
Oral Administration of Lacticaseibacillus rhamnosus CRL1505 Modulates Lung Innate Immune Response against Klebsiella pneumoniae ST25

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Animals
2.3. Respiratory Infection with K. pneumoniae LABACER 01 and LABACER 27
2.4. K. pneumoniae Counts in Lungs and Blood
2.5. Lung Damage
2.6. Leukocytes Counts
2.7. Serum Cytokines and Bronchoalveolar Lavages
2.8. Statistical Analysis
3. Results
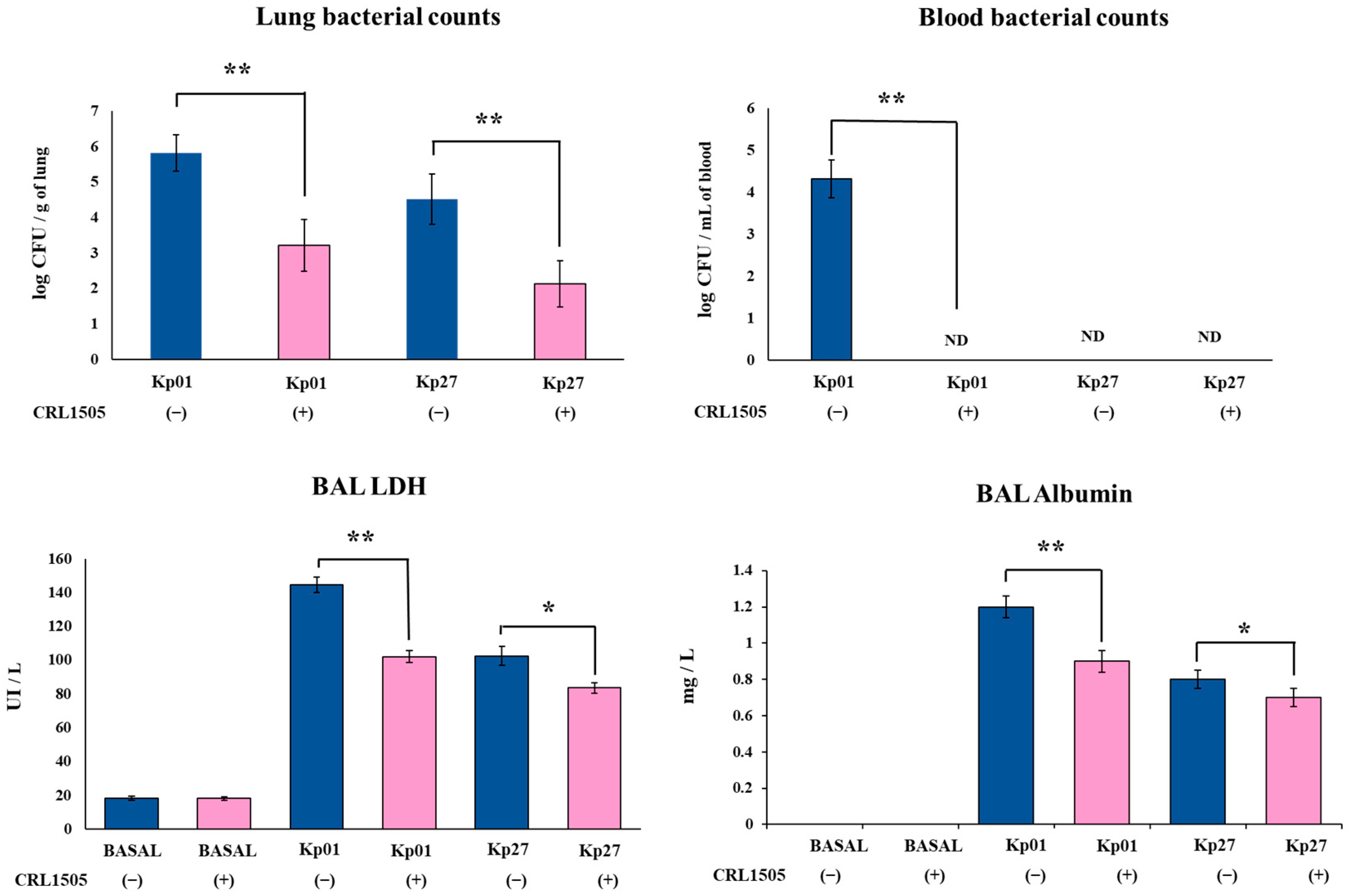
3.1. Lcb. rhamnosus CRL1505 Improves Resistance to K. pneumoniae ST25 Infection
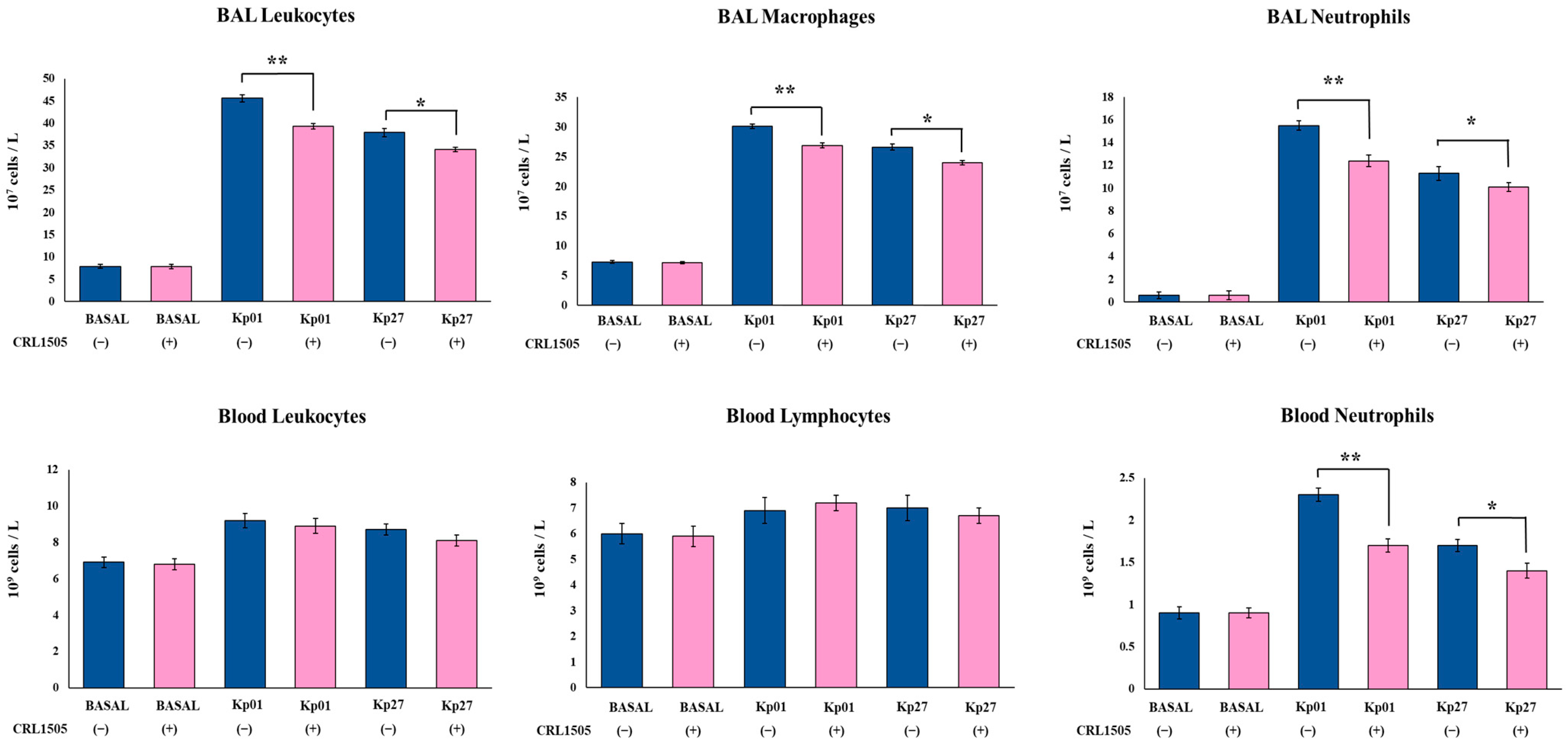
3.2. Lcb. rhamnosus CRL1505 Modulates Innate Immunity against K. pneumoniae ST25 Infection
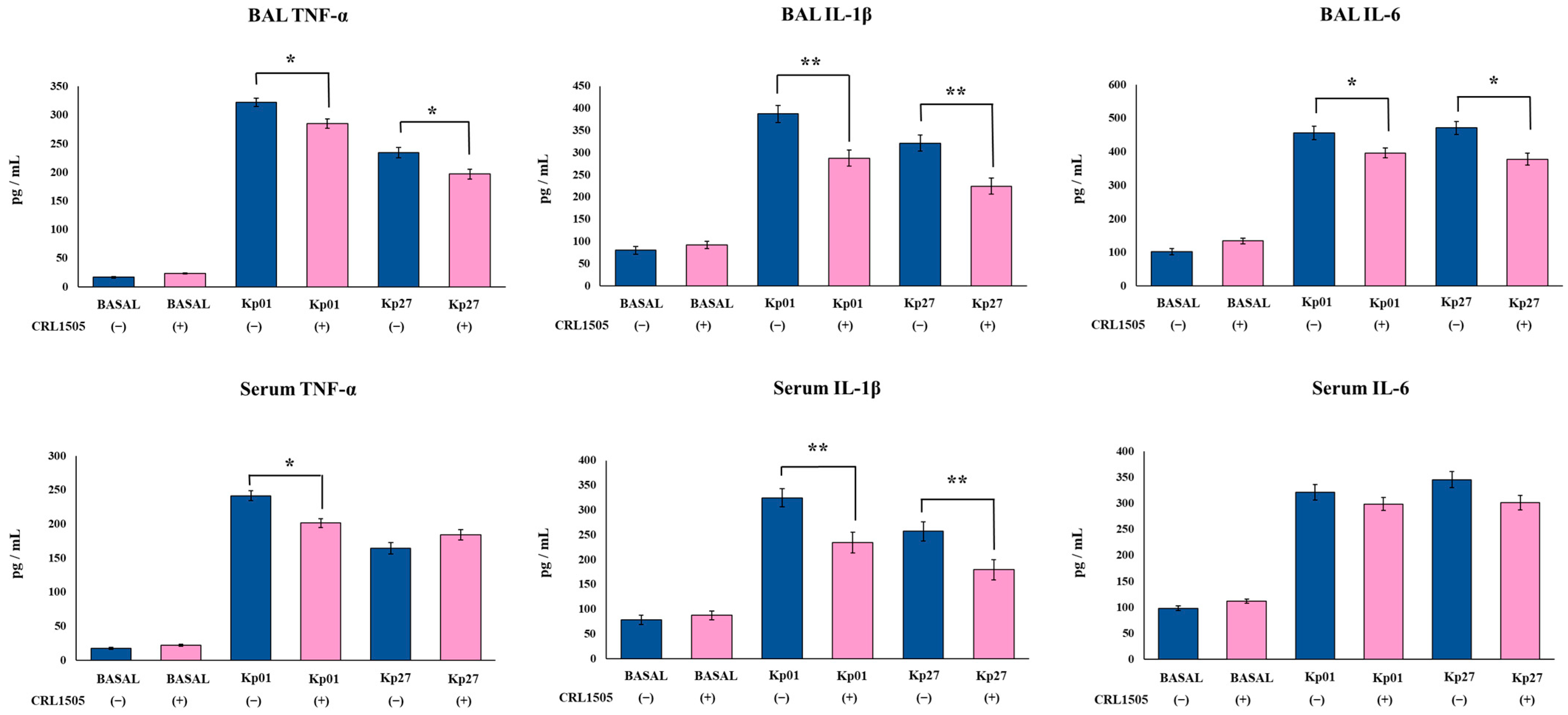
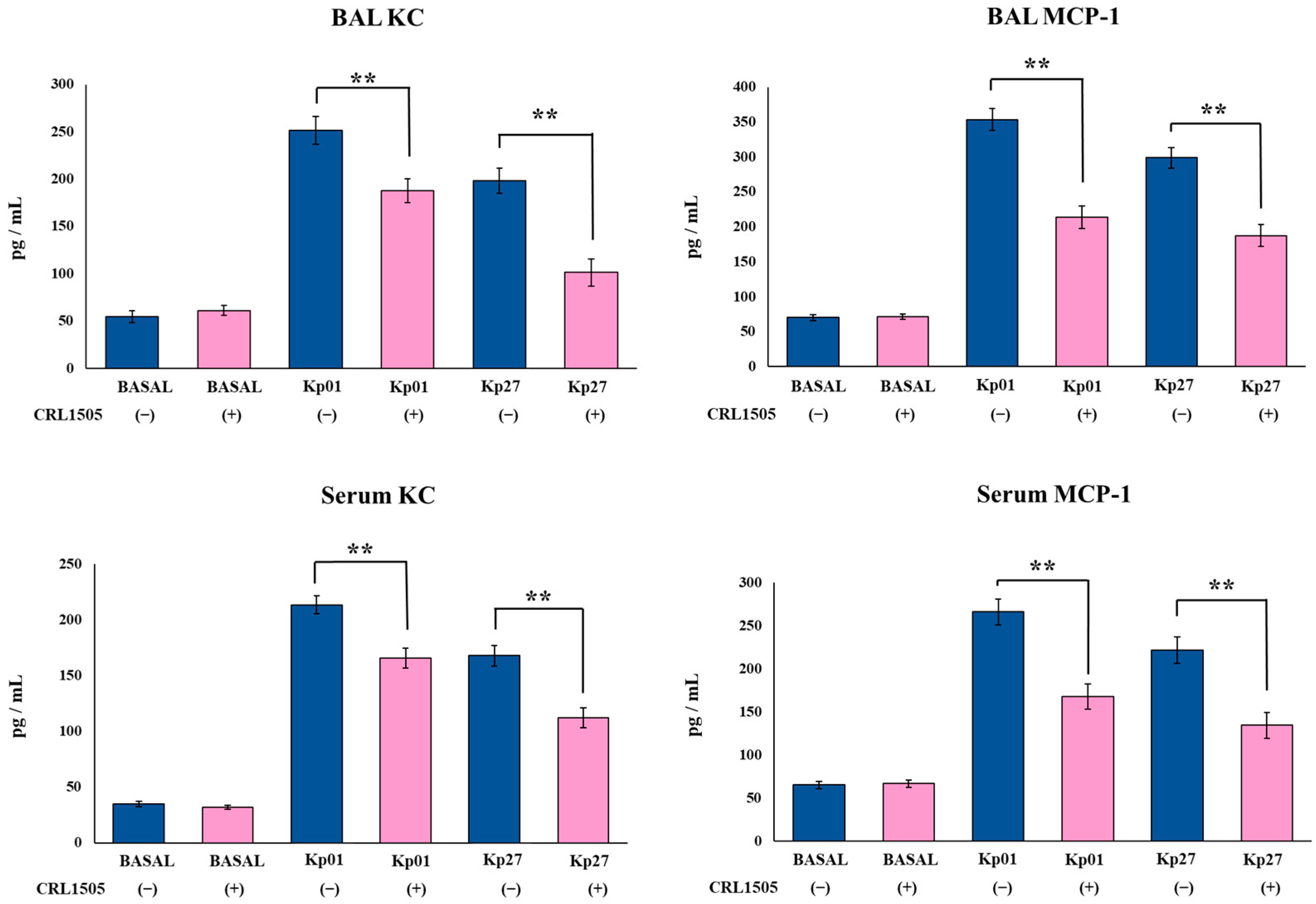
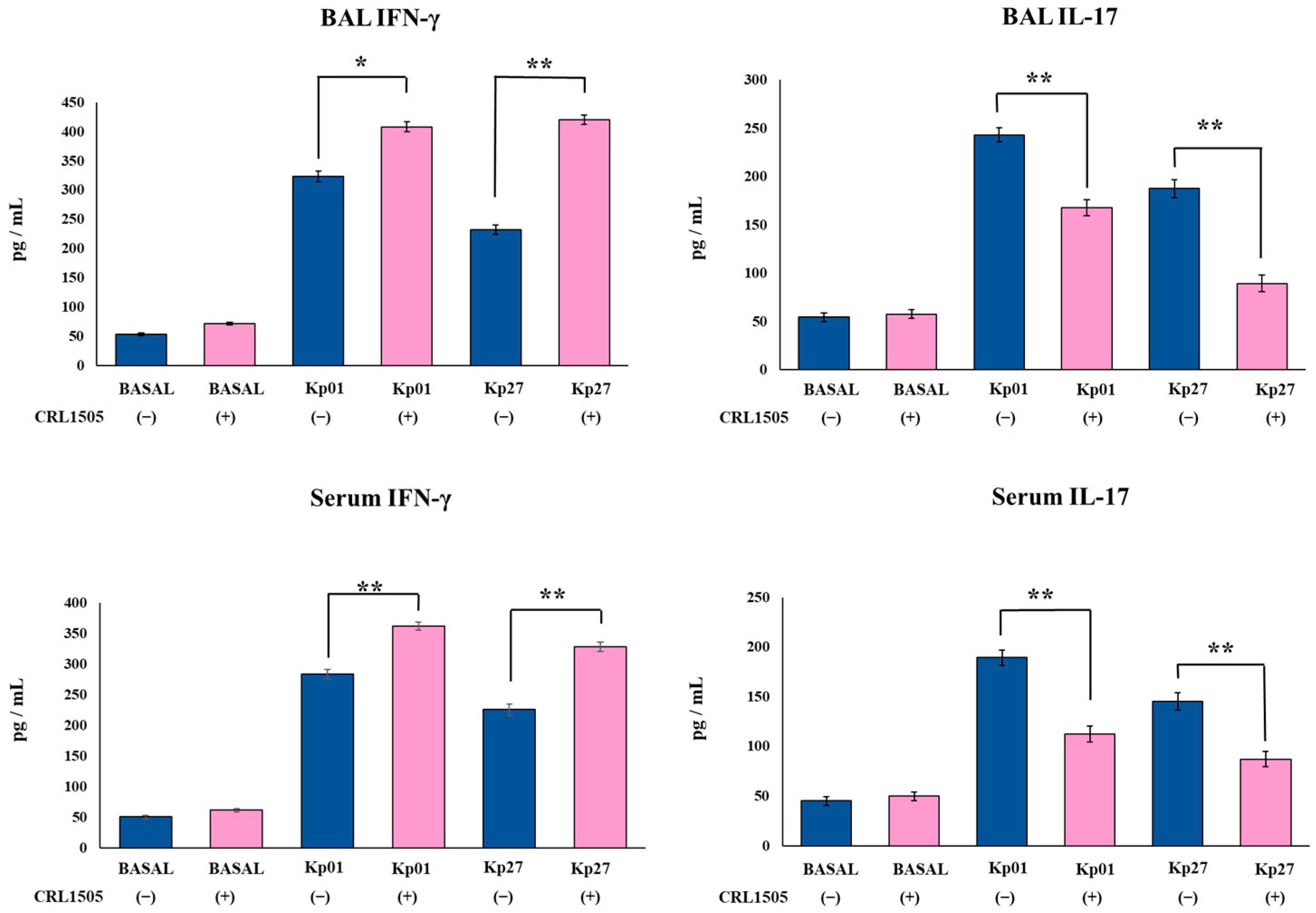
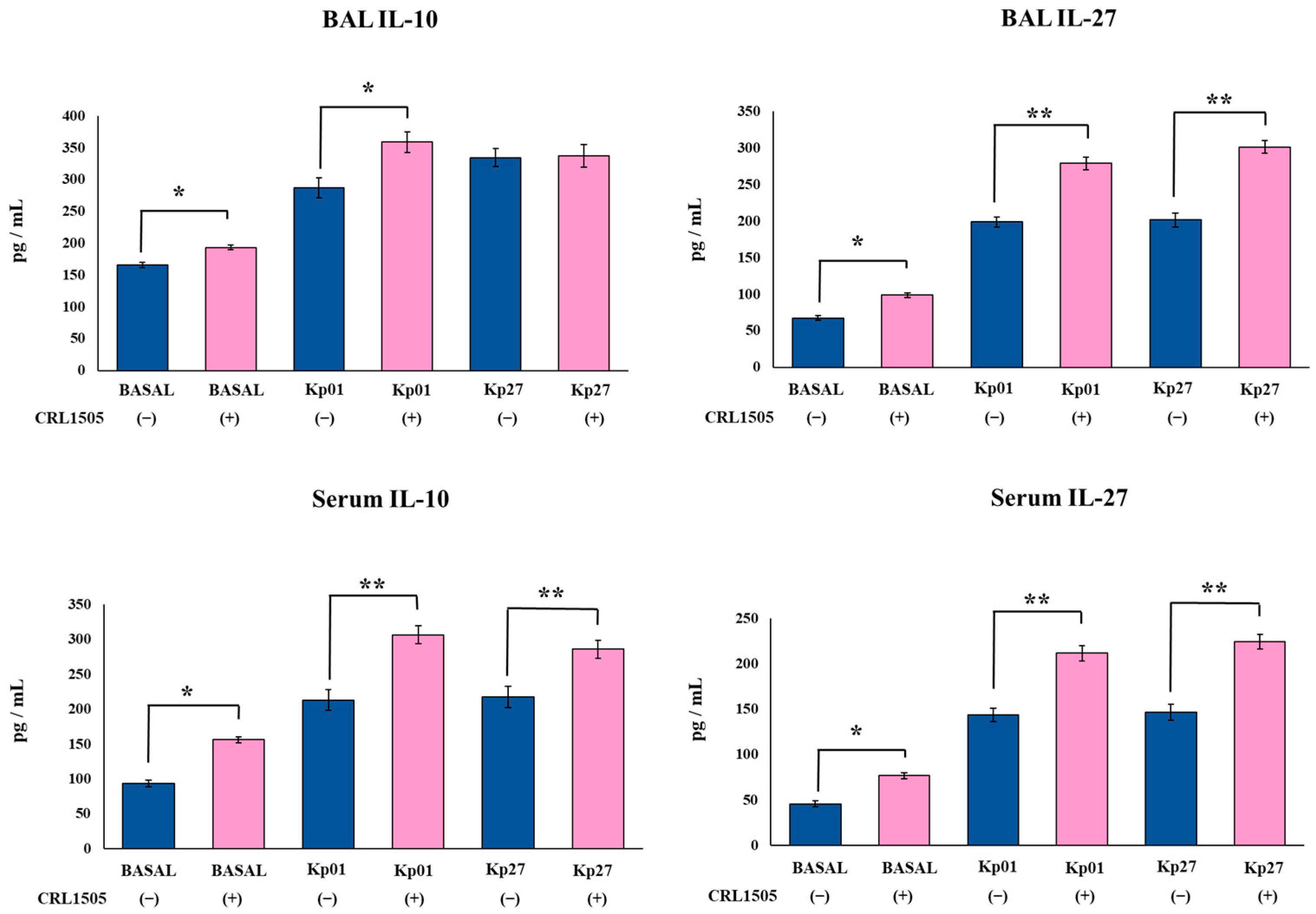
3.3. Lcb. rhamnosus CRL1505 Modulates Effector and Regulatory Cytokines Produced against K. pneumoniae ST25 Infection
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gomez, S.A.; Pasteran, F.G.; Faccone, D.; Tijet, N.; Rapoport, M.; Lucero, C.; Lastovetska, O.; Albornoz, E.; Galas, M.; Melano, R.G.; et al. Clonal dissemination of Klebsiella pneumoniae ST258 harbouring KPC-2 in Argentina. Clin. Microbiol. Infect. 2011, 17, 1520–1524. [Google Scholar] [CrossRef] [PubMed]
- Cejas, D.; Fernandez Canigia, L.; Nastro, M.; Rodríguez, C.; Tanco, A.; Rodríguez, H.; Vay, C.; Maldonado, I.; Famiglietti, A.; Giovanakis, M.; et al. Hyperendemic clone of KPC producing Klebsiella pneumoniae ST 258 in Buenos Aires hospitals. Infect. Genet. Evol. 2012, 12, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Cejas, D.; Elena, A.; Guevara Nuñez, D.; Sevilla Platero, P.; De Paulis, A.; Magariños, F.; Alfonso, C.; Berger, M.A.; Fernández-Canigia, L.; Gutkind, G.; et al. Changing epidemiology of KPC-producing Klebsiella pneumoniae in Argentina: Emergence of hypermucoviscous ST25 and high-risk clone ST307. J. Glob. Antimicrob. Resist. 2019, 18, 238–242. [Google Scholar] [CrossRef]
