

High Prevalence of Novel Sequence Types in Streptococcus pneumoniae That Caused Invasive Diseases in Kuwait in 2018



Abstract
1. Introduction
2. Materials and Methods
2.1. S. pneumoniae Isolates
2.2. DNA Preparation
2.3. PCR Assays
2.4. MLST Determination
2.5. Antibiotic Susceptibility Testing
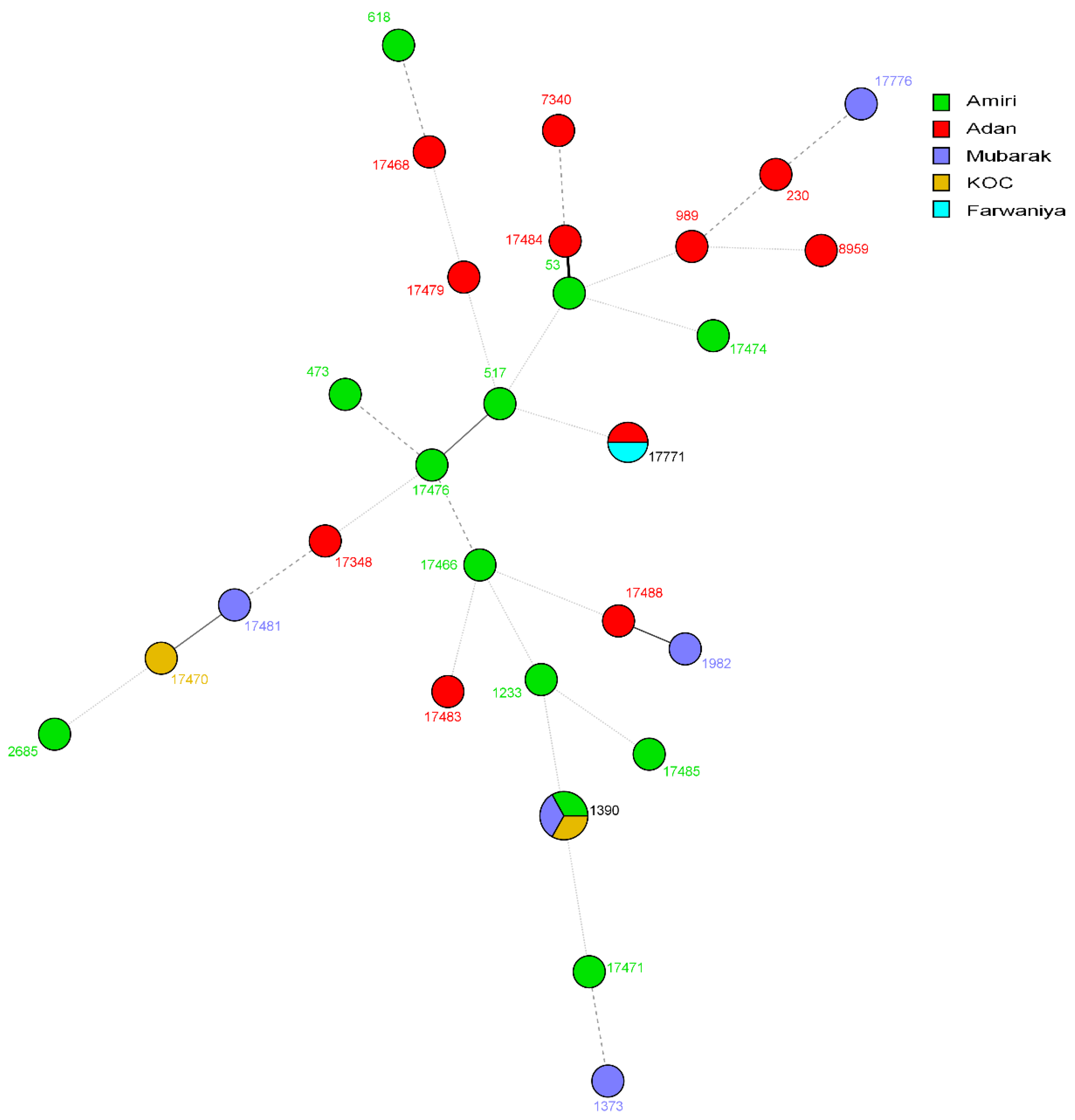
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shetty, A.K.; Maldonado, Y.A. Current trends in Streptococcus pneumoniae infections and their treatment. Curr. Pediatr. Rep. 2013, 1, 158–169. [Google Scholar] [CrossRef][Green Version]
- World Health Organization. Surveillance Vaccine Preventable. Pneumococcus; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Mokaddas, E.M.; Rotimi, V.O.; Albert, M.J. Increasing prevalence of antimicrobial resistance in Streptococcus pneumoniae in Kuwait: Implications for therapy. Microb. Drug Resist. 2007, 13, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Mokaddas, E.; Albert, M.J. Serotype distribution and penicillin non-susceptibility of Streptococcus pneumoniae causing invasive diseases in Kuwait: A 10-year study of impact of pneumococcal conjugate vaccines. Expert Rev. Vaccines 2016, 15, 1337–1345. [Google Scholar] [CrossRef] [PubMed]
- Ganaie, F.; Saad, J.S.; McGee, L.; van Tonder, A.J.; Bentley, S.D.; Lo, S.W.; Gladstone, R.A.; Turner, P.; Keenan, J.D.; Breiman, R.F.; et al. A new pneumococcal capsule type, 10D, is the 100th serotype and has a large cps fragment from an oral streptococcus. mBio 2020, 11, e00937-40. [Google Scholar] [CrossRef] [PubMed]
- Cleary, D.W.; Jones, J.; Gladstone, R.A.; Osman, K.L.; Devine, V.T.; Jefferies, J.M.; Bentley, S.D.; Faust, S.N.; Clarke, S.C. Changes in serotype prevalence of Streptococcus pneumoniae in Southampton, UK between 2006 and 2018. Sci. Rep. 2022, 12, 13332. [Google Scholar] [CrossRef] [PubMed]
- Cilloniz, C.; Garcia-Vidal, C.; Ceccati, A.; Torres, A. Antimicrobial resistance among Streptococcus pneumoniae. In Antimicrobial Resistance in the 21st Century, Emerging Infectious Diseases of the 21st Century; Fong, I.W., Shalaes, D., Drilca, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 13–38. [Google Scholar]
- Rayner, R.E.; Savill, J.; Hafner, L.M.; Huygens, F. Genotyping Streptococcus pneumoniae. Future Microbiol. 2015, 10, 653–664. [Google Scholar] [CrossRef]
- Zeng, Y.; Song, Y.; Cui, L.; Wu, Q.; Wang, C.; Coelho, A.C.; Zhang, G.; Wei, D.; Li, C.; Zhang, J.; et al. Phylogenic insights into evolutionary trajectories of multidrug resistant S. pneumoniae CC271 over a period of 14 years in China. Genome Med. 2023, 15, 46. [Google Scholar] [CrossRef]
