

Effects of LPS from Rhodobacter sphaeroides, a Purple Non-Sulfur Bacterium (PNSB), on the Gene Expression of Rice Root


Abstract
:1. Introduction
2. Materials and Methods
2.1. Rice Cultivar
2.2. PNSB Strain and LPS
2.3. Cultivation of Rice Seedlings on Agar Plates
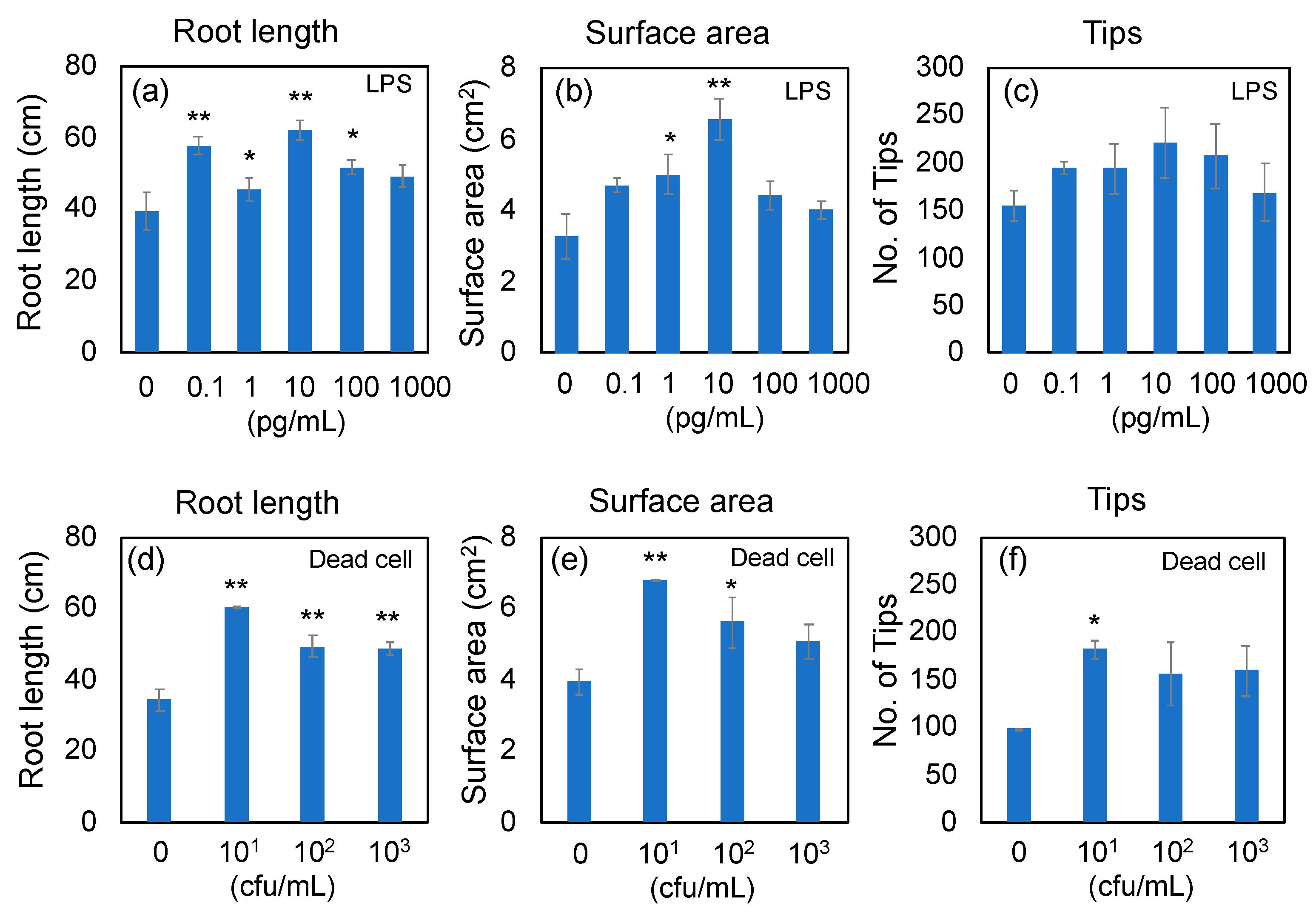
2.4. Analysis of Root Development by WinRhizo Image Analyzing System
2.5. RNA Extraction and RNA-seq
2.6. Statistical Analysis
3. Results
3.1. Effects of LPS from PNSB (R. sphaeroides NBRC 12203T) and Dead PNSB Cells on the Root Development of Rice Seedlings
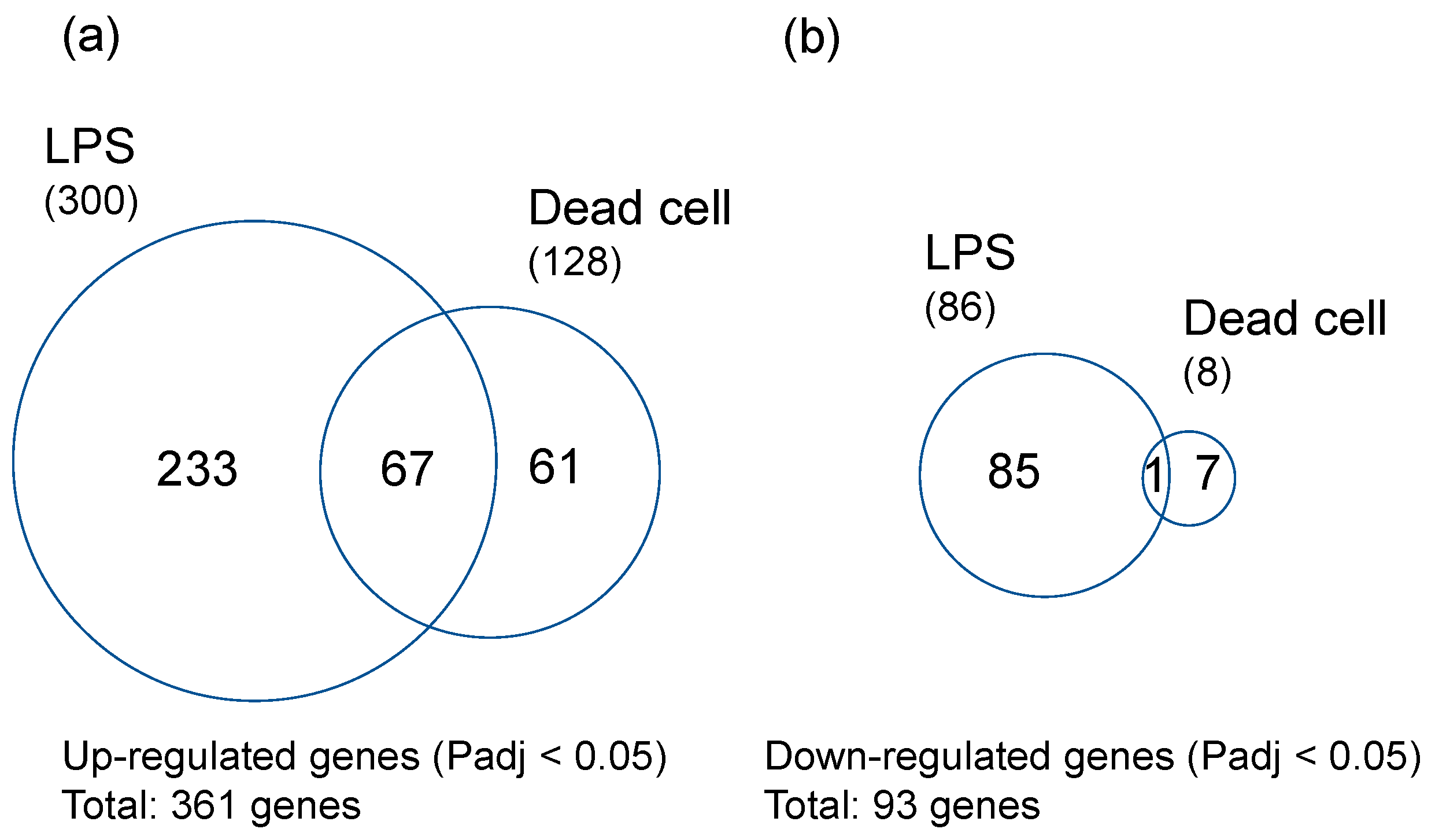
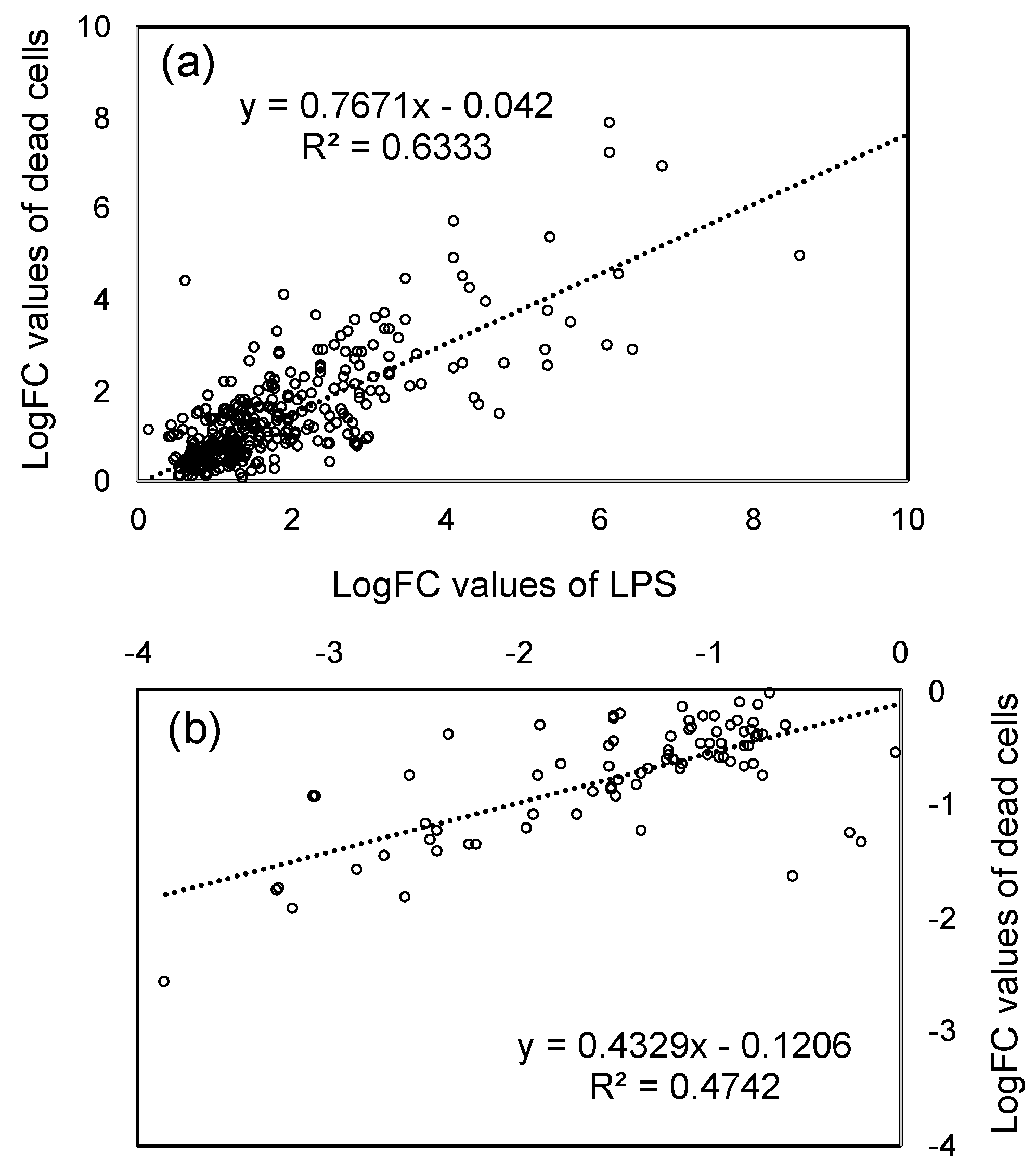
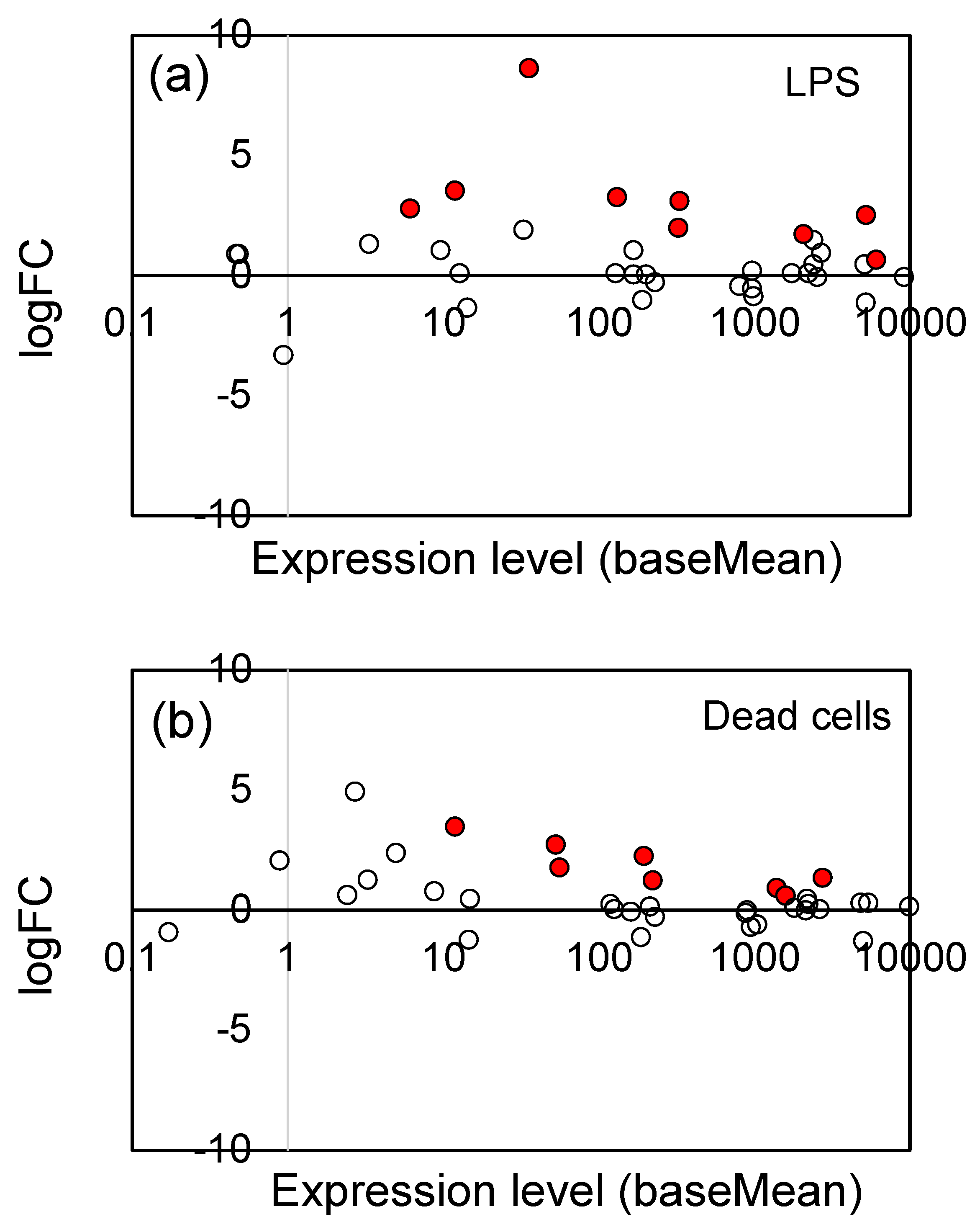
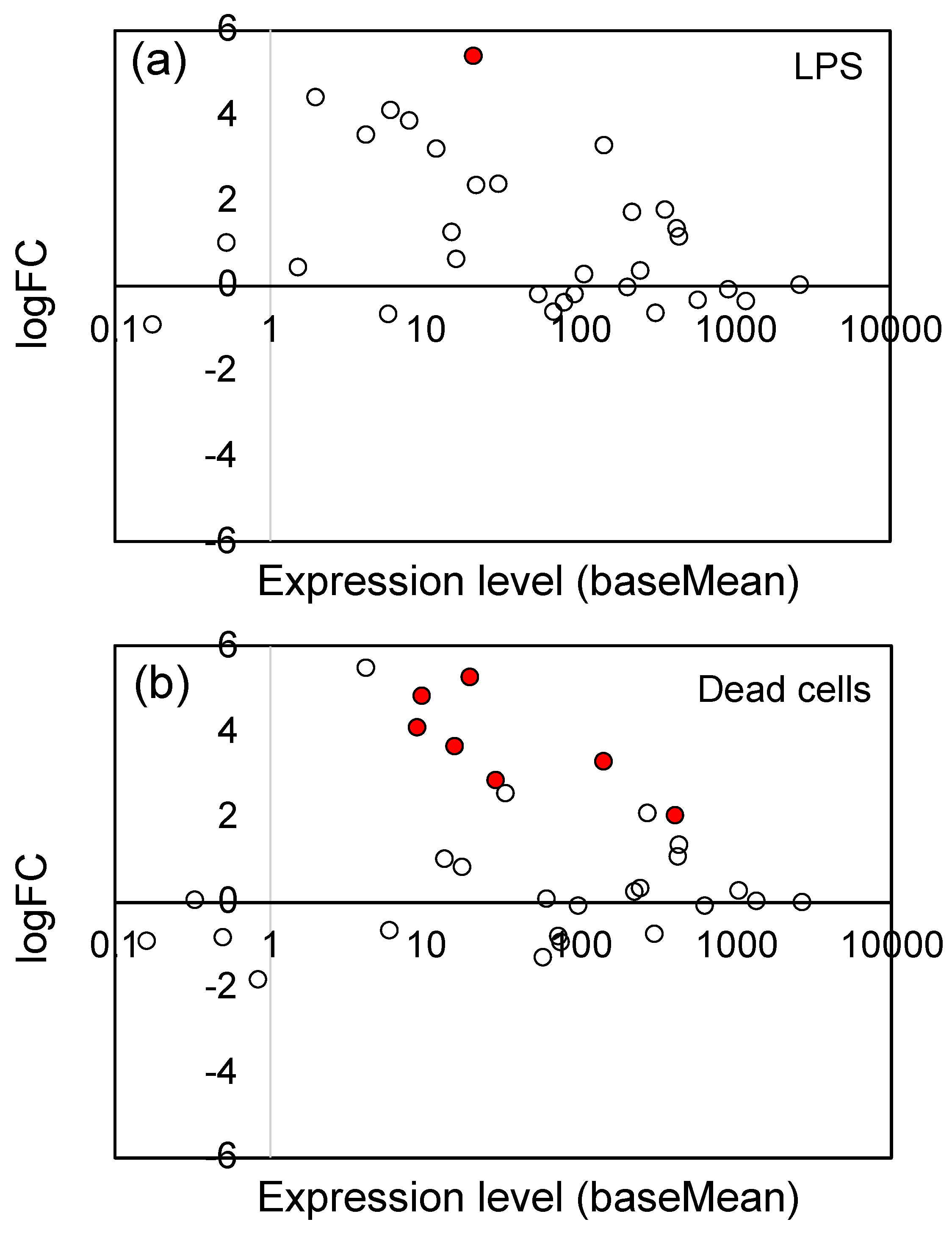
3.2. Effects of LPS from PNSB and Dead PNSB Cells on the Gene Expression of the Root of Rice Seedlings -RNA-Seq Analyses-
3.3. Effects of LPS from PNSB and Dead PNSB Cells on the Gene Expression of the Gene-Related JA Signaling Pathway
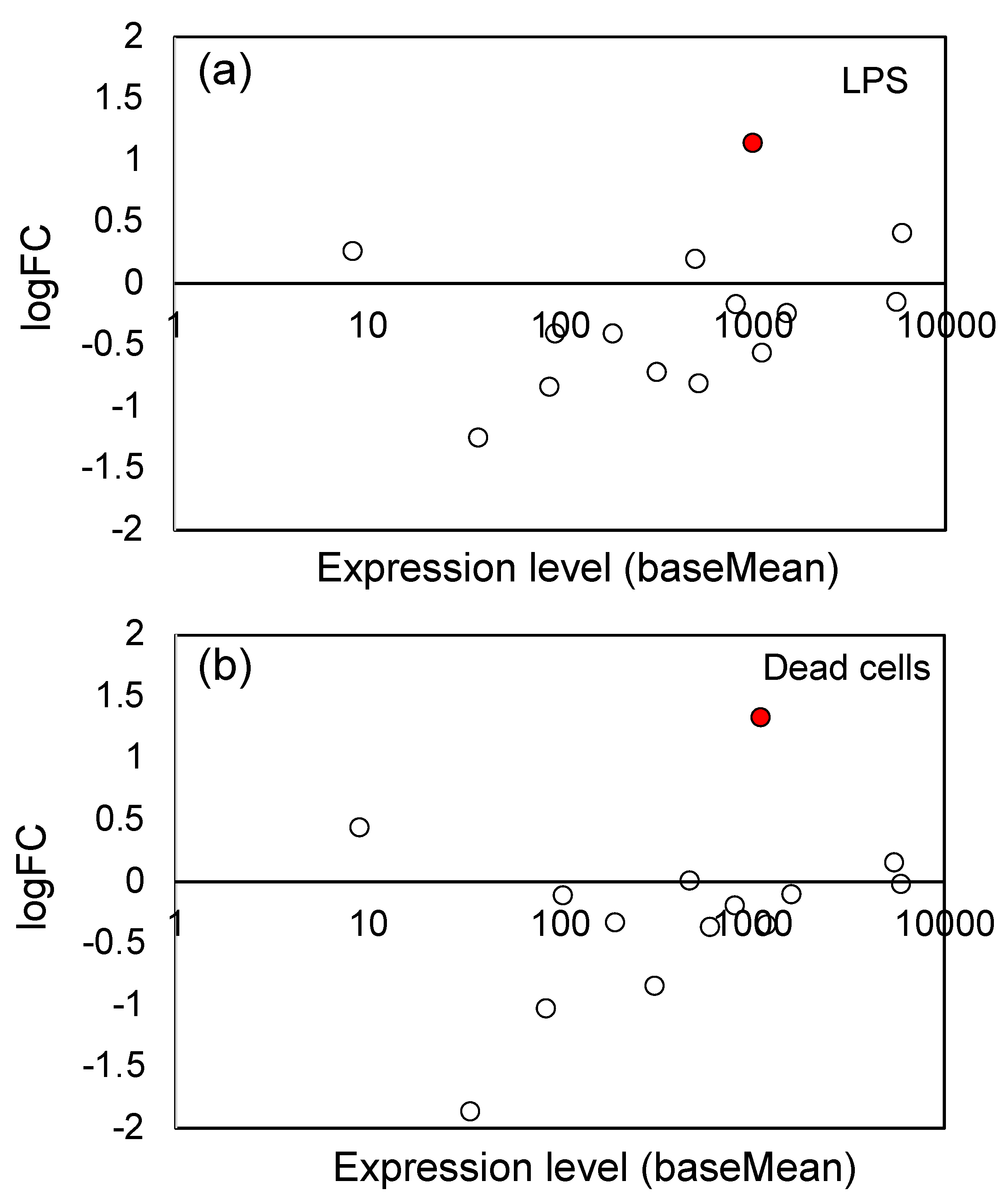
3.4. Effects of LPS from PNSB and Dead PNSB Cells on the Gene Expression of the Genes-Related Biosynthesis of Chalcone and Other Secondary Metabolites
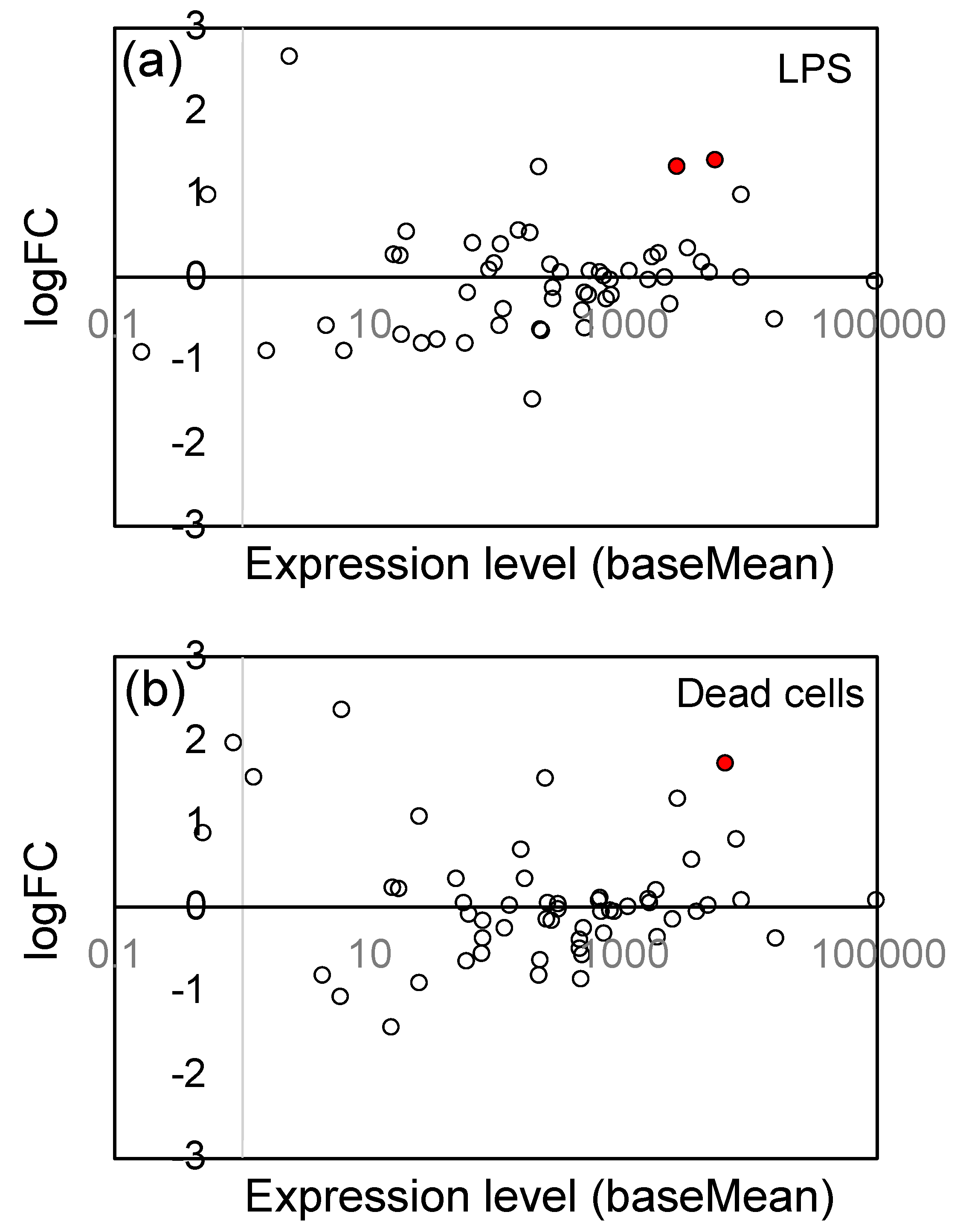
3.5. Effects of LPS from PNSB and Dead PNSB Cells on the Gene Expression of the Genes-Related Reactive Oxygen Species (ROS) Generation/Elimination
4. Discussion
4.1. Unique Property of LPS from R. sphaeroides NBRC 12203T
4.2. Comparison of the Effects of LPS and Dead Cells of PNSB on the Gene Expression of Rice Seedlings
4.3. Effects of LPS on the Expression of Genes Related to the JA Signaling Pathway
4.4. Effects of LPS on the Expression of Genes Related to Secondary Metabolism
4.5. Effects of LPS on the Expression of Genes Related to ROS Generation/Elimination
4.6. Promotion of Root Development by LPS
4.7. Stimulation of Defense Response by LPS
4.8. Practical Application of LPS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oelze, J.; Drews, G. Membranes of photosynthetic bacteria. Biochim. Biophys. Acta-Rev. Biomembr. 1972, 265, 209–239. [Google Scholar] [CrossRef]
- George, D.M.; Vincent, A.S.; Mackey, H.R. An overview of anoxygenic phototrophic bacteria and their applications in environmental biotechnology for sustainable Resource recovery. Biotechnol. Rep. 2020, 28, e00563. [Google Scholar] [CrossRef]
- Sakarika, M.; Spanoghe, J.; Sui, Y.; Wambacq, E.; Grunert, O.; Haesaert, G.; Spiller, M.; Vlaeminck, S.E. Purple non-sulphur bacteria and plant production: Benefits for fertilization, stress resistance and the environment. Microb. Biotechnol. 2020, 13, 1336–1365. [Google Scholar] [CrossRef] [Green Version]
- Wong, W.-T.; Tseng, C.-H.; Hsu, S.-H.; Lur, H.-S.; Mo, C.-W.; Huang, C.-N.; Hsu, S.-C.; Lee, K.-T.; Liu, C.-T. Promoting effects of a single Rhodopseudomonas palustris inoculant on plant growth by Brassica rapa chinensis under low fertilizer input. Microbes Environ. 2014, 29, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Sundar, L.S.; Chao, Y.Y. Potential of purple non-sulfur bacteria in sustainably enhancing the agronomic and physiological performances of rice. Agronomy 2022, 12, 2347. [Google Scholar] [CrossRef]
- Qi, Z.; Zhang, X.H.; Boon, N.; Bossier, P. Probiotics in aquaculture of China—Current state, problems and prospect. Aquaculture 2009, 290, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Miyasaka, H.; Koga, A.; Maki, T. Recent progress in the use of purple non-sulfur bacteria as probiotics in aquaculture. World J. Microbiol. Biotechnol. 2023, 39, 145. [Google Scholar] [CrossRef]
- Capson-Tojo, G.; Batstone, D.J.; Grassino, M.; Vlaeminck, S.E.; Puyol, D.; Verstraete, W.; Kleerebezem, R.; Oehmen, A.; Ghimire, A.; Pikaar, I.; et al. Purple phototrophic bacteria for resource recovery: Challenges and opportunities. Biotechnol Adv 2020, 43, 107567. [Google Scholar] [CrossRef]
- Higuchi-Takeuchi, M.; Numata, K. Marine purple photosynthetic bacteria as sustainable microbial production hosts. Front. Bioeng. Biotechnol. 2019, 7, 258. [Google Scholar]
- Kikuchi, Y.; Umekage, S. Extracellular nucleic acids of the marine bacterium Rhodovulum sulfidophilum and recombinant RNA production technology using bacteria. FEMS Microbiol. Lett. 2018, 365, fnx268. [Google Scholar] [CrossRef]
- Ghosh, S.; Dairkee, U.K.; Chowdhury, R.; Bhattacharya, P. Hydrogen from food processing wastes via photofermentation using Purple Non-sulfur Bacteria (PNSB)—A review. Energy Convers. Manag. 2017, 141, 299–314. [Google Scholar] [CrossRef]
- Sagir, E.; Alipour, S. Photofermentative hydrogen production by immobilized photosynthetic bacteria: Current perspectives and challenges. Renew. Sustain. Energy Rev. 2021, 141, 110796. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, G.; He, S.; Zhao, R.; Zhu, D. Purple non-sulfur bacteria technology: A promising and potential approach for wastewater treatment and bioresources recovery. World J. Microbiol. Biotechnol. 2021, 37, 161. [Google Scholar] [CrossRef]
