
Impacts of Arbuscular Mycorrhizal Fungi on Metabolites of an Invasive Weed Wedelia trilobata
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Experimental Design
2.3. Determination of Mycorrhization Frequency
2.4. Metabolomic Analysis
2.4.1. Metabolite Extraction
2.4.2. Chromatographic Conditions
2.4.3. Mass Spectrometric Conditions
2.4.4. Data Processing and Multivariate Data Analysis
3. Results
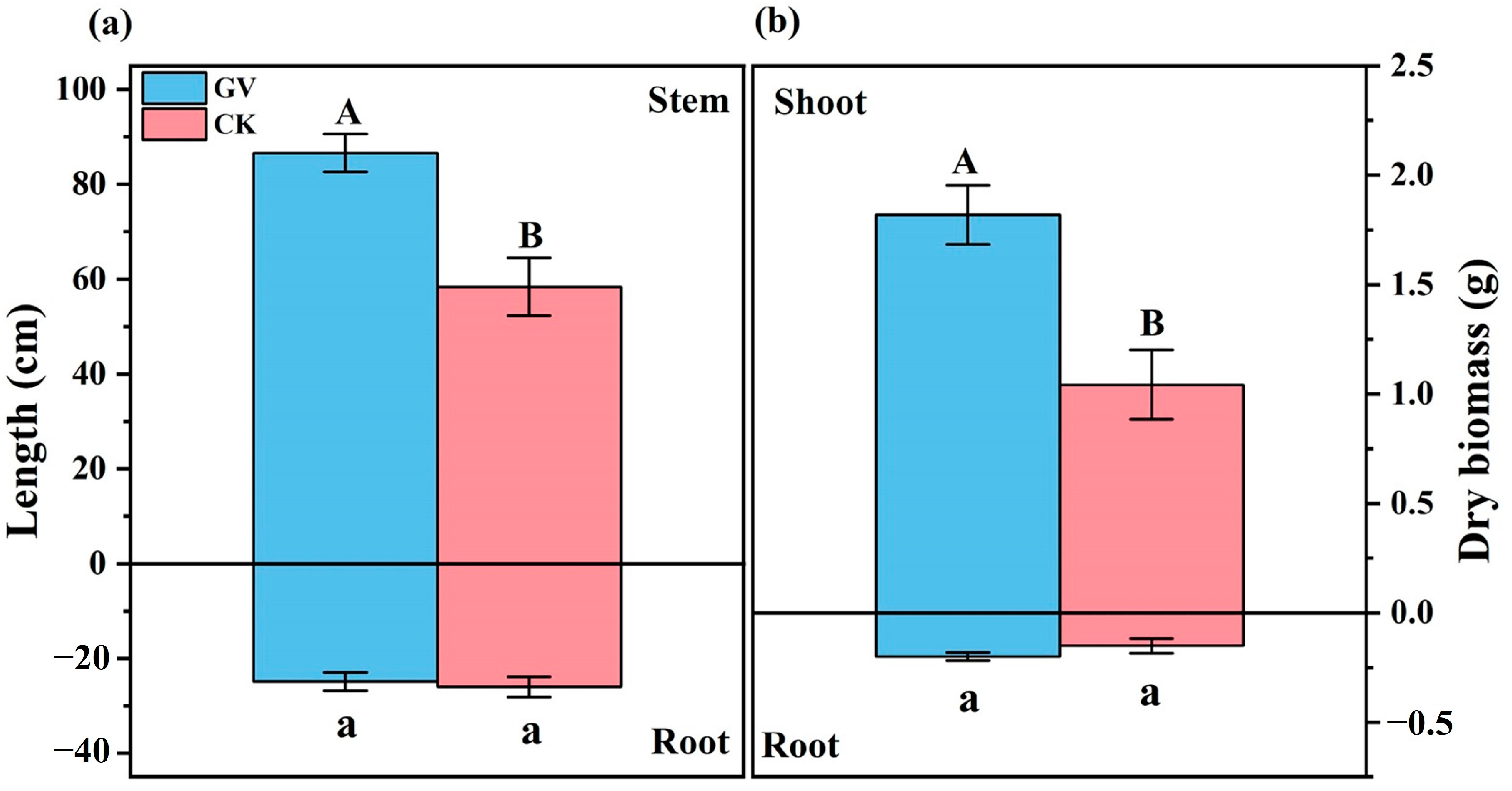
3.1. Root Colonization and Plant Growth
3.2. Metabolomics Analysis
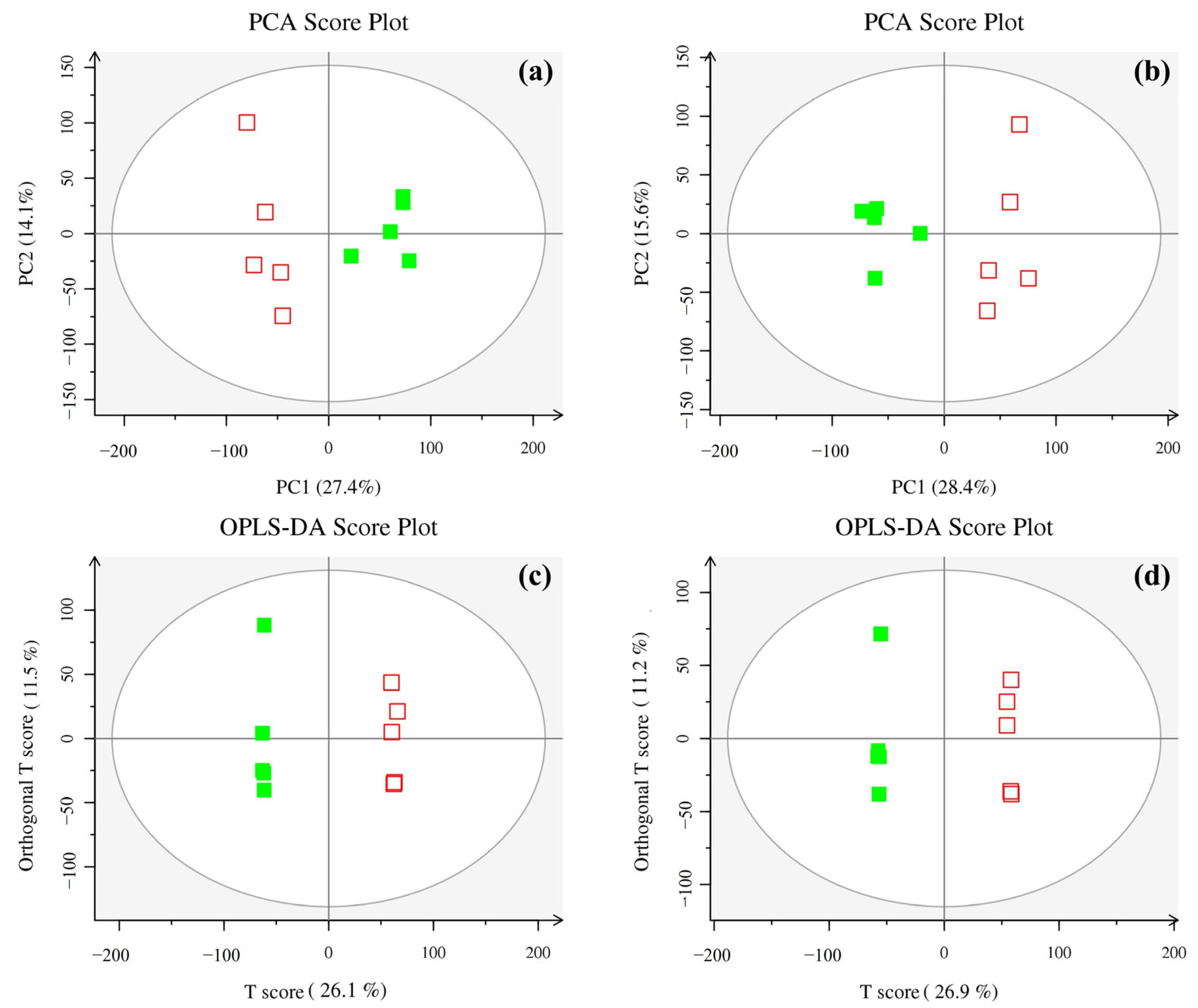
3.2.1. Multivariate Statistical Analysis
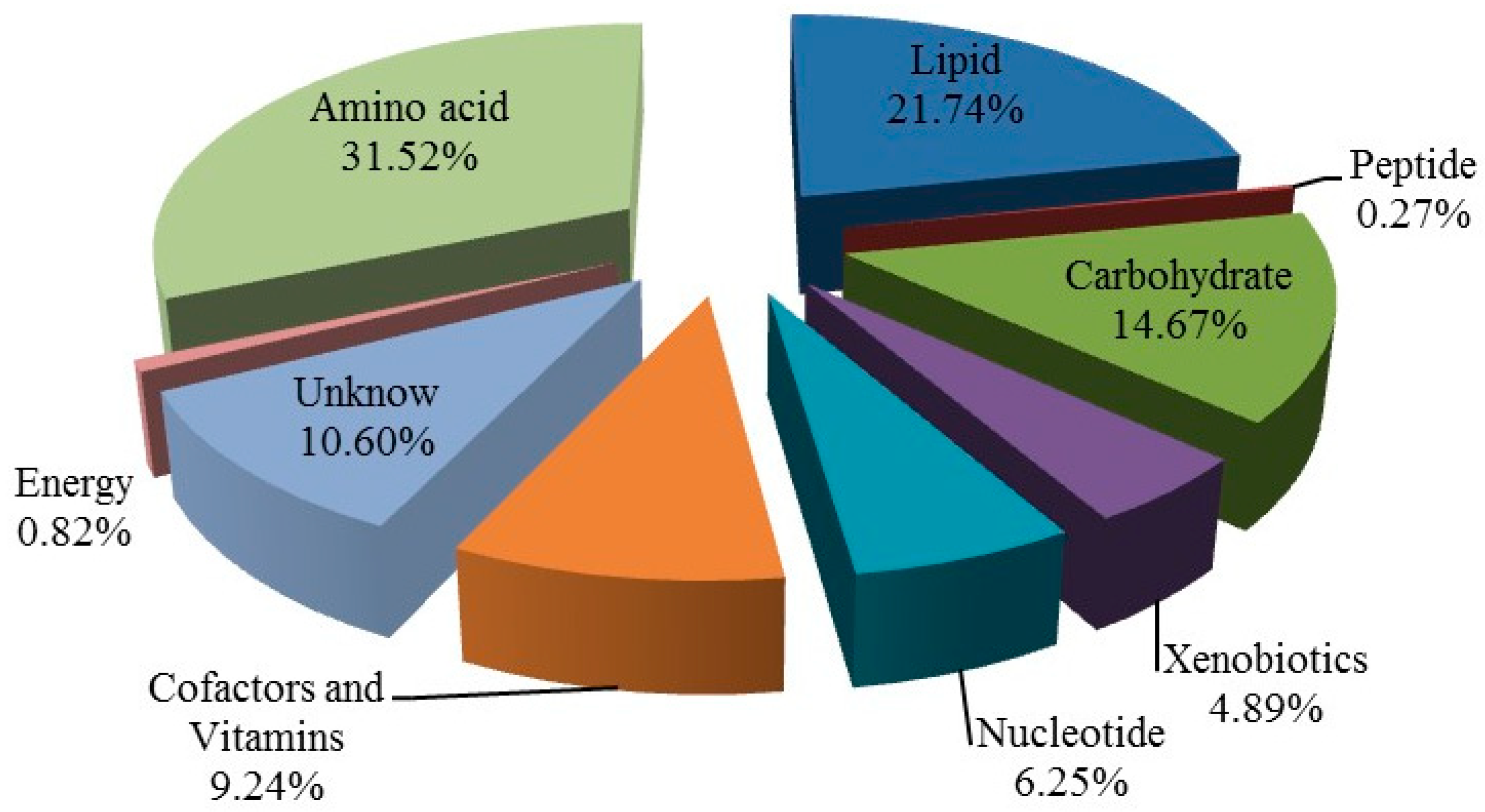
3.2.2. Comprehensive Identification of Metabolites
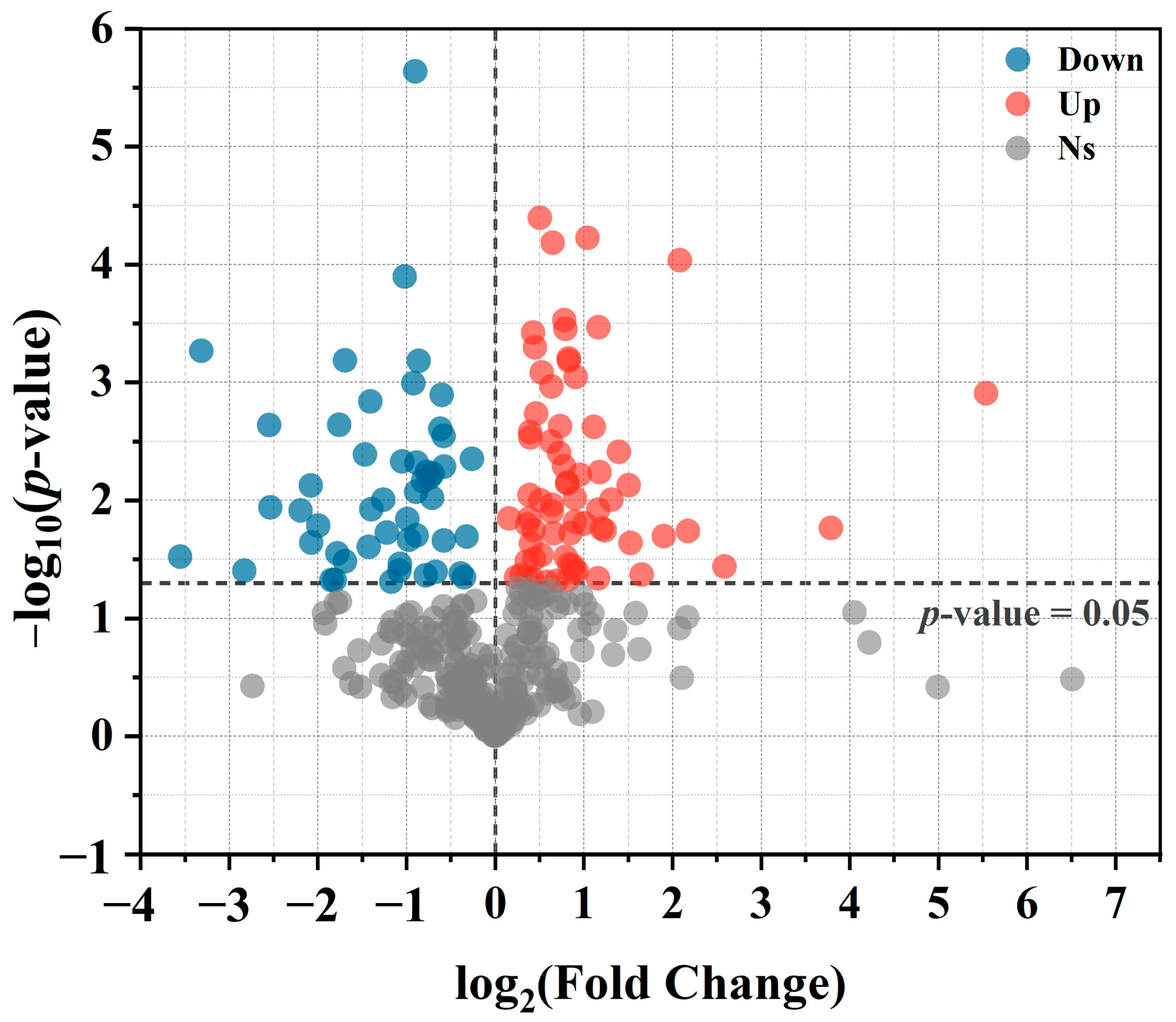
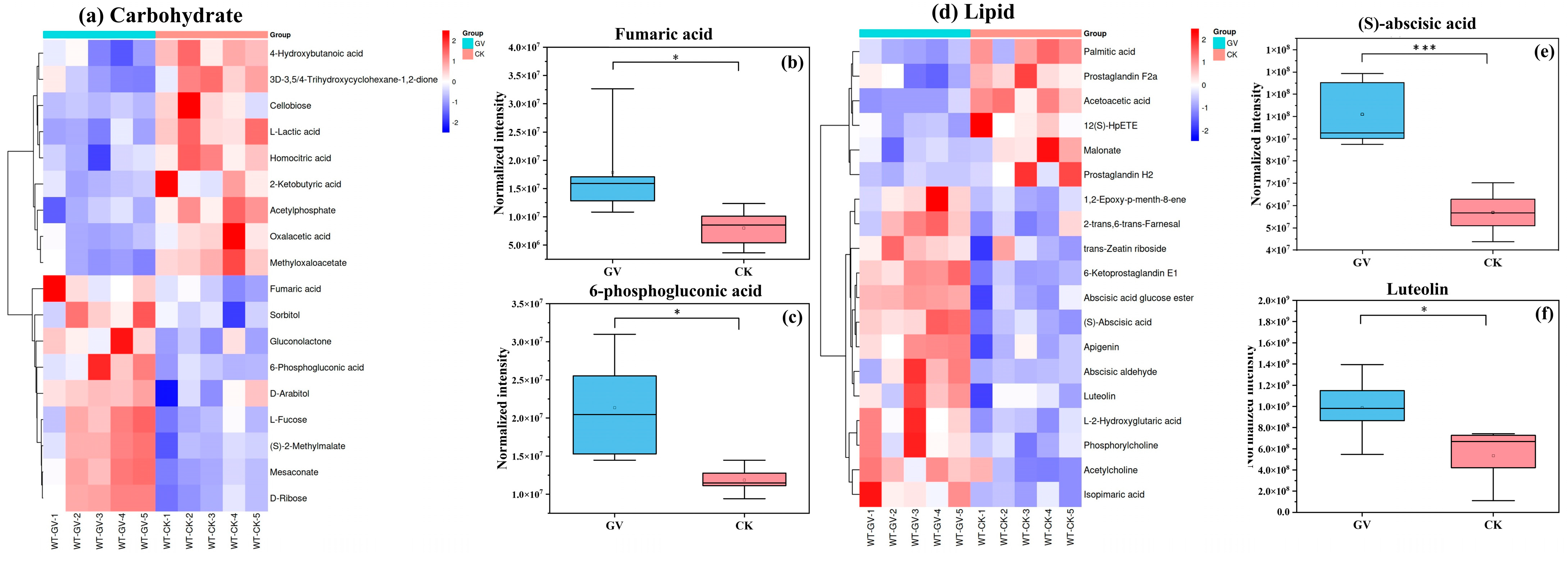
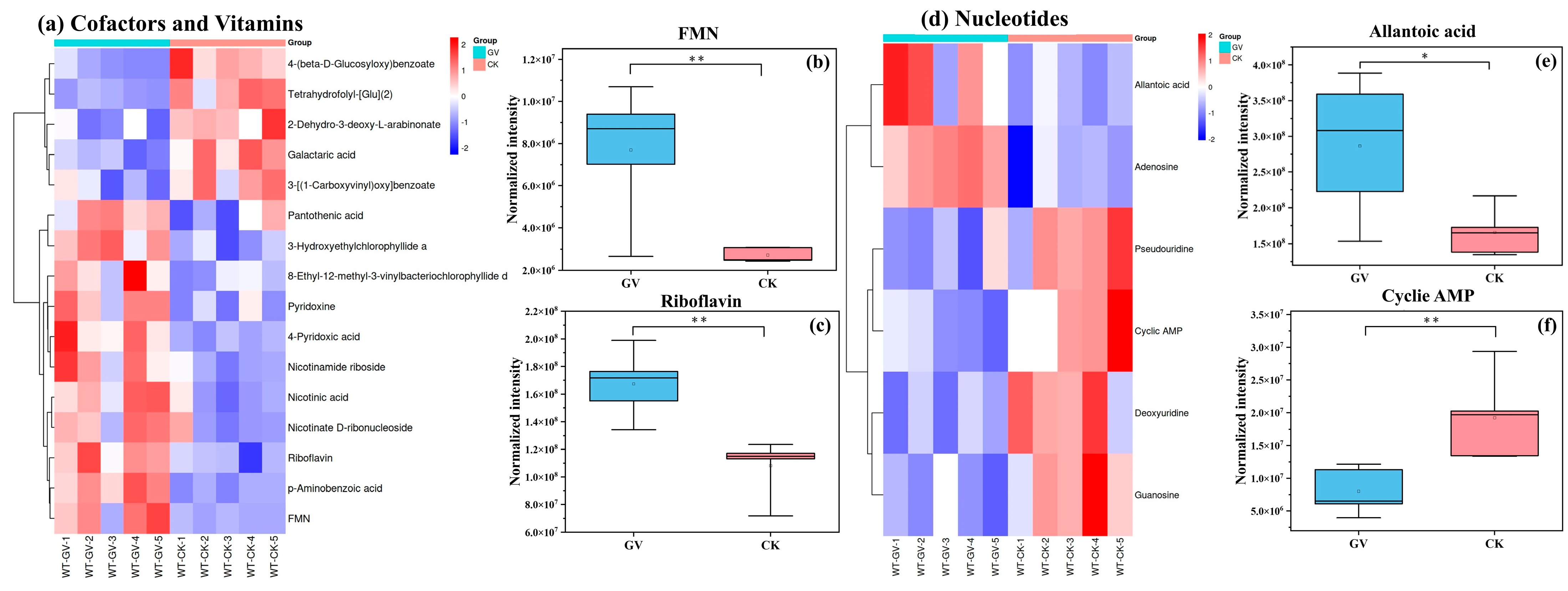
3.2.3. Differential Metabolite Analysis
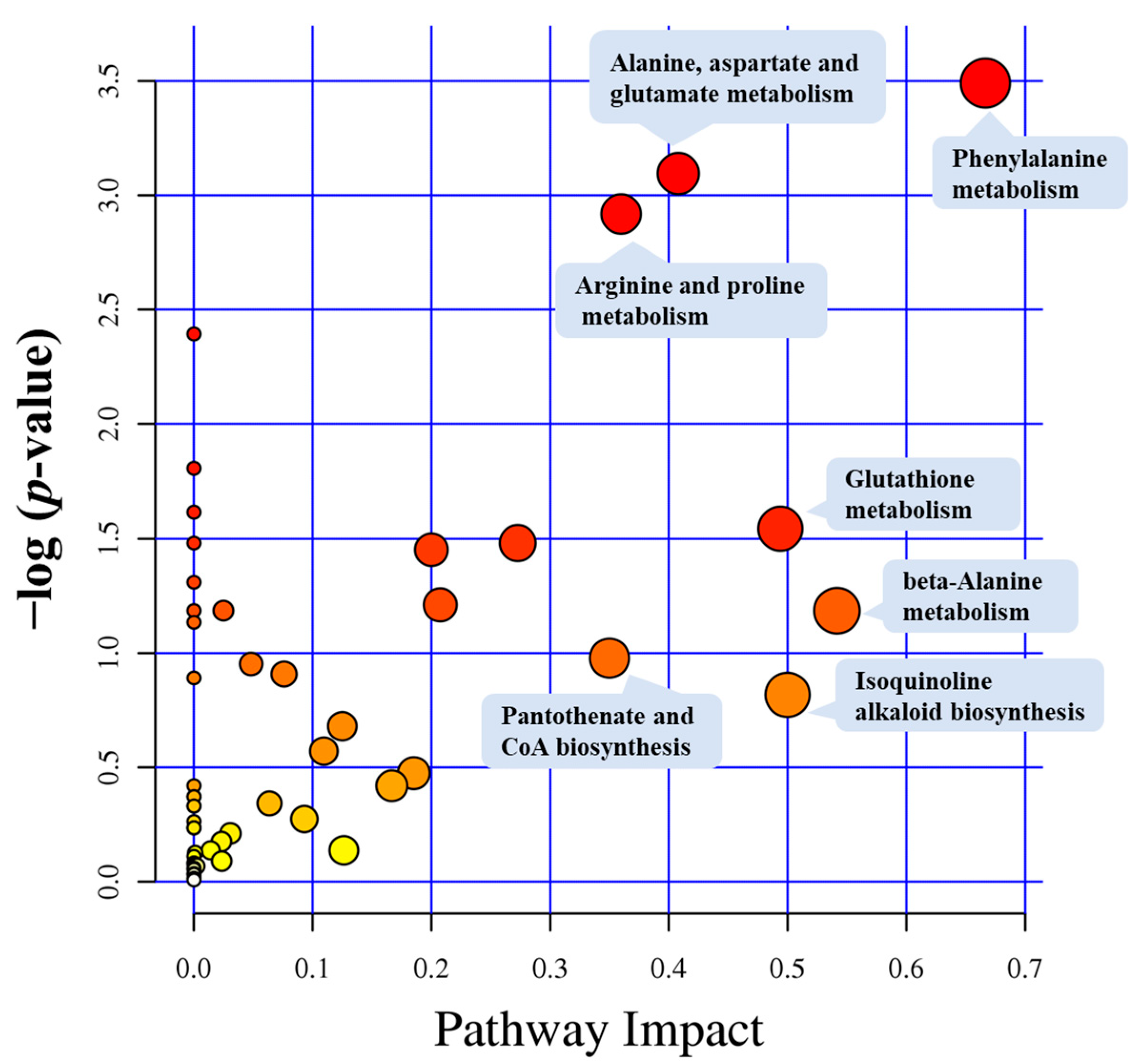
3.3. Metabolic Pathway Analysis
4. Discussion
4.1. Effects of AMF on Invasive Plant Growth
4.2. Regulation of Plant Metabolites by AMF
4.2.1. Impacts of Differential Metabolites on Plant Growth
4.2.2. Impacts of Differential Metabolites on Plant Stress Resistance
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Taylor, T.N.; Remy, W.; Hass, H.; Kerp, H. Fossil arbuscular mycorrhizae from the early devonian. Mycologia 2018, 87, 560–573. [Google Scholar] [CrossRef]
- Henkes, G.J.; Kandeler, E.; Marhan, S.; Scheu, S.; Bonkowski, M. Interactions of mycorrhiza and protists in the rhizosphere systemically alter microbial community composition, plant shoot-to-root ratio and within-root system nitrogen allocation. Front. Environ. Sci. 2018, 6, 117. [Google Scholar] [CrossRef]
- Hill, E.M.; Robinson, L.A.; Abdul-Sada, A.; Vanbergen, A.J.; Hodge, A.; Hartley, S.E. Arbuscular mycorrhizal fungi and plant chemical defence: Effects of colonisation on aboveground and belowground metabolomes. J. Chem. Ecol. 2018, 44, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Goddard, M.L.; Belval, L.; Martin, I.R.; Roth, L.; Laloue, H.; Deglene-Benbrahim, L.; Valat, L.; Bertsch, C.; Chong, J. Arbuscular mycorrhizal symbiosis triggers major changes in primary metabolism together with modification of defense responses and signaling in both roots and leaves of Vitis vinifera. Front. Plant Sci. 2021, 12, 721614. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; He, Y.; Zhang, W.; Chen, L.; Zhang, J.; Zhang, X.; Dawson, W.; Ding, J. Greater chemical signaling in root exudates enhances soil mutualistic associations in invasive plants compared to natives. New Phytol. 2022, 236, 1140–1153. [Google Scholar] [CrossRef] [PubMed]
