
The Presence of Four Pathogenic Oral Bacterial Species in Six Wild Snake Species from Southern Taiwan: Associated Factors

Abstract
1. Introduction
2. Materials and Methods
2.1. Snake Collection
2.2. Oropharyngeal Sample Collection
2.3. DNA Extraction, PCR Amplification, and Electrophoresis
2.3.1. E. faecalis
2.3.2. M. morganii
2.3.3. P. aeruginosa
2.3.4. A. hydrophila
2.3.5. Positive and Negative Controls
2.3.6. Agarose Gel Electrophoresis
2.4. Statistical Methods
3. Results
3.1. Sampled Snake Quantity by Sex
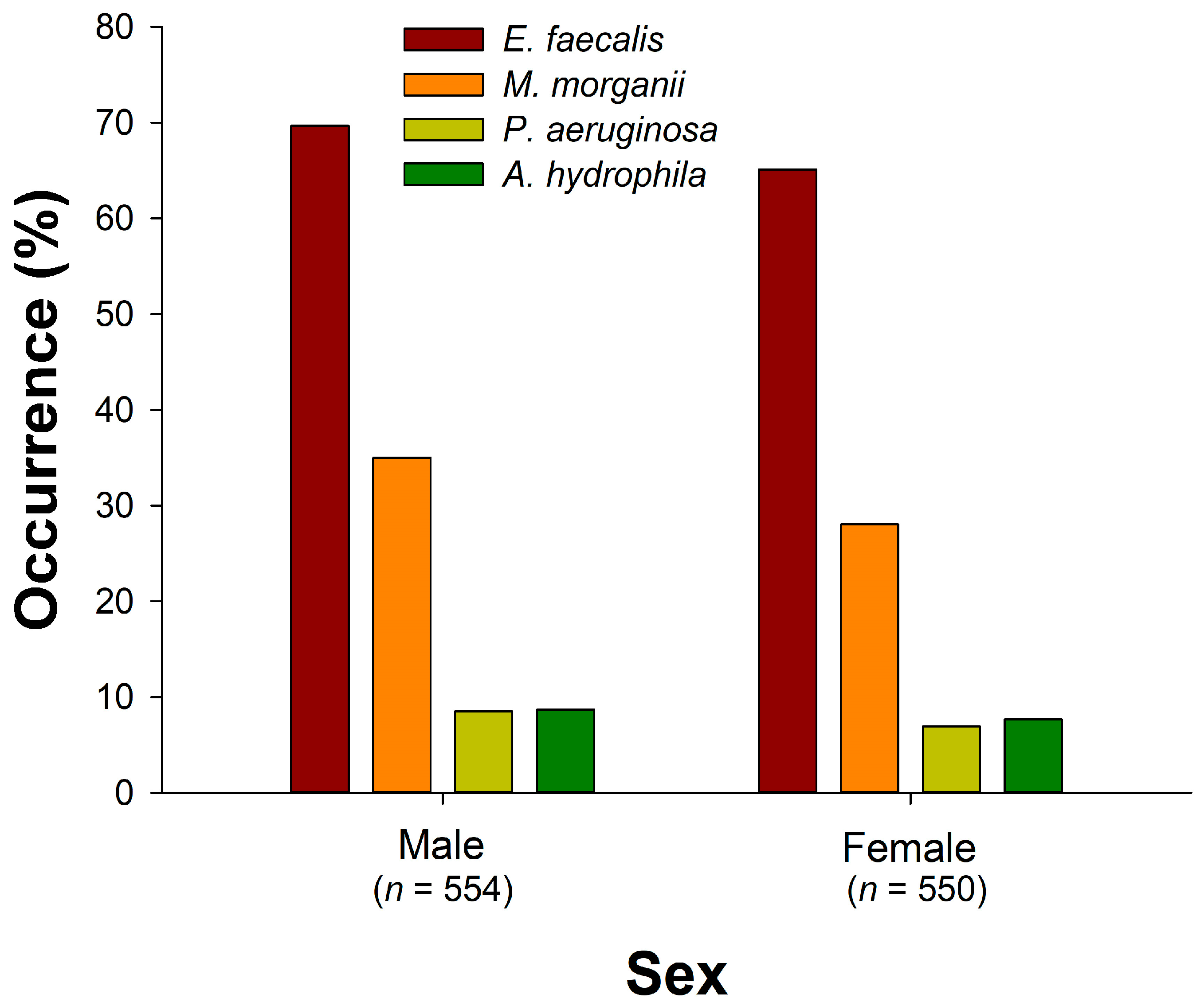
3.2. Comparison of Bacterial Occurrence by Season, Snake Species, and Sex
3.2.1. E. faecalis
3.2.2. M. morganii
3.2.3. P. aeruginosa
3.2.4. A. hydrophila
3.3. Association Factors of Bacterial Occurrence
3.3.1. E. faecalis
3.3.2. M. morganii
3.3.3. P. aeruginosa
3.3.4. A. hydrophila
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ulima, A.T.; Fawzy, A. Complication in neglected non-venomous snake bite in rural area; what we need to anticipate. Inter. J. Med. Sci. Clin. Res. Stud. 2022, 2, 1038–1043. [Google Scholar] [CrossRef]
- Sminkey, L. World report on child injury prevention. Inj. Prev. 2008, 14, 69. [Google Scholar] [CrossRef]
- Chippaux, J.P. Snake-bites: Appraisal of the global situation. Bull. World Health Organ. 1998, 76, 515–524. [Google Scholar] [PubMed]
- Noutsos, T.; Currie, B.J.; Wijewickrama, E.S.; Isbister, G.K. Snakebite associated thrombotic microangiopathy and recommendations for clinical practice. Toxins 2022, 14, 57. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Theakston, R.D.G.; Warrell, D.A. Confronting the neglected problem of snake bite envenoming: The need for a global partnership. PLoS Med. 2006, 3, e150. [Google Scholar] [CrossRef] [PubMed]
- Redewad, N.; Bhaisare, S.; Bansod, Y.; Hire, R. Management and outcome study of snake bite cases in central India. Sch. J. Appl. Med. Sci. 2014, 2, 435–441. [Google Scholar]
- Suchithra, N.; Pappachan, J.; Sujathan, P. Snakebite envenoming in Kerala, South India: Clinical profile and factors involved in adverse outcomes. Emerg. Med. J. 2008, 25, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, I.K.; Dixit, P.P.; Pawade, B.S.; Potnis-Lele, M.; Kurhe, B.P. Assessment of cultivable oral bacterial flora from important venomous snakes of India and their antibiotic susceptibilities. Curr. Microbiol. 2017, 74, 1278–1286. [Google Scholar] [CrossRef]
- Lam, K.K.; Crow, P.; Ng, K.H.; Shek, K.C.; Fung, H.T.; Ades, G.; Grioni, A.; Tan, K.S.; Yip, K.T.; Lung, D.C.; et al. A cross-sectional survey of snake oral bacterial flora from Hong Kong, SAR, China. Emerg. Med. J. 2011, 28, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Padhi, L.; Panda, S.K.; Mohapatra, P.P.; Sahoo, G. Antibiotic susceptibility of cultivable aerobic microbiota from the oral cavity of Echis carinatus from Odisha (India). Microb. Pathog. 2020, 143, 104121. [Google Scholar] [CrossRef]
- Muhammad, H.; Sadiq, H.; Nashabaru, I.; Sanda, A.; Abdullahi, F.; Bello, U.; Habib, A.G. Multidrug-resistant infection following snakebite envenoming: A case report. Nigerian J. Basic Clin. Sci. 2018, 15, 161. [Google Scholar] [CrossRef]
- Quiroga, M.; Avila-Agüero, M.; Faingezicht, I. Abscess secondary to facial snakebite. J. Venom. Anim. Toxins 2000, 6, 261–270. [Google Scholar] [CrossRef]