- Satzke, C.; Ortika, B.D.; Oftadeh, S.; Russell, F.M.; Robins-Browne, R.M.; Mulholland, E.K.; Gilbert, G.L. Molecular epidemiology of Streptococcus pneumoniae serogroup 6 isolates from Fjian children including newly identified serotypes 6C and 6D. J. Clin. Microbiol. 2010, 48, 4298–4300. [Google Scholar] [CrossRef]
- Moreno, J.; Duarte, C.; Cassiolato, A.P.; Chacón, G.C.; Alarcon, P.; Sánchez, J.; Martín, Y.N.S.; Valenzuela, C.; Castillo, W.; Gabarrot, G.G.; et al. Molecular characterization of Latin American invasive Streptococcus pneumoniae serotype 19A isolates. Vaccine 2020, 38, 3524–3530. [Google Scholar] [CrossRef]
- Pillai, D.R.; Shahinas, D.; Buzina, A.; Pollock, R.A.; Lau, R.; Khairnar, K.; Wong, A.; Farrell, D.J.; Green, K.; McGeer, A.; et al. Genome-wide dissection of globally emergent multi-drug resistant serotype 19A Streptococcus pneumoniae. BMC Genom. 2009, 10, 642. [Google Scholar] [CrossRef]
- Mokaddas, E.; Syed, S.; Albert, M.J. The 13-valent pneumococcal conjugate vaccine (PCV13) does not appear to provide much protection on combined invasive disease due to the six PCV13 non-PCV7 serotypes 1, 3, 5, 6A, 7F, and 19A in Kuwait during 2010–2019. Hum. Vaccin. Immunother. 2021, 17, 4661–4666. [Google Scholar] [CrossRef]
- Sorensen, U.B. Typing of pneumococci by using 12 pooled antisera. J. Clin. Microbiol. 1993, 31, 2097–2100. [Google Scholar] [CrossRef] [PubMed]
- Mokaddas, E.; Ahmad, S.; Abal, A.T. Molecular fingerprinting of isoniazid-resistant Mycobacterium tuberculosis isolates from chest diseases hospital in Kuwait. Microbiol. Immunol. 2002, 46, 767–771. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Enright, M.C.; Spratt, B.G. A multilocus sequence typing scheme for Streptococcus pneumoniae: Identification of clones associated with serious invasive disease. Microbiol. 1998, 144, 3049–3060. [Google Scholar] [CrossRef] [PubMed]
- Asadzadeh, M.; Ahmad, S.; Al-Sweih, N.; Khan, Z. Molecular fingerprinting studies do not support intrahospital transmission of Candida albicans among candidemia patients in Kuwait. Front. Microbiol. 2017, 8, 247. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- Li, X.-X.; Xiao, S.Z.; Gu, F.F.; Zhao, S.Y.; Xie, Q.; Sheng, Z.K.; Ni, Y.X.; Qu, J.M.; Han, L.Z. Serotype distribution, antimicrobial susceptibility, and multilocus sequence types (MLST) of Streptococcus pneumoniae from adults of three hospitals in Shanghai, China. Front. Cell. Infect. Microbiol. 2019, 9, 407. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, Z.; Zhang, L.; Kudinha, T.; An, H.; Qian, C.; Jiang, B.; Wang, Y.; Xu, Y.; Liu, Z.; et al. Serotype distribution, antimicrobial susceptibility, multilocus sequence type and virulence of invasive Streptococcus pneumoniae in China: A six-year multicenter study. Front. Microbiol. 2022, 12, 798750. [Google Scholar] [CrossRef]