- Nookongbut, P.; Kantachote, D.; Krishnan, K.; Megharaj, M. Arsenic resistance genes of As-resistant purple nonsulfur bacteria isolated from As-contaminated sites for bioremediation application. J. Basic Microbiol. 2017, 57, 316–324. [Google Scholar] [CrossRef]
- Lee, S.K.; Lur, H.S.; Liu, C. Te From lab to farm: Elucidating the beneficial roles of photosynthetic bacteria in sustainable agriculture. Microorganisms 2021, 9, 2453. [Google Scholar] [CrossRef]
- Sakpirom, J.; Kantachote, D.; Nunkaew, T.; Khan, E. Characterizations of purple non-sulfur bacteria isolated from paddy fields, and identification of strains with potential for plant growth-promotion, greenhouse gas mitigation and heavy metal bioremediation. Res. Microbiol. 2017, 168, 266–275. [Google Scholar] [CrossRef]
- Sasaki, K.; Watanabe, M.; Tanaka, T.; Tanaka, T. Biosynthesis, biotechnological production and applications of 5-aminolevulinic acid. Appl. Microbiol. Biotechnol. 2002, 58, 23–29. [Google Scholar] [CrossRef]
- Hayashi, S.; Iwamoto, Y.; Hirakawa, Y.; Mori, K.; Yamada, N.; Maki, T.; Yamamoto, S.; Miyasaka, H. Plant-growth-promoting effect by cell components of purple non-sulfur photosynthetic bacteria. Microorganisms 2022, 10, 771. [Google Scholar] [CrossRef]
- Iwai, R.; Uchida, S.; Yamaguchi, S.; Sonoda, F.; Tsunoda, K.; Nagata, H.; Nagata, D.; Koga, A.; Goto, M.; Maki, T.; et al. Effects of seed bio-priming by purple non-sulfur bacteria (PNSB) on the root development of rice. Microorganisms 2022, 10, 2197. [Google Scholar] [CrossRef]
- Kagan, J.C. Lipopolysaccharide Detection across the Kingdoms of Life. Trends Immunol. 2017, 38, 696–704. [Google Scholar] [CrossRef]
- Ranf, S. Immune sensing of lipopolysaccharide in plants and animals: Same but different. PLoS Pathog. 2016, 12, e1005596. [Google Scholar] [CrossRef]
- Desaki, Y.; Miya, A.; Venkatesh, B.; Tsuyumu, S.; Yamane, H.; Kaku, H.; Minami, E.; Shibuya, N. Bacterial lipopolysaccharides induce defense responses associated with programmed cell death in rice cells. Plant Cell Physiol. 2006, 47, 1530–1540. [Google Scholar] [CrossRef]
- Iizasa, S.; Iizasa, E.; Matsuzaki, S.; Tanaka, H.; Kodama, Y.; Watanabe, K.; Nagano, Y. Arabidopsis LBP/BPI related-1 and -2 bind to LPS directly and regulate PR1 expression. Sci. Rep. 2016, 6, 27527. [Google Scholar] [CrossRef] [Green Version]
- Shang-Guan, K.; Wang, M.; Htwe, N.M.P.S.; Li, P.; Li, Y.; Qi, F.; Zhang, D.; Cao, M.; Kim, C.; Weng, H.; et al. Lipopolysaccharides trigger two successive bursts of reactive oxygen species at distinct cellular locations. Plant Physiol. 2018, 176, 2543–2556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finnegan, T.; Steenkamp, P.A.; Piater, L.A.; Dubery, I.A. The lipopolysaccharide-induced metabolome signature in Arabidopsis thaliana reveals dynamic reprogramming of phytoalexin and phytoanticipin pathways. PLoS ONE 2016, 11, e0163572. [Google Scholar] [CrossRef] [Green Version]
- Van Peer, R.; Schippers, B. Lipopolysaccharides of plant-growth promoting Pseudomonas sp. strain WCS417r induce resistance in carnation to Fusarium wilt. Neth. J. Plant Pathol. 1992, 98, 129–139. [Google Scholar] [CrossRef]
- Reitz, M.; Rudolph, K.; Schroder, I.; Hoffmann-Hergarten, S.; Hallmann, J.; Sikora, R.A. Lipopolysaccharides of Rhizobium etli strain G12 act in potato roots as an inducing agent of systemic resistance to infection by the cyst nematode Globodera pallida. Appl. Environ. Microbiol. 2000, 66, 3515–3518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chávez-Herrera, E.; Hernández-Esquivel, A.A.; Castro-Mercado, E.; García-Pineda, E. Effect of Azospirillum brasilense Sp 245 lipopolysaccharides on wheat plant development. J. Plant Growth Regul. 2018, 37, 859–866. [Google Scholar] [CrossRef]
- Coats, S.R.; Pham, T.-T.T.; Bainbridge, B.W.; Reife, R.A.; Darveau, R.P. MD-2 mediates the ability of tetra-acylated and penta-acylated lipopolysaccharides to antagonize Escherichia coli lipopolysaccharide at the TLR4 signaling complex. J. Immunol. 2005, 175, 4490–4498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darveau, R.P.; Hancock, R.E.W. Procedure for isolation of bacterial lipopolysaccharides from both smooth and rough Pseudomonas aeruginosa and Salmonella typhimurium strains. J. Bacteriol. 1983, 155, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Pelegrini, P.B.; Franco, O.L. Plant γ-thionins: Novel insights on the mechanism of action of a multi-functional class of defense proteins. Int. J. Biochem. Cell Biol. 2005, 37, 2239–2253. [Google Scholar] [CrossRef]
