
Engineering the Metabolic Landscape of Microorganisms for Lignocellulosic Conversion

 ,
, 
Abstract
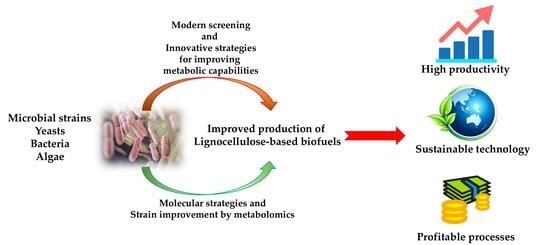
:
1. Introduction
2. Traditional Techniques for Converting Lignocellulosic Residues into High-Value Products
3. Enhancing Biomass-Based Products through Metabolic Engineering
3.1. Bacteria
3.2. Yeasts
3.3. Microalgae
4. Strategies for Advancing and Innovating Metabolic Engineering in Microorganisms for Applications
5. Combined Molecular and Evolutionary Strategies
6. Strain Improvement by Characterization of Integral Control of Metabolism
7. Non-Conventional Wastes and Auxiliary Pathways
8. A Note on Safety to Achieve the Transition from the Laboratory into the Industry
9. Prospects of Engineered Microorganisms to Produce Biomass-Based Added-Value Compounds
10. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antar, M.; Lyu, D.; Nazari, M.; Shah, A.; Zhou, X.; Smith, D.L. Biomass for a Sustainable Bioeconomy: An Overview of World Biomass Production and Utilization. Renew. Sustain. Energy Rev. 2021, 139, 110691. [Google Scholar] [CrossRef]
- Peña-Castro, J.M.; del Moral, S.; Núñez-López, L.; Barrera-Figueroa, B.E.; Amaya-Delgado, L. Biotechnological Strategies to Improve Plant Biomass Quality for Bioethanol Production. BioMed Res. Int. 2017, 2017, e7824076. [Google Scholar] [CrossRef] [PubMed]
- Adegboye, M.F.; Ojuederie, O.B.; Talia, P.M.; Babalola, O.O. Bioprospecting of Microbial Strains for Biofuel Production: Metabolic Engineering, Applications, and Challenges. Biotechnol. Biofuels 2021, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Nielsen, J. Recent Trends in Metabolic Engineering of Microbial Chemical Factories. Curr. Opin. Biotechnol. 2019, 60, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Gibson, B.; Dahabieh, M.; Krogerus, K.; Jouhten, P.; Magalhães, F.; Pereira, R.; Siewers, V.; Vidgren, V. Adaptive Laboratory Evolution of Ale and Lager Yeasts for Improved Brewing Efficiency and Beer Quality. Annu. Rev. Food Sci. Technol. 2020, 11, 23–44. [Google Scholar] [CrossRef]
- Vázquez-Núñez, E.; Peña-Castro, J.M.; Fernández-Luqueño, F.; Cejudo, E.; de la Rosa-Alvarez, M.G.; García-castañeda, M.C. A Review on Genetically Modified Plants Designed to Phytoremediate Polluted Soils: Biochemical Responses and International Regulation. Pedosphere 2018, 28, 697–712. [Google Scholar] [CrossRef]
- Joshi, A.; Verma, K.K.; D Rajput, V.; Minkina, T.; Arora, J. Recent Advances in Metabolic Engineering of Microorganisms for Advancing Lignocellulose-Derived Biofuels. Bioengineered 2022, 13, 8135–8163. [Google Scholar] [CrossRef]
- Sartaj, K.; Patel, A.; Matsakas, L.; Prasad, R. Unravelling Metagenomics Approach for Microbial Biofuel Production. Genes 2022, 13, 1942. [Google Scholar] [CrossRef]
- Miller, D.C.; Athavale, S.V.; Arnold, F.H. Combining Chemistry and Protein Engineering for New-to-Nature Biocatalysis. Nat. Synth. 2022, 1, 18–23. [Google Scholar] [CrossRef]
- Galbe, M.; Wallberg, O. Pretreatment for Biorefineries: A Review of Common Methods for Efficient Utilisation of Lignocellulosic Materials. Biotechnol. Biofuels 2019, 12, 294. [Google Scholar] [CrossRef]
- Kumar, B.; Bhardwaj, N.; Agrawal, K.; Chaturvedi, V.; Verma, P. Current Perspective on Pretreatment Technologies Using Lignocellulosic Biomass: An Emerging Biorefinery Concept. Fuel Process. Technol. 2020, 199, 106244. [Google Scholar] [CrossRef]
- Martins, M.; Ávila, P.F.; da Silva, M.F.; de Melo, A.H.F.; Lopes, A.M.M.; Goldbeck, R. Lignocellulose Bioconversion through Chemical Methods, Platform Chemicals, and New Chemicals. In Lignocellulose Bioconversion Through White Biotechnology; Wiley: Hoboken, NJ, USA, 2022; pp. 221–247. [Google Scholar]
- Zhang, F.; Xu, L.; Xu, F.; Jiang, L. Different Acid Pretreatments at Room Temperature Boost Selective Saccharification of Lignocellulose via Fast Pyrolysis. Cellulose 2021, 28, 81–90. [Google Scholar] [CrossRef]
- Mankar, A.R.; Pandey, A.; Modak, A.; Pant, K. Pretreatment of Lignocellulosic Biomass: A Review on Recent Advances. Bioresour. Technol. 2021, 334, 125235. [Google Scholar] [CrossRef]
- Dharmaraja, J.; Shobana, S.; Arvindnarayan, S.; Francis, R.R.; Jeyakumar, R.B.; Saratale, R.; Veeramuthu, A.K.; Bhatia, S.K.; Kumar, V.; Kumar, G. Lignocellulosic Biomass Conversion via Greener Pretreatment Methods towards Biorefinery Applications. Bioresour. Technol. 2022, 369, 128328. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-H.; Nižetić, S.; Sirohi, R.; Huang, Z.; Luque, R.; Papadopoulos, A.M.; Sakthivel, R.; Nguyen, X.P.; Hoang, A.T. Liquid Hot Water as Sustainable Biomass Pretreatment Technique for Bioenergy Production: A Review. Bioresour. Technol. 2022, 344, 126207. [Google Scholar] [CrossRef] [PubMed]
- Wijeyekoon, S.L.; Vaidya, A.A. Woody Biomass as a Potential Feedstock for Fermentative Gaseous Biofuel Production. World J. Microbiol. Biotechnol. 2021, 37, 134. [Google Scholar] [CrossRef]
- Rodionova, M.V.; Bozieva, A.M.; Zharmukhamedov, S.K.; Leong, Y.K.; Lan, J.C.-W.; Veziroglu, A.; Veziroglu, T.N.; Tomo, T.; Chang, J.-S.; Allakhverdiev, S.I. A Comprehensive Review on Lignocellulosic Biomass Biorefinery for Sustainable Biofuel Production. Int. J. Hydrogen Energy 2022, 47, 1481–1498. [Google Scholar] [CrossRef]
- Rosales-Calderon, O.; Arantes, V. A Review on Commercial-Scale High-Value Products That Can Be Produced alongside Cellulosic Ethanol. Biotechnol. Biofuels 2019, 12, 1–58. [Google Scholar] [CrossRef]
- Fels, U.; Gevaert, K.; Van Damme, P. Bacterial Genetic Engineering by Means of Recombineering for Reverse Genetics. Front. Microbiol. 2020, 11, 548410. [Google Scholar] [CrossRef]
