
Exopolysaccharide-Producing Bacteria Regulate Soil Aggregates and Bacterial Communities to Inhibit the Uptake of Cadmium and Lead by Lettuce


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Screening of Exopolysaccharide-Producing Bacteria
2.2. Determination of the Ability of Exopolysaccharide-Producing Bacteria to Adsorb Heavy Metals
2.3. Determination of Biological Characteristics of Functional Strains
2.4. Adsorption of Cd and Pb by Exopolysaccharide-Producing Bacteria
2.5. Mechanism of Exopolysaccharide-Producing Bacteria Adsorbing Cd and Pb
2.6. Pot Experiment with Lettuce
2.7. Effects of Exopolysaccharide-Producing Bacteria on Lettuce Growth and Cd Uptake
2.8. Effects of Exopolysaccharide-Producing Bacteria on the Structure of Rhizosphere Soil Aggregates
2.9. Determination of Bacterial Community Structure in Soil Aggregates
2.10. Data Analysis
3. Results
3.1. Screening of Exopolysaccharide-Producing Bacteria with Heavy Metal Adsorption Capacity
3.2. Biological Characteristics of the Target Bacterial Strain
3.3. Adsorption of Cd and Pb by Functional Strains
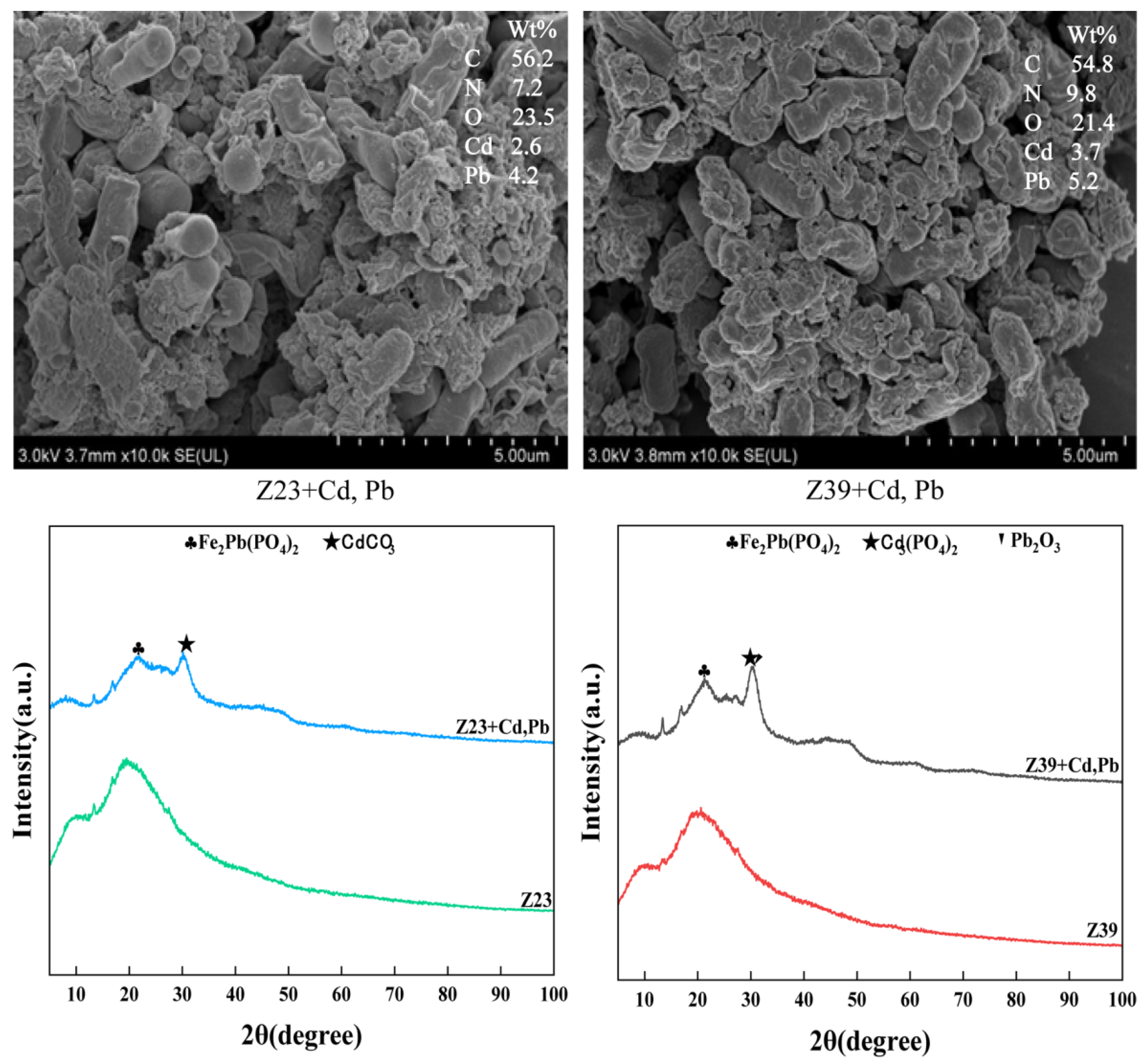
3.4. Mechanism of Functional Strains Adsorbing Cd and Pb
3.5. XPS Analysis
3.6. Dry Weight, Heavy Metal Content, and Antioxidant Enzyme Activity in Lettuce
3.7. Particle Size Distribution and Polysaccharide Content in Soil Aggregates
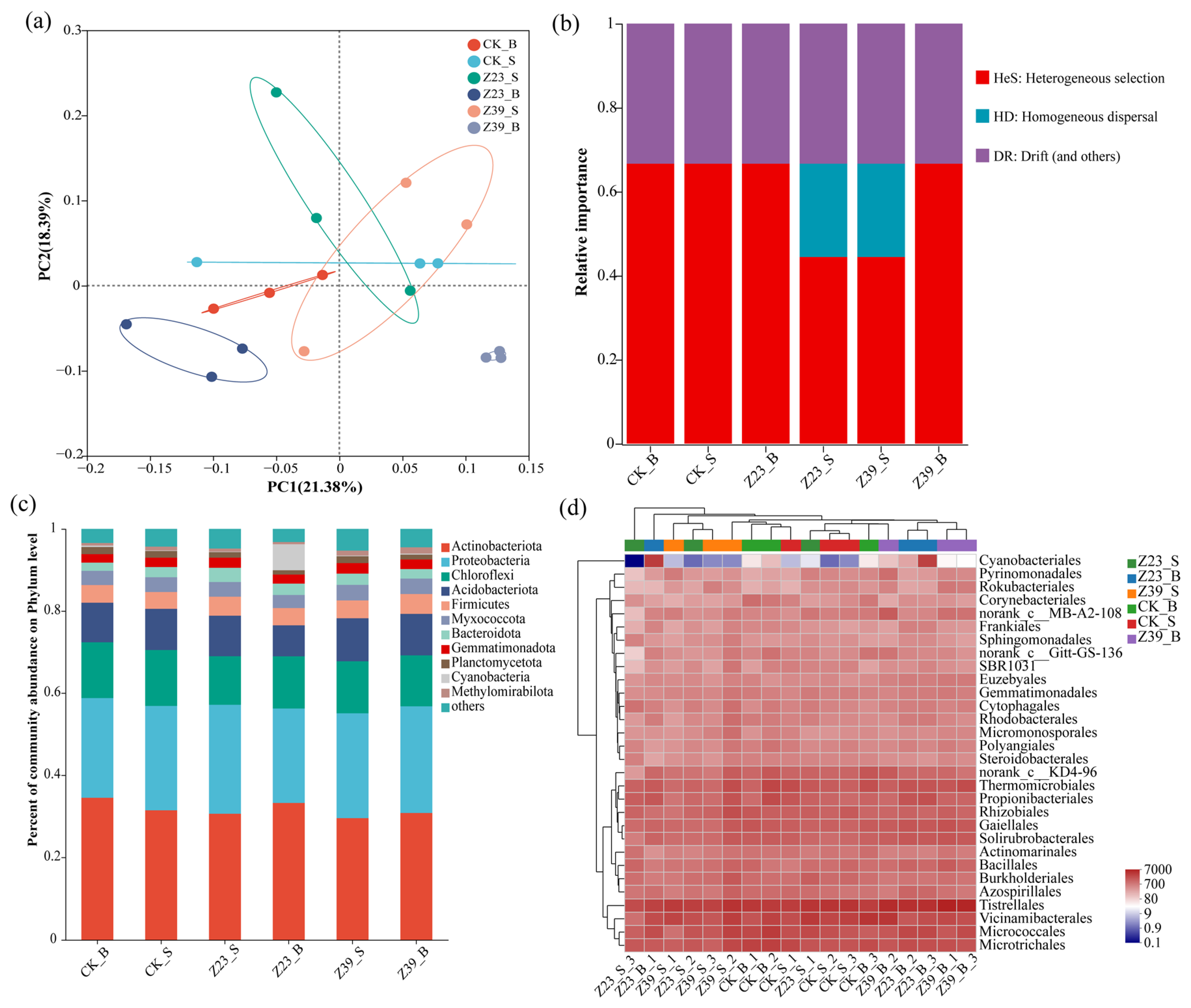
3.8. Bacterial Community Structure and Diversity of Soil Aggregates
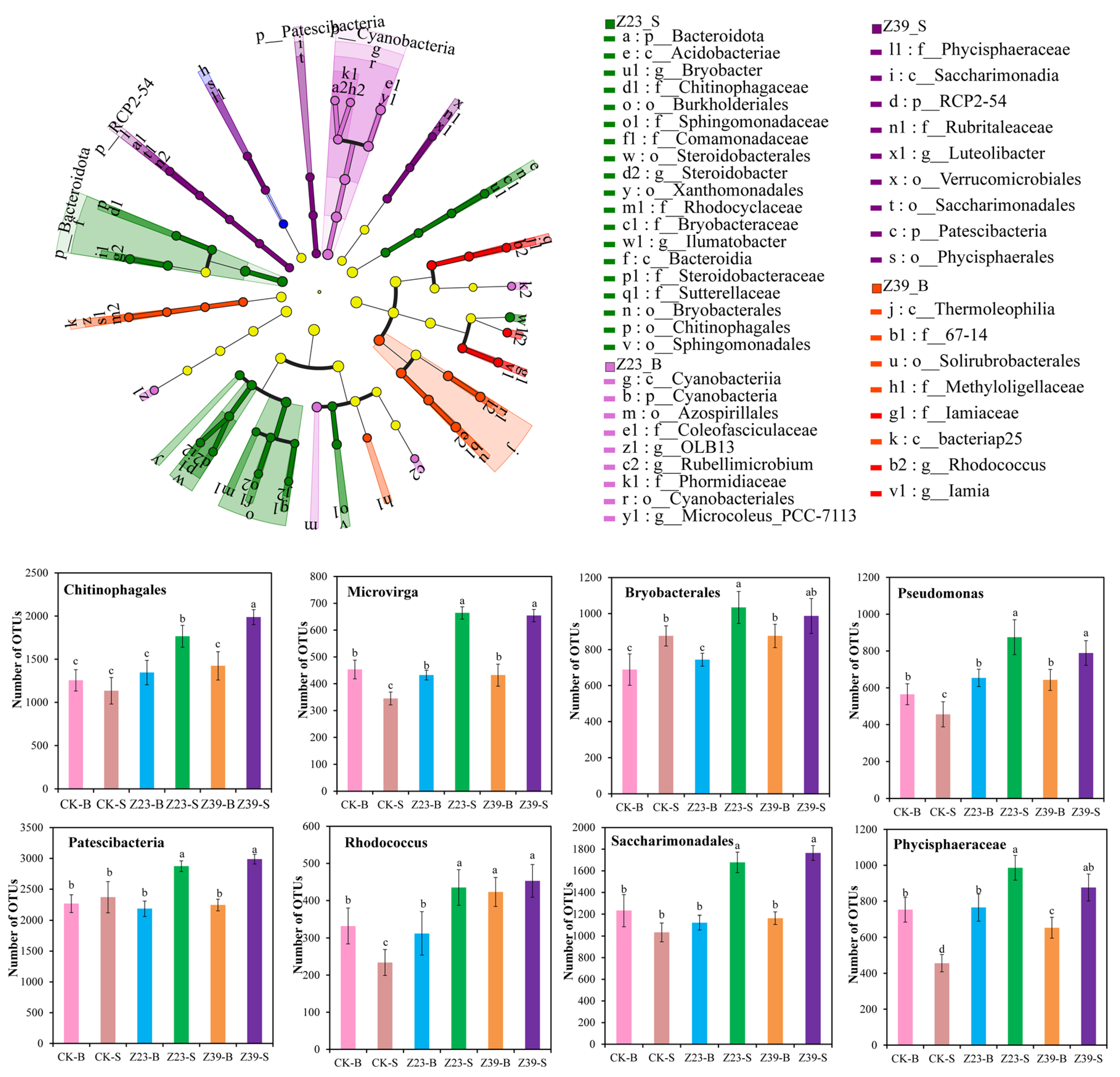
3.9. Functional Bacteria in Soil Aggregates
3.10. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhou, S.; Cheng, Q.; Weindorf, D.C.; Yang, B.; Gong, Z.; Yuan, Z. Multiple approaches for heavy metal contamination characterization and source identification of farmland soils in a metal mine impacted area. Appl. Geochem. 2024, 174, 106125. [Google Scholar] [CrossRef]
- Dong, S.; Li, L.; Chen, W.; Chen, Z.; Wang, Y.; Wang, S. Evaluation of heavy metal speciation distribution in soil and the accumulation characteristics in wild plants: A study on naturally aged abandoned farmland adjacent to tailings. Sci. Total Environ. 2024, 917, 170594. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, R.; Li, J.; Guo, Y.; Gao, T.; Guo, D.; Zhang, Q. Changes of heavy metal concentrations in farmland soils affected by non-ferrous metal smelting in China: A meta-analysis. Environ. Pollut. 2023, 336, 122442. [Google Scholar] [CrossRef] [PubMed]
- León-Cañedo, J.A.; Alarcón-Silvas, S.G.; Fierro-Sañudo, J.F.; Rodríguez-Montes de Oca, G.A.; Partida-Ruvalcaba, L.; Díaz-Valdés, T.; Páez-Osuna, F. Mercury and other trace metals in lettuce (Lactuca sativa) grown with two low-salinity shrimp effluents: Accumulation and human health risk assessment. Sci. Total Environ. 2019, 650, 2535–2544. [Google Scholar] [CrossRef] [PubMed]