- Kebede, A.; Kebede, M. In silico analysis of promoter region and regulatory elements of glucan endo-1,3-beta-glucosidase encoding genes in Solanum tuberosum: Cultivar DM 1-3 516 R44. J. Genet. Eng. Biotechnol. 2021, 19, 145. [Google Scholar] [CrossRef] [PubMed]
- Dunwell, J.M.; Gibbings, J.G.; Mahmood, T.; Saqlan Naqvi, S.M. Germin and germin-like proteins: Evolution, structure, and function. CRC Crit. Rev. Plant Sci. 2008, 27, 342–375. [Google Scholar] [CrossRef]
- Othman, T.A.; Bakar, N.A.; Abidin, R.A.Z.; Mahmood, M.; Saidi, N.B.; Shaharuddin, N.A. Potential of plant’s Bowman-Birk protease inhibitor in combating abiotic stresses: A Mini Review. Bioremediat. Sci. Technol. Res. 2015, 2, 25–33. [Google Scholar] [CrossRef]
- Tunnacliffe, A.; Wise, M.J. The continuing conundrum of the LEA proteins. Naturwissenschaften 2007, 94, 791–812. [Google Scholar] [CrossRef]
- Watanabe, K.; Praseuth, A.P.; Wang, C.C. A comprehensive and engaging overview of the type III family of polyketide synthases. Curr. Opin. Chem. Biol. 2007, 11, 279–286. [Google Scholar] [CrossRef]
- Wang, J.D.; Li, Z.; Liang, Y.; Zheng, J.; Gong, Z.; Zhou, G.; Xu, Y.; Li, X. Genome-wide identification and expression reveal the involvement of the FCS-like zinc finger (FLZ) gene family in Gossypium hirsutum at low temperature. PeerJ 2023, 11, e14690. [Google Scholar] [CrossRef]
- Huang, H.; Liu, B.; Liu, L.; Song, S. Jasmonate action in plant growth and development. J. Exp. Bot. 2017, 68, 1349–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, H.T.; To, H.T.M.; Lebrun, M.; Bellafiore, S.; Champion, A. Jasmonates—The master regulator of rice development, adaptation and defense. Plants 2019, 8, 339. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Li, Q.; Wang, Q.; Lu, D.; Zhang, H.; Wang, J.; Fu, R. Transcriptional profiling provides new insights into the role of nitric oxide in enhancing Ganoderma oregonense resistance to heat stress. Sci. Rep. 2017, 7, 15694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, H.; He, L.F. Crosstalk between melatonin and nitric oxide in plant development and stress responses. Physiol. Plant 2020, 170, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Chutoam, P.; Charoensawan, V.; Wongtrakoongate, P.; Kum-arth, A.; Buphamalai, P.; Tungpradabkul, S. RpoS and oxidative stress conditions regulate succinyl-CoA: 3-ketoacid-coenzyme A transferase (SCOT) expression in Burkholderia pseudomallei. Microbiol. Immunol. 2013, 57, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Field, B. Green magic: Regulation of the chloroplast stress response by (p)ppGpp in plants and algae. J. Exp. Bot. 2018, 69, 2797–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Z.H.; He, D.; Kohorn, B.D. Requirement for the induced expression of a cell wall associated receptor kinase for survival during the pathogen response. Plant J. 1998, 14, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Jez, J.M. Connecting primary and specialized metabolism: Amino acid conjugation of phytohormones by GRETCHEN HAGEN 3 (GH3) acyl acid amido synthetases. Curr. Opin. Plant Biol. 2022, 66, 102194. [Google Scholar] [CrossRef]
- Kaur, H.; Chowrasia, S.; Gaur, V.S.; Mondal, T.K. Allantoin: Emerging role in plant abiotic stress tolerance. Plant Mol. Biol. Report. 2016, 39, 648–661. [Google Scholar] [CrossRef]
- Mutuku, J.M.; Yoshida, S.; Shimizu, T.; Ichihashi, Y.; Wakatake, T.; Takahashi, A.; Seo, M.; Shirasu, K. The WRKY45-dependent signaling pathway is required for resistance against Striga hermonthica parasitism. Plant Physiol. 2015, 168, 1152–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perotti, M.F.; Arce, A.L.; Chan, R.L. The underground life of homeodomain-leucine zipper transcription factors. J. Exp. Bot. 2021, 72, 4005–4021. [Google Scholar] [CrossRef]
- Stotz, H.U.; Mueller, S.; Zoeller, M.; Mueller, M.J.; Berger, S. TGA transcription factors and jasmonate-independent COI1 signalling regulate specific plant responses to reactive oxylipins. J. Exp. Bot. 2013, 64, 963–975. [Google Scholar] [CrossRef]
- Domsalla, A.; Melzig, M.F. Occurrence and properties of proteases in plant latices. Planta Med. 2008, 74, 699–711. [Google Scholar] [CrossRef] [Green Version]
- Lin, K.; Zeng, M.; Williams, D.V.; Hu, W.; Shabala, S.; Zhou, M.; Cao, F. Integration of transcriptome and metabolome analyses reveals the mechanistic basis for cadmium accumulation in maize. iScience 2022, 25, 105484. [Google Scholar] [CrossRef] [PubMed]
- Beier, M.P.; Obara, M.; Taniai, A.; Sawa, Y.; Ishizawa, J.; Yoshida, H.; Tomita, N.; Yamanaka, T.; Ishizuka, Y.; Kudo, S.; et al. Lack of ACTPK1, an STY kinase, enhances ammonium uptake and use, and promotes growth of rice seedlings under sufficient external ammonium. Plant J. 2018, 93, 992–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onyemaobi, O.; Sangma, H.; Garg, G.; Wallace, X.; Kleven, S.; Dolferus, R. Transcriptome profiling of the chilling response in wheat spikes: I, acclimation response to long-term chilling treatment. Curr. Plant Biol. 2022, 31, 100255. [Google Scholar] [CrossRef]