- Hodge, A.; Storer, K. Arbuscular mycorrhiza and nitrogen: Implications for individual plants through to ecosystems. Plant Soil 2014, 386, 1–19. [Google Scholar] [CrossRef]
- Qi, S.S.; Dai, Z.C.; Miao, S.L.; Zhai, D.L.; Si, C.C.; Huang, P.; Wang, R.P.; Du, D.L. Light limitation and litter of an invasive clonal plant, Wedelia trilobata, inhibit its seedling recruitment. Ann. Bot. 2014, 114, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Luque, G.M.; Bellard, C.; Bertelsmeier, C.; Bonnaud, E.; Genovesi, P.; Simberloff, D.; Courchamp, F. The 100th of the world’s worst invasive alien species. Biol. Invasions 2013, 16, 981–985. [Google Scholar] [CrossRef]
- Qi, S.S.; Dai, Z.C.; Zhai, D.L.; Chen, S.C.; Si, C.C.; Huang, P.; Wang, R.P.; Zhong, Q.X.; Du, D.L. Curvilinear effects of invasive plants on plant diversity: Plant community invaded by Sphagneticola trilobata. PLoS ONE 2014, 9, e113964. [Google Scholar] [CrossRef]
- Vila, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarosik, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pysek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Dickie, I.A.; Bufford, J.L.; Cobb, R.C.; Desprez-Loustau, M.L.; Grelet, G.; Hulme, P.E.; Klironomos, J.; Makiola, A.; Nunez, M.A.; Pringle, A.; et al. The emerging science of linked plant-fungal invasions. N. Phytol. 2017, 215, 1314–1332. [Google Scholar] [CrossRef] [PubMed]
- Pringle, A.; Bever, J.D.; Gardes, M.; Parrent, J.L.; Rillig, M.C.; Klironomos, J.N. Mycorrhizal symbioses and plant invasions. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 699–715. [Google Scholar] [CrossRef]
- Shah, M.A.; Reshi, Z.A.; Khasa, D.P. Arbuscular mycorrhizas: Drivers or passengers of alien plant invasion. Bot. Rev. 2009, 75, 397–417. [Google Scholar] [CrossRef]
- Sun, D.; Yang, X.; Wang, Y.; Fan, Y.; Ding, P.; Song, X.; Yuan, X.; Yang, X. Stronger mutualistic interactions with arbuscular mycorrhizal fungi help Asteraceae invaders outcompete the phylogenetically related natives. N. Phytol. 2022, 236, 1487–1496. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; Wang, J.; Wan, L.; Dai, Z.; da Silva Matos, D.M.; Du, D.; Egan, S.; Bonser, S.P.; Thomas, T.; Moles, A.T. Arbuscular mycorrhizal fungi contribute to phosphorous uptake and allocation strategies of Solidago canadensis in a phosphorous-deficient environment. Front. Plant Sci. 2022, 13, 831654. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wu, W.W.; Qi, S.S.; Cheng, H.; Li, Q.; Ran, Q.; Dai, Z.C.; Du, D.L.; Egan, S.; Thomas, T. Arbuscular mycorrhizal fungi improve the growth and disease resistance of the invasive plant Wedelia trilobata. J. Appl. Microbiol. 2021, 130, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Tsiokanos, E.; Cartabia, A.; Tsafantakis, N.; Lalaymia, I.; Termentzi, A.; Miguel, M.; Declerck, S.; Fokialakis, N. The metabolic profile of Anchusa officinalis L. differs according to its associated arbuscular mycorrhizal fungi. Metabolites 2022, 12, 573. [Google Scholar] [CrossRef] [PubMed]