- Suhani, I.; Sahab, S.; Srivastava, V.; Singh, R.P. Impact of cadmium pollution on food safety and human health. Curr. Opin. Toxicol. 2021, 27, 1–7. [Google Scholar] [CrossRef]
- Han, H.; Wu, X.; Yao, L.; Chen, Z. Heavy metal-immobilizing bacteria combined with calcium polypeptides reduced the uptake of Cd in wheat and shifted the rhizosphere bacterial communities. Environ. Pollut. 2020, 267, 115432. [Google Scholar] [CrossRef]
- Pan, W.; Lu, Q.; Xu, Q.-R.; Zhang, R.-R.; Li, H.-Y.; Yang, Y.-H.; Liu, H.-J.; Du, S.-T. Abscisic acid-generating bacteria can reduce Cd concentration in pakchoi grown in Cd-contaminated soil. Ecotoxicol. Environ. Saf. 2019, 177, 100–107. [Google Scholar] [CrossRef]
- Siedt, M.; Schäffer, A.; Smith, K.E.C.; Nabel, M.; Roß-Nickoll, M.; van Dongen, J.T. Comparing straw, compost, and biochar regarding their suitability as agricultural soil amendments to affect soil structure, nutrient leaching, microbial communities, and the fate of pesticides. Sci. Total Environ. 2021, 751, 141607. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Q.; Gao, C.; Lu, Y.; Sheng, Y.; Xiao, M.; Yun, Y.; Selvaraj, J.N.; Zhang, X.; Li, Y.; et al. Endophytic bacteria for Cd remediation in rice: Unraveling the Cd tolerance mechanisms of Cupriavidus metallidurans CML2. J. Hazard. Mater. 2024, 469, 133846. [Google Scholar] [CrossRef]
- Rawat, P.; Sharma, A.; Shankhdhar, D.; Shankhdhar, S.C. Improvement of phosphorus uptake, phosphorus use efficiency, and grain yield of upland rice (Oryza sativa L.) in response to phosphate-solubilizing bacteria blended with phosphorus fertilizer. Pedosphere 2022, 32, 752–763. [Google Scholar] [CrossRef]
- Disi, Z.A.; Attia, E.; Ahmad, M.I.; Zouari, N. Immobilization of heavy metals by microbially induced carbonate precipitation using hydrocarbon-degrading ureolytic bacteria. Biotechnol. Rep. 2022, 35, e00747. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, H.; Xin, C.; Yuan, Y.; Sun, Q.; Cao, C.; Chao, H.; Wu, T.; Zheng, S. Insight into adsorption of Pb(II) with wild resistant bacteria TJ6 immobilized on biochar composite: Roles of bacterial cell and biochar. Sep. Purif. Technol. 2024, 331, 125660. [Google Scholar] [CrossRef]
- Priyadarshanee, M.; Das, S. Spectra metrology for interaction of heavy metals with extracellular polymeric substances (EPS) of Pseudomonas aeruginosa OMCS-1 reveals static quenching and complexation dynamics of EPS with heavy metals. J. Hazard. Mater. 2024, 466, 133617. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.V.; Ranjit, P.; Priyanka, E.; Maddela, N.R.; Prasad, R. Bioremediation of heavy metals-contaminated sites by microbial extracellular polymeric substances—A critical view. Environ. Chem. Ecotoxicol. 2024, 6, 408–421. [Google Scholar] [CrossRef]
