
Single and Combined Effects of Phenanthrene and Silver Nanoparticles on Denitrification Processes in Coastal Marine Sediments
Abstract
:1. Introduction
2. Materials and Methods
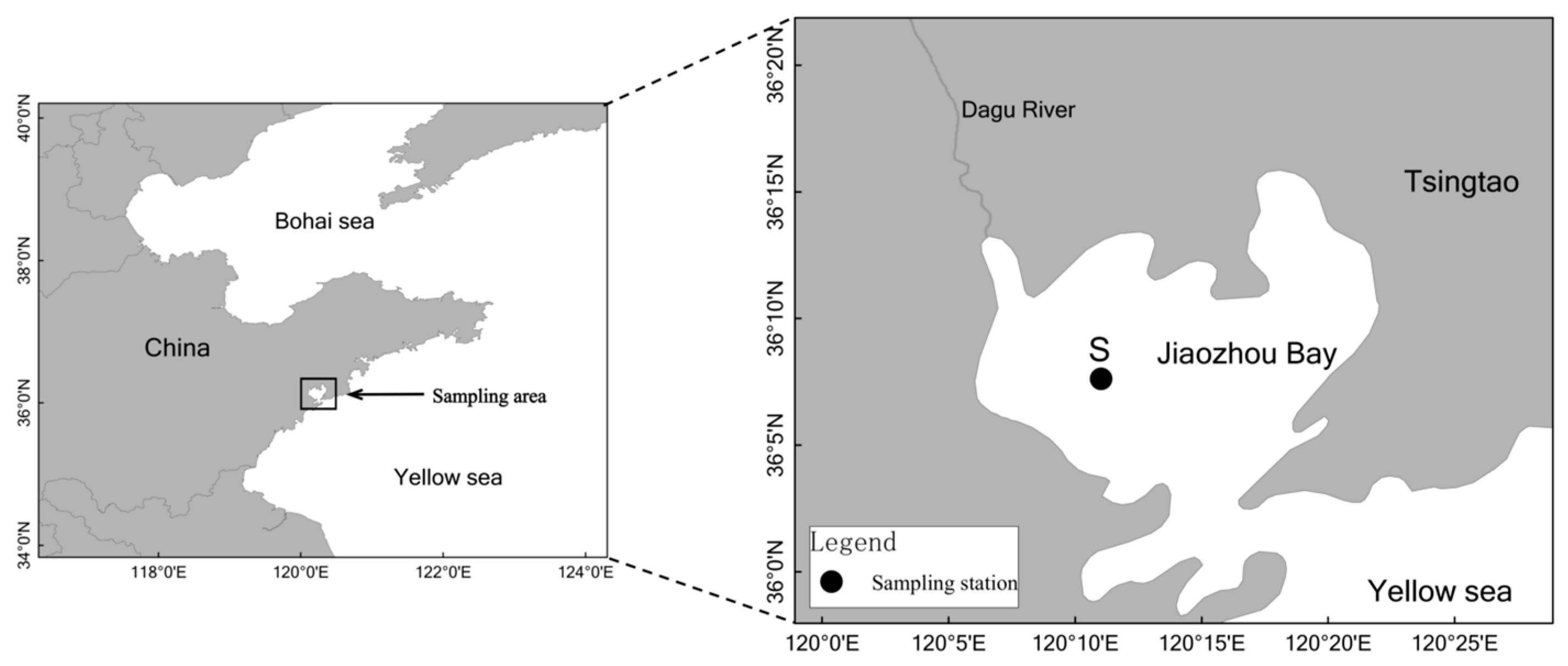