- Nacoon, S.; Jogloy, S.; Riddech, N.; Mongkolthanaruk, W.; Kuyper, T.W.; Boonlue, S. Interaction between phosphate solubilizing bacteria and arbuscular mycorrhizal fungi on growth promotion and tuber inulin content of Helianthus tuberosus L. Sci. Rep. 2020, 10, 4916. [Google Scholar] [CrossRef] [PubMed]
- Sangwan, S.; Prasanna, R. Mycorrhizae helper bacteria: Unlocking their potential as bioenhancers of plant-arbuscular mycorrhizal fungal associations. Microb. Ecol. 2022, 84, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Schillaci, M.; Roessner, U. Metabolomics as an emerging tool to study plant-microbe interactions. Emerg. Top. Life Sci. 2022, 6, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Fu, X. Reprogramming of plant central metabolism in response to abiotic stresses: A metabolomics view. Int. J. Mol. Sci. 2022, 23, 5716. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Jian, T.; Liu, X.; Lv, Q.; Zhang, G.; Ling, J. Application of metabolomics in fungal research. Molecules 2022, 27, 7365. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, K.; Ibrahim, B.; Ahmad Bawadikji, A.; Lim, J.W.; Tong, W.Y.; Leong, C.R.; Khaw, K.Y.; Tan, W.N. Recent developments in metabolomics studies of endophytic fungi. J. Fungi 2021, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Rozmos, M.; Bukovska, P.; Hrselova, H.; Kotianova, M.; Dudas, M.; Gancarcikova, K.; Jansa, J. Organic nitrogen utilisation by an arbuscular mycorrhizal fungus is mediated by specific soil bacteria and a protist. ISME J. 2022, 16, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Ma, R.; Chen, X.L. Advances of metabolomics in fungal pathogen-plant interactions. Metabolites 2019, 9, 169. [Google Scholar] [CrossRef] [PubMed]
- Shtark, O.; Puzanskiy, R.; Avdeeva, G.; Yemelyanov, V.; Shavarda, A.; Romanyuk, D.; Kliukova, M.; Kirpichnikova, A.; Tikhonovich, I.; Zhukov, V.; et al. Metabolic alterations in Pisum sativum roots during plant growth and arbuscular mycorrhiza development. Plants 2021, 10, 1033. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Suseela, V. Unraveling arbuscular mycorrhiza-Induced changes in plant primary and secondary metabolome. Metabolites 2020, 10, 335. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.K.; Sadhukhan, S. Unearthing the alteration in plant volatiles induced by mycorrhizal fungi: A shield against plant pathogens. Physiol. Plant. 2023, 175, e13845. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.S.; Manoharan, B.; Dhandapani, V.; Jegadeesan, S.; Rutherford, S.; Wan, J.S.H.; Huang, P.; Dai, Z.C.; Du, D.L. Pathogen resistance in Sphagneticola trilobata (Singapore daisy): Molecular associations and differentially expressed genes in response to disease from a widespread fungus. Genetica 2022, 150, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.S.; Liu, Y.J.; Dai, Z.C.; Wan, L.Y.; Du, D.L.; Ju, R.T.; Wan, J.S.H.; Bonser, S.P. Allelopathy confers an invasive Wedelia higher resistance to generalist herbivore and pathogen enemies over its native congener. Oecologia 2020, 192, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Xing, Z.; Ma, T.; Wu, L.; Zhang, Z.; Ding, J.; Siemann, E. Foliar herbivory modifies arbuscular mycorrhizal fungal colonization likely through altering root flavonoids. Funct. Ecol. 2023, 38, 259–271. [Google Scholar] [CrossRef]
- Gao, X.; Guo, H.; Zhang, Q.; Guo, H.; Zhang, L.; Zhang, C.; Gou, Z.; Liu, Y.; Wei, J.; Chen, A.; et al. Arbuscular mycorrhizal fungi (AMF) enhanced the growth, yield, fiber quality and phosphorus regulation in upland cotton (Gossypium hirsutum L.). Sci. Rep. 2020, 10, 2084. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, W.; Yang, J.; Qin, S.; Yang, Y.; Sun, C.; Pei, G.; Zeeshan, M.; Liao, H.; Liu, L.; et al. Mycorrhizal symbiosis promotes the nutrient content accumulation and affects the root exudates in maize. BMC Plant Biol. 2022, 22, 64. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.N.; Qin, Q.Y.; Ma, W.Y.; Zhou, L.J.; Wu, Q.S.; Xu, Y.J.; Kuca, K.; Hashem, A.; Al-Arjani, A.F.; Almutairi, K.F.; et al. Metabolomics reveals arbuscular mycorrhizal fungi-mediated tolerance of walnut to soil drought. BMC Plant Biol. 2023, 23, 118. [Google Scholar] [CrossRef] [PubMed]
- Majewska, M.L.; Rola, K.; Zubek, S. The growth and phosphorus acquisition of invasive plants Rudbeckia laciniata and Solidago gigantea are enhanced by arbuscular mycorrhizal fungi. Mycorrhiza 2017, 27, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Tu, C.; Chen, X.; Hu, S. Arbuscular mycorrhizal fungi enhance P uptake and alter plant morphology in the invasive plant Microstegium vimineum. Biol. Invasions 2013, 16, 1083–1093. [Google Scholar] [CrossRef]
- Osunkoya, O.O.; Bayliss, D.; Panetta, F.D.; Vivian-Smith, G. Leaf trait co-ordination in relation to construction cost, carbon gain and resource-use efficiency in exotic invasive and native woody vine species. Ann. Bot. 2010, 106, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, J.M.; Sánchez, C.; Navarro, L. Can asexual reproduction by plant fragments help to understand the invasion of the NW Iberian coast by Spartina patens? Flora 2019, 257, 151410. [Google Scholar] [CrossRef]