- He, M.X.; Wu, B.; Qin, H.; Ruan, Z.Y.; Tan, F.R.; Wang, J.L.; Shui, Z.X.; Dai, L.C.; Zhu, Q.L.; Pan, K. Zymomonas mobilis: A Novel Platform for Future Biorefineries. Biotechnol. Biofuels 2014, 7, 101. [Google Scholar] [CrossRef]
- Mendonça, A.A.; da Silva, P.K.N.; Calazans, T.L.S.; de Souza, R.B.; Elsztein, C.; de Morais Junior, M.A. Gene Regulation of the Lactobacillus Vini in Response to Industrial Stress in the Fuel Ethanol Production. Microbiol. Res. 2020, 236, 126450. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.R.; Klein-Marcuschamer, D.; Stephanopoulos, G. Selection and Optimization of Microbial Hosts for Biofuels Production. Metab. Eng. 2008, 10, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Connor, M.R.; Cann, A.F.; Liao, J.C. 3-Methyl-1-Butanol Production in Escherichia coli: Random Mutagenesis and Two-Phase Fermentation. Appl. Microbiol. Biotechnol. 2010, 86, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Nino, A.; Luna, C.; Luna, D.; Marcos, A.T.; Canovas, D.; Mellado, E. Selection and Characterization of Biofuel-Producing Environmental Bacteria Isolated from Vegetable Oil-Rich Wastes. PLoS ONE 2014, 9, e104063. [Google Scholar] [CrossRef]
- Tahar, I.B.; Fickers, P. Metabolic Engineering of Microorganisms for Urban Waste Valorization. Case Stud. Chem. Environ. Eng. 2021, 4, 100148. [Google Scholar] [CrossRef]
- Majidian, P.; Tabatabaei, M.; Zeinolabedini, M.; Naghshbandi, M.P.; Chisti, Y. Metabolic Engineering of Microorganisms for Biofuel Production. Renew. Sustain. Energy Rev. 2018, 82, 3863–3885. [Google Scholar] [CrossRef]
- Lu, H.; Yadav, V.; Zhong, M.; Bilal, M.; Taherzadeh, M.J.; Iqbal, H.M. Bioengineered Microbial Platforms for Biomass-Derived Biofuel Production–A Review. Chemosphere 2022, 288, 132528. [Google Scholar] [CrossRef]
- Wang, H.; Cao, S.; Wang, W.T.; Wang, K.T.; Jia, X. Very High Gravity Ethanol and Fatty Acid Production of Zymomonas mobilis without Amino Acid and Vitamin. J. Ind. Microbiol. Biotechnol. 2016, 43, 861–871. [Google Scholar] [CrossRef]
- Zhou, S.; Yomano, L.; Shanmugam, K.; Ingram, L. Fermentation of 10%(w/v) Sugar to D (−)-Lactate by Engineered Escherichia coli B. Biotechnol. Lett. 2005, 27, 1891–1896. [Google Scholar] [CrossRef]
- Ajit, A.; Sulaiman, A.Z.; Chisti, Y. Production of Bioethanol by Zymomonas mobilis in High-Gravity Extractive Fermentations. Food Bioprod. Process. 2017, 102, 123–135. [Google Scholar] [CrossRef]
- Maleki, F.; Changizian, M.; Zolfaghari, N.; Rajaei, S.; Noghabi, K.A.; Zahiri, H.S. Consolidated Bioprocessing for Bioethanol Production by Metabolically Engineered Bacillus Subtilis Strains. Sci. Rep. 2021, 11, 13731. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Si, T.; Dai, J. Whole-Genome Regulation for Yeast Metabolic Engineering. Small Methods 2020, 4, 1900640. [Google Scholar] [CrossRef]
- Lu, R.; Cao, L.; Wang, K.; Ledesma-Amaro, R.; Ji, X.-J. Engineering Yarrowia Lipolytica to Produce Advanced Biofuels: Current Status and Perspectives. Bioresour. Technol. 2021, 341, 125877. [Google Scholar] [CrossRef] [PubMed]
- Qiao, K.; Wasylenko, T.M.; Zhou, K.; Xu, P.; Stephanopoulos, G. Lipid Production in Yarrowia Lipolytica Is Maximized by Engineering Cytosolic Redox Metabolism. Nat. Biotechnol. 2017, 35, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Qiao, Y.; Li, F.; Xu, Y.; Yan, Y.; Madzak, C.; Yan, J. Subcellular Engineering of Lipase Dependent Pathways Directed towards Lipid Related Organelles for Highly Effectively Compartmentalized Biosynthesis of Triacylglycerol Derived Products in Yarrowia Lipolytica. Metab. Eng. 2019, 55, 231–238. [Google Scholar] [CrossRef]
- Ledesma-Amaro, R.; Dulermo, R.; Niehus, X.; Nicaud, J.-M. Combining Metabolic Engineering and Process Optimization to Improve Production and Secretion of Fatty Acids. Metab. Eng. 2016, 38, 38–46. [Google Scholar] [CrossRef]
- Xue, J.; Balamurugan, S.; Li, T.; Cai, J.-X.; Chen, T.-T.; Wang, X.; Yang, W.-D.; Li, H.-Y. Biotechnological Approaches to Enhance Biofuel Producing Potential of Microalgae. Fuel 2021, 302, 121169. [Google Scholar] [CrossRef]
- Dębowski, M.; Zieliński, M.; Kazimierowicz, J.; Kujawska, N.; Talbierz, S. Microalgae Cultivation Technologies as an Opportunity for Bioenergetic System Development—Advantages and Limitations. Sustainability 2020, 12, 9980. [Google Scholar] [CrossRef]
- Sarwer, A.; Hamed, S.M.; Osman, A.I.; Jamil, F.; Al-Muhtaseb, A.H.; Alhajeri, N.S.; Rooney, D.W. Algal Biomass Valorization for Biofuel Production and Carbon Sequestration: A Review. Environ. Chem. Lett. 2022, 20, 2797–2851. [Google Scholar] [CrossRef]
- Kang, N.K.; Kim, E.K.; Kim, Y.U.; Lee, B.; Jeong, W.-J.; Jeong, B.; Chang, Y.K. Increased Lipid Production by Heterologous Expression of AtWRI1 Transcription Factor in Nannochloropsis Salina. Biotechnol. Biofuels 2017, 10, 1–14. [Google Scholar] [CrossRef]
- Yang, D.-W.; Syn, J.-W.; Hsieh, C.-H.; Huang, C.-C.; Chien, L.-F. Genetically Engineered Hydrogenases Promote Biophotocatalysis-Mediated H2 Production in the Green Alga Chlorella Sp. DT. Int. J. Hydrogen Energy 2019, 44, 2533–2545. [Google Scholar] [CrossRef]
- Sahoo, S.; Mahapatra, S.R.; Das, N.; Parida, B.K.; Rath, S.; Misra, N.; Suar, M. Functional Elucidation of Hypothetical Proteins Associated with Lipid Accumulation: Prioritizing Genetic Engineering Targets for Improved Algal Biofuel Production. Algal Res. 2020, 47, 101887. [Google Scholar] [CrossRef]
- Chen, Z.; Wu, Y.; Huang, J.; Liu, D. Metabolic Engineering of Klebsiella Pneumoniae for the de Novo Production of 2-Butanol as a Potential Biofuel. Bioresour. Technol. 2015, 197, 260–265. [Google Scholar] [CrossRef]