2.1. Sample Collection
2.2. Experiment Setups
2.2.1. Exposure to Phe and AgNPs
2.2.2. Six Days Batch Exposure Experiment
2.2.3. Denitrification Activity
2.2.4. Determination of Denitrifying Reductase
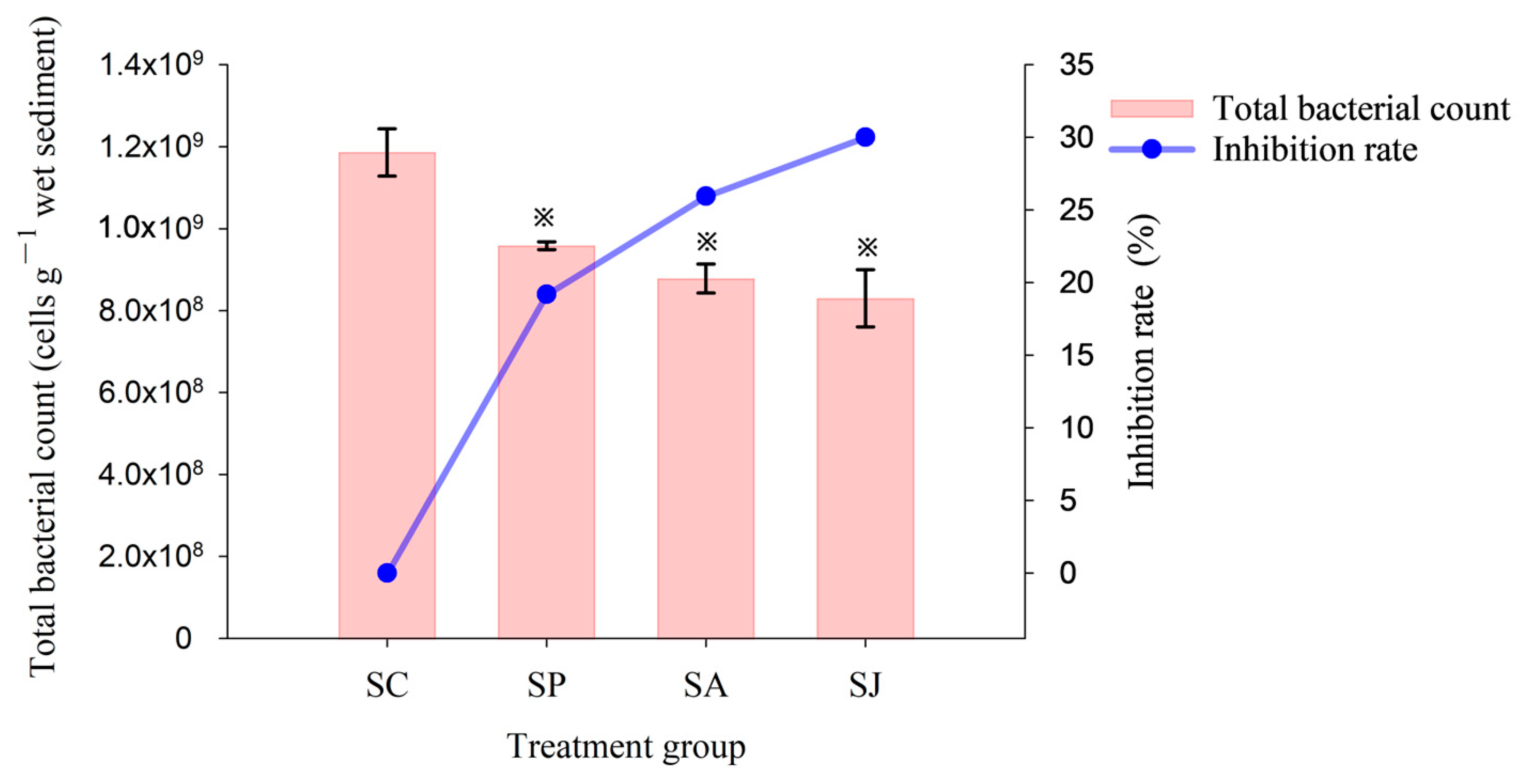
