
Interactions of Different Species of Phytophthora with Cacao Induce Genetic, Biochemical, and Morphological Plant Alterations




Abstract
:1. Introduction
2. Materials and Methods
2.1. String Construction and Selection for Database Searches
2.2. Literature Search
2.3. Literature Selection
2.4. Data Extraction
3. Results
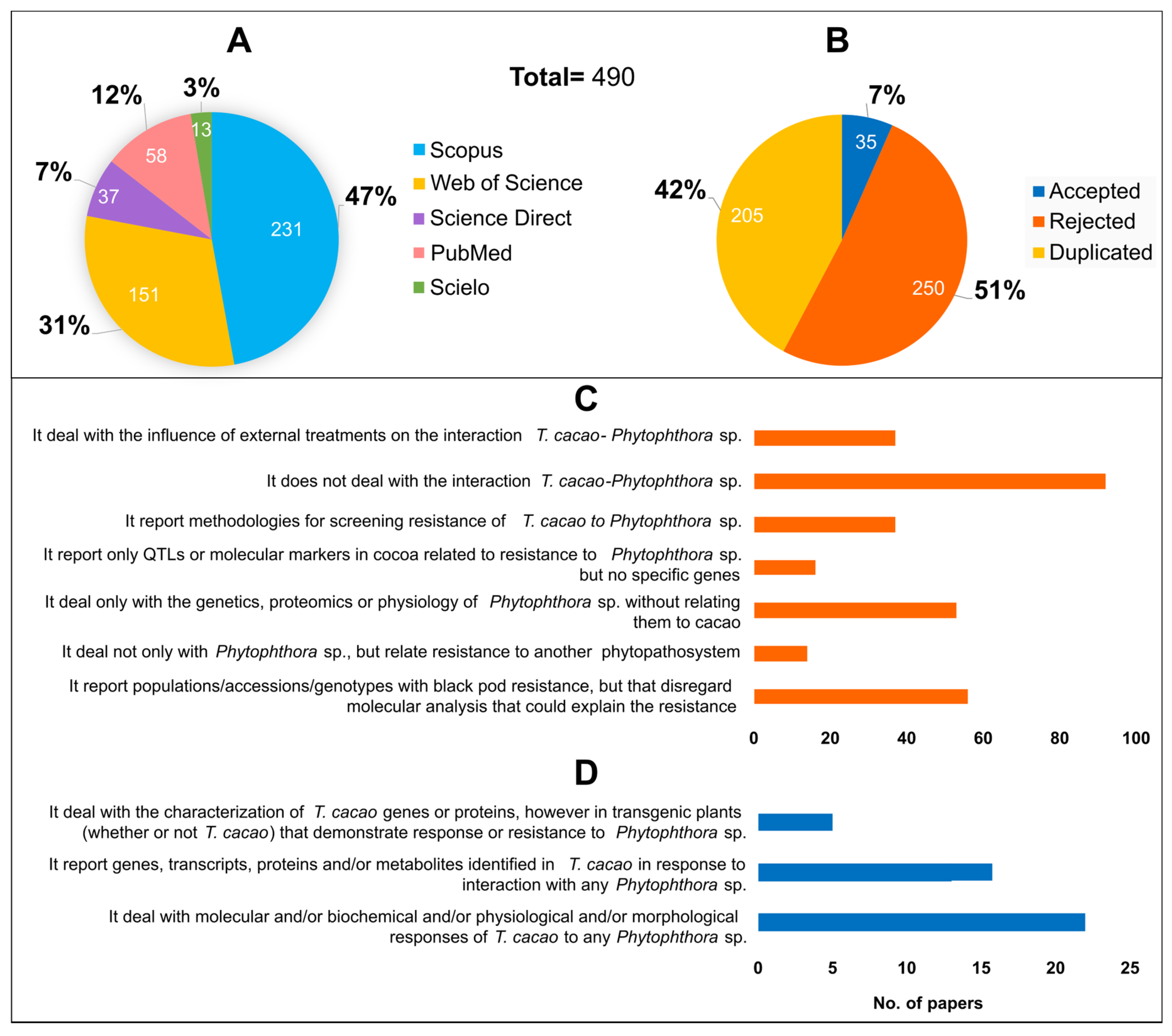
3.1. Subsection Literature Search and Selection
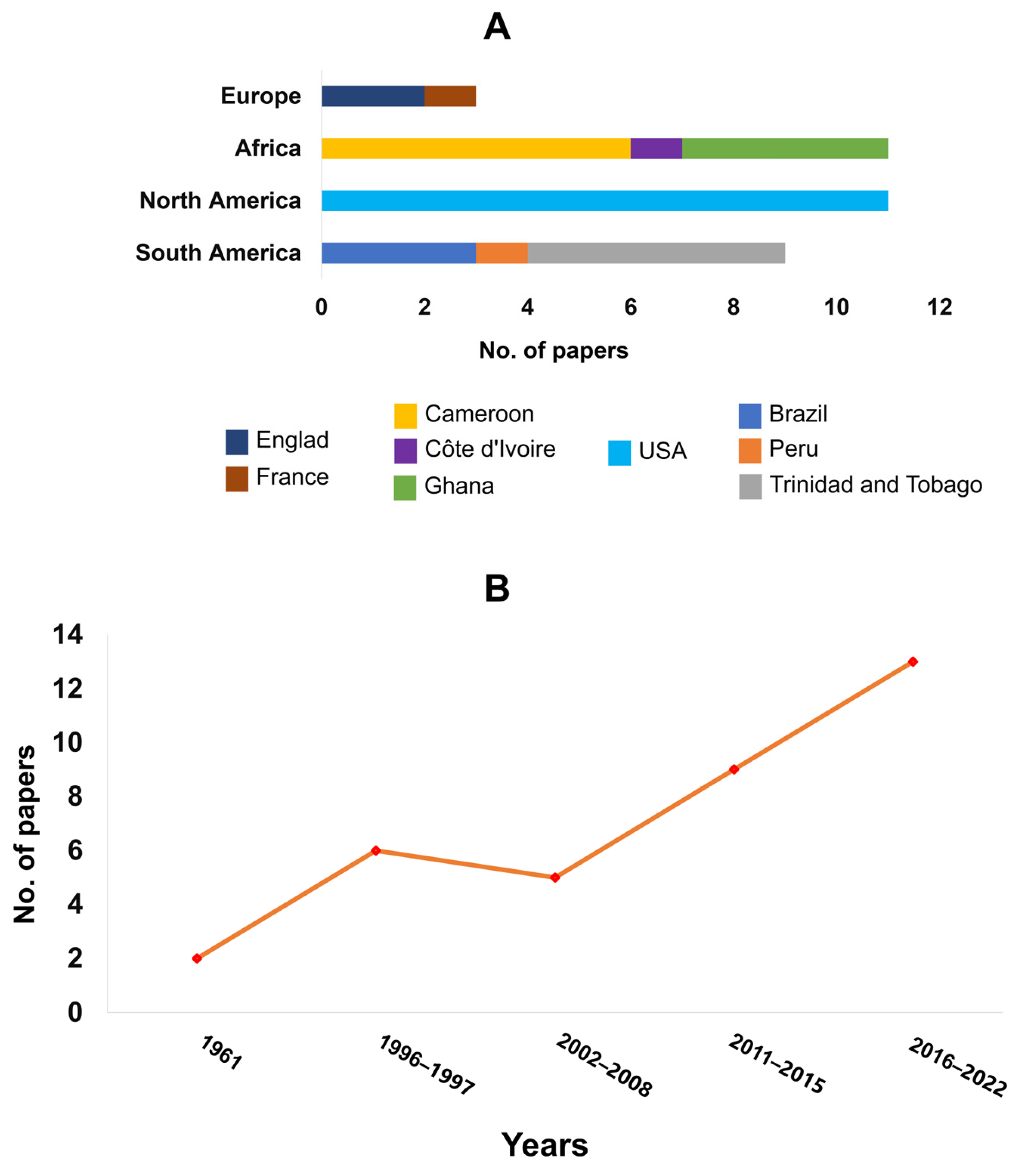
3.2. Countries of Origin and Year of Publication of Papers Accepted for Data Extraction
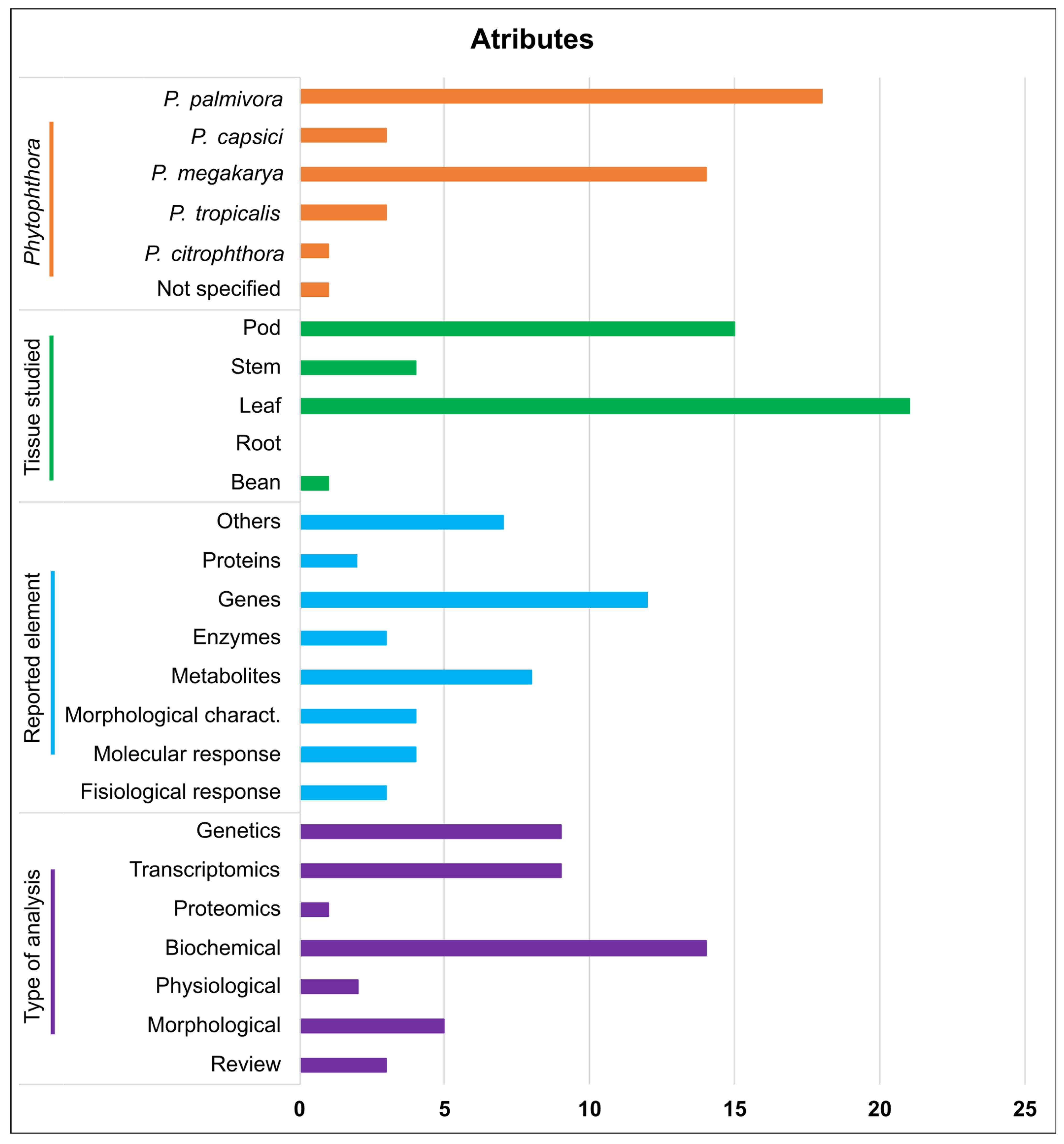
3.3. Data Extraction
3.4. Elements That Participate in the Interaction with Species of Phytophthora

{kind=link}
{kind=link}
{kind=link}
| Enzymes | |||||||
| Enzyme activity | Level of activity | Time after inoculation | Effect on T. cacao | Methodology | Tissue studied | Species | References |
| Polyphenol oxidase | High | 6 h | Resistance | Tissue maceration and dimming | Fruit/Beans | Ppal | [27] |
| Peroxidase | High | 12 weeks | Resistance | Spectrophotometry | Stem | Ppal | [28] |
| Polyphenoloxidase | High | 12 weeks | Resistance | Spectrophotometry | Stem | Ppal | [28] |
| Phenylalanine ammonium-lyase | High | 12 weeks | Resistance | Spectrophotometry | Stem | Ppal | [28] |
| Polyphenoloxidase (isoforms) | High | 6 days | Resistance | Spectrophotometry | Fruit | Pmeg | [29] |
| Metabolites | |||||||
| Metabolite | Content | Time after Infection | Effect on T. cacao | Methodology | Tissue studied | Species | References |
| Tannins | High | 6 h | Resistance | Tissue maceration and dimming | Fruit | Ppal | [27] |
| Flavonol | High | 4 days | Resistance | Spectrophotometry | Fruit | Pmeg | [30] |
| Hydroxynamic derivatives | High | 4 days | Resistance | Spectrophotometry | Fruit | Pmeg | [30] |
| Phenol | Presence | 6 weeks | Resistance | Ferric chloride test | Stem | Ppal | [31] |
| Terpenoides | Absence | 6 weeks | Resistance | Test of 2,4-DNP | Stem | Ppal | [31] |
| Glicosides | Absence | 6 weeks | Resistance | Picric acid test | Stem | Ppal | [31] |
| Soluble phenolics compounds | High | 6 days | Resistance | Spectrophotometry | Leaf | Pmeg | [32] |
| Luteolin derivatives | High | 6 days | Resistance | HPLC | Leaf | Pmeg | [32] |
| Apigenin derivaties | High | 6 days | Resistance | HPLC | Leaf | Pmeg | [32] |
| Derivatives of hydroxycincamic acids | High | 6 days | Resistance | HPLC | Leaf | Pmeg | [32] |
| Soluble phenolic compounds | High | 6 days | Resistance | Spectrophotometry | Leaf | Pmeg | [33] |