- Martinez-Medina, A.; Flors, V.; Heil, M.; Mauch-Mani, B.; Pieterse, C.M.J.; Pozo, M.J.; Ton, J.; van Dam, N.M.; Conrath, U. Recognizing Plant Defense Priming. Trends Plant Sci. 2016, 21, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Mauch-Mani, B.; Baccelli, I.; Luna, E.; Flors, V. Defense Priming: An Adaptive Part of Induced Resistance. Annu. Rev. Plant Biol. 2017, 68, 485–512. [Google Scholar] [CrossRef] [PubMed]
- Desimone, M.; Catoni, E.; Ludewig, U.; Hilpert, M.; Schneider, A.; Kunze, R.; Tegeder, M.; Frommer, W.B.; Schumacher, K. A novel superfamily of transporters for allantoin and other oxo derivatives of nitrogen heterocyclic compounds in Arabidopsis. Plant Cell 2002, 14, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Pélissier, H.C.; Frerich, A.; Desimone, M.; Schumacher, K.; Tegeder, M. PvUPS1, an Allantoin Transporter in Nodulated Roots of French Bean. Plant Physiol. 2004, 134, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Redillas, M.; Bang, S.W.; Lee, D.K.; Kim, Y.S.; Jung, H.; Chung, P.J.; Suh, J.W.; Kim, J.K. Allantoin accumulation through overexpression of ureide permease1 improves rice growth under limited nitrogen conditions. Plant Biotechnol. J. 2019, 17, 1289–1301. [Google Scholar] [CrossRef] [PubMed]
- Keymer, A.; Pimprikar, P.; Wewer, V.; Huber, C.; Brands, M.; Bucerius, S.L.; Delaux, P.M.; Klingl, V.; Ropenack-Lahaye, E.V.; Wang, T.L.; et al. Lipid transfer from plants to arbuscular mycorrhiza fungi. eLife 2017, 6, e29107. [Google Scholar] [CrossRef] [PubMed]
- Salmeron-Santiago, I.A.; Martinez-Trujillo, M.; Valdez-Alarcon, J.J.; Pedraza-Santos, M.E.; Santoyo, G.; Pozo, M.J.; Chavez-Barcenas, A.T. An Updated Review on the Modulation of Carbon Partitioning and Allocation in Arbuscular Mycorrhizal Plants. Microorganisms 2022, 10, 75. [Google Scholar] [CrossRef] [PubMed]
- Chia, D.W.; Yoder, T.J.; Reiter, W.-D.; Gibson, S.I. Fumaric acid: An overlooked form of fixed carbon in Arabidopsis and other plant species. Planta 2000, 211, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Araujo, W.L.; Nunes-Nesi, A.; Fernie, A.R. Fumarate: Multiple functions of a simple metabolite. Phytochemistry 2011, 72, 838–843. [Google Scholar] [CrossRef] [PubMed]
- Al-Arjani, A.F.; Hashem, A.; Abd Allah, E.F. Arbuscular mycorrhizal fungi modulates dynamics tolerance expression to mitigate drought stress in Ephedra foliata Boiss. Saudi J. Biol. Sci. 2020, 27, 380–394. [Google Scholar] [CrossRef]
- Jia, T.; Wang, J.; Chang, W.; Fan, X.; Sui, X.; Song, F. Proteomics analysis of E. angustifolia seedlings inoculated with arbuscular mycorrhizal fungi under salt stress. Int. J. Mol. Sci. 2019, 20, 788. [Google Scholar] [CrossRef] [PubMed]