- Liu, R.; Zhu, F.; Lu, L.; Fu, A.; Lu, J.; Deng, Z.; Liu, T. Metabolic Engineering of Fatty Acyl-ACP Reductase-Dependent Pathway to Improve Fatty Alcohol Production in Escherichia coli. Metab. Eng. 2014, 22, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Huang, J.; Wu, Y.; Wu, W.; Zhang, Y.; Liu, D. Metabolic Engineering of Corynebacterium Glutamicum for the Production of 3-Hydroxypropionic Acid from Glucose and Xylose. Metab. Eng. 2017, 39, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Zeng, Q.; Han, S.; Wang, Z.; Dong, Q.; Bi, Y.; Zhao, Y. High-Efficient n-Butanol Production by Co-Culturing Clostridium Acetobutylicum and Saccharomyces Cerevisiae Integrated with Butyrate Fermentative Supernatant Addition. World J. Microbiol. Biotechnol. 2017, 33, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wu, Y.; Wu, W.; Zhang, Y.; Liu, D.; Chen, Z. Cofactor Recycling for Co-Production of 1,3-Propanediol and Glutamate by Metabolically Engineered Corynebacterium Glutamicum. Sci. Rep. 2017, 7, srep42246. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Sun, H.; Huang, J.; Wu, Y.; Liu, D. Metabolic Engineering of Klebsiella Pneumoniae for the Production of 2-Butanone from Glucose. PLoS ONE 2015, 10, e0140508. [Google Scholar] [CrossRef]
- Raj, S.M.; Rathnasingh, C.; Jo, J.-E.; Park, S. Production of 3-Hydroxypropionic Acid from Glycerol by a Novel Recombinant Escherichia coli BL21 Strain. Process Biochem. 2008, 43, 1440–1446. [Google Scholar] [CrossRef]
- Ko, J.K.; Enkh-Amgalan, T.; Gong, G.; Um, Y.; Lee, S. Improved Bioconversion of Lignocellulosic Biomass by Saccharomyces Cerevisiae Engineered for Tolerance to Acetic Acid. GCB Bioenergy 2020, 12, 90–100. [Google Scholar] [CrossRef]
- Wang, X.; Liao, B.; Li, Z.; Liu, G.; Diao, L.; Qian, F.; Yang, J.; Jiang, Y.; Zhao, S.; Li, Y. Reducing Glucoamylase Usage for Commercial-Scale Ethanol Production from Starch Using Glucoamylase Expressing Saccharomyces Cerevisiae. Bioresour. Bioprocess. 2021, 8, 1–14. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, Y.; Cai, W.; Zeng, J.; Liu, N.; Wan, Y.; Fu, G. Research Progress of Anti-Environmental Factor Stress Mechanism and Anti-Stress Tolerance Way of Saccharomyces Cerevisiae during the Brewing Process. Crit. Rev. Food Sci. Nutr. 2022. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ha, S.-J.; Kim, H.J.; Galazka, J.M.; Cate, J.H.; Jin, Y.-S.; Zhao, H. Investigation of the Functional Role of Aldose 1-Epimerase in Engineered Cellobiose Utilization. J. Biotechnol. 2013, 168, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Cunha, J.T.; Soares, P.O.; Romaní, A.; Thevelein, J.M.; Domingues, L. Xylose Fermentation Efficiency of Industrial Saccharomyces Cerevisiae Yeast with Separate or Combined Xylose Reductase/Xylitol Dehydrogenase and Xylose Isomerase Pathways. Biotechnol. Biofuels 2019, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ruchala, J.; Kurylenko, O.O.; Dmytruk, K.V.; Sibirny, A.A. Construction of Advanced Producers of First-and Second-Generation Ethanol in Saccharomyces Cerevisiae and Selected Species of Non-Conventional Yeasts (Scheffersomyces stipitis, Ogataea Polymorpha). J. Ind. Microbiol. Biotechnol. 2020, 47, 109–132. [Google Scholar] [CrossRef] [PubMed]
- Demeke, M.M.; Dietz, H.; Li, Y.; Foulquié-Moreno, M.R.; Mutturi, S.; Deprez, S.; Abt, T.D.; Bonini, B.M.; Lidén, G.; Dumortier, F.; et al. Development of a D-Xylose Fermenting and Inhibitor Tolerant Industrial Saccharomyces Cerevisiae Strain with High Performance in Lignocellulose Hydrolysates Using Metabolic and Evolutionary Engineering. Biotechnol. Biofuels 2013, 6, 89. [Google Scholar] [CrossRef] [PubMed]
- Karhumaa, K.; Sanchez, R.G.; Hahn-Hägerdal, B.; Gorwa-Grauslund, M.-F. Comparison of the Xylose Reductase-Xylitol Dehydrogenase and the Xylose Isomerase Pathways for Xylose Fermentation by Recombinant Saccharomyces Cerevisiae. Microb. Cell Fact. 2007, 6, 5. [Google Scholar] [CrossRef]
- Sato, T.K.; Tremaine, M.; Parreiras, L.S.; Hebert, A.S.; Myers, K.S.; Higbee, A.J.; Sardi, M.; McIlwain, S.J.; Ong, I.M.; Breuer, R.J. Directed Evolution Reveals Unexpected Epistatic Interactions That Alter Metabolic Regulation and Enable Anaerobic Xylose Use by Saccharomyces Cerevisiae. PLoS Genet. 2016, 12, e1006372. [Google Scholar]
- Nag Dasgupta, C.; Nayaka, S.; Toppo, K.; Singh, A.K.; Deshpande, U.; Mohapatra, A. Draft Genome Sequence and Detailed Characterization of Biofuel Production by Oleaginous Microalga Scenedesmus Quadricauda LWG002611. Biotechnol. Biofuels 2018, 11, 1–15. [Google Scholar] [CrossRef]
- Balamurugan, S.; Wang, X.; Wang, H.-L.; An, C.-J.; Li, H.; Li, D.-W.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. Occurrence of Plastidial Triacylglycerol Synthesis and the Potential Regulatory Role of AGPAT in the Model Diatom Phaeodactylum tricornutum. Biotechnol. Biofuels 2017, 10, 1–14. [Google Scholar] [CrossRef]
- Chen, C.; Kao, A.; Tsai, Z.; Chow, T.; Chang, H.; Zhao, X.; Chen, P.; Su, H.; Chang, J. Expression of Type 2 Diacylglycerol Acyltransferse Gene DGTT1 from Chlamydomonas Reinhardtii Enhances Lipid Production in Scenedesmus Obliquus. Biotechnol. J. 2016, 11, 336–344. [Google Scholar] [CrossRef]
- Li, D.-W.; Cen, S.-Y.; Liu, Y.-H.; Balamurugan, S.; Zheng, X.-Y.; Alimujiang, A.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. A Type 2 Diacylglycerol Acyltransferase Accelerates the Triacylglycerol Biosynthesis in Heterokont Oleaginous Microalga Nannochloropsis Oceanica. J. Biotechnol. 2016, 229, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Balduyck, L. Lipolytic Stability during Wet Storage of Autotrophic Microalgae. 2018. Available online: https://kuleuven.limo.libis.be/discovery/search?query=any,contains,LIRIAS3355010&tab=LIRIAS&search_scope=lirias_profile&vid=32KUL_KUL:Lirias&offset=0 (accessed on 19 July 2023).