- Lin, J.; Huang, Y.; Qian, L.; Pan, X.; Song, Y. Liver Abscess Combined with Endogenous Endophthalmitis Caused by Genotype ST25 Serotype K2 Hypervirulent Klebsiella pneumoniae: A Case Report. Infect. Drug Resist. 2022, 15, 4557–4561. [Google Scholar] [CrossRef] [PubMed]
- Vargas, J.M.; Moreno Mochi, M.P.; Nuñez, J.M.; Cáceres, M.; Mochi, S.; del Campo Moreno, R.; Jure, M.A. Virulence factors and clinical patterns of multiple-clone hypermucoviscous KPC-2 producing K. pneumoniae. Heliyon 2019, 5, e01829. [Google Scholar] [CrossRef] [PubMed]
- Jure, M.A.; Castillo, M.E.; Musa, H.E.; López, C.G.; Cáceres, M.A.; Mochi, S.D.; Bousquet, A.A.; Genel, N.A.; Arlet, G.A.; Decré, D.C. Novel patterns in the molecular epidemiology of KPC-producing Klebsiella pneumoniae in Tucumán, Argentina. J. Glob. Antimicrob. Resist. 2019, 19, 183–187. [Google Scholar] [CrossRef]
- Jure, M.A.; Albarracin, L.; Vargas, J.M.; Maidana, S.D.; Zamar, J.C.; Kitazawa, H.; Villena, J. Draft genome sequences of two hypermucoviscous carbapenem-resistant ST25 Klebsiella pneumoniae strains causing respiratory and systemic infections. J. Glob. Antimicrob. Resist. 2021, 26, 174–176. [Google Scholar] [CrossRef]
- Villena, J.; Kitazawa, H. The Modulation of Mucosal Antiviral Immunity by Immunobiotics: Could They Offer Any Benefit in the SARS-CoV-2 Pandemic? Front. Physiol. 2020, 11, 699. [Google Scholar] [CrossRef]
- Raras, T.Y.M.; Firdausy, A.F.; Kinanti, I.R.; Noorhamdani, N. Anti-Biofilm Activity of Lactic Acid Bacteria Isolated from Kefir against Multidrug-Resistant Klebsiella pneumoniae. J. Pure Appl. Microbiol. 2019, 13, 983–992. [Google Scholar] [CrossRef]