- Nakashima, K.; Takasaki, H.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. NAC transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta-Gene Regul. Mech. 2012, 1819, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Feys, B.J.; Parker, J.E. Interplay of signaling pathways in plant disease resistance. Trends Genet. 2000, 16, 449–455. [Google Scholar] [CrossRef]
- Jia, J.; Fu, J.; Zheng, J.; Zhou, X.; Huai, J.; Wang, J.; Wang, M.; Zhang, Y.; Chen, X.; Zhang, J.; et al. Annotation and expression profile analysis of 2073 full-length cDNAs from stress-induced maize (Zea mays L.) seedlings. Plant J. 2006, 48, 710–727. [Google Scholar] [CrossRef]
- Al-Momany, B.; Abu-Romman, S. Cloning and molecular characterization of a flavin-dependent oxidoreductase gene from barley. J. Appl. Genet. 2014, 55, 457–468. [Google Scholar] [CrossRef]
- Collins, C.; Keane, T.M.; Turner, D.J.; O’Keeffe, G.; Fitzpatrick, D.A.; Doyle, S. Genomic and proteomic dissection of the ubiquitous plant pathogen, Armillaria mellea: Toward a new infection model system. J. Proteome Res. 2013, 12, 2552–2570. [Google Scholar] [CrossRef]
- James, D.; Tarafdar, A.; Biswas, K.; Sathyavathi, T.C.; Padaria, J.C.; Kumar, P.A. Development and characterization of a high temperature stress responsive subtractive cDNA library in Pearl Millet Pennisetum glaucum (L.) R. Br. Indian J. Exp. Biol. 2015, 53, 543–550. [Google Scholar]
- Li, C.; Xu, M.; Cai, X.; Han, Z.; Si, J.; Chen, D. Jasmonate signaling pathway modulates plant defense, growth, and their trade-offs. Int. J. Mol. Sci. 2022, 23, 3945. [Google Scholar] [CrossRef]
- Miyamoto, K.; Shimizu, T.; Okada, K. Transcriptional regulation of the biosynthesis of phytoalexin: A lesson from specialized metabolites in rice. Plant Biotechnol. 2014, 31, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Austin, M.B.; Noel, J.P. The chalcone synthase superfamily of type III polyketide synthases. Nat. Prod. Rep. 2003, 20, 79–110. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.T.H.; Linthorst, H.J.M.; Verpoorte, R. Chalcone synthase and its functions in plant resistance. Phytochem. Rev. 2011, 10, 397–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, A.; Pühler, A.; Niehaus, K. The lipopolysaccharides of the phytopathogen Xanthomonas campestris pv. campestris induce an oxidative burst reaction in cell cultures of Nicotiana tabacum. Planta 2001, 213, 214–222. [Google Scholar] [CrossRef]
- Silipo, A.; Molinaro, A.; Sturiale, L.; Dow, J.M.; Erbs, G.; Lanzetta, R.; Newman, M.A.; Parrilli, M. The elicitation of plant innate immunity by lipooligosaccharide of Xanthomonas campestris. J. Biol. Chem. 2005, 280, 33660–33668. [Google Scholar] [CrossRef] [Green Version]
- Madala, N.E.; Molinaro, A.; Dubery, I.A. Distinct carbohydrate and lipid-based molecular patterns within lipopolysaccharides from Burkholderia cepacia contribute to defense-associated differential gene expression in Arabidopsis thaliana. Innate Immun. 2012, 18, 140–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigida, E.N.; Fedonenko, Y.P.; Shashkov, A.S.; Toukach, P.V.; Shelud’ko, A.V.; Zdorovenko, E.L.; Knirel, Y.A.; Konnova, S.A. Structural studies of O-specific polysaccharide(s) and biological activity toward plants of the lipopolysaccharide from Azospirillum brasilense SR8. Int. J. Biol. Macromol. 2019, 126, 246–253. [Google Scholar] [CrossRef]
- Oblak, A.; Jerala, R. The molecular mechanism of species-specific recognition of lipopolysaccharides by the MD-2/TLR4 receptor complex. Mol. Immunol. 2015, 63, 134–142. [Google Scholar] [CrossRef]
- Steimle, A.; Autenrieth, I.B.; Frick, J.S. Structure and function: Lipid A modifications in commensals and pathogens. Int. J. Med. Microbiol. 2016, 306, 290–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirey, K.A.; Lai, W.; Scott, A.J.; Lipsky, M.; Mistry, P.; Pletneva, L.M.; Karp, C.L.; McAlees, J.; Gioannini, T.L.; Weiss, J.; et al. The TLR4 antagonist Eritoran protects mice from lethal influenza infection. Nature 2013, 497, 498–502. [Google Scholar] [CrossRef] [Green Version]
- Rossignol, D.P.; Lynn, M. TLR4 antagonists for endotoxemia and beyond. Curr. Opin. Investig. Drugs 2005, 6, 496–502. [Google Scholar] [PubMed]
- Zeidler, D.; Zähringer, U.; Gerber, I.; Dubery, I.; Hartung, T.; Bors, W.; Hutzler, P.; Durner, J. Innate immunity in Arabidopsis thaliana: Lipopolysaccharides activate nitric oxide synthase (NOS) and induce defense genes. Proc. Natl. Acad. Sci. USA 2004, 101, 15811–15816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishina, T.E.; Zeier, J. Pathogen-associated molecular pattern recognition rather than development of tissue necrosis contributes to bacterial induction of systemic acquired resistance in Arabidopsis. Plant J. 2007, 50, 500–513. [Google Scholar] [CrossRef]