| Luteolin derivatives | High | 6 days | Resistance | HPLC | Leaf | Pmeg | [33] |
| Apigenin derivatives | High | 6 days | Resistance | HPLC | Leaf | Pmeg | [33] |
| Derivatives of hydroxycinnamic acids | High | 6 days | Resistance | HPLC | Leaf | Pmeg | [33] |
| Soluble phenolic compounds | High | 6 days | Resistance | Spectrophotometry | Leaf | Pmeg | [34] |
| Total polyphenols | High | 6 days | Resistance | Folin-Ciocalteu reagent method | Leaf/Fruit | Pmeg | [7] |
| Flavonoides | High | 6 days | Resistance | Aluminum chloride method | Leaf/Fruit | Pmeg | [7] |
| Tannins | High | 6 days | Resistance | Vanillin method | Leaf/Fruit | Pmeg | [7] |
| Caffeoyl-DOPA (clovamide) | High | Basal | Resistance | LC-MS/MS | Leaf | Ppal | [22] |
| Coumaroyl-DOPA | High | Basal | Resistance | LC-MS/MS | Leaf | Ppal | [22] |
| Coumaroyl-Tyrosine | High | Basal | Resistance | LC-MS/MS | Leaf | Ppal | [22] |
| Sinapoyl-Tyrosine | High | Basal | Resistance | LC-MS/MS | Leaf | Ppal | [22] |
| Caffeoyl-Tryptophan | High | Basal | Resistance | LC-MS/MS | Leaf | Ppal | [22] |
| Caffeoyl-DOPA, Alkyl-Sulfated | High | Basal | Resistance | LC-MS/MS | Leaf | Ppal | [22] |
| Feruloyl-DOPA | High | Basal | Resistance | LC-MS/MS | Leaf | Ppal | [22] |
| Feruloyl-DOPA, Aryl-Sulfated | High | Basal | Resistance | LC-MS/MS | Leaf | Ppal | [22] |
| Caffeoyl-Phenylalanine | High | Basal | Resistance | LC-MS/MS | Leaf | Ppal | [22] |
| Caffeoyl-Tyrosine, Alkyl-Sulfated | High | Basal | Resistance | LC-MS/MS | Leaf | Ppal | [22] |
| Clovamide | Decrease | 72 h | Resistance | LC-MS/MS | Fruit | Ppal | [22] |
| Aryl-sulfatedclovamide | Decrease | 72 h | Resistance | LC-MS/MS | Fruit | Ppal | [22] |
| Feruloyl-DOPA | Decrease | 72 h | Resistance | LC-MS/MS | Fruit | Ppal | [22] |
| Arylsulfatedferuloyl-DOPA | Decrease | 72 h | Resistance | LC-MS/MS | Fruit | Ppal | [22] |
| Proteíns | |||||||
| Protein name | Regulation | Time after inoculation | Resistance | Methodology | Tissue studied | Species | References |
| Vacuolar morphogenesis protein 7 (domain type: VAM7p-PX) | Expression | 3 days | Resistance | Transiente transformation/Western blot | Leaf | Ptrop/Ppal | [35] |
| Hepatocyte growth factor-regulated tyrosine kinase substrate (domain type: Hrs-2xFYVE) | Expression | 4 days | Resistance | Transiente transformation/Western blot | Leaf | Ptrop/Ppal | [35] |