- Poliner, E.; Pulman, J.A.; Zienkiewicz, K.; Childs, K.; Benning, C.; Farré, E.M. A Toolkit for Nannochloropsis Oceanica CCMP 1779 Enables Gene Stacking and Genetic Engineering of the Eicosapentaenoic Acid Pathway for Enhanced Long-chain Polyunsaturated Fatty Acid Production. Plant Biotechnol. J. 2018, 16, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Miao, X. Short Chain Fatty Acid Biosynthesis in Microalgae Synechococcus Sp. PCC 7942. Mar. Drugs 2019, 17, 255. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Yuan, C.; Jin, Y.; Hu, G.-R.; Li, F.-L. Characterization of 3-Ketoacyl-CoA Synthase in a Nervonic Acid Producing Oleaginous Microalgae Mychonastes Afer. Algal Res. 2018, 31, 225–231. [Google Scholar] [CrossRef]
- Wang, X.; Dong, H.-P.; Wei, W.; Balamurugan, S.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. Dual Expression of Plastidial GPAT1 and LPAT1 Regulates Triacylglycerol Production and the Fatty Acid Profile in Phaeodactylum tricornutum. Biotechnol. Biofuels 2018, 11, 1–14. [Google Scholar] [CrossRef]
- Zhulin, I.B. Databases for Microbiologists. J. Bacteriol. 2015, 197, 2458–2467. [Google Scholar] [CrossRef]
- Naas, A.E.; Solden, L.M.; Norbeck, A.D.; Brewer, H.; Hagen, L.H.; Heggenes, I.M.; McHardy, A.C.; Mackie, R.I.; Paša-Tolić, L.; Arntzen, M.Ø.; et al. “Candidatus Paraporphyromonas Polyenzymogenes” Encodes Multi-Modular Cellulases Linked to the Type IX Secretion System. Microbiome 2018, 6, 44. [Google Scholar] [CrossRef]
- Meneses, C.; Silva, B.; Medeiros, B.; Serrato, R.; Johnston-Monje, D. A Metagenomic Advance for the Cloning and Characterization of a Cellulase from Red Rice Crop Residues. Molecules 2016, 21, 831. [Google Scholar] [CrossRef]
- Kim, S.-K.; Russell, J.; Cha, M.; Himmel, M.E.; Bomble, Y.J.; Westpheling, J. Coexpression of a β-d-Xylosidase from Thermotoga Maritima and a Family 10 Xylanase from Acidothermus Cellulolyticus Significantly Improves the Xylan Degradation Activity of the Caldicellulosiruptor bescii Exoproteome. Appl. Environ. Microbiol. 2021, 87, e0052421. [Google Scholar] [CrossRef]
- Takeda, M.; Baba, S.; Okuma, J.; Hirose, Y.; Nishimura, A.; Takata, M.; Oda, K.; Shibata, D.; Kumasaka, T.; Kondo, Y. Metagenomic Mining and Structure-Function Studies of a Hyper-Thermostable Cellobiohydrolase from Hot Spring Sediment. Commun. Biol. 2022, 5, 247. [Google Scholar] [CrossRef] [PubMed]
- Kreuzenbeck, N.B.; Seibel, E.; Schwitalla, J.W.; Fricke, J.; Conlon, B.H.; Schmidt, S.; Hammerbacher, A.; Köllner, T.G.; Poulsen, M.; Hoffmeister, D.; et al. Comparative Genomic and Metabolomic Analysis of Termitomyces Species Provides Insights into the Terpenome of the Fungal Cultivar and the Characteristic Odor of the Fungus Garden of Macrotermes Natalensis Termites. mSystems 2022, 7, e0121421. [Google Scholar] [CrossRef]
- Reichart, N.J.; Bowers, R.M.; Woyke, T.; Hatzenpichler, R. High Potential for Biomass-Degrading Enzymes Revealed by Hot Spring Metagenomics. Front. Microbiol. 2021, 12, 668238. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.H.; Lai, A.G. Mixed Evolutionary Origins of Endogenous Biomass-Depolymerizing Enzymes in Animals. BMC Genom. 2018, 19, 483. [Google Scholar] [CrossRef]
- Piao, H.; Froula, J.; Du, C.; Kim, T.-W.; Hawley, E.R.; Bauer, S.; Wang, Z.; Ivanova, N.; Clark, D.S.; Klenk, H.-P.; et al. Identification of Novel Biomass-Degrading Enzymes from Genomic Dark Matter: Populating Genomic Sequence Space with Functional Annotation. Biotechnol. Bioeng. 2014, 111, 1550–1565. [Google Scholar] [CrossRef] [PubMed]
- DeAngelis, K.M.; Allgaier, M.; Chavarria, Y.; Fortney, J.L.; Hugenholtz, P.; Simmons, B.; Sublette, K.; Silver, W.L.; Hazen, T.C. Characterization of Trapped Lignin-Degrading Microbes in Tropical Forest Soil. PLoS ONE 2011, 6, e19306. [Google Scholar] [CrossRef]
- Morya, R.; Kumar, M.; Singh, S.S.; Thakur, I.S. Genomic Analysis of Burkholderia Sp. ISTR5 for Biofunneling of Lignin-Derived Compounds. Biotechnol. Biofuels 2019, 12, 277. [Google Scholar] [CrossRef]
- Pan, H.; Wang, J.; Wu, H.; Li, Z.; Lian, J. Synthetic Biology Toolkit for Engineering Cupriviadus Necator H16 as a Platform for CO2 Valorization. Biotechnol. Biofuels 2021, 14, 212. [Google Scholar] [CrossRef]
- Braga, L.P.P.; Pereira, R.V.; Martins, L.F.; Moura, L.M.S.; Sanchez, F.B.; Patané, J.S.L.; da Silva, A.M.; Setubal, J.C. Genome-Resolved Metagenome and Metatranscriptome Analyses of Thermophilic Composting Reveal Key Bacterial Players and Their Metabolic Interactions. BMC Genom. 2021, 22, 652. [Google Scholar] [CrossRef]
- Tahir, A.A.; Mohd Barnoh, N.F.; Yusof, N.; Mohd Said, N.N.; Utsumi, M.; Yen, A.M.; Hashim, H.; Mohd Noor, M.J.M.; Akhir, F.N.M.; Mohamad, S.E.; et al. Microbial Diversity in Decaying Oil Palm Empty Fruit Bunches (OPEFB) and Isolation of Lignin-Degrading Bacteria from a Tropical Environment. Microbes Environ. 2019, 34, 161–168. [Google Scholar] [CrossRef]
- Tan, F.; Cheng, J.; Zhang, Y.; Jiang, X.; Liu, Y. Genomics Analysis and Degradation Characteristics of Lignin by Streptomyces Thermocarboxydus Strain DF3-3. Biotechnol. Biofuels Bioprod. 2022, 15, 78. [Google Scholar] [CrossRef]