- El-Mokhtar, M.A.; Hassanein, K.M.; Ahmed, A.S.; Gad, G.F.M.; Amin, M.M.; Hassanein, O.F.E. Antagonistic Activities of Cell-Free Supernatants of Lactobacilli Against Extended-Spectrum β-Lactamase Producing Klebsiella pneumoniae and Pseudomonas aeruginosa. Infect. Drug Resist. 2020, 13, 543–552. [Google Scholar] [CrossRef]
- Yan, R.; Lu, Y.; Wu, X.; Yu, P.; Lan, P.; Wu, X.; Jiang, Y.; Li, Q.; Pi, X.; Liu, W.; et al. Anticolonization of Carbapenem-Resistant Klebsiella pneumoniae by Lactobacillus plantarum LP1812 through Accumulated Acetic Acid in Mice Intestinal. Front. Cell. Infect. Microbiol. 2021, 11, 1276. [Google Scholar] [CrossRef] [PubMed]
- Lagrafeuille, R.; Miquel, S.; Balestrino, D.; Vareille-Delarbre, M.; Chain, F.; Langella, P.; Forestier, C. Opposing effect of Lactobacillus on in vitro Klebsiella pneumoniae in biofilm and in an in vivo intestinal colonisation model. Benef. Microbes 2017, 9, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Ermolenko, E.; Rybalchenko, O.; Borshev, Y.; Tarasova, E.; Kramskaya, T.; Leontieva, G.; Kotyleva, M.; Orlova, O.; Abdurasulova, I.; Suvorov, A. Influence of monostrain and multistrain probiotics on immunity, intestinal ultrastructure and microbiota in experimental dysbiosis. Benef. Microbes 2018, 9, 937–949. [Google Scholar] [CrossRef] [PubMed]
- Savinova, O.S.; Glazunova, O.A.; Moiseenko, K.V.; Begunova, A.V.; Rozhkova, I.V.; Fedorova, T.V. Exoproteome analysis of antagonistic interactions between the probiotic bacteria Limosilactobacillus reuteri LR1 and Lacticaseibacillus rhamnosus F and multidrug resistant strain of klebsiella pneumonia. Int. J. Mol. Sci. 2021, 22, 10999. [Google Scholar] [CrossRef]