- Song, M.C.; Beom, R.K.; Song, H.H.; Anderson, A.J.; Park, J.Y.; Lee, Y.H.; Baik, H.C.; Yang, K.Y.; Ryu, C.M.; Kim, Y.C. 2R,3R-butanediol, a bacterial volatile produced by Pseudomonas chlororaphis O6, is involved in induction of systemic tolerance to drought in Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2008, 21, 1067–1075. [Google Scholar] [CrossRef] [Green Version]
- Silipo, A.; Sturiale, L.; Garozzo, D.; Erbs, G.; Jensen, T.T.; Lanzetta, R.; Dow, J.M.; Parrilli, M.; Newman, M.A.; Molinaro, A. The acylation and phosphorylation pattern of lipid A from Xanthomonas campestris strongly influence its ability to trigger the innate immune response in Arabidopsis. ChemBioChem 2008, 9, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Madala, N.E.; Leone, M.R.; Molinaro, A.; Dubery, I.A. Deciphering the structural and biological properties of the lipid A moiety of lipopolysaccharides from Burkholderia cepacia strain ASP B 2D, in Arabidopsis thaliana. Glycobiology 2011, 21, 184–194. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, K.H.; Daniel, T.; Aurélien, D.; El-Maarouf-Bouteau, H.; Rafik, E.; Arbelet-Bonnin, D.; Biligui, B.; Florence, V.; Mustapha, E.M.; François, B. Deciphering the dual effect of lipopolysaccharides from plant pathogenic Pectobacterium. Plant Signal. Behav. 2015, 10, e1000160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melotto, M.; Underwood, W.; Koczan, J.; Nomura, K.; He, S.Y. Plant stomata function in innate immunity against bacterial invasion. Cell 2006, 126, 969–980. [Google Scholar] [CrossRef] [Green Version]
- Iizasa, S.; Iizasa, E.; Watanabe, K.; Nagano, Y. Transcriptome analysis reveals key roles of AtLBR-2 in LPS-induced defense responses in plants. BMC Genom. 2017, 18, 995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shilina, J.V.; Gushcha, M.I.; Molozhava, O.S.; Litvinov, S.V.; Dmitriev, A.P. Induction of Arabidopsis thaliana resistance to pathogenic bacteria by lipopolysaccharide and salicylic acid. Cytol. Genet. 2018, 52, 169–173. [Google Scholar] [CrossRef]
- Tinte, M.M.; Steenkamp, P.A.; Piater, L.A.; Dubery, I.A. Lipopolysaccharide perception in Arabidopsis thaliana: Diverse LPS chemotypes from Burkholderia cepacia, Pseudomonas syringae and Xanthomonas campestris trigger differential defence-related perturbations in the metabolome. Plant Physiol. Biochem. 2020, 156, 267–277. [Google Scholar] [CrossRef]
- Hussan, R.H.; Dubery, I.A.; Piater, L.A. Identification of mamp-responsive plasma membrane-associated proteins in Arabidopsis thaliana following challenge with different LPS chemotypes from Xanthomonas campestris. Pathogens 2020, 9, 787. [Google Scholar] [CrossRef] [PubMed]
- Girija, A.M.; Kinathi, B.K.; Madhavi, M.B.; Ramesh, P.; Vungarala, S.; Patel, H.K.; Sonti, R.V. Rice leaf transcriptional profiling suggests a functional interplay between Xanthomonas oryzae pv. oryzae lipopolysaccharide and extracellular polysaccharide in modulation of defense responses during infection. Mol. Plant-Microbe Interact. 2017, 30, 16–27. [Google Scholar] [CrossRef] [Green Version]
- Desaki, Y.; Kouzai, Y.; Ninomiya, Y.; Iwase, R.; Shimizu, Y.; Seko, K.; Molinaro, A.; Minami, E.; Shibuya, N.; Kaku, H.; et al. OsCERK1 plays a crucial role in the lipopolysaccharide-induced immune response of rice. New. Phytol. 2018, 217, 1042–1049. [Google Scholar] [CrossRef] [Green Version]
- Vallejo-Ochoa, J.; López-Marmolejo, M.; Hernández-Esquivel, A.A.; Méndez-Gómez, M.; Suárez-Soria, L.N.; Castro-Mercado, E.; García-Pineda, E. Early plant growth and biochemical responses induced by Azospirillum brasilense Sp 245 lipopolysaccharides in wheat (Triticum aestivum L.) seedlings are attenuated by procyanidin B2. Protoplasma 2018, 255, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Coventry, H.S.; Dubery, I.A. Lipopolysaccharides from Burkholderia cepacia contribute to an enhanced defensive capacity and the induction of pathogenesis-related proteins in Nicotianae tabacum. Physiol. Mol. Plant Pathol. 2001, 58, 149–158. [Google Scholar] [CrossRef]
- Gerber, I.B.; Zeidler, D.; Durner, J.; Dubery, I.A. Early perception responses of Nicotiana tabacum cells in response to lipopolysaccharides from Burkholderia cepacia. Planta 2004, 218, 647–657. [Google Scholar] [CrossRef]
- Braun, S.G.; Meyer, A.; Holst, O.; Pühler, A.; Niehaus, K. Characterization of the Xanthomonas campestris pv. campestris lipopolysaccharide substructures essential for elicitation of an oxidative burst in tobacco cells. Mol. Plant-Microbe Interact. 2005, 18, 674–681. [Google Scholar] [CrossRef] [Green Version]