- Mezzina, M.P.; Manoli, M.T.; Prieto, M.A.; Nikel, P.I. Engineering Native and Synthetic Pathways in Pseudomonas Putida for the Production of Tailored Polyhydroxyalkanoates. Biotechnol. J. 2021, 16, e2000165. [Google Scholar] [CrossRef] [PubMed]
- Habe, H.; Sato, Y.; Kirimura, K. Microbial and Enzymatic Conversion of Levulinic Acid, an Alternative Building Block to Fermentable Sugars from Cellulosic Biomass. Appl. Microbiol. Biotechnol. 2020, 104, 7767–7775. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Heredia, S.; Peña-Castro, J.M.; Aguilar-Uscanga, M.G.; Olvera, C.; Nolasco-Hipólito, C.; del Moral, S. AmyJ33, a Truncated Amylase with Improved Catalytic Properties. Biotechnol. Lett. 2022, 44, 1447–1463. [Google Scholar] [CrossRef] [PubMed]
- Balagurunathan, B.; Jonnalagadda, S.; Tan, L.; Srinivasan, R. Reconstruction and Analysis of a Genome-Scale Metabolic Model for Scheffersomyces stipitis. Microb. Cell Factories 2012, 11, 27. [Google Scholar] [CrossRef]
- Johnson, C.W.; Beckham, G.T. Aromatic Catabolic Pathway Selection for Optimal Production of Pyruvate and Lactate from Lignin. Metab. Eng. 2015, 28, 240–247. [Google Scholar] [CrossRef]
- Keller, M.W.; Lipscomb, G.L.; Loder, A.J.; Schut, G.J.; Kelly, R.M.; Adams, M.W.W. A Hybrid Synthetic Pathway for Butanol Production by a Hyperthermophilic Microbe. Metab. Eng. 2015, 27, 101–106. [Google Scholar] [CrossRef]
- You, S.; Li, J.; Zhang, F.; Bai, Z.-Y.; Shittu, S.; Herman, R.-A.; Zhang, W.-X.; Wang, J. Loop Engineering of a Thermostable GH10 Xylanase to Improve Low-Temperature Catalytic Performance for Better Synergistic Biomass-Degrading Abilities. Bioresour. Technol. 2021, 342, 125962. [Google Scholar] [CrossRef]
- Guarnieri, M.T.; Ann Franden, M.; Johnson, C.W.; Beckham, G.T. Conversion and Assimilation of Furfural and 5-(Hydroxymethyl)Furfural by Pseudomonas Putida KT2440. Metab. Eng. Commun. 2017, 4, 22–28. [Google Scholar] [CrossRef]
- Hobbs, J.K.; Lee, S.M.; Robb, M.; Hof, F.; Barr, C.; Abe, K.T.; Hehemann, J.-H.; McLean, R.; Abbott, D.W.; Boraston, A.B. KdgF, the Missing Link in the Microbial Metabolism of Uronate Sugars from Pectin and Alginate. Proc. Natl. Acad. Sci. USA 2016, 113, 6188–6193. [Google Scholar] [CrossRef]
- Borjigin, Q.; Yu, X.; Gao, J.; Zhang, B.; Wang, Z.; Hu, S.; Han, S.; Sun, J.; Hu, W. Taxonomic Structure and Function of the Corn Stover Degradative Microbial Consortium GF-20 Following Growth on Different Sources of Nitrogen. Ann. Appl. Biol. 2022, 180, 236–246. [Google Scholar] [CrossRef]
- Moreno, A.D.; Carbone, A.; Pavone, R.; Olsson, L.; Geijer, C. Evolutionary Engineered Candida Intermedia Exhibits Improved Xylose Utilization and Robustness to Lignocellulose-Derived Inhibitors and Ethanol. Appl. Microbiol. Biotechnol. 2019, 103, 1405–1416. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.; Jones, A.D.; Sousa, L.; Piotrowski, J.; Jin, M.; Sarks, C.; Dale, B.E.; Balan, V. Water-Soluble Phenolic Compounds Produced from Extractive Ammonia Pretreatment Exerted Binary Inhibitory Effects on Yeast Fermentation Using Synthetic Hydrolysate. PLoS ONE 2018, 13, e0194012. [Google Scholar] [CrossRef] [PubMed]
- Cecchini, D.A.; Pepe, O.; Pennacchio, A.; Fagnano, M.; Faraco, V. Directed Evolution of the Bacterial Endo-β-1,4-Glucanase from Streptomyces Sp. G12 towards Improved Catalysts for Lignocellulose Conversion. AMB Express 2018, 8, 74. [Google Scholar] [CrossRef]
- HamediRad, M.; Lian, J.; Li, H.; Zhao, H. RNAi Assisted Genome Evolution Unveils Yeast Mutants with Improved Xylose Utilization. Biotechnol. Bioeng. 2018, 115, 1552–1560. [Google Scholar] [CrossRef]
- Sana, B.; Chia, K.H.B.; Raghavan, S.S.; Ramalingam, B.; Nagarajan, N.; Seayad, J.; Ghadessy, F.J. Development of a Genetically Programed Vanillin-Sensing Bacterium for High-Throughput Screening of Lignin-Degrading Enzyme Libraries. Biotechnol. Biofuels 2017, 10, 32. [Google Scholar] [CrossRef]
- Wohlbach, D.J.; Kuo, A.; Sato, T.K.; Potts, K.M.; Salamov, A.A.; Labutti, K.M.; Sun, H.; Clum, A.; Pangilinan, J.L.; Lindquist, E.A.; et al. Comparative Genomics of Xylose-Fermenting Fungi for Enhanced Biofuel Production. Proc. Natl. Acad. Sci. USA 2011, 108, 13212–13217. [Google Scholar] [CrossRef]
- Sherkhanov, S.; Korman, T.P.; Chan, S.; Faham, S.; Liu, H.; Sawaya, M.R.; Hsu, W.-T.; Vikram, E.; Cheng, T.; Bowie, J.U. Isobutanol Production Freed from Biological Limits Using Synthetic Biochemistry. Nat. Commun. 2020, 11, 4292. [Google Scholar] [CrossRef]
- Gassler, T.; Baumschabl, M.; Sallaberger, J.; Egermeier, M.; Mattanovich, D. Adaptive Laboratory Evolution and Reverse Engineering Enhances Autotrophic Growth in Pichia Pastoris. Metab. Eng. 2022, 69, 112–121. [Google Scholar] [CrossRef]
- Wang, J.; Anderson, K.; Yang, E.; He, L.; Lidstrom, M.E. Enzyme Engineering and in Vivo Testing of a Formate Reduction Pathway. Synth. Biol. 2021, 6, ysab020. [Google Scholar] [CrossRef]
- Schada von Borzyskowski, L.; Schulz-Mirbach, H.; Troncoso Castellanos, M.; Severi, F.; Gómez-Coronado, P.A.; Paczia, N.; Glatter, T.; Bar-Even, A.; Lindner, S.N.; Erb, T.J. Implementation of the β-Hydroxyaspartate Cycle Increases Growth Performance of Pseudomonas Putida on the PET Monomer Ethylene Glycol. Metab. Eng. 2023, 76, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Tremaine, M.; Grass, J.A.; Purdy, H.M.; Landick, R.; Kiley, P.J.; Reed, J.L. Systems Metabolic Engineering of Escherichia coli Improves Coconversion of Lignocellulose-Derived Sugars. Biotechnol. J. 2019, 14, e1800441. [Google Scholar] [CrossRef]