- Li, X.; Yu, Y.; Li, Y.; Yin, Z.; Liu, X.; Lian, B. Molecular mechanism of increasing extracellular polysaccharide production of Paenibacillus mucilaginosus K02 by adding mineral powders. Int. Biodeterior. Biodegrad. 2022, 167, 105340. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, L.; Han, Y.; Zhou, Z. Biosorption of Pb2+ and Zn2+ by Ca-alginate immobilized and free extracellular polysaccharides produced by Leuconostoc citreum B-2. Int. J. Biol. Macromol. 2021, 193, 2365–2373. [Google Scholar] [CrossRef]
- Lahiri, D.; Nag, M.; Dutta, B.; Dey, A.; Ray, R.R. Chapter 1—Bacterial extracellular polysaccharides in biofilm formation and function. In Application of Biofilms in Applied Microbiology; Shah, M.P., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 1–23. [Google Scholar]
- Ajao, V.; Nam, K.; Chatzopoulos, P.; Spruijt, E.; Bruning, H.; Rijnaarts, H.; Temmink, H. Regeneration and reuse of microbial extracellular polymers immobilised on a bed column for heavy metal recovery. Water Res. 2020, 171, 408–421. [Google Scholar] [CrossRef]
- Ali, M.; Cybulska, J.; Frąc, M.; Zdunek, A. Application of polysaccharides for the encapsulation of beneficial microorganisms for agricultural purposes: A review. Int. J. Biol. Macromol. 2023, 244, 125366. [Google Scholar] [CrossRef]
- Ghosh, P.K.; Maiti, T.K. Structure of Extracellular Polysaccharides (EPS) Produced by Rhizobia and their Functions in Legume–Bacteria Symbiosis:—A Review. Achiev. Life Sci. 2016, 10, 136–143. [Google Scholar] [CrossRef]
- Yan, C.; Ji, S.; Wu, R.; Li, M.; He, K.; Shi, H.; Wang, C.; Yang, H.; Guo, J.; Wu, J. Structural properties and biological activities of the extracellular polysaccharide of Bacillus subtilis LZ13-4. Int. J. Biol. Macromol. 2024, 259, 129176. [Google Scholar] [CrossRef]
- Mukherjee, P.; Mitra, A.; Roy, M. Halomonas Rhizobacteria of Avicennia marina of Indian Sundarbans Promote Rice Growth Under Saline and Heavy Metal Stresses Through Exopolysaccharide Production. Front. Microbiol. 2019, 10, 1207. [Google Scholar] [CrossRef] [PubMed]
- Mota, R.; Rossi, F.; Andrenelli, L.; Pereira, S.B.; De Philippis, R.; Tamagnini, P. Released polysaccharides (RPS) from Cyanothece sp. CCY 0110 as biosorbent for heavy metals bioremediation: Interactions between metals and RPS binding sites. Appl. Microbiol. Biotechnol. 2016, 100, 7765–7775. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xia, S.; Zhang, J.; Nguyen, B.T.; Zhang, Z. Insight into the influences of pH value on Pb(II) removal by the biopolymer extracted from activated sludge. Chem. Eng. J. 2017, 308, 1098–1104. [Google Scholar] [CrossRef]
- Shahid, M.; Altaf, M.; Ali, S.; Tyagi, A. Isolation and assessment of the beneficial effect of exopolysaccharide-producing PGPR in Triticum aestivum (L.) plants grown under NaCl and Cd -stressed conditions. Plant Physiol. Biochem. 2024, 215, 108973. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; He, L.-Y.; Wang, Q.; Sheng, X.-F. Synergistic effects of plant growth-promoting Neorhizobium huautlense T1-17 and immobilizers on the growth and heavy metal accumulation of edible tissues of hot pepper. J. Hazard. Mater. 2016, 312, 123–131. [Google Scholar] [CrossRef]