| Pleckstrin homology domain-containing family A member 4 (domain type: PEPP1-PH) | Expression | 5 days | Resistance | Transiente transformation/Western blot | Leaf | Ptrop/Ppal | [35] |
| PH domain-containing protein (domain type: GmPh1-PH) | Expression | 6 days | Resistance | Transiente transformation/Western blot | Leaf | Ptrop/Ppal | [35] |
| List of differentially accumulated proteins (Table S2) | Differential | 48 h | Resistance | LC-MS/MS | Leaf | Ppal | [24] |
| Molecular Responses | |||||||
| Responses | Regulation | Time after inoculation | Resistance | Methodology | Tissue studied | Species | References |
| Signal transduction inducing a gene defense response | Induced | NS | Resistance | Macroarray | Leaf | Pmeg | [36] |
| Biosynthesis of phenylpropanoids | Induced | 24, 48 e 72 h | Resistance | RNA-Seq/RT-qPCR/KEGG | Leaf/Fruit | Ppal/Pmeg | [37] |
| Biosynthesis and action of ethylene | Induced | 24, 48 e 72 h | Resistance | RNA-Seq/RT-qPCR/KEGG | Leaf/Fruit | Ppal/Pmeg | [37] |
| Biosynthesis and action of jasmonic acid | Induced | 24, 48 e 72 h | Resistance | RNA-Seq/RT-qPCR/KEGG | Leaf/Fruit | Ppal/Pmeg | [37] |
| Defense signal transduction | Induced | 24, 48 e 72 h | Resistance | RNA-Seq/RT-qPCR/KEGG | Leaf/Fruit | Ppal/Pmeg | [37] |
| Endocytosis | Induced | 24, 48 e 72 h | Resistance | RNA-Seq/RT-qPCR/KEGG | Leaf/Fruit | Ppal/Pmeg | [37] |
| Response to stimuli and stress | Induced | 24 h | Resistance | RNA-Seq/BLAST2GO | Leaf | Pmeg | [21] |
| Signaling | Induced | 24 h | Resistance | RNA-Seq/BLAST2GO | Leaf | Pmeg | [21] |
| Morphological Characteristics | |||||||
| Characteristics | Attribute | Time after inoculation | Resistance | Methodology | Tissue studied | Species | References |
| Shell thickness | Bigger | Basal | Resistance | Measuring with scale | Fruit | Ppal | [38] |
| Shell hardness | Bigge | 2, 4, 6, 8, 10, 12 weeks | Resistance | Tensile/compression test | Root | Ppal | [39] |
| Epicarp thickness | Bigge | Basal | Resistance | Microscopy | Fruit | Ppal/Pmeg | [40] |
| Phloem fiber thickness | Bigge | Basal | Resistance | Microscopy | Fruit | Ppal/Pmeg | [40] |
| Length of vascular bundles | Bigge | Basal | Resistance | Microscopy | Fruit | Ppal/Pmeg | [40] |
| Distance between adjacent vascular bundles | Bigge | Basal | Resistance | Microscopy | Fruit | Ppal/Pmeg | [40] |
| Distance between vascular bundles and epicarp | Bigge | Basal | Resistance | Microscopy | Fruit | Ppal/Pmeg | [40] |
| Number of cells in the epicarp | Bigge | Basal | Resistance | Microscopy | Fruit | Ppal/Pmeg | [40] |