- Souza, L.A.; Camargos, L.S.; Schiavinato, M.A.; Andrade, S.A.L. Mycorrhization alters foliar soluble amino acid composition and influences tolerance to Pb in Calopogonium mucunoides. Theor. Exp. Plant Physiol. 2014, 26, 211–216. [Google Scholar] [CrossRef]
- Ben Rejeb, K.; Abdelly, C.; Savoure, A. Proline, a multifunctional amino-acid involved in plant adaptation to environmental constraints. Biol. Aujourdhui 2012, 206, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, M.; Upadhyay, R.S. Regulation of L-proline biosynthesis, signal transduction, transport, accumulation and its vital role in plants during variable environmental conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Bi, S.; Meng, J.; Liu, T.; Li, P.; Yu, C.; Peng, X. Arbuscular mycorrhizal fungi enhanced rice proline metabolism under low temperature with nitric oxide involvement. Front. Plant Sci. 2022, 13, 962460. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.J.; Reddy, M.S.; Wang, L. The phenylpropanoid pathway and plant defence—A genomics perspective. Mol. Plant Pathol 2002, 3, 371–390. [Google Scholar] [CrossRef]
- Chen, Q.; Man, C.; Li, D.; Tan, H.; Xie, Y.; Huang, J. Arogenate dehydratase isoforms differentially regulate anthocyanin biosynthesis in Arabidopsis thaliana. Mol. Plant 2016, 9, 1609–1619. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal. Behav. 2010, 5, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Aseel, D.G.; Rashad, Y.M.; Hammad, S.M. Arbuscular mycorrhizal fungi trigger transcriptional expression of flavonoid and chlorogenic acid biosynthetic pathways genes in tomato against Tomato Mosaic Virus. Sci. Rep. 2019, 9, 9692. [Google Scholar] [CrossRef] [PubMed]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [PubMed]
- Ran, Z.; Ding, W.; Cao, S.; Fang, L.; Zhou, J.; Zhang, Y. Arbuscular mycorrhizal fungi: Effects on secondary metabolite accumulation of traditional chinese medicines. Plant Biol. 2022, 24, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, R.; Muller, C. Leaf metabolome in arbuscular mycorrhizal symbiosis. Curr. Opin. Plant Biol. 2015, 26, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Michaeli, S.; Fromm, H. Closing the loop on the GABA shunt in plants: Are GABA metabolism and signaling entwined? Front. Plant Sci. 2015, 6, 419. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yu, J.; Peng, Y.; Huang, B. Metabolic pathways regulated by γ-aminobutyric acid (GABA) contributing to heat tolerance in creeping bentgrass (Agrostis stolonifera). Sci. Rep. 2016, 6, 30338. [Google Scholar] [CrossRef]