- Habib, A. Tetanus complicating snakebite in northern Nigeria: Clinical presentation and public health implications. Acta Trop. 2003, 85, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Blaylock, R. The identification and syndromic management of snakebite in South Africa. S. Afr. Fam. Pract. 2005, 47, 46–53. [Google Scholar] [CrossRef]
- Goldstein, E.; Agyare, E.; Vagvolgyi, A.; Halpern, M. Aerobic bacterial oral flora of garter snakes: Development of normal flora and pathogenic potential for snakes and humans. J. Clin. Microbiol. 1981, 13, 954–956. [Google Scholar] [CrossRef]
- Lin, W.H.; Tsai, T.S. Comparisons of the oral microbiota from seven species of wild venomous snakes in Taiwan using the high-throughput amplicon sequencing of the rull-length 16S rRNA gene. Biology 2023, 12, 1206. [Google Scholar] [CrossRef]
- Abubakar, S.; Habib, A.; Mathew, J. Amputation and disability following snakebite in Nigeria. Trop. Doct. 2010, 40, 114–116. [Google Scholar] [CrossRef]
- Gold, B.S.; Dart, R.C.; Barish, R.A. Bites of venomous snakes. N. Engl. J. Med. 2002, 347, 347–356. [Google Scholar] [CrossRef]
- Snow, R.W.; Bronzan, R.; Roques, T.; Nyamawi, C.; Murphy, S.; Marsh, K. The prevalence and morbidity of snake bite and treatment-seeking behaviour among a rural Kenyan population. Ann. Trop. Med. Parasitol. 1994, 88, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Auerbach, P.S.; Cushing, T.A.; Harris, N.S. Auerbach’s Wilderness Medicine; Elsevier Health Sciences: London, UK, 2016. [Google Scholar]
- Blaylock, R. Normal oral bacterial flora from some southern African snakes. Onderstepoort J. Vet. Res. 2001, 68, 175–182. [Google Scholar]
- Sadeghi, M.; Barazandeh, M.; Zakariaei, Z.; Davoodi, L.; Tabaripour, R.; Fakhar, M.; Zakariaei, A. Massive cutaneous complications due to snakebite: A case report and literature review. Clin. Case Rep. 2021, 9, e04129. [Google Scholar] [CrossRef]
- Chen, C.-M.; Wu, K.-G.; Chen, C.-J.; Wang, C.-M. Bacterial infection in association with snakebite: A 10-year experience in a northern Taiwan medical center. J. Microbiol. Immunol. Infect. 2011, 44, 456–460. [Google Scholar] [CrossRef]
- Theakston, R.; Phillips, R.; Looareesuwan, S.; Echeverria, P.; Makin, T.; Warrell, D. Bacteriological studies of the venom and mouth cavities of wild Malayan pit vipers (Calloselasma rhodostoma) in southern Thailand. Trans. R. Soc. Trop. Med. Hyg. 1990, 84, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Artavia-Leon, A.; Romero-Guerrero, A.; Sancho-Blanco, C.; Rojas, N.; Umana-Castro, R. Diversity of aerobic bacteria isolated from oral and cloacal cavities from free-living snakes species in Costa Rica rainforest. Int. Sch. Res. Notices 2017, 2017, 8934285. [Google Scholar] [CrossRef] [PubMed]
- Jho, Y.-S.; Park, D.-H.; Lee, J.-H.; Cha, S.-Y.; Han, J.S. Identification of bacteria from the oral cavity and cloaca of snakes imported from Vietnam. Lab. Anim. Res. 2011, 27, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, M.S.; Ferretti, J.J. The thin line between gut commensal and pathogen. Science 2003, 299, 1999–2002. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.Y.; Weng, L.L.; Tseng, S.Y.; Huang, C.C.; Cheng, C.C.; Mao, Y.C.; Tung, K.C. Colistin resistance of Pseudomonas aeruginosa isolated from snakes in Taiwan. Can. J. Infect. Dis. Med. Microbiol. 2017, 2017, 7058396. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [PubMed]