- Sener, B.; McGee, L.; Pinar, A.; Eser, O. Genomic background of drug-resistant Streptococcus pneumoniae in Ankara, Turkey: Identification of emerging new clones. Microb. Drug Resist. 2006, 12, 109–114. [Google Scholar] [CrossRef]
- Candeias, C.; Felix, S.; Handem, S.; de Lencastre, H.; Sa-Leao, R. Clonal changes in the pneumococcal population carried by Portuguese children during six years of private use of the 13-valent pneumococcal conjugate vaccine: The relative contribution of clonal expansion, clonal emergence, and capsular switch events. Microbiol. Spectr. 2023, 11, e02909-22. [Google Scholar] [CrossRef]
- Gagetti, P.; Lo, S.W.; Hawkins, P.A.; Gladstone, R.A.; Regueira, M.; Faccone, D.; Klugman, K.P.; Breiman, R.F.; McGee, L.; Bentley, S.D.; et al. Population genetic structure, serotype distribution and antibiotic resistance of Streptococcus pneumoniae causing invasive disease in children in Argentina. Microb. Genom. 2021, 7, 000636. [Google Scholar] [CrossRef]
- Chewapreecha, C. Dense genomic sampling identifies highways of pneumococcal recombination. Nat Genet. 2014, 46, 305–309. [Google Scholar] [CrossRef]
- Straume, D.; Stamsas, D.A.; Havarstein, L.S. Natural transformation and genome evolution in Streptococcus pneumoniae. Infect. Genet. Evol. 2015, 33, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Croucher, N.J.; Harris, S.R.; Fraser, C.; Quail, M.A.; Burton, J.; van der Linden, M.; McGee, L.; von Gottberg, A.; Song, J.H.; Ko, K.S.; et al. Rapid pneumococcal evolution in response to clinical interventions. Science 2011, 331, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Croucher, N.J.; Finkelstein, J.A.; Pelton, S.I.; Mitchell, P.K.; Lee, G.M.; Parkhill, J.; Bentley, S.D.; Hanage, W.P.; Lipsitch, M. Population genomics of post-vaccine changes in pneumococcal epidemiology. Nat. Genet. 2013, 45, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Torumkuney, D.; Behbehani, N.; Hasselt, J.; Hamouda, M.; Keles, N. Country data on AMR in Kuwait in the context of community-acquired respiratory tract infections: Links between antibiotic susceptibility, local and international antibiotic prescribing guidelines, access to medicines, and clinical outcome. J. Antimicrob. Chemother. 2022, 77 (Suppl. S1), i77–i83. [Google Scholar] [CrossRef] [PubMed]