- Bown, A.W.; Shelp, B.J. Plant GABA: Not just a metabolite. Trends Plant Sci. 2016, 21, 811–813. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.M.; Alabdallah, N.M.; Alharbi, B.M.; Waseem, M.; Yao, G.; Liu, X.D.; Abd El-Gawad, H.G.; El-Yazied, A.A.; Ibrahim, M.F.M.; Jahan, M.S.; et al. GABA: A key player in drought stress resistance in plants. Int. J. Mol. Sci. 2021, 22, 10136. [Google Scholar] [CrossRef] [PubMed]
- Hijaz, F.; Nehela, Y.; Killiny, N. Application of gamma-aminobutyric acid increased the level of phytohormones in Citrus sinensis. Planta 2018, 248, 909–918. [Google Scholar] [CrossRef] [PubMed]
- Tarkowski, L.P.; Signorelli, S.; Hofte, M. γ-Aminobutyric acid and related amino acids in plant immune responses: Emerging mechanisms of action. Plant Cell Environ. 2020, 43, 1103–1116. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Jalil, S.U.; Chopra, P.; Chhillar, H.; Ferrante, A.; Khan, N.A.; Ansari, M.I. Role of GABA in plant growth, development and senescence. Plant Gene 2021, 26, 100283. [Google Scholar] [CrossRef]
- Shelp, B.; Bown, A.; Zarei, A. 4-Aminobutyrate (GABA): A metabolite and signal with practical significance. Botany 2017, 95, 1015–1032. [Google Scholar] [CrossRef]
- Michaeli, S.; Fait, A.; Lagor, K.; Nunes-Nesi, A.; Grillich, N.; Yellin, A.; Bar, D.; Khan, M.; Fernie, A.R.; Turano, F.J.; et al. A mitochondrial GABA permease connects the GABA shunt and the TCA cycle, and is essential for normal carbon metabolism. Plant J. 2011, 67, 485–498. [Google Scholar] [CrossRef]
- Zhang, D.; Wei, X.; Liu, Z.; Wu, X.; Bao, C.; Sun, Y.; Su, N.; Cui, J. Transcriptome analysis reveals the molecular mechanism of GABA accumulation during quinoa (Chenopodium quinoa Willd.) germination. J. Agric. Food. Chem. 2021, 69, 12171–12186. [Google Scholar] [CrossRef] [PubMed]
- Hoeksema, J.D.; Bever, J.D.; Chakraborty, S.; Chaudhary, V.B.; Gardes, M.; Gehring, C.A.; Hart, M.M.; Housworth, E.A.; Kaonongbua, W.; Klironomos, J.N.; et al. Evolutionary history of plant hosts and fungal symbionts predicts the strength of mycorrhizal mutualism. Commun. Biol. 2018, 1, 116. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, S.; Radhakrishnan, D.; Prasad, K.; Chini, A. Fungal production and manipulation of plant hormones. Curr. Med. Chem. 2018, 25, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Vierheilig, H.; Steinkellner, S.; Khaosaad, T.; Garcia-Garrido, J.M. The biocontrol effect of mycorrhization on soilborne fungal pathogens and the autoregulation of the AM symbiosis: One mechanism, two effects? In Mycorrhiza; Springer: Berlin/Heidelberg, Germany, 2008; pp. 307–320. [Google Scholar]