- Mehler, S.J.; Bennett, R.A. Upper alimentary tract disease. In Reptile Medicine and Surgery, 2nd ed.; Mader, D.R., Ed.; Saunders/Elsevier: St. Louis, MO, USA, 2006; pp. 924–930. [Google Scholar]
- Hedley, J. Anatomy and disorders of the oral cavity of reptiles and amphibians. Vet. Clin. N. Am. Exot. Anim. Pract. 2016, 19, 689–706. [Google Scholar] [CrossRef] [PubMed]
- Mehler, S.J.; Bennett, R.A. Oral, dental, and beak disorders of reptiles. Vet. Clin. N. Am. Exot. Anim. Pract. 2003, 6, 477–503. [Google Scholar] [CrossRef]
- Abba, Y.; Ilyasu, Y.M.; Noordin, M.M. Isolation and identification of bacterial populations of zoonotic importance from captive non-venomous snakes in Malaysia. Microb. Pathog. 2017, 108, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, J. Selected infectious diseases of wild reptiles and amphibians. J. Exot. Pet Med. 2006, 15, 18–24. [Google Scholar] [CrossRef]
- Mitchell, M.; Tully, T.N. Manual of Exotic Pet Practice; Elsevier Health Sciences: Saint Louis, MO, USA, 2008. [Google Scholar]
- Lowel, A. Diagnostics procedures: Hematology. In The Biology, Husbandry and Health Care of Reptiles; TFH Publications, Inc.: Neptune, NJ, USA, 1998; Volume III. [Google Scholar]
- Baron, S.E. Medical Microbiology, 4th ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Orozova, P.; Sirakov, I.; Petkov, I.; Crumlish, M.; Austin, B. Recovery of Aeromonas hydrophila associated with bacteraemia in captive snakes. FEMS Microbiol. Lett. 2012, 334, 22–26. [Google Scholar] [CrossRef]
- Hedley, J.; Whitehead, M.L.; Munns, C.; Pellett, S.; Abou-Zahr, T.; Calvo, D.C.; Wissink-Argilaga, N. Antibiotic stewardship for reptiles. J. Small Anim. Pract. 2021, 62, 829–839. [Google Scholar] [CrossRef]
- Fonseca, M.G.; Moreira, W.; Cunha, K.; Ribeiro, A.; Almeida, M. Oral microbiota of Brazilian captive snakes. J. Venom. Anim. Toxins Incl. Trop. Dis. 2009, 15, 54–60. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Egas, C.; Nunes, O.C.; Manaia, C.M. Culture-dependent and culture-independent diversity surveys target different bacteria: A case study in a freshwater sample. Antonie Van. Leeuwenhoek 2011, 100, 245–257. [Google Scholar] [CrossRef]
- Amann, R.I.; Ludwig, W.; Schleifer, K.-H. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar] [CrossRef]
- Vartoukian, S.R.; Palmer, R.M.; Wade, W.G. Strategies for culture of ‘unculturable’ bacteria. FEMS Microbiol. Lett. 2010, 309, 1–7. [Google Scholar] [CrossRef]
- Shek, K.; Tsui, K.; Lam, K.; Crow, P.; Ng, K.; Ades, G.; Yip, K.; Grioni, A.; Tan, K.; Lung, D.C. Oral bacterial flora of the Chinese cobra (Naja atra) and bamboo pit viper (Trimeresurus albolabris) in Hong Kong SAR, China. Hong Kong Med. J. 2009, 15, 183–190. [Google Scholar]
- Krishnankutty, S.P.; Muraleedharan, M.; Perumal, R.C.; Michael, S.; Benny, J.; Balan, B.; Kumar, P.; Manazhi, J.; Kumar, B.D.; Santhosh, S.; et al. Next-generation sequencing analysis reveals high bacterial diversity in wild venomous and non-venomous snakes from India. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 41. [Google Scholar] [CrossRef]