- Wang, Y.; Jing, Y.; Leng, F.; Wang, S.; Wang, F.; Zhuang, Y.; Liu, X.; Wang, X.; Ma, X. Establishment and Application of a Method for Rapid Determination of Total Sugar Content Based on Colorimetric Microplate. Sugar Tech 2016, 19, 424–431. [Google Scholar] [CrossRef]
- Teng, Z.; Shao, W.; Zhang, K.; Huo, Y.; Li, M. Characterization of phosphate solubilizing bacteria isolated from heavy metal contaminated soils and their potential for lead immobilization. J. Environ. Manag. 2019, 231, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.-y.; Sheng, X.-f.; Qian, M.; Wang, Q.-y. Isolation and characterization of a heavy metal-resistant Burkholderia sp. from heavy metal-contaminated paddy field soil and its potential in promoting plant growth and heavy metal accumulation in metal-polluted soil. Chemosphere 2008, 72, 157–164. [Google Scholar] [CrossRef]
- Rajkumar, M.; Nagendran, R.; Lee, K.J.; Lee, W.H.; Kim, S.Z. Influence of plant growth promoting bacteria and Cr6+ on the growth of Indian mustard. Chemosphere 2006, 62, 741–748. [Google Scholar] [CrossRef]
- Belimov, A.A.; Hontzeas, N.; Safronova, V.I.; Demchinskaya, S.V.; Piluzza, G.; Bullitta, S.; Glick, B.R. Cadmium-tolerant plant growth-promoting bacteria associated with the roots of Indian mustard (Brassica juncea L. Czern.). Soil Biol. Biochem. 2005, 37, 241–250. [Google Scholar] [CrossRef]
- Han, H.; Wang, Q.; He, L.-y.; Sheng, X.-f. Increased biomass and reduced rapeseed Cd accumulation of oilseed rape in the presence of Cd-immobilizing and polyamine-producing bacteria. J. Hazard. Mater. 2018, 353, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Yang, J.; Liu, W.; Yan, Y.; Wang, Y. Hydroxyapatite as a passivator for safe wheat production and its impacts on soil microbial communities in a Cd-contaminated alkaline soil. J. Hazard. Mater. 2021, 404, 124005. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Y.; Li, L.; Yang, Y.; Xu, C.; Luo, Y.; Wang, Y.; Liang, X.; Sun, J. Immobilization of Cd by mercapto-palygorskite in typical calcareous and acidic soil aggregates: Performance and differences. Chemosphere 2023, 323, 138223. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Cai, H.; Wang, X.; Hu, X.; Chen, Z.; Yao, L. Heavy metal-immobilizing bacteria increase the biomass and reduce the Cd and Pb uptake by pakchoi (Brassica chinensis L.) in heavy metal-contaminated soil. Ecotoxicol. Environ. Saf. 2020, 195, 110375. [Google Scholar] [CrossRef] [PubMed]
- Fisher, K.A.; Yarwood, S.A.; James, B.R. Soil urease activity and bacterial ureC gene copy numbers: Effect of pH. Geoderma 2017, 285, 1–8. [Google Scholar] [CrossRef]