- Kao, M.-R.; Yu, S.-M.; Ho, T.-H.U.D. Improvements of the Productivity and Saccharification Efficiency of the Cellulolytic β-Glucosidase D2-BGL in Pichia Pastoris via Directed Evolution. Biotechnol. Biofuels 2021, 14, 126. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.A.; Gladden, J.M.; Kimbrel, J.A.; Simmons, B.A.; Singer, S.W.; Thelen, M.P. Guanidine Riboswitch-Regulated Efflux Transporters Protect Bacteria against Ionic Liquid Toxicity. J. Bacteriol. 2019, 201, e00069-19. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhang, C.; Yang, K.-L.; He, J. Unique Genetic Cassettes in a Thermoanaerobacterium Contribute to Simultaneous Conversion of Cellulose and Monosugars into Butanol. Sci. Adv. 2018, 4, e1701475. [Google Scholar] [CrossRef]
- Cheng, K.-K.; Lee, B.-S.; Masuda, T.; Ito, T.; Ikeda, K.; Hirayama, A.; Deng, L.; Dong, J.; Shimizu, K.; Soga, T.; et al. Global Metabolic Network Reorganization by Adaptive Mutations Allows Fast Growth of Escherichia coli on Glycerol. Nat. Commun. 2014, 5, 3233. [Google Scholar] [CrossRef]
- Yao, Z.; Guo, Y.; Wang, H.; Chen, Y.; Wang, Q.; Nielsen, J.; Dai, Z. A Highly Efficient Transcriptome-Based Biosynthesis of Non-Ethanol Chemicals in Crabtree Negative Saccharomyces Cerevisiae. Biotechnol. Biofuels Bioprod. 2023, 16, 37. [Google Scholar] [CrossRef]
- Goedegebuur, F.; Dankmeyer, L.; Gualfetti, P.; Karkehabadi, S.; Hansson, H.; Jana, S.; Huynh, V.; Kelemen, B.R.; Kruithof, P.; Larenas, E.A.; et al. Improving the Thermal Stability of Cellobiohydrolase Cel7A from Hypocrea Jecorina by Directed Evolution. J. Biol. Chem. 2017, 292, 17418–17430. [Google Scholar] [CrossRef]
- Johnson, C.W.; Abraham, P.E.; Linger, J.G.; Khanna, P.; Hettich, R.L.; Beckham, G.T. Eliminating a Global Regulator of Carbon Catabolite Repression Enhances the Conversion of Aromatic Lignin Monomers to Muconate in Pseudomonas Putida KT2440. Metab. Eng. Commun. 2017, 5, 19–25. [Google Scholar] [CrossRef]
- Rodionov, D.A.; Rodionova, I.A.; Rodionov, V.A.; Arzamasov, A.A.; Zhang, K.; Rubinstein, G.M.; Tanwee, T.N.N.; Bing, R.G.; Crosby, J.R.; Nookaew, I.; et al. Transcriptional Regulation of Plant Biomass Degradation and Carbohydrate Utilization Genes in the Extreme Thermophile Caldicellulosiruptor bescii. mSystems 2021, 6, e0134520. [Google Scholar] [CrossRef]
- Elmore, J.R.; Dexter, G.N.; Salvachúa, D.; O’Brien, M.; Klingeman, D.M.; Gorday, K.; Michener, J.K.; Peterson, D.J.; Beckham, G.T.; Guss, A.M. Engineered Pseudomonas Putida Simultaneously Catabolizes Five Major Components of Corn Stover Lignocellulose: Glucose, Xylose, Arabinose, p-Coumaric Acid, and Acetic Acid. Metab. Eng. 2020, 62, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Mazzoli, R.; Olson, D.G.; Concu, A.M.; Holwerda, E.K.; Lynd, L.R. In Vivo Evolution of Lactic Acid Hyper-Tolerant Clostridium thermocellum. New Biotechnol. 2022, 67, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Gleizer, S.; Ben-Nissan, R.; Bar-On, Y.M.; Antonovsky, N.; Noor, E.; Zohar, Y.; Jona, G.; Krieger, E.; Shamshoum, M.; Bar-Even, A.; et al. Conversion of Escherichia coli to Generate All Biomass Carbon from CO2. Cell 2019, 179, 1255–1263.e12. [Google Scholar] [CrossRef] [PubMed]
- Ceron-Chafla, P.; Chang, Y.-T.; Rabaey, K.; van Lier, J.B.; Lindeboom, R.E.F. Directional Selection of Microbial Community Reduces Propionate Accumulation in Glycerol and Glucose Anaerobic Bioconversion Under Elevated PCO2. Front. Microbiol. 2021, 12, 675763. [Google Scholar] [CrossRef]
- Kurgan, G.; Onyeabor, M.; Holland, S.C.; Taylor, E.; Schneider, A.; Kurgan, L.; Billings, T.; Wang, X. Directed Evolution of Zymomonas mobilis Sugar Facilitator Glf to Overcome Glucose Inhibition. J. Ind. Microbiol. Biotechnol. 2022, 49, kuab066. [Google Scholar] [CrossRef]
- Sievert, C.; Nieves, L.M.; Panyon, L.A.; Loeffler, T.; Morris, C.; Cartwright, R.A.; Wang, X. Experimental Evolution Reveals an Effective Avenue to Release Catabolite Repression via Mutations in XylR. Proc. Natl. Acad. Sci. USA 2017, 114, 7349–7354. [Google Scholar] [CrossRef]
- Zou, L.; Jin, X.; Tao, Y.; Zheng, Z.; Ouyang, J. Unraveling the Mechanism of Furfural Tolerance in Engineered Pseudomonas Putida by Genomics. Front. Microbiol. 2022, 13, 1035263. [Google Scholar] [CrossRef]
- Rodríguez, J.E.G.; Brojanigo, S.; Basaglia, M.; Favaro, L.; Casella, S. Efficient Production of Polyhydroxybutyrate from Slaughterhouse Waste Using a Recombinant Strain of Cupriavidus necator DSM 545. Sci. Total Environ. 2021, 794, 148754. [Google Scholar] [CrossRef]
- Cai, R.-F.; Liu, L.; Chen, F.-F.; Li, A.; Xu, J.-H.; Zheng, G.-W. Reductive Amination of Biobased Levulinic Acid to Unnatural Chiral γ-Amino Acid Using an Engineered Amine Dehydrogenase. ACS Sustain. Chem. Eng. 2020, 8, 17054–17061. [Google Scholar] [CrossRef]
- Tan, G.-Y.A.; Chen, C.-L.; Ge, L.; Li, L.; Tan, S.N.; Wang, J.-Y. Bioconversion of Styrene to Poly(Hydroxyalkanoate) (PHA) by the New Bacterial Strain Pseudomonas Putida NBUS12. Microbes Environ. 2015, 30, 76–85. [Google Scholar] [CrossRef]