- Vareille-Delarbre, M.; Miquel, S.; Garcin, S.; Bertran, T.; Balestrino, D.; Evrard, B.; Forestier, C. Immunomodulatory effects of Lactobacillus plantarum on inflammatory response induced by Klebsiella pneumoniae. Infect. Immun. 2019, 87, e00570-19. [Google Scholar] [CrossRef]
- Vieira, A.T.; Rocha, V.M.; Tavares, L.; Garcia, C.C.; Teixeira, M.M.; Oliveira, S.C.; Cassali, G.D.; Gamba, C.; Martins, F.S.; Nicoli, J.R. Control of Klebsiella pneumoniae pulmonary infection and immunomodulation by oral treatment with the commensal probiotic Bifidobacterium longum 51A. Microbes Infect. 2016, 18, 180–189. [Google Scholar] [CrossRef]
- Raya Tonetti, F.; Clua, P.; Fukuyama, K.; Marcial, G.; Sacur, J.; Marranzino, G.; Tomokiyo, M.; Vizoso-Pinto, G.; Garcia-Cancino, A.; Kurata, S.; et al. The ability of postimmunobiotics from L. rhamnosus CRL1505 to Protect against respiratory syncytial virus and pneumococcal super-infection is a strain-dependent characteristic. Microorganisms 2022, 10, 2185. [Google Scholar] [CrossRef]
- Kitazawa, H.; Villena, J. Modulation of respiratory TLR3-anti-viral response by probiotic microorganisms: Lessons learned from Lactobacillus rhamnosus CRL1505. Front. Immunol. 2014, 5, 201. [Google Scholar] [CrossRef]
- Garcia-Castillo, V.; Tomokiyo, M.; Raya Tonetti, F.; Islam, M.A.; Takahashi, H.; Kitazawa, H.; Villena, J. Alveolar Macrophages Are Key Players in the Modulation of the Respiratory Antiviral Immunity Induced by Orally Administered Lacticaseibacillus rhamnosus CRL1505. Front. Immunol. 2020, 11, 568636. [Google Scholar] [CrossRef]
- Albarracin, L.; Garcia-Castillo, V.; Masumizu, Y.; Indo, Y.; Islam, M.A.; Suda, Y.; Garcia-Cancino, A.; Aso, H.; Takahashi, H.; Kitazawa, H.; et al. Efficient Selection of New Immunobiotic Strains With Antiviral Effects in Local and Distal Mucosal Sites by Using Porcine Intestinal Epitheliocytes. Front. Immunol. 2020, 11, 543. [Google Scholar] [CrossRef]