- Fernandez, I.; Merlos, M.; Lopez-Raez, J.A.; Martinez-Medina, A.; Ferrol, N.; Azcon, C.; Bonfante, P.; Flors, V.; Pozo, M.J. Defense related phytohormones regulation in arbuscular mycorrhizal symbioses depends on the partner genotypes. J. Chem. Ecol. 2014, 40, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Davies, W.J.; Kudoyarova, G.; Hartung, W. Long-distance ABA signaling and its relation to other signaling pathways in the detection of soil drying and the mediation of the plant’s response to drought. J. Plant Growth Regul. 2005, 24, 285–295. [Google Scholar] [CrossRef]
- Ondzighi-Assoume, C.A.; Chakraborty, S.; Harris, J.M. Environmental nitrate stimulates abscisic acid accumulation in arabidopsis root tips by releasing it from inactive stores. Plant Cell 2016, 28, 729–745. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway ID | Description | Number | Metabolites | Kegg ID |
|---|---|---|---|---|
| ath00360 | Phenylalanine metabolism | 3 | L-phenylalanine | C00079 |
| Trans-cinnamate | C00423 | |||
| Phenylacetaldehyde | C00601 | |||
| ath00250 | Alanine, aspartate and glutamate metabolism | 5 | L-glutamic acid | C00025 |
| 2-keto-glutaramic acid | C00940 | |||
| Oxalacetic acid | C00036 | |||
| Fumaric acid | C00122 | |||
| γ-aminobutyric acid | C00334 | |||
| ath00330 | Arginine and proline metabolism | 7 | L-glutamic acid | C00025 |
| N-acetylglutamic acid | C00624 | |||
| L-proline | C00148 | |||
| L-glutamic gamma-semialdehyde | C01165 | |||
| Fumaric acid | C00122 | |||
| Oxalacetic acid | C00436 | |||
| γ-aminobutyric acid | C00334 | |||
| ath00480 | Glutathione metabolism | 4 | Glutathione | C00051 |
| γ-glutamylcysteine | C00669 | |||
| Pyroglutamic acid | C01879 | |||
| L-glutamic acid | C00025 | |||
| ath00410 | beta-Alanine metabolism | 2 | β-alanine | C00099 |
| Pantothenic acid | C00864 | |||
| ath00950 | Isoquinoline alkaloid biosynthesis | 1 | L-tyrosine | C00082 |
| L-tyrosine | Pantothenate and CoA biosynthesis | 2 | β-alanine | C00099 |
| Pantothenic acid | C008640 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, X.; Chen, D.; Zhang, Y.; Naz, M.; Dai, Z.; Qi, S.; Du, D. Impacts of Arbuscular Mycorrhizal Fungi on Metabolites of an Invasive Weed Wedelia trilobata. Microorganisms 2024, 12, 701. https://doi.org/10.3390/microorganisms12040701
Jiang X, Chen D, Zhang Y, Naz M, Dai Z, Qi S, Du D. Impacts of Arbuscular Mycorrhizal Fungi on Metabolites of an Invasive Weed Wedelia trilobata. Microorganisms. 2024; 12(4):701. https://doi.org/10.3390/microorganisms12040701
Chicago/Turabian StyleJiang, Xinqi, Daiyi Chen, Yu Zhang, Misbah Naz, Zhicong Dai, Shanshan Qi, and Daolin Du. 2024. "Impacts of Arbuscular Mycorrhizal Fungi on Metabolites of an Invasive Weed Wedelia trilobata" Microorganisms 12, no. 4: 701. https://doi.org/10.3390/microorganisms12040701