- Palermo, G.C.d.L.; Coutouné, N.; Bueno, J.G.R.; Maciel, L.F.; Dos Santos, L.V. Exploring Metal Ion Metabolisms to Improve Xylose Fermentation in Saccharomyces Cerevisiae. Microb. Biotechnol. 2021, 14, 2101–2115. [Google Scholar] [CrossRef] [PubMed]
- Akberdin, I.R.; Collins, D.A.; Hamilton, R.; Oshchepkov, D.Y.; Shukla, A.K.; Nicora, C.D.; Nakayasu, E.S.; Adkins, J.N.; Kalyuzhnaya, M.G. Rare Earth Elements Alter Redox Balance in Methylomicrobium alcaliphilum 20ZR. Front. Microbiol. 2018, 9, 2735. [Google Scholar] [CrossRef] [PubMed]
- Yuzawa, S.; Mirsiaghi, M.; Jocic, R.; Fujii, T.; Masson, F.; Benites, V.T.; Baidoo, E.E.K.; Sundstrom, E.; Tanjore, D.; Pray, T.R.; et al. Short-Chain Ketone Production by Engineered Polyketide Synthases in Streptomyces Albus. Nat. Commun. 2018, 9, 4569. [Google Scholar] [CrossRef]
- Valencia, L.E.; Incha, M.R.; Schmidt, M.; Pearson, A.N.; Thompson, M.G.; Roberts, J.B.; Mehling, M.; Yin, K.; Sun, N.; Oka, A.; et al. Engineering Pseudomonas Putida KT2440 for Chain Length Tailored Free Fatty Acid and Oleochemical Production. Commun. Biol. 2022, 5, 1363. [Google Scholar] [CrossRef] [PubMed]
- Stirling, F.; Silver, P.A. Controlling the Implementation of Transgenic Microbes: Are We Ready for What Synthetic Biology Has to Offer? Mol. Cell 2020, 78, 614–623. [Google Scholar] [CrossRef]
- Jefferson, C.; Lentzos, F.; Marris, C. Synthetic Biology and Biosecurity: Challenging the “Myths”. Front. Public Health 2014, 2, 115. [Google Scholar] [CrossRef]
- Ciesielski, P.N.; Pecha, M.B.; Lattanzi, A.M.; Bharadwaj, V.S.; Crowley, M.F.; Bu, L.; Vermaas, J.V.; Steirer, K.X.; Crowley, M.F. Advances in Multiscale Modeling of Lignocellulosic Biomass. ACS Sustain. Chem. Eng. 2020, 8, 3512–3531. [Google Scholar] [CrossRef]
- Beckham, G.T.; Matthews, J.F.; Peters, B.; Bomble, Y.J.; Himmel, M.E.; Crowley, M.F. Molecular-Level Origins of Biomass Recalcitrance: Decrystallization Free Energies for Four Common Cellulose Polymorphs. J. Phys. Chem. B 2011, 115, 4118–4127. [Google Scholar] [CrossRef]
- Ciesielski, P.N.; Pecha, M.B.; Thornburg, N.E.; Crowley, M.F.; Gao, X.; Oyedeji, O.; Sitaraman, H.; Brunhart-Lupo, N. Bridging Scales in Bioenergy and Catalysis: A Review of Mesoscale Modeling Applications, Methods, and Future Directions. Energy Fuels 2021, 35, 14382–14400. [Google Scholar] [CrossRef]
- Vermaas, J.V.; Crowley, M.F.; Beckham, G.T. A Quantitative Molecular Atlas for Interactions Between Lignin and Cellulose. ACS Sustain. Chem. Eng. 2019, 7, 19570–19583. [Google Scholar] [CrossRef]
- Sarkar, D.; Santiago, I.J.; Vermaas, J.V. Atomistic Origins of Biomass Recalcitrance in Organosolv Pretreatment. Chem. Eng. Sci. 2023, 272, 118587. [Google Scholar] [CrossRef]
- Mottiar, Y.; Vanholme, R.; Boerjan, W.; Ralph, J.; Mansfield, S.D. Designer Lignins: Harnessing the Plasticity of Lignification. Curr. Opin. Biotechnol. 2016, 37, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Kishani, S.; Benselfelt, T.; Wågberg, L.; Wohlert, J. Entropy Drives the Adsorption of Xyloglucan to Cellulose Surfaces—A Molecular Dynamics Study. J. Colloid Interface Sci. 2021, 588, 485–493. [Google Scholar] [CrossRef] [PubMed]
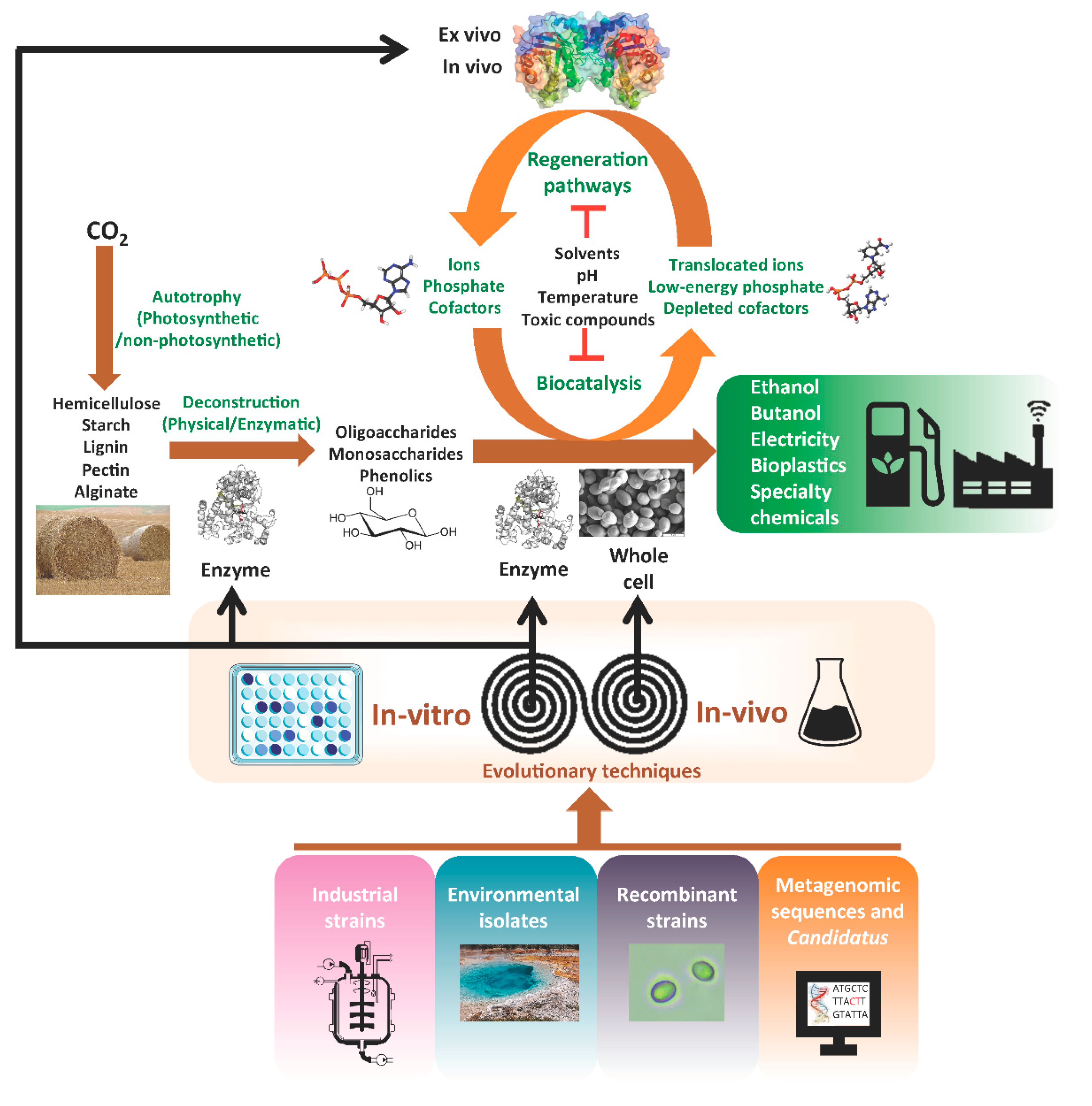

{kind=link}
{kind=link}
| Microorganism | Substrate | Product | Pathway | Improvement of Modified Strain | References | |
|---|---|---|---|---|---|---|
| Bacteria | ||||||
| Klebsiella pneumoniae | Glucose | 2-Butanol | Meso-2,3-butanediol synthesis | 320 mg/L of 2-butanol 720 mg/L by knocking the IdhA gene and adding coenzyme B12 1030 mg/L by engineering the diol hydratase | [44] | |
| Escherichia coli | Synthetic medium | Fatty alcohol | Fatty acyl-ACP reductase-dependent | 0.75 g/L of fatty alcohol | [45] | |
| Corynebacterium glutamicum | Glucose, xylose | 3-hydroxypropionic (HP) acid | Glycerol | 62.6 g/L 3-HP | [46] | |
| Clostridium acetobutylicum and Saccharomyces cerevisiae (yeast) | Glucose, corn, corn stover, and starch | n-Butanol | Clostridial acetoacetyl-CoA-derived pathway | 16.3 g/L of butanol | [47] | |
| Corynebacterium glutamicum | Glycerol | 1,3-propanediol (1,3-PDO) | Glutamate fermentation | Conversion of glycerol into 1,3-PDO of 1.0 mol/mol glycerol | [48] | |
| Klebsiella pneumoniae | Glucose | 2-butanone | 2, 3-butanediol synthesis pathway | 450 mg/L of 2-butanone | [49] | |
| Escherichia coli | Glycerol | 3-hydroxypropionic acid (3-HP) | Expression of dhaB and aldH | 31 g/L of 3-HP | [50] | |
| Escherichia coli | Glucose | 3-hydroxypropionic acid (3-HP) | Modulation of malonyl-CoA reductase (MCR) activity | 40.6 g/L of 3-HP | ||
| Yeast | ||||||
| Saccharomyces cerevisiae strain XUSAE57 | Xylose and Glucose | Ethanol | Xylose-isomerase pathway | 0.43–0.50 g ethanol/g xylose | [51] | |
| Saccharomyces cerevisiae | Raw corn or casaba starch | Ethanol | Glucoamylase expression | Better fermentation performance, observing a reduction of at least 40% in the dose of glucoamylase | [52] | |
| Saccharomyces cerevisiae | Sucrose | Ethanol | Modification to resist chemical stress | Increased the ability to resist stress factors by changing the cell membrane components, expressing transcriptional regulatory factors, activating the anti-stress metabolic pathway, and enhancing ROS scavenging ability | [53] | |