| Number of cells in the mesocarp | Bigge | Basal | Resistance | Microscopy | Fruit | Ppal/Pmeg | [40] |
| Number of vascular bundles | Smaller | Basal | Resistance | Microscopy | Fruit | Ppal/Pmeg | [40] |
| Stomatal frequency | Smalle | 72 h | Resistance | Microscopy | Fruit | Ppal | [41] |
| Pore length | Smalle | 72 h | Resistance | Microscopy | Fruit | Ppal | [41] |
| Physiological Responses | |||||||
| Responses | Attribute | Time after inoculation | Resistance | Methodology | Tissue studied | Species | References |
| Moisture content | Bigger | 2, 4, 6, 8, 10, 12 weeks | Susceptibility | Dry dough method | Root | Ppal | [39] |
| Moisture content | Variable | 72 h | Unrelated | Dry dough method | Fruit | Ppal | [41] |
| Moisture content | Low | Basal | Resistance | Dry dough method | Fruit | Ppal | [38] |
| Others | |||||||
| Compound | Content | Time after inoculation | Resistance | Methodology | Tissue studied | Species | References |
| Lignin | High | 2, 4, 6, 8, 10, 12 weeks | Resistance | Phloroglucinol-HCl test | Root | Ppal | [39] |
| Proline | Increase | 5 days | Resistance | Spectrophotometry | Fruit | Pmeg | [42] |
| Tyrosine | Increase | 5 days | Resistance | Thin layer chromatography | Fruit | Pmeg | [42] |
| Aspartate | Increase | 5 days | Resistance | Thin layer chromatography | Fruit | Pmeg | [42] |
| Soluble carbohydrates | Decrease | 5 days | Resistance | Spectrophotometry | Fruit | Pmeg | [42] |
| Proline | Presence | 6 days | Resistance | Thin layer chromatography | Leaf | Pmeg | [34] |
| Leucine | Presence | 6 days | Resistance | Thin layer chromatography | Leaf | Pmeg | [34] |
| Sucrose | Absence | 6 days | Resistance | Thin layer chromatography | Leaf | Pmeg | [34] |
| Synthetic peptides (US Patent # 5597945) | Expression | 3 days | Resistance | Transgenic | Leaf | Pcap/Ppal | [43] |
| Wax | High | 4 days | Resistance | Phenol wax wash | Leaf/Fruit | Ppal/Pmeg | [40] |
| Lignin | High | 6 days | Resistance | Thioglycolic acid test | Leaf/Fruit | Pmeg | [7] |
| Totals proteins | Increase | 6 days | Resistance | Nitrogen Content × 6.25 | Leaf/Fruit | Pmeg | [7] |
| Soluble and insoluble sugars | Increase | 6 days | Resistance | Phenol Sulfuric Method | Leaf/Fruit | Pmeg | [7] |
| Caffeine | High | NS | Resistance | RP-HPLC | Beans | NS | [44] |
4. Discussion
4.1. Few Primary Studies Address the Molecular and Morphological Aspects of the Interaction of T. cacao with Phytophthora spp.