- Chiba, E.; Tomosada, Y.; Vizoso-Pinto, M.G.; Salva, S.; Takahashi, T.; Tsukida, K.; Kitazawa, H.; Alvarez, S.; Villena, J. Immunobiotic Lactobacillus rhamnosus improves resistance of infant mice against respiratory syncytial virus infection. Int. Immunopharmacol. 2013, 17, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Salva, S.; Villena, J.; Alvarez, S. Immunomodulatory activity of Lactobacillus rhamnosus strains isolated from goat milk: Impact on intestinal and respiratory infections. Int. J. Food Microbiol. 2010, 141, 82–89. [Google Scholar] [CrossRef]
- Dentice Maidana, S.; Ortiz Moyano, R.; Vargas, J.M.; Fukuyama, K.; Kurata, S.; Melnikov, V.; Jure, M.Á.; Kitazawa, H.; Villena, J. Respiratory Commensal Bacteria Increase Protection against Hypermucoviscous Carbapenem-Resistant Klebsiella pneumoniae ST25 Infection. Pathogens 2022, 11, 1063. [Google Scholar] [CrossRef] [PubMed]
- Albarracin, L.; Ortiz Moyano, R.; Vargas, J.M.; Andrade, B.G.N.; Zamar, J.C.; Dentice Maidana, S.; Fukuyama, K.; Kurata, S.; Jure, M.Á.; Kitazawa, H.; et al. Genomic and Immunological Characterization of Hypermucoviscous Carbapenem-Resistant Klebsiella pneumoniae ST25 Isolates from Northwest Argentina. Int. J. Mol. Sci. 2022, 23, 7361. [Google Scholar] [CrossRef] [PubMed]
- Lou, W.; Venkataraman, S.; Zhong, G.; Ding, B.; Tan, J.P.K.; Xu, L.; Fan, W.; Yang, Y.Y. Antimicrobial polymers as therapeutics for treatment of multidrug-resistant Klebsiella pneumoniae lung infection. Acta Biomater. 2018, 78, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Ge, Y.; Wu, T.; Zhao, K.; Chen, Y.; Wu, B.; Zhu, F.; Zhu, B.; Cui, L. Co-infection with respiratory pathogens among COVID-2019 cases. Virus Res. 2020, 285, 198005. [Google Scholar] [CrossRef] [PubMed]