- Yang, Y.; Wikieł, A.J.; Dall’Agnol, L.T.; Eloy, P.; Genet, M.J.; Moura, J.J.G.; Sand, W.; Dupont-Gillain, C.C.; Rouxhet, P.G. Proteins dominate in the surface layers formed on materials exposed to extracellular polymeric substances from bacterial cultures. Biofouling 2016, 32, 95–108. [Google Scholar] [CrossRef]
- Ye, S.; Ma, Z.; Liu, Z.; Liu, Y.; Zhang, M.; Wang, J. Effects of carbohydrate sources on biosorption properties of the novel exopolysaccharides produced by Arthrobacter ps-5. Carbohydr. Polym. 2014, 112, 615–621. [Google Scholar] [CrossRef]
- Lian, Z.; Yang, Z.; Song, W.; Sun, M.; Gan, Y.; Bai, X. Characteristics of EPS from Pseudomonas aeruginosa and Alcaligenes faecalis under Cd(II) stress: Changes in chemical components and adsorption performance. Environ. Sci. Pollut. Res. 2022, 29, 75883–75895. [Google Scholar] [CrossRef]
- Abd-El-Haleem, D. Alpha-glucan: A novel bacterial polysaccharide and its application as a biosorbent for heavy metals. J. Genet. Eng. Biotechnol. 2023, 21, 133. [Google Scholar] [CrossRef]
- Qu, C.; Duan, C.; Li, W.; Wu, X.; Liu, Z.; Feng, F.; Tang, X.; Chai, X.; Tang, C.-J. Understanding the slight inhibition of high As(III) stress on nitritation process: Insights from arsenic speciation and microbial community analyses. J. Hazard. Mater. 2022, 435, 128957. [Google Scholar] [CrossRef]
- Pan, X.; Zhang, S.; Li, T.; Ouyang, J.; Gong, G.; Wang, G.; Xu, X.; Pu, Y.; Long, L.; Jia, Y. Response of microbiomes with different abundances to removal of metal fractions by soil washing. Ecotoxicol. Environ. Saf. 2022, 242, 113862. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Zhang, P.; He, Y.; Zeng, F.; Xu, J.; He, L. Enantioselective effects of cyflumetofen on microbial community and related nitrogen cycle gene function in acid-soil. Sci. Total Environ. 2021, 771, 144831. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Rensing, C.; Han, D.; Xiao, K.-Q.; Dai, Y.; Tang, Z.; Liesack, W.; Peng, J.; Cui, Z.; Zhang, F.; et al. Genome-Resolved Metagenomics Reveals Distinct Phosphorus Acquisition Strategies between Soil Microbiomes. mSystems 2022, 7, e0110721. [Google Scholar] [CrossRef] [PubMed]
- Dedysh, S.N.; Beletsky, A.V.; Kulichevskaya, I.S.; Mardanov, A.V.; Ravin, N.V.; Cameron Thrash, J. Complete Genome Sequence of Paludibaculum fermentans P105T, a Facultatively Anaerobic Acidobacterium Capable of Dissimilatory Fe(III) Reduction. Microbiol. Resour. Announc. 2021, 10, e01313-20. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, L.; Sun, Y.; Xie, L.; Liu, S.; Li, M.; Yu, Q. Combined microbe-plant remediation of cadmium in saline-alkali soil assisted by fungal mycelium-derived biochar. Environ. Res. 2024, 240, 117424. [Google Scholar] [CrossRef]
- Shan, S.; Guo, Z.; Lei, P.; Li, Y.; Wang, Y.; Zhang, M.; Cheng, W.; Wu, S.; Wu, M.; Du, D. Increased biomass and reduced tissue cadmium accumulation in rice via indigenous Citrobacter sp. XT1-2-2 and its mechanisms. Sci. Total Environ. 2020, 708, 135224. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, C.; Wang, J.; Zou, S.; Long, X. Phosphorus solubilizing bacteria Bacillus thuringiensis and Pantoea ananatis simultaneously promote soil inorganic phosphate dissolution and soil Pb immobilization. Rhizosphere 2021, 20, 100448. [Google Scholar] [CrossRef]
