUFAs | Liver | 4.21 a | 2.46 b | 1.99 b | 0.287 | <0.01 |
| Longissimus dorsi | 1.75 a | 0.75 b | 0.49 c | 0.145 | <0.01 | |
| n-6 PUFAs | Liver | 25.49 b | 27.15 a | 27.55 a | 0.509 | <0.01 |
| Longissimus dorsi | 29.22 b | 30.54 b | 33.95 a | 0.702 | 0.022 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, F.; Piao, M.; Yang, C.; Diao, Q.; Tu, Y. Effects of Coconut Oil and Palm Oil on Growth, Rumen Microbiota, and Fatty Acid Profile of Suckling Calves. Microorganisms 2023, 11, 655. https://doi.org/10.3390/microorganisms11030655
Hu F, Piao M, Yang C, Diao Q, Tu Y. Effects of Coconut Oil and Palm Oil on Growth, Rumen Microbiota, and Fatty Acid Profile of Suckling Calves. Microorganisms. 2023; 11(3):655. https://doi.org/10.3390/microorganisms11030655
Chicago/Turabian StyleHu, Fengming, Minyu Piao, Chuntao Yang, Qiyu Diao, and Yan Tu. 2023. "Effects of Coconut Oil and Palm Oil on Growth, Rumen Microbiota, and Fatty Acid Profile of Suckling Calves" Microorganisms 11, no. 3: 655. https://doi.org/10.3390/microorganisms11030655
APA StyleHu, F., Piao, M., Yang, C., Diao, Q., & Tu, Y. (2023). Effects of Coconut Oil and Palm Oil on Growth, Rumen Microbiota, and Fatty Acid Profile of Suckling Calves. Microorganisms, 11(3), 655. https://doi.org/10.3390/microorganisms11030655