- Chaguza, C.; Cornick, J.E.; Everett, D.B. Mechanisms and impact of genetic recombination in the evolution of Streptococcus pneumoniae. Comput. Struct. Biotech J. 2015, 13, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Sandgren, A.; Sjöström, K.; Olsson-Liljequist, B.; Christensson, B.; Samuelsson, A.; Kronvall, G.; Normark, B.H. Effect of clonal and serotype-specific properties on the invasive capacity of Streptococcus pneumoniae. J. Infect. Dis. 2004, 189, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Dagerhamn, J.; Blomberg, C.; Browall, S.; Sjöström, K.; Morfeldt, E.; Henriques-Normark, B. Determination of accessory gene patterns predicts the same relatedness among strains of Streptococcus pneumoniae as sequencing of housekeeping genes does and represents a novel approach in molecular epidemiology. J. Clin. Microbiol. 2008, 46, 863–868. [Google Scholar] [CrossRef]
- Ktari, S.; Ben Ayed, N.; Ben Rbeh, I.; Garbi, N.; Maalej, S.; Mnif, B.; Rhimi, F.; Hammami, A. Antibiotic resistance pattern, capsular types, and molecular characterization of invasive isolates of Streptococcus pneumoniae in the south of Tunisia from 2012 to 2018. BMC. Microbiol. 2023, 23, 36. [Google Scholar] [CrossRef]
- Wambugu, P.; Shah, M.-M.; Nguyen, H.-A.; Le, K.-A.; Le, H.-H.; Vo, H.-M.; Toizumi, M.; Bui, M.-X.; Dang, D.-A.; Yoshida, L.-M. Molecular epidemiology of Streptococcus pneumoniae detected in hospitalized pediatric acute respiratory infection cases in Central Vietnam. Pathogens 2023, 12, 943. [Google Scholar] [CrossRef]
- Bricio-Moreno, L.; Ebruke, C.; Chaguza, C.; Cornick, J.; Kwambana-Adams, B.; Yang, M.; Mackenzie, G.; Wren, B.W.; Everett, D.; Antonio, M.; et al. Comparative genomic analysis and in vivo modelling of streptococcus pneumoniae ST3081 and ST618 isolates reveal key genetic and phenotypic differences contributing to clonal replacement of serotype 1 in the Gambia. J. Infect. Dis. 2017, 216, 1318–1327. [Google Scholar] [CrossRef] [PubMed]
- del Amo, E.; Esteva, C.; Hernandez-Bou, S.; Galles, C.; Navarro, M.; Sauca, G.; Diaz, A.; Gassiot, P.; Marti, C.; Larrosa, N.; et al. Serotypes and clonal diversity of Streptococcus pneumoniae causing invasive disease in the era of PCV13 in Catalonia, Spain. PLoS ONE 2016, 11, e0151125. [Google Scholar] [CrossRef] [PubMed]
- Nurse-Lucas, M.; McGee, L.; Hawkins, P.A.; Swanston, W.H.; Akpaka, P.E. Serotypes and genotypes of Streptococcus pneumoniae isolated from Trinidad and Tobago. Int. J. Infect. Dis. 2016, 46, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Guo, J.; Qin, T.; Ren, H.; Xu, Y.; Wang, C.; Xu, X. Serotype and MLST-based inference of population structure of clinical Streptococcus pneumoniae from invasive and noninvasive pneumococcal disease. Infect. Gen. Evol. 2017, 55, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Cherazard, R.; Epstein, M.; Doan, T.L.; Salim, T.; Bharti, S.; Smith, M.A. Antimicrobial resistant Streptococcus pneumoniae: Prevalence, mechanisms, and clinical implications. Am. J. Ther. 2017, 24, e361–e369. [Google Scholar] [CrossRef]