- Huang, L.-W.; Wang, J.-D.; Huang, J.-A.; Hu, S.-Y.; Wang, L.-M.; Tsan, Y.-T. Wound infections secondary to snakebite in central Taiwan. J. Venom. Anim. Toxins Incl. Trop. Dis. 2012, 18, 272–276. [Google Scholar] [CrossRef]
- Mao, Y.-C.; Liu, P.-Y.; Chiang, L.-C.; Lai, C.-S.; Lai, K.-L.; Ho, C.-H.; Wang, T.-H.; Yang, C.-C. Naja atra snakebite in Taiwan. Clin. Toxicol. 2018, 56, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Chaou, C.H.; Tseng, C.Y. An investigation of snakebite antivenom usage in Taiwan. J. Formos. Med. Assoc. 2016, 115, 672–677. [Google Scholar] [CrossRef] [PubMed]
- Chuang, P.C.; Lin, W.H.; Chen, Y.C.; Chien, C.C.; Chiu, I.M.; Tsai, T.S. Oral bacteria and their antibiotic susceptibilities in Taiwanese venomous snakes. Microorganisms 2022, 10, 951. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.C.; Chuang, H.N.; Shih, C.H.; Hsieh, H.H.; Jiang, Y.H.; Chiang, L.C.; Lin, W.L.; Hsiao, T.H.; Liu, P.Y. An investigation of conventional microbial culture for the Naja atra bite wound, and the comparison between culture-based 16S Sanger sequencing and 16S metagenomics of the snake oropharyngeal bacterial microbiota. PLoS Negl. Trop. Dis. 2021, 15, e0009331. [Google Scholar] [CrossRef] [PubMed]
- McKiernan, H.E.; Danielson, P.B. Molecular diagnostic applications in forensic science. In Molecular Diagnostics; Elsevier: London, UK, 2017; pp. 371–394. [Google Scholar]
- Rocas, I.N.; Siqueira, J.F., Jr.; Santos, K.R. Association of Enterococcus faecalis with different forms of periradicular diseases. J. Endod. 2004, 30, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, C.; Ricci, G.; Borgo, F.; Fortina, M.G. Species-specific DNA probe and development of a quantitative PCR assay for the detection of Morganella morganii. Lett. Appl. Microbiol. 2012, 54, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Altaai, M.E.; Aziz, I.H.; Marhoon, A.A. Identification Pseudomonas aeruginosa by 16s rRNA gene for differentiation from other Pseudomonas species that isolated from patients and environment. Baghdad Sci. J. 2014, 11, 1028–1034. [Google Scholar]
- Chu, W.H.; Lu, C.P. Multiplex PCR assay for the detection of pathogenic Aeromonas hydrophila. J. Fish Dis. 2005, 28, 437–441. [Google Scholar] [CrossRef]
- Micek, S.T.; Lloyd, A.E.; Ritchie, D.J.; Reichley, R.M.; Fraser, V.J.; Kollef, M.H. Pseudomonas aeruginosa bloodstream infection: Importance of appropriate initial antimicrobial treatment. Antimicrob. Agents Chemother. 2005, 49, 1306–1311. [Google Scholar] [CrossRef]
- Peig, J.; Green, A.J. New perspectives for estimating body condition from mass/length data: The scaled mass index as an alternative method. Oikos 2009, 118, 1883–1891. [Google Scholar] [CrossRef]
- Peig, J.; Green, A.J. The paradigm of body condition: A critical reappraisal of current methods based on mass and length. Funct. Ecol. 2010, 24, 1323–1332. [Google Scholar] [CrossRef]
- Goss, M.; Swain, D.L.; Abatzoglou, J.T.; Sarhadi, A.; Kolden, C.A.; Williams, A.P.; Diffenbaugh, N.S. Climate change is increasing the likelihood of extreme autumn wildfire conditions across California. Environ. Res. Lett. 2020, 15, 094016. [Google Scholar] [CrossRef]