4.2. Expression Patterns of Pattern Recognition Receptors (PRRs) Are Crucial in Cacao Resistance to Phytophthora spp.
4.3. Expression Patterns of Genes Encoding Pathogenesis-Related (PRs) Proteins Differ between Resistant and Susceptible Genotypes
4.4. Phenolic Compounds Play an Essential Role in Preformed Defenses
4.5. Possible Gene-to-Gene Interaction May Be a Factor in the Resistance of T. cacao to Phytophthora spp.
4.6. Genes Proposed via QTL Analysis Were Confirmed in Transcriptomic Studies
4.7. Accumulation of Proline May Be Involved in Cell Wall Integrity
4.8. A Study at the Level of Proteomics Reveals Crucial Proteins in the Interaction of Cacao and P. palmivora
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marelli, J.-P.; Guest, D.I.; Bailey, B.A.; Evans, H.C.; Brown, J.K.; Junaid, M.; Barreto, R.W.; Lisboa, D.O.; Puig, A.S. Chocolate under Threat from Old and New Cacao Diseases. Phytopathology 2019, 109, 1331–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, B.A.; Ali, S.S.; Akrofi, A.Y.; Meinhardt, L.W. Phytophthora megakarya, a Causal Agent of Black Pod Rot in Africa. In Cacao Diseases: A History of Old Enemies and New Encounters; Bailey, B.A., Meinhardt, L.W., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 267–303. ISBN 978-3-319-24789-2. [Google Scholar]
- Decloquement, J.; Ramos-Sobrinho, R.; Elias, S.G.; Britto, D.S.; Puig, A.S.; Reis, A.; da Silva, R.A.F.; Honorato-Júnior, J.; Luz, E.D.M.N.; Pinho, D.B.; et al. Phytophthora theobromicola sp. Nov.: A New Species Causing Black Pod Disease on Cacao in Brazil. Front. Microbiol. 2021, 12, 537399. [Google Scholar] [CrossRef] [PubMed]
- Guest, D. Black Pod: Diverse Pathogens with a Global Impact on Cocoa Yield. Phytopathology 2007, 97, 1650–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surujdeo-Maharaj, S.; Sreenivasan, T.N.; Motilal, L.A.; Umaharan, P. Black Pod and Other Phytophthora Induced Diseases of Cacao: History, Biology, and Control. In Cacao Diseases: A History of Old Enemies and New Encounters; Bailey, B.A., Meinhardt, L.W., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 213–266. ISBN 978-3-319-24789-2. [Google Scholar]
- Barreto, M.A.; Santos, J.C.S.; Corrêa, R.X.; Luz, E.D.M.N.; Marelli, J.; Souza, A.P. Detection of Genetic Resistance to Cocoa Black Pod Disease Caused by Three Phytophthora Species. Euphytica 2015, 206, 677–687. [Google Scholar] [CrossRef]
- Nyadanu, D.; Akromah, R.; Adomako, B.; Kwoseh, C.; Lowor, S.T.; Dzahini-Obiatey, H.; Akrofi, A.Y.; Ansah, F.O.; Asiama, Y.O.; Assuah, M.K. Biochemical Mechanisms of Resistance to Black Pod Disease in Cocoa (Theobroma cacao L.). Am. J. Biochem. Mol. Biol. 2013, 3, 20–37. [Google Scholar] [CrossRef] [Green Version]
- Barreto, M.A.; Rosa, J.R.B.F.; Holanda, I.S.A.; Cardoso-Silva, C.B.; Vildoso, C.I.A.; Ahnert, D.; Souza, M.M.; Corrêa, R.X.; Royaert, S.; Marelli, J.; et al. QTL Mapping and Identification of Corresponding Genomic Regions for Black Pod Disease Resistance to Three Phytophthora Species in Theobroma cacao L. Euphytica 2018, 214, 188. [Google Scholar] [CrossRef]
- Risterucci, A.M.; Paulin, D.; Ducamp, M.; N’Goran, J.A.K.; Lanaud, C. Identification of QTLs Related to Cocoa Resistance to Three Species of Phytophthora. Theor. Appl. Genet. 2003, 108, 168–174. [Google Scholar] [CrossRef]
- Argout, X.; Salse, J.; Aury, J.-M.; Guiltinan, M.J.; Droc, G.; Gouzy, J.; Allegre, M.; Chaparro, C.; Legavre, T.; Maximova, S.N.; et al. The Genome of Theobroma cacao. Nat. Genet. 2011, 43, 101–108. [Google Scholar] [CrossRef]
- Araújo, I.S.; De Souza Filho, G.A.; Pereira, M.G.; Faleiro, F.G.; De Queiroz, V.T.; Guimarães, C.T.; Moreira, M.A.; De Barros, E.G.; Machado, R.C.R.; Pires, J.L.; et al. Mapping of Quantitative Trait Loci for Butter Content and Hardness in Cocoa Beans (Theobroma cacao L.). Plant Mol. Biol. Rep. 2009, 27, 177–183. [Google Scholar] [CrossRef]
- Clement, D.; Risterucci, A.M.; Motamayor, J.C.; N’Goran, J.; Lanaud, C. Mapping QTL for Yield Components, Vigor, and Resistance to Phytophthora palmivora in Theobroma cacao L. Genome 2003, 46, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Crouzillat, D.; Phillips, W.; Fritz, P.J.; Pétiard, V. Quantitative Trait Loci Analysis in Theobroma cacao Using Molecular Markers. Inheritance of Polygenic Resistance to Phytophthora palmivora in Two Related Cacao Populations. Euphytica 2000, 114, 25–36. [Google Scholar] [CrossRef]