- McGee, L.; McDougal, L.; Zhou, J.; Spratt, B.G.; Tenover, F.C.; George, R.; Hakenbeck, R.; Hryniewicz, W.; Lefévre, J.C.; Tomasz, A.; et al. Nomenclature of major antimicrobial-resistant clones of Streptococcus pneumoniae defined by the pneumococcal molecular epidemiology network. J. Clin. Microbiol. 2001, 39, 2565–2571. [Google Scholar] [CrossRef]
- Hansen, C.B.; Fuursted, K.; Valentiner-Branth, P.; Dalby, T.; Jorgensen, C.S.; Slotved, H.C. Molecular characterization and epidemiology of Streptococcus pneumoniae serotype 8 in Denmark. BMC. Infect. Dis. 2021, 21, 421. [Google Scholar] [CrossRef]
- Isturiz, R.; Sings, H.L.; Hilton, B.; Arguedas, A.; Reinert, R.R.; Jodar, L. Streptococcus pneumoniae serotype 19A: Worldwide epidemiology. Expert Rev. Vaccines 2017, 16, 1007–1027. [Google Scholar] [CrossRef]
- Desmet, S.; Theeten, H.; Laenen, L.; Cuypers, L.; Maes, P.; Bossuyt, W.; Van Heirstraeten, L.; Peetermans, W.E.; Lagrou, K. Characterization of emerging serotype 19A pneumococcal strains in invasive disease and carriage, Belgium. Emerg. Infect. Dis. 2022, 28, 1606–1614. [Google Scholar] [CrossRef]
- Muller, A.; Kleynhans, J.; de Gouveia, L.; Meiring, S.; Cohen, C.; Hathaway, L.J.; von Gottberg, A.; for GERMS-SA. Streptococcus pneumoniae serotypes associated with death, South Africa, 2012–2018. Emerg. Infect. Dis. 2022, 28, 166–179. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Isolate No. | Specimen | Patient | Hospital | Serotype | Sequence Type | |
|---|---|---|---|---|---|---|
| Age (y/m/d) | Gender | (ST) | ||||
| 1949 | Blood | 11 m | M | Adan | 9N | 17479 * |
| 1963 | Blood | 53 y | M | Amiri | 9V | 17466 * |
| 1964 | Blood | 2 y | M | Amiri | 28A | 17474 * |
| 1967 | Blood | 78 y | F | Amiri | 33D | 473 |
| 1968 | Blood | 38 y | F | Amiri | 9N | 517 |
| 1998 | CSF | 38 y | M | Adan | 19F | 17348 |
| 2005 | Blood | 10 d | F | Adan | 12F | 989 |
| 2006 | Blood | 21 y | F | Adan | 15A | 17483 * |
| 2017 | Blood | 2 y | F | Adan | 24F | 230 |
| 2018 | Blood | 11 y | M | Adan | 15A | 7340 |
| 2028 | Blood | 25 y | F | Amiri | 19A | 17471 * |
| 2040 | Blood | 32 y | F | Amiri | 8 | 53 |
| 2050 | Blood | 71 y | F | KOC | 22A | 17470 * |
| 2055 | Blood | 60 y | M | Amiri | 18C | 1233 |
| 2070 | Blood | 21 y | M | Mubarak | 22A | 17481 * |
| 2076 | Blood | 29 y | F | Adan | 23A | 17771 * |
| 2080 | Blood | 59 y | M | Mubarak | 6C | 1390 |
| 2087 | Blood | 4 y | M | Mubarak | 34F | 17776 * |