- Sperry, J.H.; Peak, R.G.; Cimprich, D.A.; Weatherhead, P.J. Snake activity affects seasonal variation in nest predation risk for birds. J. Avian Biol. 2008, 39, 379–383. [Google Scholar] [CrossRef]
- Jones-Dias, D.; Clemente, L.; Moura, I.B.; Sampaio, D.A.; Albuquerque, T.; Vieira, L.; Manageiro, V.; Caniça, M. Draft genomic analysis of an avian multidrug resistant Morganella morganii isolate carrying qnrD1. Front. Microbiol. 2016, 7, 1660. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Chen, Y.C.; Goh, Z.N.L.; Seak, C.K.; Seak, J.C.; Shi-Ying, G.; Seak, C.J.; Spot, I. Wound infections of snakebites from the venomous Protobothrops mucrosquamatus and Viridovipera stejnegeri in Taiwan: Bacteriology, antibiotic susceptibility, and predicting the need for antibiotics-A BITE study. Toxins 2020, 12, 575. [Google Scholar] [CrossRef] [PubMed]
- Yeh, H.; Gao, S.Y.; Lin, C.C. Wound Infections from Taiwan cobra (Naja atra) bites: Determining bacteriology, antibiotic susceptibility, and the use of antibiotics-A cobra BITE Study. Toxins 2021, 13, 183. [Google Scholar] [CrossRef]
- Samy, R.P.; Kandasamy, M.; Gopalakrishnakone, P.; Stiles, B.G.; Rowan, E.G.; Becker, D.; Shanmugam, M.K.; Sethi, G.; Chow, V.T. Wound healing activity and mechanisms of action of an antibacterial protein from the venom of the eastern diamondback rattlesnake (Crotalus adamanteus). PLoS ONE 2014, 9, e80199. [Google Scholar] [CrossRef] [PubMed]
- Vargas, L.J.; Londoño, M.; Quintana, J.C.; Rua, C.; Segura, C.; Lomonte, B.; Núñez, V. An acidic phospholipase A2 with antibacterial activity from Porthidium nasutum snake venom. Comp. Biochem. Physiol. B-Biochem. Mol. Biol. 2012, 161, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Ciscotto, P.; De Avila, R.M.; Coelho, E.; Oliveira, J.; Diniz, C.; Farías, L.; De Carvalho, M.; Maria, W.; Sanchez, E.; Borges, A. Antigenic, microbicidal and antiparasitic properties of an L-amino acid oxidase isolated from Bothrops jararaca snake venom. Toxicon 2009, 53, 330–341. [Google Scholar] [CrossRef] [PubMed]
- Schulte, L.; Damm, M.; Avella, I.; Uhrig, L.; Erkoc, P.; Schiffmann, S.; Furst, R.; Timm, T.; Lochnit, G.; Vilcinskas, A.; et al. Venomics of the milos viper (Macrovipera schweizeri) unveils patterns of venom composition and exochemistry across blunt-nosed viper venoms. Front. Mol. Biosci. 2023, 10, 1254058. [Google Scholar] [CrossRef]
- Talan, D.A.; Citron, D.M.; Overturf, G.D.; Singer, B.; Froman, P.; Goldstein, E.J. Antibacterial activity of crotalid venoms against oral snake flora and other clinical bacteria. J. Infect. Dis. 1991, 164, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Cushing, A.; Pinborough, M.; Stanford, M. Review of bacterial and fungal culture and sensitivity results from reptilian samples submitted to a UK laboratory. Vet. Rec. 2011, 169, 390. [Google Scholar] [CrossRef] [PubMed]
- Al-Asmari, A.K.; Abbasmanthiri, R.; Osman, N.M.A.; Siddiqui, Y.; Al-Bannah, F.A.; Al-Rawi, A.M.; Al-Asmari, S.A. Assessment of the antimicrobial activity of few Saudi Arabian snake venoms. Open Microbiol. J. 2015, 9, 18. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.C.; Liu, P.Y.; Hung, D.Z.; Lai, W.C.; Huang, S.T.; Hung, Y.M.; Yang, C.C. Bacteriology of Naja atra snakebite wound and its implications for antibiotic therapy. Am. J. Trop. Med. Hyg. 2016, 94, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Morandi, S.; Brasca, M.; Alfieri, P.; Lodi, R.; Tamburini, A. Influence of pH and temperature on the growth of Enterococcus faecium and Enterococcus faecalis. Lait 2005, 85, 181–192. [Google Scholar] [CrossRef]