- Flament, M.-H.; Kebe, I.; Clément, D.; Pieretti, I.; Risterucci, A.-M.; N’Goran, J.-A.-K.; Cilas, C.; Despréaux, D.; Lanaud, C. Genetic Mapping of Resistance Factors to Phytophthora palmivora in Cocoa. Génome 2001, 44, 79–85. [Google Scholar] [CrossRef]
- Lanaud, C.; Fouet, O.; Clément, D.; Boccara, M.; Risterucci, A.M.; Surujdeo-Maharaj, S.; Legavre, T.; Argout, X. A Meta–QTL Analysis of Disease Resistance Traits of Theobroma cacao L. Mol. Breed. 2009, 24, 361–374. [Google Scholar] [CrossRef]
- Santos, E.S.L. Thesis Abstract Characterization of the Resistance Gradient of Cacao Genotypes (F2 Sca6 x ICS1) to Black Pod Disease Using Phytopathometric, Genetic and Molecular Approaches. Genet. Mol. Res. 2009, 8, 1307–1308. [Google Scholar] [CrossRef]
- Schnell, R.J.; Kuhn, D.N.; Brown, J.S.; Olano, C.T.; Phillips-Mora, W.; Amores, F.M.; Motamayor, J.C. Development of a Marker Assisted Selection Program for Cacao. Phytopathology 2007, 97, 1664–1669. [Google Scholar] [CrossRef] [Green Version]
- Nyassé, S.; Cilas, C.; Herail, C.; Blaha, G. Leaf Inoculation as an Early Screening Test for Cocoa (Theobroma cacao L.) Resistance to Phytophthora Black Pod Disease. Crop Prot. 1995, 14, 657–663. [Google Scholar] [CrossRef]
- Clément, D.; Lanaud, C.; Sabau, X.; Fouet, O.; Le Cunff, L.; Ruiz, E.; Risterucci, A.M.; Glaszmann, J.C.; Piffanelli, P. Creation of BAC Genomic Resources for Cocoa (Theobroma cacao L.) for Physical Mapping of RGA Containing BAC Clones. Theor. Appl. Genet. 2004, 108, 1627–1634. [Google Scholar] [CrossRef] [PubMed]
- Pugh, T.; Fouet, O.; Risterucci, A.M.; Brottier, P.; Abouladze, M.; Deletrez, C.; Courtois, B.; Clement, D.; Larmande, P.; N’Goran, J.A.K.; et al. A New Cacao Linkage Map Based on Codominant Markers: Development and Integration of 201 New Microsatellite Markers. Theor. Appl. Genet. 2004, 108, 1151–1161. [Google Scholar] [CrossRef]
- Pokou, D.N.; Fister, A.S.; Winters, N.; Tahi, M.; Klotioloma, C.; Sebastian, A.; Marden, J.H.; Maximova, S.N.; Guiltinan, M.J. Resistant and Susceptible Cacao Genotypes Exhibit Defense Gene Polymorphism and Unique Early Responses to Phytophthora megakarya Inoculation. Plant Mol. Biol. 2019, 99, 499–516. [Google Scholar] [CrossRef]
- Knollenberg, B.J.; Li, G.-X.; Lambert, J.D.; Maximova, S.N.; Guiltinan, M.J. Clovamide, a Hydroxycinnamic Acid Amide, Is a Resistance Factor Against Phytophthora spp. in Theobroma cacao. Front. Plant Sci. 2020, 11, 617520. [Google Scholar] [CrossRef]
- Bailey, B.A.; Bae, H.; Strem, M.D.; Antúnez De Mayolo, G.; Guiltinan, M.J.; Verica, J.A.; Maximova, S.N.; Bowers, J.H. Developmental Expression of Stress Response Genes in Theobroma cacao Leaves and Their Response to Nep1 Treatment and a Compatible Infection by Phytophthora megakarya. Plant Physiol. Biochem. 2005, 43, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Rego, A.P.B.; Mora-Ocampo, I.Y.; Pirovani, C.P.; Luz, E.D.M.N.; Corrêa, R.X. Protein Level Defense Responses of Theobroma cacao Interaction with Phytophthora palmivora. Front. Agron. 2022, 4, 836360. [Google Scholar] [CrossRef]
- Baruah, I.K.; Shahin, S.A.; Shao, J.; Lary, D.; Bailey, B.A. Changes in Gene Expression in Leaves of Cacao Genotypes Resistant and Susceptible to Phytophthora palmivora Infection. Front. Plant Sci. 2022, 12, 780805. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, S.; Silva, C.; Hernandes, E.; Octaviano, F.; Di Thommazo, A.; Belgamo, A. Improvements in the StArt Tool to Better Support the Systematic Review Process. In Proceedings of the 20th International Conference on Evaluation and Assessment in Software Engineering, Limerick, Ireland, 1–3 June 2016; ACM: New York, NY, USA, 2016; pp. 1–5. [Google Scholar]
- Spence, J.A. Black-Pod Disease of Cocoa: II. A Study of Host-Parasite Relations. Ann. Appl. Biol. 1961; 49, 723–734. [Google Scholar] [CrossRef]
- Okey, E.N.; Duncan, E.J.; Sirju-Charran, G.; Sreenivasan, T.N. Phytophthora Canker Resistance in Cacao: Role of Peroxidase, Polyphenoloxidase and Phenylalanine Ammonia-Lyase. J. Phytopathol. 1997, 145, 295–299. [Google Scholar] [CrossRef]
- Simo, C.; Djocgoue, P.F.; Mbouobda, H.D.; Effa, P.O.; Boudjeko, T.; Omokolo, D.N. Variation and Heritability of Polyphenoloxidasic Activities in Two Hybrid Families of Theobroma cacao L. after Cocoa Pods Inoculation with Phytophtora megakarya Bras. et Grif. Plant Pathol. J. 2011, 10, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Ndoumou, O.D.; Ndzomo, T.G.; Djocgoue, P.F. Changes in Carbohydrate, Amino Acid and Phenol Contents in Cocoa Pods from Three Clones after Infection with Phytophthora megakarya Bra. and Grif. Ann. Bot. 1996, 77, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Okey, E.N.; Duncan, E.J.; Sirju-Charran, G.; Sreenvisasan, T.N. Zoospore Germination and Growth of Phytophthora palmivora in Stem Extracts as Criteria for Assessing Cacao Resistance to Canker. Mycol. Res. 1997, 101, 683–686. [Google Scholar] [CrossRef]