| Saccharomyces cerevisiae | Non-glucose sugars and cellulose | Ethanol | Xylose isomerase and 1-epimerase expression | Improved cellobiose utilization | [54] | |
| Saccharomyces cerevisiae | Cellulose | Ethanol | Xylose isomerase expression | It was found to improve ethanol production from non-detoxified hemicellulosic hydrolysates | [55] | |
| Scheffersomyces stipitis | Glucose, xylose, arabinose | Ethanol | Xylose reductase | Improvements of xylose fermentation on lignocellulose, showing defects in glucose catabolite repression and are more resistance to inhibitors present in hydrolysates | [56] | |
| Saccharomyces cerevisiae | Corn cob hemicellulosic hydrolysate | Ethanol | Introduction of Xylose isomerase (XI) and xylose reductase/xylitol dehydrogenase (XR/XDH) pathways | High ethanol productivities and yields from xylose | [55] | |
| Saccharomyces cerevisiae | Lignocellulose hydrolysates of Arundo donax | Ethanol | Expression cassette containing 13 genes including Clostridium phytofermentans XylA, encoding D-xylose isomerase (XI), and enzymes of the pentose phosphate pathway | Increased ethanol titer of 5.8% (v/v) | [57] | |
| Saccharomyces cerevisiae TMB3400 | Xylose | Ethanol | Pentose fermentation XR/XDH (S. stipitis XYL1, XYL2) by SsXYL1, SsXYL2 + XKS1↑, random mutagenesis | Increased ethanol values up to 0.33 g/g | [58] | |
| Saccharomyces cerevisiae GLBRCY87 | Glucose and xylose | Ethanol | SsXYL1, SsXYL2, SsXYL3, evolved on xylose and hydrolysate inhibitors | Increased ethanol values up to 0.34 g/g | [59] | |
| Microalgae | ||||||
| Scenedesmus quadricauda | Beijerinck medium | Triacylglycerols (TAGs) | Elevating intracellular malonyl-CoA and glycerol-3-phosphate (G3P) by overexpression of Acetyl-CoA carboxylase (ACCase) genes | It was evaluated in S. quadricauda LWG002611 which exhibits high biomass as well as high lipid productivity, to improve it via molecular engineering | [60] | |
| Phaeodactylum tricornutum | F/2 medium without Na2SiO3·9H2O | Improvement in lipid accumulation | Overexpression of lysophosphatidic acid acyltransferases (LPAATases) | Increase of 1.81-fold in polyunsaturated fatty acids | [61] | |
| Scenedesmus obliquus | BG11 medium | Improvement in lipid content | Overexpression of the type 2 diacylglycerol acyltransferase (DGAT) | 127.8%, 20.0%, and 232.6% higher production of lipid content, biomass concentration, and biomass productivity, respectively, compared to wild-type strain | [62] | |
| Nannochloropsis oceanica | F/2 liquid medium or F/2 agar plates | Improvement in triacylglycerol | Overexpression of the diacylglycerol acyltransferase (DGAT) | Increase of 69% in neutral lipid content | [63] | |
| Phaeodactylum tricornutum | Algal and humus media | Improvement in total lipids | Deletion of multifunctional lipase | Rapid increase in free fatty acid (FFA) content in engineered microalgae | [64] | |
| Nannochloropsis salina | F2N medium | Fatty acids | Heterologous expression of AP2 type TF Wrinkled1 of Arabidopsis (AtWRI1 TF) | Increase in total lipid contents of 44.7% | [41] | |
| Nannochloropsis oceanica | F/2 medium | Fatty acids | Construction and overexpression of multiple fatty acid desaturases (FAD) in N. oceanica CCM P1779 | Enhanced ω 3 long-chain polyunsaturated fatty acids (LC-PUFAs) and eicosapentaenoic acid (EPA) production | [65] | |
| Synechococcus sp. | BG11 medium | Short chain fatty acids (SCFAs) | Antisense expression of Synpcc7942_0537 (fabB/F) and Synpcc7942_1455 (fabH) | Fatty acid composition analysis showed C14 increased by 65.19% and 130%, respectively, when fabB/F and fabH were antisense expressed. | [66] | |
| Mychonastes afer | BG-11 medium | Lipids | Cloning and expression of 3-ketoacyl-coA synthase gene from M. afer (MaKCS) in Saccharomyces cerevisiae BY4741 | Increased lipid content, especially nervonic acid, under stress conditions of high light and low nitrogen. | [67] | |
| Phaeodactylum tricornutum | f/2 medium | Lipids | Overexpression of glycerol-3-phosphate acyltransferase (GPAT1) and lysophosphatidic acid acyltransferase (LPAT1) | Increase in photosynthetic activity and lipid content without compromising growth | [68] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peña-Castro, J.M.; Muñoz-Páez, K.M.; Robledo-Narvaez, P.N.; Vázquez-Núñez, E. Engineering the Metabolic Landscape of Microorganisms for Lignocellulosic Conversion. Microorganisms 2023, 11, 2197. https://doi.org/10.3390/microorganisms11092197
Peña-Castro JM, Muñoz-Páez KM, Robledo-Narvaez PN, Vázquez-Núñez E. Engineering the Metabolic Landscape of Microorganisms for Lignocellulosic Conversion. Microorganisms. 2023; 11(9):2197. https://doi.org/10.3390/microorganisms11092197
Chicago/Turabian StylePeña-Castro, Julián Mario, Karla M. Muñoz-Páez, Paula N. Robledo-Narvaez, and Edgar Vázquez-Núñez. 2023. "Engineering the Metabolic Landscape of Microorganisms for Lignocellulosic Conversion" Microorganisms 11, no. 9: 2197. https://doi.org/10.3390/microorganisms11092197
APA StylePeña-Castro, J. M., Muñoz-Páez, K. M., Robledo-Narvaez, P. N., & Vázquez-Núñez, E. (2023). Engineering the Metabolic Landscape of Microorganisms for Lignocellulosic Conversion. Microorganisms, 11(9), 2197. https://doi.org/10.3390/microorganisms11092197