- Sanabria, N.M.; Dubery, I.A. Differential display profiling of the Nicotiana response to LPS reveals elements of plant basal resistance. Biochem. Biophys. Res. Commun. 2006, 344, 1001–1007. [Google Scholar] [CrossRef]
- Sanabria, N.M.; van Heerden, H.; Dubery, I.A. Molecular characterisation and regulation of a Nicotiana tabacum S-domain receptor-like kinase gene induced during an early rapid response to lipopolysaccharides. Gene 2012, 501, 39–48. [Google Scholar] [CrossRef]
- Mhlongo, M.I.; Piater, L.A.; Madala, N.E.; Steenkamp, P.A.; Dubery, I.A. Phenylpropanoid defences in Nicotiana tabacum cells: Overlapping metabolomes indicate common aspects to priming responses induced by lipopolysaccharides, chitosan and flagellin-22. PLoS ONE 2016, 11, e0151350. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.A.; Von Roepenack-Lahaye, E.; Parr, A.; Daniels, M.J.; Dow, J.M. Prior exposure to lipopolysaccharide potentiates expression of plant defenses in response to bacteria. Plant J. 2002, 29, 487–495. [Google Scholar] [CrossRef]
- Mareya, C.R.; Tugizimana, F.; Di Lorenzo, F.; Silipo, A.; Piater, L.A.; Molinaro, A.; Dubery, I.A. Adaptive defence-related changes in the metabolome of Sorghum bicolor cells in response to lipopolysaccharides of the pathogen Burkholderia andropogonis. Sci. Rep. 2020, 10, 7626. [Google Scholar] [CrossRef] [PubMed]
- Rapicavoli, J.N.; Blanco-Ulate, B.; Muszyński, A.; Figueroa-Balderas, R.; Morales-Cruz, A.; Azadi, P.; Dobruchowska, J.M.; Castro, C.; Cantu, D.; Roper, M.C. Lipopolysaccharide O-antigen delays plant innate immune recognition of Xylella fastidiosa. Nat. Commun. 2018, 9, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nürnberger, T.S.D. Signal transmission in the plant immune response. Trends Plant Sci. 2018, 6, 372–379. [Google Scholar] [CrossRef]
- Bennett, R.N.; Wallsgrove, R.M. Secondary metabolites in plant defence mechanisms. New. Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef]
- Lei, K.J.; Zhang, L.; Du, X.Y.; An, Y.; Chang, G.H.; An, G.Y. A chalcone synthase controls the verticillium disease resistance response in both Arabidopsis thaliana and cotton. Eur. J. Plant Pathol. 2018, 152, 769–781. [Google Scholar] [CrossRef]
- Newman, M.A.; Dow, M.J.; Molinaro, A.; Parrilli, M. Priming, induction and modulation of plant defence responses by bacterial lipopolysaccharides. J. Endotoxin Res. 2007, 13, 69–84. [Google Scholar] [CrossRef] [Green Version]
- Van Loon, L.C.; Bakker, P.A.H.M.; Van Der Heijdt, W.H.W.; Wendehenne, D.; Pugin, A. Early responses of tobacco suspension cells to rhizobacterial elicitors of induced systemic resistance. Mol. Plant-Microbe Interact. 2008, 21, 1609–1621. [Google Scholar] [CrossRef] [Green Version]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.F.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.G.; et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 422, 442–446. [Google Scholar] [CrossRef]
- Takeda, S.; Gapper, C.; Kaya, H.; Bell, E.; Kuchitsu, K.; Dolan, L. Local positive feedback regulation determines cell shape in root hair cells. Science 2008, 319, 1241–1244. [Google Scholar] [CrossRef]
- Wongkantrakorn, N.; Sunohara, Y.; Matsumoto, H. Mechanism of growth amelioration of NaCl-stressed rice (Oryza sativa L.) by δ-aminolevulinic acid. J. Pestic. Sci. 2009, 34, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Su, P.; Tan, X.; Li, C.; Zhang, D.; Cheng, J.; Zhang, S.; Zhou, X.; Yan, Q.; Peng, J.; Zhang, Z.; et al. Photosynthetic bacterium Rhodopseudomonas palustris GJ-22 induces systemic resistance against viruses. Microb. Biotechnol. 2017, 10, 612–624. [Google Scholar] [CrossRef] [PubMed]
- De Geyter, N.; Gholami, A.; Goormachtig, S.; Goossens, A. Transcriptional machineries in jasmonate-elicited plant secondary metabolism. Trends Plant Sci. 2012, 17, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Memelink, J. Jasmonate-responsive transcription factors regulating plant secondary metabolism. Biotechnol. Adv. 2016, 34, 441–449. [Google Scholar] [CrossRef]
- Nabi, N.; Singh, S.; Saffeullah, P. Responses of in vitro cell cultures to elicitation: Regulatory role of jasmonic acid and methyl jasmonate: A review. In Vitro Cell. Dev. Biol.-Plant 2021, 57, 341–355. [Google Scholar] [CrossRef]
- Ho, T.; Murthy, H.N.; Park, S. Methyl jasmonate induced oxidative stress and accumulation of secondary metabolites in plant cell and organ cultures. Int. J. Mol. Sci. 2020, 21, 716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Jones, A.D.; Howe, G.A. Constitutive activation of the jasmonate signaling pathway enhances the production of secondary metabolites in tomato. FEBS Lett. 2006, 580, 2540–2546. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LPS | Dead Cells | |||||
|---|---|---|---|---|---|---|
| Gene ID | logFC | Gene Description | Stress Response | JA * | Reference | |