- Djocgoue, P.F.; Boudjeko, T.; Mbouobda, H.D.; Nankeu, D.J.; Hadrami, I.E.; Omokolo, N.D. Heritability of Phenols in the Resistance of Theobroma cacao against Phytophthora megakarya, the Causal Agent of Black Pod Disease. J. Phytopathol. 2007, 155, 519–525. [Google Scholar] [CrossRef]
- Boudjeko, T.; Djocgoue, P.F.; Nankeu, J.D.; Mbouobda, H.D.; Omokolo, D.N.; El Hadrami, I. Luteolin Derivatives and Heritability of Resistance to Phytophthora megakarya in Theobroma cacao. Aust. Plant Pathol. 2007, 36, 56. [Google Scholar] [CrossRef]
- Djocgoue, P.F.; Mbouobda, H.D.; Boudjeko, T.; Effa, P.O.; Omokolo, D.N. Amino Acids, Carbohydrates and Heritability of Resistance in the Theobroma cacao/Phythophthora megakarya Interaction. Phytopathol. Mediterr. 2011, 50, 370–383. [Google Scholar]
- Helliwell, E.E.; Vega-Arreguín, J.; Shi, Z.; Bailey, B.; Xiao, S.; Maximova, S.N.; Tyler, B.M.; Guiltinan, M.J. Enhanced Resistance in Theobroma cacao against Oomycete and Fungal Pathogens by Secretion of Phosphatidylinositol-3-Phosphate-Binding Proteins. Plant Biotechnol. J. 2016, 14, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Legavre, T.; Ducamp, M.; Sabau, X.; Argout, X.; Fouet, O.; Dedieu, F.; Surujdeo-Maharaj, S.; Garcia, D.; Paulin, D.; Lanaud, C. Identification of Theobroma cacao Genes Differentially Expressed during Phytophthora megakarya Infection. Physiol. Mol. Plant Pathol. 2015, 92, 1–13. [Google Scholar] [CrossRef]
- Ali, S.S.; Shao, J.; Lary, D.J.; Strem, M.D.; Meinhardt, L.W.; Bailey, B.A. Phytophthora megakarya and P. palmivora, Causal Agents of Black Pod Rot, Induce Similar Plant Defense Responses Late during Infection of Susceptible Cacao Pods. Front. Plant Sci. 2017, 8, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyadanu, D.; Assuah, M.K.; Adomako, B.; Asiama, Y.O.; Adu-Ampoma, Y. Thickness of the Cocoa Pod Husk and Its Moisture Content as Resistance Factors to Phytophthora Pod Rot. Int. J. Agric. Res. 2011, 6, 310–322. [Google Scholar] [CrossRef] [Green Version]
- Okey, E.N.; Duncan, E.J.; Sirju-Charran, G.; Sreenivasan, T.N. Wound-healing in Cocoa (Theobroma cacao L.) Stems and Its Effect on Canker Caused by Phytophthora palmivora (Butl.) Butler. Int. J. Pest Manag. 1995, 41, 224–228. [Google Scholar] [CrossRef]
- Nyadanu, D.; Akromah, R.; Adomako, B.; Kwoseh, C.; Lowor, S.T.; Dzahini-Obiatey, H.; Akrofi, A.Y.; Assuah, M.K. Inheritance and General Combining Ability Studies of Detached Pod, Leaf Disc and Natural Field Resistance to Phytophthora palmivora and Phytophthora megakarya in Cacao (Theobroma cacao L.). Euphytica 2012, 188, 253–264. [Google Scholar] [CrossRef]
- Iwaro, A.D.; Sreenivasan, T.N.; Spence, J.A. Studies on Black Pod Disease in Trinidad. In Proceedings of the International Workshop on the Contribution of Disease Resistance to Cocoa Variety Improvement, Salvador, Bahia, Brazil, 24–26 November 1996; Volume 1999, pp. 67–74. [Google Scholar]
- Omokolo, N.D.; Nankeu, D.J.; Niemenak, N.; Djocgoue, P.F. Analysis of Amino Acids and Carbohydrates in the Cortex of Nine Clones of Theobroma cacao L. in Relation to Their Susceptibility to Phytophthora megakarya Bra. and Grif. Crop Prot. 2002, 21, 395–402. [Google Scholar] [CrossRef]
- Mejía, L.C.; Guiltinan, M.J.; Shi, Z.; Landherr, L.; Maximova, S.N. Expression of Designed Antimicrobial Peptides in Theobroma cacao L. Trees Reduces Leaf Necrosis Caused by Phytophthora spp.; Rajasekaran, K., Cary, J.W., Jaynes, J.M., Montesinos, E., Eds.; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2012; Volume 1095, pp. 379–395. ISBN 978-0-8412-2748-4. [Google Scholar]
- Kieck, J.S.; Zug, K.L.M.; Huamaní Yupanqui, H.A.; Gómez Aliaga, R.; Cierjacks, A. Plant Diversity Effects on Crop Yield, Pathogen Incidence, and Secondary Metabolism on Cacao Farms in Peruvian Amazonia. Agric. Ecosyst. Environ. 2016, 222, 223–234. [Google Scholar] [CrossRef]
- Report Shows United States Leads in Science and Technology as China Rapidly Advances. Available online: https://www.sciencedaily.com/releases/2018/01/180124113951.htm (accessed on 28 March 2023).