- Huang, H.; Jia, Q.; Jing, W.; Dahms, H.-U.; Wang, L. Screening strains for microbial biosorption technology of cadmium. Chemosphere 2020, 251, 126428. [Google Scholar] [CrossRef]
- Yu, H.; Li, C.; Yan, J.; Ma, Y.; Zhou, X.; Yu, W.; Kan, H.; Meng, Q.; Xie, R.; Dong, P. A review on adsorption characteristics and influencing mechanism of heavy metals in farmland soil. RSC Adv. 2023, 13, 3505–3519. [Google Scholar] [CrossRef]
- Zhang, Y.-H.; Liu, C.-S.; Tian, Y.; Wang, J.; Xin, S.; Sheng, X. An eco-friendly photo-responsive hyaluronic acid-based supramolecular polysaccharide hybrid hydrogels for plant growth regulation and heavy metal ions adsorption. Int. J. Biol. Macromol. 2023, 242, 125194. [Google Scholar] [CrossRef]
- Sun, J.; He, X.; Le, Y.; Al-Tohamy, R.; Ali, S.S. Potential applications of extremophilic bacteria in the bioremediation of extreme environments contaminated with heavy metals. J. Environ. Manag. 2024, 352, 120081. [Google Scholar] [CrossRef] [PubMed]
- Karthik, C.; Oves, M.; Thangabalu, R.; Sharma, R.; Santhosh, S.B.; Indra Arulselvi, P. Cellulosimicrobium funkei-like enhances the growth of Phaseolus vulgaris by modulating oxidative damage under Chromium(VI) toxicity. J. Adv. Res. 2016, 7, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Li, F.; Wu, C.; Yu, R.; Wu, X.; Shen, L.; Liu, Y.; Qiu, G.; Li, J. Role of extracellular polymeric substance (EPS) in toxicity response of soil bacteria Bacillus sp. S3 to multiple heavy metals. Bioprocess Biosyst. Eng. 2019, 43, 153–167. [Google Scholar] [CrossRef] [PubMed]
- K, K.R.; Sardar, U.R.; Bhargavi, E.; Devi, I.; Bhunia, B.; Tiwari, O.N. Advances in exopolysaccharides based bioremediation of heavy metals in soil and water: A critical review. Carbohydr. Polym. 2018, 199, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Musarurwa, H.; Tavengwa, N.T. Application of carboxymethyl polysaccharides as bio-sorbents for the sequestration of heavy metals in aquatic environments. Carbohydr. Polym. 2020, 237, 116142. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Li, Y.; Noguera, D.R.; Zhao, N.; Song, Y.; Ding, J.; Zhao, Q.; Cui, F. Adsorption of Cu2+ and Zn2+ by extracellular polymeric substances (EPS) in different sludges: Effect of EPS fractional polarity on binding mechanism. J. Hazard. Mater. 2017, 321, 473–483. [Google Scholar] [CrossRef]
- Iyer, A.; Mody, K.; Jha, B. Accumulation of hexavalent chromium by an exopolysaccharide producing marine Enterobacter cloaceae. Mar. Pollut. Bull. 2004, 49, 974–977. [Google Scholar] [CrossRef]