| Os06g0514100 | 6.17 | 7.82 | Thionin; antimicrobial peptide | X | [31] | |
| Os10g0177300 | 5.38 | 5.28 | Chalcone and stilbene synthases | X | [36] | |
| Os03g0183500 | 4.54 | 3.88 | FCS-like zinc finger protein | X | X | [37,38] |
| Os11g0603000 | 4.34 | 4.17 | Basic helix-loop-helix dimerization region bHLH domain containing protein | X | X | [38,39] |
| Os02g0137700 | 4.25 | 2.53 | NAD(P)-binding domain containing protein | X | [40] | |
| Os01g0946600 | 3.65 | 2.74 | Glucan endo-1,3-beta-glucosidase | X | [32] | |
| Os10g0392400 | 3.55 | 2.01 | Tify domain containing protein | X | X | [38,39] |
| Os07g0153000 | 3.51 | 3.48 | Jasmonate ZIM-domain (JAZ) family | X | X | [38,39] |
| Os06g0513781 | 3.41 | 3.09 | Thionin; antimicrobial peptide | X | [31] | |
| Os03g0741100 | 3.29 | 2.67 | Basic helix-loop-helix transcription factor, drought tolerance | X | X | [38,39] |
| Os11g0684000 | 3.25 | 1.76 | Myb transcription factor | X | X | [38,39] |
| Os01g0946700 | 3.17 | 1.93 | Glucan endo-1,3-beta-glucosidase | X | [32] | |
| Os03g0142600 | 3.10 | 3.52 | Myb transcription factor | X | X | [38,39] |
| Os03g0180900 | 3.09 | 2.22 | Jasmonate ZIM-domain containing protein | X | X | [38,39] |
| Os10g0118200 | 3.08 | 2.94 | Acetylserotonin O-methyltransferase | X | [41] | |
| Os01g0124700 | 3.04 | 1.93 | Bowman–Birk type protease inhibitor | X | [34] | |
| Os03g0181100 | 2.92 | 2.78 | Tify domain containing protein | X | X | [38,39] |
| Os12g0478400 | 2.90 | 2.46 | EGF-type aspartate/asparagine hydroxylation site domain containing protein | |||
| Os01g0225500 | 2.85 | 3.47 | 3-methyl-2-oxobutanoate hydroxymethyltransferase | X | [42] | |
| Os01g0124650 | 2.91 | 1.84 | Bowman–Birk type protease inhibitor | X | [34] | |
| Os05g0161500 | 2.73 | 2.75 | Calcium-activated (p)ppGpp synthetase | X | [43] | |
| Os02g0808000 | 2.57 | 1.98 | Wall-associated receptor kinase 2 | X | [44] | |
| Os05g0586200 | 2.53 | 1.35 | Gretchen hagen 3 (GH3) | X | [45] | |
| Os08g0190100 | 2.38 | 2.12 | Germin-like protein | X | [33] | |
| Os12g0503000 | 2.08 | 2.08 | Allantoin transporter | X | [46] | |
| Os09g0439200 | 1.97 | 1.24 | Jasmonate ZIM-domain protein | X | X | [38,39] |
| Os01g0108600 | 1.76 | 1.51 | Basic helix-loop-helix dimerization region bHLH domain containing protein | X | X | [38,39] |
| Os02g0181300 | 1.75 | 1.56 | WRKY transcription factor | X | X | [38,47] |
| Os03g0402800 | 1.71 | 0.89 | TIFY family protein, JASMONATE-ZIM domain (JAZ) protein | X | X | [38,39] |
| Os03g0198600 | 1.71 | 1.56 | Homeodomain-leucine zipper transcription factor; regulation of panicle exertion | X | X | [38,48] |
| Os12g0138800 | 1.59 | 1.92 | Six-bladed beta-propeller, TolB-like domain containing protein | X | X | [38,49] |
| Os12g0548401 | 1.58 | 1.64 | Proteinase inhibitor | X | [50] | |
| Os06g0231600 | 1.53 | 1.13 | RING-H2 finger protein ATL1Q | X | [51] | |
| Os07g0475900 | 1.50 | 1.52 | ACT domain containing protein kinase | X | [52] | |
| Os12g0267200 | 1.46 | 1.32 | Cyclopropane-fatty-acyl-phospholipid synthase | X | [53] | |
| Os09g0401300 | 1.45 | 0.57 | JASMONATE ZIM-domain (JAZ) | X | X | [38,39] |
| Os07g0138200 | 1.37 | 1.51 | NAC transcription factor; ABA-induced leaf senescence and tillering | X | [54] | |
| Os01g0314800 | 1.32 | 1.11 | Late embryogenesis abundant protein 3 | X | [35] | |
| Os11g0644700 | 1.29 | 1.61 | Plant disease-resistance response protein | X | [55] | |
| Os06g0112100 | 1.17 | 1.03 | Nucleoside phosphorylase | |||
| Os05g0126800 | 1.15 | 1.02 | Mss4-like domain containing protein | X | [56] | |
| Os01g0370000 | 1.14 | 1.33 | NADH:flavin oxidoreductase/NADH oxidase | X | [57] | |
| Os05g0542200 | 1.06 | 0.69 | Alpha/beta hydrolase fold-1 domain containing protein | X | [58] | |
| Os09g0248900 | 1.04 | 1.30 | Myb/SANT-like domain containing protein | X | X | [38,39] |
| Os07g0633400 | 0.90 | 1.51 | IQ calmodulin-binding region domain containing protein | X | [59] | |
| Os12g0626400 | 0.82 | 0.84 | Phytoene synthase 1 | |||
| Os04g0517100 | 0.74 | 0.61 | SG2-type MYB transcription factor; cold tolerance; resistance to fungal and bacterial pathogens | X | X | [38,39] |
| Plant | Bacteria | Concentration (μg/mL) | Response to LPS | Reference |
|---|---|---|---|---|
| Arabidopsis thaliana | Burkholderia cepacia | 100 | Activation of nitric oxide synthase (NOS) and induction of defense genes | [72] |
| Arabidopsis thaliana | Xanthomonas campestris | 50 | Elicitation of innate immunity | [65] |
| Arabidopsis thaliana | Pseudomonas syringae E. coli | 100 | Induction of systemic acquired resistance (SAR) | [73] |
| Arabidopsis thaliana | Pseudomonas chlororaphis O6 | 100 | Stomatal closure and induction of systemic tolerance to drought | [74] |