- Mucherino Muñoz, J.J.; De Melo, C.A.F.; Santana Silva, R.J.; Luz, E.D.M.N.; Corrêa, R.X. Structural and Functional Genomics of the Resistance of Cacao to Phytophthora palmivora. Pathogens 2021, 10, 961. [Google Scholar] [CrossRef]
- Lim, C.W.; Yang, S.H.; Shin, K.H.; Lee, S.C.; Kim, S.H. The AtLRK10L1.2, Arabidopsis Ortholog of Wheat LRK10, Is Involved in ABA-Mediated Signaling and Drought Resistance. Plant Cell Rep. 2015, 34, 447–455. [Google Scholar] [CrossRef]
- Tang, D.; Wang, G.; Zhou, J.-M. Receptor Kinases in Plant-Pathogen Interactions: More Than Pattern Recognition. Plant Cell 2017, 29, 618–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fister, A.S.; Mejia, L.C.; Zhang, Y.; Herre, E.A.; Maximova, S.N.; Guiltinan, M.J. Theobroma cacao L. Pathogenesis-Related Gene Tandem Array Members Show Diverse Expression Dynamics in Response to Pathogen Colonization. BMC Genom. 2016, 17, 363. [Google Scholar] [CrossRef] [Green Version]
- Fister, A.S.; Shi, Z.; Zhang, Y.; Helliwell, E.E.; Maximova, S.N.; Guiltinan, M.J. Protocol: Transient Expression System for Functional Genomics in the Tropical Tree Theobroma cacao L. Plant Methods 2016, 12, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of Inducible Defense-Related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fister, A.S.; O’Neil, S.T.; Shi, Z.; Zhang, Y.; Tyler, B.M.; Guiltinan, M.J.; Maximova, S.N. Two Theobroma cacao Genotypes with Contrasting Pathogen Tolerance Show Aberrant Transcriptional and ROS Responses after Salicylic Acid Treatment. J. Exp. Bot. 2015, 66, 6245–6258. [Google Scholar] [CrossRef] [Green Version]
- Lev-Yadun, S.; Gould, K.S. Role of Anthocyanins in Plant Defence. In Anthocyanins; Winefield, C., Davies, K., Gould, K., Eds.; Springer: New York, NY, USA, 2008; pp. 22–28. ISBN 978-0-387-77334-6. [Google Scholar]
- Marinova, K.; Pourcel, L.; Weder, B.; Schwarz, M.; Barron, D.; Routaboul, J.-M.; Debeaujon, I.; Klein, M. The Arabidopsis MATE Transporter TT12 Acts as a Vacuolar Flavonoid/H+-Antiporter Active in Proanthocyanidin-Accumulating Cells of the Seed Coat. Plant Cell 2007, 19, 2023–2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballvora, A.; Ercolano, M.R.; Weiss, J.; Meksem, K.; Bormann, C.A.; Oberhagemann, P.; Salamini, F.; Gebhardt, C. The R1 Gene for Potato Resistance to Late Blight (Phytophthora infestans) Belongs to the Leucine Zipper/NBS/LRR Class of Plant Resistance Genes. Plant J. 2002, 30, 361–371. [Google Scholar] [CrossRef]
- Van Der Vossen, E.; Sikkema, A.; Hekkert, B.T.L.; Gros, J.; Stevens, P.; Muskens, M.; Wouters, D.; Pereira, A.; Stiekema, W.; Allefs, S. An Ancient R Gene from the Wild Potato Species Solanum bulbocastanum Confers Broad-Spectrum Resistance to Phytophthora infestans in Cultivated Potato and Tomato. Plant J. 2003, 36, 867–882. [Google Scholar] [CrossRef]
- Kishor, P.B.K. Role of Proline in Cell Wall Synthesis and Plant Development and Its Implications in Plant Ontogeny. Front. Plant Sci. 2015, 6, 544. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, R.; Barchi, B.; Turlapati, S.A.; Gagne, M.; Minocha, R.; Long, S.; Minocha, S.C. Glutamate, Ornithine, Arginine, Proline, and Polyamine Metabolic Interactions: The Pathway Is Regulated at the Post-Transcriptional Level. Front. Plant Sci. 2016, 7, 78. [Google Scholar] [CrossRef] [Green Version]
- Bae, H.; Kim, S.-H.; Kim, M.S.; Sicher, R.C.; Lary, D.; Strem, M.D.; Natarajan, S.; Bailey, B.A. The Drought Response of Theobroma cacao (Cacao) and the Regulation of Genes Involved in Polyamine Biosynthesis by Drought and Other Stresses. Plant Physiol. Biochem. 2008, 46, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Perl, K.; Ushakov, K.; Pozniak, Y.; Yizhar-Barnea, O.; Bhonker, Y.; Shivatzki, S.; Geiger, T.; Avraham, K.B.; Shamir, R. Reduced Changes in Protein Compared to MRNA Levels across Non-Proliferating Tissues. BMC Genom. 2017, 18, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, C.; Marcotte, E.M. Insights into the Regulation of Protein Abundance from Proteomic and Transcriptomic Analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
| String Tested on Scopus a | Found Articles | Useful Articles for Answering the Question | Sentinel Articles in the First 20 Articles on the List b |
|---|---|---|---|
| (cacao OR cocoa) AND (Phytophthora) | 819 | 2 | 2 |
| ((cacao OR cocoa) AND (Phytophthora)) | 385 | 3 | 2 |
| ((cacao OR cocoa) AND (Phytophthora) AND (resistance OR response)) | 145 | 4 | 3 |
| ((cacao OR cocoa) AND (resistance OR response* OR interaction OR defense) AND (Phytophthora)) | 165 | 4 | 3 |
| ((cacao OR cocoa) AND (resistance OR response* OR interaction OR defense OR protein* OR gene*) AND (Phytophthora)) | 231 | 4 | 4 |
| Type of analysis | Genetic |
| Transcriptomic | |
| Proteomic | |
| Biochemical | |
| Physiological | |
| Morphological | |
| Plant tissue studied | Fruit |
| Stem | |
| Leaf | |
| Root | |
| Seed | |
| Reported element of T. cacao (resistant and susceptible) that participates in the interaction with species of Phytophthora | Protein |
| Gene | |
| Enzyme | |
| Metabolite | |
| Morphological structure | |
| Molecular response | |
| Physiological response | |
| Others | |
| Species of Phytophthora | |
| Country where he was accomplished the study | |
| Year of publication |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rêgo, A.P.B.; Mora-Ocampo, I.Y.; Corrêa, R.X. Interactions of Different Species of Phytophthora with Cacao Induce Genetic, Biochemical, and Morphological Plant Alterations. Microorganisms 2023, 11, 1172. https://doi.org/10.3390/microorganisms11051172
Rêgo APB, Mora-Ocampo IY, Corrêa RX. Interactions of Different Species of Phytophthora with Cacao Induce Genetic, Biochemical, and Morphological Plant Alterations. Microorganisms. 2023; 11(5):1172. https://doi.org/10.3390/microorganisms11051172
Chicago/Turabian StyleRêgo, Angra Paula Bomfim, Irma Yuliana Mora-Ocampo, and Ronan Xavier Corrêa. 2023. "Interactions of Different Species of Phytophthora with Cacao Induce Genetic, Biochemical, and Morphological Plant Alterations" Microorganisms 11, no. 5: 1172. https://doi.org/10.3390/microorganisms11051172