- Zancolli, G.; Mahsberg, D.; Sickel, W.; Keller, A. Reptiles as reservoirs of bacterial infections: Real threat or methodological bias? Microb. Ecol. 2015, 70, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Koyuncu, S.; Fırat, O. Morganella morganii osteomyelitis complicated by secondary septic knee arthritis: A case report. Acta Orthop. Traumatol. Turc. 2012, 46, 464–467. [Google Scholar] [CrossRef]
- Samonis, G.; Anatoliotaki, M.; Apostolakou, H.; Souglakos, J.; Georgoulias, V. Fatal septicemia and meningitis due to Morganella morganii in a patient with Hodgkin’s disease. Scand. J. Infect. Dis. 2001, 33, 553–555. [Google Scholar] [CrossRef]
- Emerson, J.D.; DeSantis, D.L.; Mata-Silva, V.; Wagler, A.E.; Johnson, J.D. Movement, home range size, and habitat use of Eastern Black-tailed Rattlesnakes (Crotalus ornatus) in the northern Chihuahuan Desert. Herpetologica 2022, 78, 110–118. [Google Scholar] [CrossRef]
- Natusch, D.; Lyons, J.; Shine, R. Spatial ecology, activity patterns, and habitat use by giant pythons (Simalia amethistina) in tropical Australia. Sci. Rep. 2022, 12, 5274. [Google Scholar] [CrossRef] [PubMed]
- Crone, S.; Vives-Flórez, M.; Kvich, L.; Saunders, A.M.; Malone, M.; Nicolaisen, M.H.; Martínez-García, E.; Rojas-Acosta, C.; Catalina, M.G.P.; Calum, H. The environmental occurrence of Pseudomonas aeruginosa. APMIS 2020, 128, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Johansen, H.; Høiby, N. Seasonal onset of initial colonisation and chronic infection with Pseudomonas aeruginosa in patients with cystic fibrosis in Denmark. Thorax 1992, 47, 109–111. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.-Y.; Shi, Z.-Y.; Shyu, C.-L.; Wu, Z.-Y.; Lai, K.-L.; Chang, C.-Y.; Chen, Y.-J.; Huang, J.-A.; Mao, Y.-C.; Tung, K.-C. Cobra bite wound infection caused by Shewanella algae. Int. J. Infect. Dis. 2014, 20, 11–12. [Google Scholar] [CrossRef] [PubMed]
- Aundhakar, S.; Bhardiya, A.; Panpalia, N.; Bhalla, G.; Mane, M. Snake bite complicated by bacterial meningitis. Int. J. Med. Toxicol. Forens. Med. 2013, 3, 34–37. [Google Scholar] [CrossRef]
- Garg, A.; Sujatha, S.; Garg, J.; Acharya, N.S.; Parija, S.C. Wound infections secondary to snakebite. J. Infect. Dev. Ctries. 2009, 3, 221–223. [Google Scholar] [CrossRef]
- Visser, L.; Kyei-Faried, S.; Belcher, D. Protocol and monitoring to improve snake bite outcomes in rural Ghana. Trans. R. Soc. Trop. Med. Hyg. 2004, 98, 278–283. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | B. kraepelini | B. multicinctus | N. atra | E. t. freisi | P. mucrosquamatus | T. stejnegeri | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Min | Max | Min | Max | Min | Max | Min | Max | Min | Max | |
| Snout-vent length (cm) | 29.8 | 120 | 21.3 | 126 | 29.5 | 135.2 | 32.9 | 192 | 21.3 | 92.6 | 21.1 | 73 |
| Scaled mass index a | 26.18 | 819.43 | 44.66 | 7315.94 | 30.88 | 11,762.84 | 12.61 | 10,798.92 | 59.58 | 878.69 | 67.51 | 545.94 |