| Arabidopsis thaliana | Xanthomonas campestris | 50 (lipid A) | Induction of pathogenesis-related 1 (PR1) gene | [75] |
| Arabidopsis thaliana | Burkholderia cepacia | 20 (lipid A). | Induction of defense-related genes | [76] |
| Arabidopsis thaliana | Burkholderia cepacia | 100 (LPS) 20 (lipid A) | Induction of defense-related metabolites synthesis | [66] |
| Arabidopsis thaliana | Pectobacterium atrosepticum Pectobacterium carotovorum subsp. carotovorum | 10 to 100 | Induction of defense response | [77] |
| Arabidopsis thaliana | Burkholderia cepacia | 80 | Induction of phytoalexin synthesis | [25] |
| Arabidopsis thaliana | Pseudomonas aeruginosa | 100 | Stomatal closure | [78] |
| Arabidopsis thaliana | E. coli Pseudomonas aeruginosa | LPS (E.) 10 LPS (P.) 100 | ROS generation | [23] |
| Arabidopsis thaliana | Pseudomonas aeruginosa | 100 | Induction of defense response | [79] |
| Arabidopsis thaliana | Pseudomonas aeruginosa | LPS 25 Lipid A 10 | ROS generation Inhibition of seedling growth | [24] |
| Arabidopsis thaliana | Pseudomonas aeruginosa | 100 | Enhanced resistance to pathogen Activation of SA signaling pathway | [80] |
| Arabidopsis thaliana | Burkholderia cepacian Pseudomonas syringae Xanthomonas campestris | 100 | Induction of defense-related metabolites synthesis | [81] |
| Arabidopsis thaliana | Xanthomonas campestris | 100 | Induction of defense response | [82] |
| Oryza sativa | Xanthomonas oryzae | 50 | ROS generation | [22] |
| Oryza sativa | Xanthomonas oryzae | 100 | Induction of defense response | [83] |
| Oryza sativa | Pseudomonas aeruginosa, E. coli | 50 | Induction of immune response | [84] |
| Solanum tuberosum | Rhizobium etli strain G12 | 100 to 1000 | Resistance to nematode infection | [27] |
| Triticum aestivum | Azospirillum brasilense Sp245 | 100 | Growth promotion, ROS generation | [85] |
| Triticum aestivum | Azospirillum brasilense Sp245 | 2–5 | Promotion of plant development (plant aging, spike formation, and size) | [28] |
| Triticum aestivum | Azospirillum brasilense SR8 | 1000 | Root hair deformations | [67] |
| Nicotiana tabacum | Burkholderia cepacia | 100 | Induction of defense response | [86] |
| Nicotiana tabacum | Burkholderia cepacia | 100 | ROS generation | [87] |
| Nicotiana tabacum | Xanthomonas campestris | 10 (5 to 500) | ROS generation | [64] |
| Nicotiana tabacum | Xanthomonas campestris pv. campestris | 20 | ROS generation | [88] |
| Nicotiana tabacum | Burkholderia cepacia | 100 | Induction of innate immunity ROS generation | [89] |
| Nicotiana tabacum | Burkholderia cepacia | 100 | Induction of defense response S-domain receptor-like kinase (RLK) | [90] |
| Nicotiana tabacum | Burkholderia cepacia | 100 | Induction of phenylpropanoid biosynthesis | [91] |
| Capsicum annuum | Xanthomonas axonopodis pv. Vesicatoria X. campestris pv. campestris | 50 | Accumulation of salicylic acid (SA), coumaroyl-tyramine (CT), and feruloyl-tyramine (FT) | [92] |
| Sorghum bicolor | Burkholderia andropogonis | 100 | Induction of secondary metabolites synthesis | [93] |
| Vitis vinifera | Xylella fastidiosa | 50 | Induction of defense response against pathogens | [94] |
| PNSB | Concentration | |||
| Brassica rapa var. perviridis | Rhodobacter sphaeroides NBRC 12203T | 10 pg/mL | Growth promotion | [18] |
| Oryza sativa | R. sphaeroides NBRC 12203T | 5 ng/mL | Promotion of root development | [19] |
| Oryza sativa | R. sphaeroides NBRC 12203T | 10 pg/mL | Stimulation of JA signaling pathway Induction of secondary metabolites synthesis | Present study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iwai, R.; Uchida, S.; Yamaguchi, S.; Nagata, D.; Koga, A.; Hayashi, S.; Yamamoto, S.; Miyasaka, H. Effects of LPS from Rhodobacter sphaeroides, a Purple Non-Sulfur Bacterium (PNSB), on the Gene Expression of Rice Root. Microorganisms 2023, 11, 1676. https://doi.org/10.3390/microorganisms11071676
Iwai R, Uchida S, Yamaguchi S, Nagata D, Koga A, Hayashi S, Yamamoto S, Miyasaka H. Effects of LPS from Rhodobacter sphaeroides, a Purple Non-Sulfur Bacterium (PNSB), on the Gene Expression of Rice Root. Microorganisms. 2023; 11(7):1676. https://doi.org/10.3390/microorganisms11071676
Chicago/Turabian StyleIwai, Ranko, Shunta Uchida, Sayaka Yamaguchi, Daiki Nagata, Aoi Koga, Shuhei Hayashi, Shinjiro Yamamoto, and Hitoshi Miyasaka. 2023. "Effects of LPS from Rhodobacter sphaeroides, a Purple Non-Sulfur Bacterium (PNSB), on the Gene Expression of Rice Root" Microorganisms 11, no. 7: 1676. https://doi.org/10.3390/microorganisms11071676
APA StyleIwai, R., Uchida, S., Yamaguchi, S., Nagata, D., Koga, A., Hayashi, S., Yamamoto, S., & Miyasaka, H. (2023). Effects of LPS from Rhodobacter sphaeroides, a Purple Non-Sulfur Bacterium (PNSB), on the Gene Expression of Rice Root. Microorganisms, 11(7), 1676. https://doi.org/10.3390/microorganisms11071676