- Russo, T.A.; Marr, C.M. Hypervirulent Klebsiella pneumoniae. Clin. Microbiol. Rev. 2019, 32, e00001-19. [Google Scholar] [CrossRef]
- CDCP. Antibiotic Resistance Threats in the United States, 2019; CDC: Atlanta, GA, USA, 2019. [CrossRef]
- Wasbotten, R.K.; Dahler, A.A.; Mackel, J.J.; Smith, C.M.; Rosen, D.A. Murine Respiratory Tract Infection with Classical Klebsiella pneumoniae Induces Bronchus-Associated Lymphoid Tissue. Infect. Immun. 2022, 90, e00596-21. [Google Scholar] [CrossRef]
- Ahn, D.; Bhushan, G.; McConville, T.H.; Annavajhala, M.K.; Soni, R.K.; Wong Fok Lung, T.; Hofstaedter, C.E.; Shah, S.S.; Chong, A.M.; Castano, V.G.; et al. An acquired acyltransferase promotes Klebsiella pneumoniae ST258 respiratory infection. Cell Rep. 2021, 35, 109196. [Google Scholar] [CrossRef]
- Happel, K.I.; Dubin, P.J.; Zheng, M.; Ghilardi, N.; Lockhart, C.; Quinton, L.J.; Odden, A.R.; Shellito, J.E.; Bagby, G.J.; Nelson, S.; et al. Divergent roles of IL-23 and IL-12 in host defense against Klebsiella pneumoniae. J. Exp. Med. 2005, 202, 761–769. [Google Scholar] [CrossRef]
- Moore, T.A.; Lau, H.Y.; Cogen, A.L.; Standiford, T.J. Defective innate antibacterial host responses during murine Klebsiella pneumoniae bacteremia: Tumor necrosis factor (TNF) receptor 1 deficiency versus therapy with anti-TNF-alpha. Clin. Infect. Dis. 2005, 41 (Suppl. 3), S213–S217. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Zhang, X.; Yang, R.; Jing, H.; Yuan, Y.; Chen, Z.; Gou, Q.; Zhao, Z.; Zhang, J.; Wang, X. Host Immune Response to Clinical Hypervirulent Klebsiella pneumoniae Pulmonary Infections via Transcriptome Analysis. J. Immunol. Res. 2022, 2022, 5336931. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Guo, J.; Cao, C.; Qin, T.; Zhao, Y.; Song, X.; Lv, M.; Hu, L.; Zhang, L.; Zhou, D.; et al. Time-Course Transcriptome Analysis of Lungs From Mice Infected With Hypervirulent Klebsiella pneumoniae via Aerosolized Intratracheal Inoculation. Front. Cell. Infect. Microbiol. 2022, 12, 386. [Google Scholar] [CrossRef] [PubMed]
- Fagundes, C.T.; Amaral, F.A.; Vieira, A.T.; Soares, A.C.; Pinho, V.; Nicoli, J.R.; Vieira, L.Q.; Teixeira, M.M.; Souza, D.G. Transient TLR activation restores inflammatory response and ability to control pulmonary bacterial infection in germfree mice. J. Immunol. 2012, 188, 1411–1420. [Google Scholar] [CrossRef]
- Schuijt, T.J.; Lankelma, J.M.; Scicluna, B.P.; De Sousa E Melo, F.; Roelofs, J.J.T.H.; De Boer, J.D.; Hoogendijk, A.J.; De Beer, R.; De Vos, A.; Belzer, C.; et al. The gut microbiota plays a protective role in the host defence against pneumococcal pneumonia. Gut 2016, 65, 575–583. [Google Scholar] [CrossRef]
- Ichinohe, T.; Pang, I.K.; Kumamoto, Y.; Peaper, D.R.; Ho, J.H.; Murray, T.S.; Iwasaki, A. Microbiota regulates immune defense against respiratory tract influenza a virus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 5354–5359. [Google Scholar] [CrossRef]
- Abt, M.C.; Osborne, L.C.; Monticelli, L.A.; Doering, T.A.; Alenghat, T.; Sonnenberg, G.F.; Paley, M.A.; Antenus, M.; Williams, K.L.; Erikson, J.; et al. Commensal bacteria calibrate the activation threshold of innate antiviral immunity. Immunity 2012, 37, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Peñaloza, H.F.; Noguera, L.P.; Ahn, D.; Vallejos, O.P.; Castellanos, R.M.; Vazquez, Y.; Salazar-Echegarai, F.J.; González, L.; Suazo, I.; Pardo-Roa, C.; et al. Interleukin-10 produced by Myeloid-Derived suppressor cells provides protection to Carbapenem-Resistant Klebsiella pneumoniae sequence type 258 by enhancing its clearance in the airways. Infect. Immun. 2019, 87, e00665-18. [Google Scholar] [CrossRef]