- Cui, D.; Tan, C.; Deng, H.; Gu, X.; Pi, S.; Chen, T.; Zhou, L.; Li, A. Biosorption Mechanism of Aqueous Pb2+, Cd2+, and Ni2+ Ions on Extracellular Polymeric Substances (EPS). Archaea 2020, 2020, 8891543. [Google Scholar] [CrossRef]
- Vimalnath, S.; Subramanian, S. Studies on the biosorption of Pb(II) ions from aqueous solution using extracellular polymeric substances (EPS) of Pseudomonas aeruginosa. Colloids Surf. B Biointerfaces 2018, 172, 60–67. [Google Scholar] [CrossRef]
- Zainab, N.; Amna; Din, B.U.; Javed, M.T.; Afridi, M.S.; Mukhtar, T.; Kamran, M.A.; Ul Ain, O.; Khan, A.A.; Ali, J.; et al. Deciphering metal toxicity responses of flax (Linum usitatissimum L.) with exopolysaccharide and ACC-deaminase producing bacteria in industrially contaminated soils. Plant Physiol. Biochem. 2020, 152, 90–99. [Google Scholar] [CrossRef]
- Al-Dhabi, N.A.; Esmail, G.A.; Valan Arasu, M. Sustainable conversion of palm juice wastewater into extracellular polysaccharides for absorption of heavy metals from Saudi Arabian wastewater. J. Clean. Prod. 2020, 277, 124252. [Google Scholar] [CrossRef]
- Morillo Pérez, J.A.; García-Ribera, R.; Quesada, T.; Aguilera, M.; Ramos-Cormenzana, A.; Monteoliva-Sánchez, M. Biosorption of heavy metals by the exopolysaccharide produced by Paenibacillus jamilae. World J. Microbiol. Biotechnol. 2008, 24, 2699–2704. [Google Scholar] [CrossRef]
- Vandana; Das, S. Genetic regulation, biosynthesis and applications of extracellular polysaccharides of the biofilm matrix of bacteria. Carbohydr. Polym. 2022, 291, 119536. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Ma, L.; Cheng, H.; Liu, Y.; Xu, D.; Li, B.; Liu, F.; Ren, Y.; Liu, Z.; Zhao, C.; et al. Characterization of the particle size fraction associated heavy metals in tropical arable soils from Hainan Island, China. J. Geochem. Explor. 2014, 139, 109–114. [Google Scholar] [CrossRef]
- Huang, B.; Yuan, Z.; Li, D.; Zheng, M.; Nie, X.; Liao, Y. Effects of soil particle size on the adsorption, distribution, and migration behaviors of heavy metal(loid)s in soil: A review. Environ. Sci. Process. Impacts 2020, 22, 1596–1615. [Google Scholar] [CrossRef]
- Song, J.; Brookes, P.C.; Shan, S.; Xu, J.; Liu, X. Effects of remediation agents on microbial community structure and function in soil aggregates contaminated with heavy metals. Geoderma 2022, 425, 116030. [Google Scholar] [CrossRef]
- Wu, N.; Pan, H.-X.; Qiu, D.; Zhang, Y.-M. Feasibility of EPS-producing bacterial inoculation to speed up the sand aggregation in the Gurbantunggut Desert, Northwestern China. J. Basic Microbiol. 2014, 54, 1378–1386. [Google Scholar] [CrossRef]
- Zhao, R.; Kuzyakov, Y.; Zhang, H.; Wang, Z.; Li, T.; Shao, L.; Jiang, L.; Wang, R.; Li, M.; Jianxin Sun, O.; et al. Labile carbon inputs offset nitrogen-induced soil aggregate destabilization via enhanced growth of saprophytic fungi in a meadow steppe. Geoderma 2024, 443, 116841. [Google Scholar] [CrossRef]