| Artificial ground area (m2) b | 5128.98 | 31,415.96 | 2501.6 | 31,415.96 | 6943.17 | 31,415.96 | 4838.05 | 31,415.96 | 2557.07 | 31,415.96 | 4333.86 | 31,415.96 |
| Temperature (°C) | 19.1 | 31.3 | 18.6 | 31.3 | 17.7 | 31.6 | 15 | 31.5 | 15.3 | 30.3 | 15.8 | 31.3 |
| Rainfall (mm) | 0 | 13.3 | 0 | 12.6 | 0 | 15.7 | 0 | 14.3 | 0 | 7.8 | 0 | 18 |
| Elevation (m) | 4 | 430 | 3 | 320 | 1 | 320 | 3 | 252 | 7 | 248 | 2 | 283 |
| Duration (days) c | 1 | 26 | 0 | 22 | 2 | 25 | 0 | 26 | 2 | 27 | 0 | 21 |
| Factor | B | S.E. | Wald | d.f. | p | Exp(B) | 95% C.I. | |
|---|---|---|---|---|---|---|---|---|
| Lower | Upper | |||||||
| Snake species (B. kraepelini) | 129.590 | 5 | <0.001 | |||||
| N. atra | −1.966 | 0.500 | 15.436 | 1 | <0.001 | 0.140 | 0.052 | 0.373 |
| B. multicinctus | −1.092 | 0.528 | 4.283 | 1 | 0.038 | 0.335 | 0.119 | 0.944 |
| T. stejnegeri | −3.284 | 0.508 | 41.800 | 1 | <0.001 | 0.037 | 0.014 | 0.101 |
| P. mucrosquamatus | −2.582 | 0.522 | 24.480 | 1 | <0.001 | 0.076 | 0.027 | 0.210 |
| E. t. friesi | −0.500 | 0.520 | 0.925 | 1 | 0.336 | 0.607 | 0.219 | 1.680 |
| Temperature | 0.128 | 0.039 | 10.729 | 1 | 0.001 | 1.137 | 1.053 | 1.227 |
| Elevation | 0.007 | 0.003 | 6.915 | 1 | 0.009 | 1.007 | 1.002 | 1.012 |
| Seasons (Winter 2021) | 20.033 | 4 | <0.001 | |||||
| Autumn 2021 | −0.203 | 0.365 | 0.311 | 1 | 0.577 | 0.816 | 0.399 | 1.668 |
| Spring 2022 | −0.618 | 0.361 | 2.940 | 1 | 0.086 | 0.539 | 0.266 | 1.093 |
| Summer 2022 | −0.657 | 0.445 | 2.175 | 1 | 0.140 | 0.518 | 0.217 | 1.241 |
| Autumn 2022 | 0.332 | 0.375 | 0.786 | 1 | 0.375 | 1.394 | 0.669 | 2.906 |
| Presence of M. morganii | 1.549 | 0.192 | 65.060 | 1 | <0.001 | 4.709 | 3.231 | 6.862 |
| Presence of A. hydrophila | 0.950 | 0.414 | 5.266 | 1 | 0.022 | 2.585 | 1.149 | 5.819 |
| Presence of P. aeruginosa | 1.836 | 0.547 | 11.277 | 1 | <0.001 | 6.272 | 2.148 | 18.314 |
| Factor | B | S.E. | Wald | d.f. | P | Exp(B) | 95% C.I. | |
|---|---|---|---|---|---|---|---|---|
| Lower | Upper | |||||||
| Snake species (B. kraepelini) | 120.133 | 5 | <0.001 | |||||
| N. atra | 2.164 | 0.353 | 37.496 | 1 | <0.001 | 8.702 | 4.354 | 17.393 |
| B. multicinctus | 0.636 | 0.378 | 2.829 | 1 | 0.093 | 1.888 | 0.900 | 3.961 |
| T. stejnegeri | 2.184 | 0.379 | 33.301 | 1 | <0.001 | 8.885 | 4.231 | 18.659 |
| P. mucrosquamatus | 0.716 | 0.422 | 2.878 | 1 | 0.090 | 2.047 | 0.895 | 4.683 |
| E. t. friesi | 0.054 | 0.400 | 0.018 | 1 | 0.892 | 1.056 | 0.482 | 2.310 |
| Sex-Females (Males) | −0.365 | 0.150 | 5.887 | 1 | 0.015 | 0.694 | 0.517 | 0.932 |
| Temperature | −0.087 | 0.038 | 5.205 | 1 | 0.023 | 0.917 | 0.851 | 0.988 |
| Seasons (Winter 2021) | 16.695 | 4 | 0.002 | |||||
| Autumn 2021 | −0.027 | 0.366 | 0.005 | 1 | 0.942 | 0.974 | 0.475 | 1.996 |
| Spring 2022 | 0.100 | 0.353 | 0.080 | 1 | 0.777 | 1.105 | 0.553 | 2.210 |
| Summer 2022 | −0.155 | 0.438 | 0.125 | 1 | 0.723 | 0.857 | 0.363 | 2.020 |
| Autumn 2022 | 0.631 | 0.357 | 3.119 | 1 | 0.077 | 1.880 | 0.933 | 3.789 |
| Presence of E. faecalis | 1.534 | 0.190 | 64.990 | 1 | <0.001 | 4.639 | 3.194 | 6.737 |
| Presence of A. hydrophila | 1.156 | 0.264 | 19.099 | 1 | <0.001 | 3.177 | 1.892 | 5.335 |
| Presence of P. aeruginosa | 0.741 | 0.269 | 7.595 | 1 | 0.006 | 2.099 | 1.239 | 3.556 |