| 2093 | CSF | 64 y | F | Adan | 29 | 17488 * |
| 2102 | CSF | 2 y | M | Farwaniya | 23A | 17771 * |
| 2111 | Blood | 85 y | F | Amiri | 35F | 17476 * |
| 2112 | CSF | 50 y | M | Amiri | 6C/6D | 1390 |
| 2116 | Blood | 32 y | M | Adan | 19A | 17468 * |
| 2117 | Blood | 75 y | M | KOC | 6C/6D | 1390 |
| 2122 | Blood | 69 y | M | Amiri | 11A | 2685 |
| 2126 | Blood | 3 m | M | Adan | 23B | 8959 |
| 2127 | Blood | 4 y | F | Mubarak | 23B | 1373 |
| 2128 | Blood | 6 d | F | Adan | 8 | 17484 * |
| 2129 | Blood | 9 m | M | Mubarak | 29 | 1982 |
| 2133 | Blood | 49 y | M | Amiri | 1 | 618 |
| 2134 | Blood | 72 y | M | Amiri | 17F | 17485 * |
| Specimen No. | Allele for Gene | New ST | ||||||
|---|---|---|---|---|---|---|---|---|
| aroE | gdh | gki | recP | spi | xpt | ddl | ||
| 1949 | 2 | 720 | 174 | 4 | 7 | 435 | 5 | 17479 |
| 1963 | 15 | 17 | 4 | 16 | 6 | 1 | 271 | 17466 |
| 1964 | 16 | 44 | 1 | 4 | 9 | 70 | 17 | 17474 |
| 2006 | 4 | 41 | 47 | 16 | 6 | 14 | 2 | 17483 |
| 2028 | 8 | 13 | 8 | 6 | 461 | 6 | 18 | 17471 |
| 2050 | 118 | 16 | 751 | 4 | 15 | 1 | 9 | 17470 |
| 2070 | 7 | 16 | 751 | 1 | 15 | 1 | 14 | 17481 |
| 2076 | 54 | 5 | 9 | 546 * | 6 | 273 | 8 | 17771 |
| 2087 | 5 | 19 | 2 | 547 * | 121 | 22 | 265 | 17776 |
| 2093 | 296 | 12 | 4 | 44 | 14 | 77 | 97 | 17488 |
| 2102 | 54 | 5 | 9 | 546 * | 6 | 273 | 8 | 17771 |
| 2111 | 7 | 5 | 4 | 4 | 6 | 1 | 79 | 17476 |
| 2116 | 2 | 18 | 4 | 1 | 7 | 232 | 9 | 17468 |
| 2128 | 2 | 5 | 1 | 11 | 16 | 3 | 271 | 17484 |
| 2134 | 10 | 11 | 426 | 5 | 6 | 88 | 9 | 17485 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mokaddas, E.; Asadzadeh, M.; Syed, S.; Albert, M.J. High Prevalence of Novel Sequence Types in Streptococcus pneumoniae That Caused Invasive Diseases in Kuwait in 2018. Microorganisms 2024, 12, 225. https://doi.org/10.3390/microorganisms12010225
Mokaddas E, Asadzadeh M, Syed S, Albert MJ. High Prevalence of Novel Sequence Types in Streptococcus pneumoniae That Caused Invasive Diseases in Kuwait in 2018. Microorganisms. 2024; 12(1):225. https://doi.org/10.3390/microorganisms12010225
Chicago/Turabian StyleMokaddas, Eiman, Mohammad Asadzadeh, Shabeera Syed, and M. John Albert. 2024. "High Prevalence of Novel Sequence Types in Streptococcus pneumoniae That Caused Invasive Diseases in Kuwait in 2018" Microorganisms 12, no. 1: 225. https://doi.org/10.3390/microorganisms12010225
APA StyleMokaddas, E., Asadzadeh, M., Syed, S., & Albert, M. J. (2024). High Prevalence of Novel Sequence Types in Streptococcus pneumoniae That Caused Invasive Diseases in Kuwait in 2018. Microorganisms, 12(1), 225. https://doi.org/10.3390/microorganisms12010225