- Mohite, B.V.; Koli, S.H.; Patil, S.V. Heavy Metal Stress and Its Consequences on Exopolysaccharide (EPS)-Producing Pantoea agglomerans. Appl. Biochem. Biotechnol. 2018, 186, 199–216. [Google Scholar] [CrossRef]
- Zhang, K.; Chen, S.; Ping, Y.; Song, F.; Fan, X.; Wang, L.; Zang, C. Coinoculation of arbuscular mycorrhizal fungi and rhizobia stimulates atrazine dissipation by changing the atrazine-degrading bacterial community at the soil aggregate scale. Environ. Pollut. 2024, 345, 123507. [Google Scholar] [CrossRef]
- Fontana, M.B.; Novelli, L.E.; Sterren, M.A.; Uhrich, W.G.; Rondán, G.A.; Barbagelata, P.A.; Benintende, S.M. Cover crop benefit bacteria and increase aggregate-associate soil C and N storage. Geoderma Reg. 2024, 36, e00743. [Google Scholar] [CrossRef]
- Wang, X.; Huang, P.; Ma, M.; Shan, K.; Wu, S. Effects of riparian pioneer plants on soil aggregate stability: Roles of root traits and rhizosphere microorganisms. Sci. Total Environ. 2024, 940, 173584. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, E.; Piazza, G.; Helgason, T.; Ercoli, L. Microbiome structure and interconnection in soil aggregates across conservation and conventional agricultural practices allow to identify main prokaryotic and fungal taxa related to soil functioning. Soil Biol. Biochem. 2022, 175, 108833. [Google Scholar] [CrossRef]
- Sarder, M.P.; Kamruzzaman, M.; Siddique, M.A.B.; Halder, M. Stability and heavy metals accumulation of soil aggregates under different land uses in the southwest coastal Bangladesh. Heliyon 2024, 10, e37806. [Google Scholar] [CrossRef]
- Mann, C.; Lynch, D.; Fillmore, S.; Mills, A. Relationships between field management, soil health, and microbial community composition. Appl. Soil Ecol. 2019, 144, 12–21. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Wang, K.; Liu, X.; Yao, L.; Chen, Z.; Han, H. Exopolysaccharide-Producing Bacteria Regulate Soil Aggregates and Bacterial Communities to Inhibit the Uptake of Cadmium and Lead by Lettuce. Microorganisms 2024, 12, 2112. https://doi.org/10.3390/microorganisms12112112
Zhang H, Wang K, Liu X, Yao L, Chen Z, Han H. Exopolysaccharide-Producing Bacteria Regulate Soil Aggregates and Bacterial Communities to Inhibit the Uptake of Cadmium and Lead by Lettuce. Microorganisms. 2024; 12(11):2112. https://doi.org/10.3390/microorganisms12112112
Chicago/Turabian StyleZhang, Heyun, Ke Wang, Xinru Liu, Lunguang Yao, Zhaojin Chen, and Hui Han. 2024. "Exopolysaccharide-Producing Bacteria Regulate Soil Aggregates and Bacterial Communities to Inhibit the Uptake of Cadmium and Lead by Lettuce" Microorganisms 12, no. 11: 2112. https://doi.org/10.3390/microorganisms12112112
APA StyleZhang, H., Wang, K., Liu, X., Yao, L., Chen, Z., & Han, H. (2024). Exopolysaccharide-Producing Bacteria Regulate Soil Aggregates and Bacterial Communities to Inhibit the Uptake of Cadmium and Lead by Lettuce. Microorganisms, 12(11), 2112. https://doi.org/10.3390/microorganisms12112112