- Branchett, W.J.; Lloyd, C.M. Regulatory cytokine function in the respiratory tract. Mucosal Immunol. 2019, 12, 589–600. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dentice Maidana, S.; Imamura, Y.; Elean, M.; Albarracín, L.; Nishiyama, K.; Suda, Y.; Kurata, S.; Jure, M.Á.; Kitazawa, H.; Villena, J. Oral Administration of Lacticaseibacillus rhamnosus CRL1505 Modulates Lung Innate Immune Response against Klebsiella pneumoniae ST25. Microorganisms 2023, 11, 1148. https://doi.org/10.3390/microorganisms11051148
Dentice Maidana S, Imamura Y, Elean M, Albarracín L, Nishiyama K, Suda Y, Kurata S, Jure MÁ, Kitazawa H, Villena J. Oral Administration of Lacticaseibacillus rhamnosus CRL1505 Modulates Lung Innate Immune Response against Klebsiella pneumoniae ST25. Microorganisms. 2023; 11(5):1148. https://doi.org/10.3390/microorganisms11051148
Chicago/Turabian StyleDentice Maidana, Stefania, Yoshiya Imamura, Mariano Elean, Leonardo Albarracín, Keita Nishiyama, Yoshihito Suda, Shoichiro Kurata, María Ángela Jure, Haruki Kitazawa, and Julio Villena. 2023. "Oral Administration of Lacticaseibacillus rhamnosus CRL1505 Modulates Lung Innate Immune Response against Klebsiella pneumoniae ST25" Microorganisms 11, no. 5: 1148. https://doi.org/10.3390/microorganisms11051148
APA StyleDentice Maidana, S., Imamura, Y., Elean, M., Albarracín, L., Nishiyama, K., Suda, Y., Kurata, S., Jure, M. Á., Kitazawa, H., & Villena, J. (2023). Oral Administration of Lacticaseibacillus rhamnosus CRL1505 Modulates Lung Innate Immune Response against Klebsiella pneumoniae ST25. Microorganisms, 11(5), 1148. https://doi.org/10.3390/microorganisms11051148