| Factor | B | S.E. | Wald | d.f. | p | Exp(B) | 95% C.I. | |
|---|---|---|---|---|---|---|---|---|
| Lower | Upper | |||||||
| Seasons (Winter 2021) | 19.347 | 4 | <0.001 | |||||
| Autumn 2021 | −0.133 | 0.458 | 0.084 | 1 | 0.771 | 0.875 | 0.357 | 2.149 |
| Spring 2022 | −0.731 | 0.516 | 2.009 | 1 | 0.156 | 0.481 | 0.175 | 1.323 |
| Summer 2022 | 0.527 | 0.419 | 1.584 | 1 | 0.208 | 1.693 | 0.746 | 3.846 |
| Autumn 2022 | −1.093 | 0.527 | 4.295 | 1 | 0.038 | 0.335 | 0.119 | 0.942 |
| Presence of E. faecalis | 2.045 | 0.529 | 14.943 | 1 | <0.001 | 7.730 | 2.741 | 21.802 |
| Presence of M. morganii | 0.520 | 0.250 | 4.317 | 1 | 0.038 | 1.683 | 1.030 | 2.749 |
| Presence of A. hydrophila | 1.503 | 0.292 | 26.520 | 1 | <0.001 | 4.496 | 2.537 | 7.966 |
| Intercept | −4.659 | 0.729 | 40.859 | 1 | <0.001 | 0.009 | ||
| Factor | B | S.E. | Wald | d.f. | p | Exp(B) | 95% C.I. | |
|---|---|---|---|---|---|---|---|---|
| Lower | Upper | |||||||
| Snake species (B. kraepelini) | 21.561 | 5 | <0.001 | |||||
| N. atra | −1.842 | 0.531 | 12.047 | 1 | <0.001 | 0.159 | 0.056 | 0.449 |
| B. multicinctus | −0.140 | 0.435 | 0.103 | 1 | 0.748 | 0.870 | 0.371 | 2.040 |
| T. stejnegeri | −1.286 | 0.561 | 5.257 | 1 | 0.022 | 0.276 | 0.092 | 0.830 |
| P. mucrosquamatus | −0.722 | 0.531 | 1.849 | 1 | 0.174 | 0.486 | 0.172 | 1.375 |
| E. t. friesi | −1.372 | 0.624 | 4.838 | 1 | 0.028 | 0.253 | 0.075 | 0.861 |
| Presence of E. faecalis | 1.055 | 0.397 | 7.073 | 1 | 0.008 | 2.871 | 1.320 | 6.244 |
| Presence of M. morganii | 1.257 | 0.278 | 20.495 | 1 | <0.001 | 3.515 | 2.040 | 6.056 |
| Presence of P. aeruginosa | 1.307 | 0.301 | 18.872 | 1 | <0.001 | 3.694 | 2.049 | 6.660 |
| Factor | E. faecalis | M. morganii | P. aeruginosa | A. hydrophila |
|---|---|---|---|---|
| Snake species (B. kraepelini) | ||||
| N. atra | ↓↓↓ | ↑↑↑ | ↓↓↓ | |
| B. multicinctus | ↓ | |||
| T. stejnegeri | ↓↓↓ | ↑↑↑ | ↓ | |
| P. mucrosquamatus | ↓↓↓ | |||
| E. t. friesi | ↓ | |||
| Sex (Males) | ||||
| Females | ↓ | |||
| Seasons (Winter 2021) | ||||
| Summer 2022 | ↓ | |||
| Temperature | ↑↑↑ | ↓ | ||
| Elevation | (↑) * | |||
| Coexisting bacteria | ↑↑↑ | ↑↑↑ | ↑↑↑ | ↑↑↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, W.-H.; Tsai, T.-S.; Chuang, P.-C. The Presence of Four Pathogenic Oral Bacterial Species in Six Wild Snake Species from Southern Taiwan: Associated Factors. Microorganisms 2024, 12, 263. https://doi.org/10.3390/microorganisms12020263
Lin W-H, Tsai T-S, Chuang P-C. The Presence of Four Pathogenic Oral Bacterial Species in Six Wild Snake Species from Southern Taiwan: Associated Factors. Microorganisms. 2024; 12(2):263. https://doi.org/10.3390/microorganisms12020263
Chicago/Turabian StyleLin, Wen-Hao, Tein-Shun Tsai, and Po-Chun Chuang. 2024. "The Presence of Four Pathogenic Oral Bacterial Species in Six Wild Snake Species from Southern Taiwan: Associated Factors" Microorganisms 12, no. 2: 263. https://doi.org/10.3390/microorganisms12020263
APA StyleLin, W.-H., Tsai, T.-S., & Chuang, P.-C. (2024). The Presence of Four Pathogenic Oral Bacterial Species in Six Wild Snake Species from Southern Taiwan: Associated Factors. Microorganisms, 12(2), 263. https://doi.org/10.3390/microorganisms12020263