2.3. Total Bacterial Count
2.4. DNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
2.5. Gene Amplification, High Throughput Sequencing and Data Analysis
3. Results and Discussion
3.1. Effect of Single and Combined Contamination of Phe and AgNPs on Denitrification Process
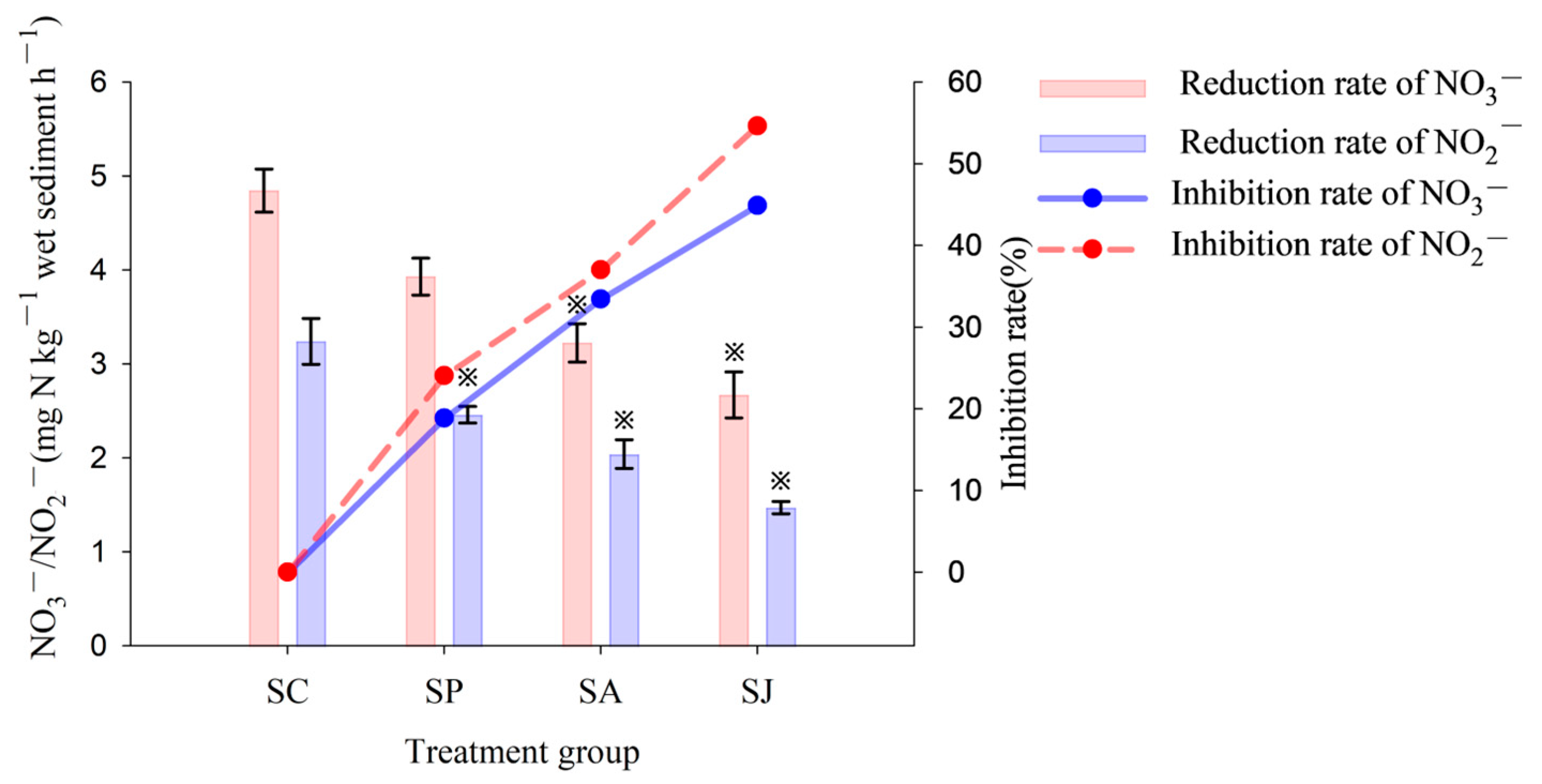
3.1.1. Nitrate and Nitrite Reduction
3.1.2. Potential Denitrification Activity
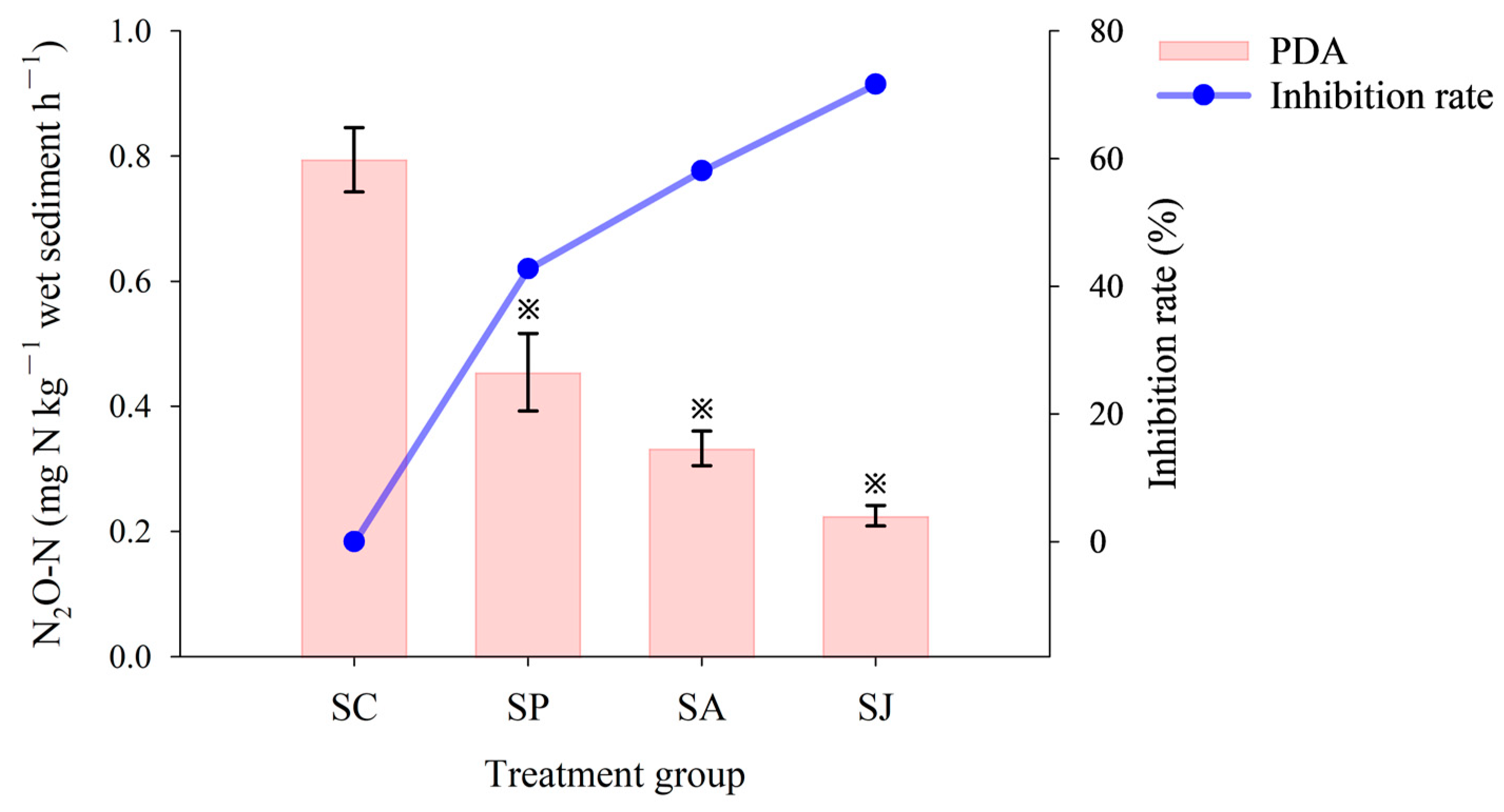
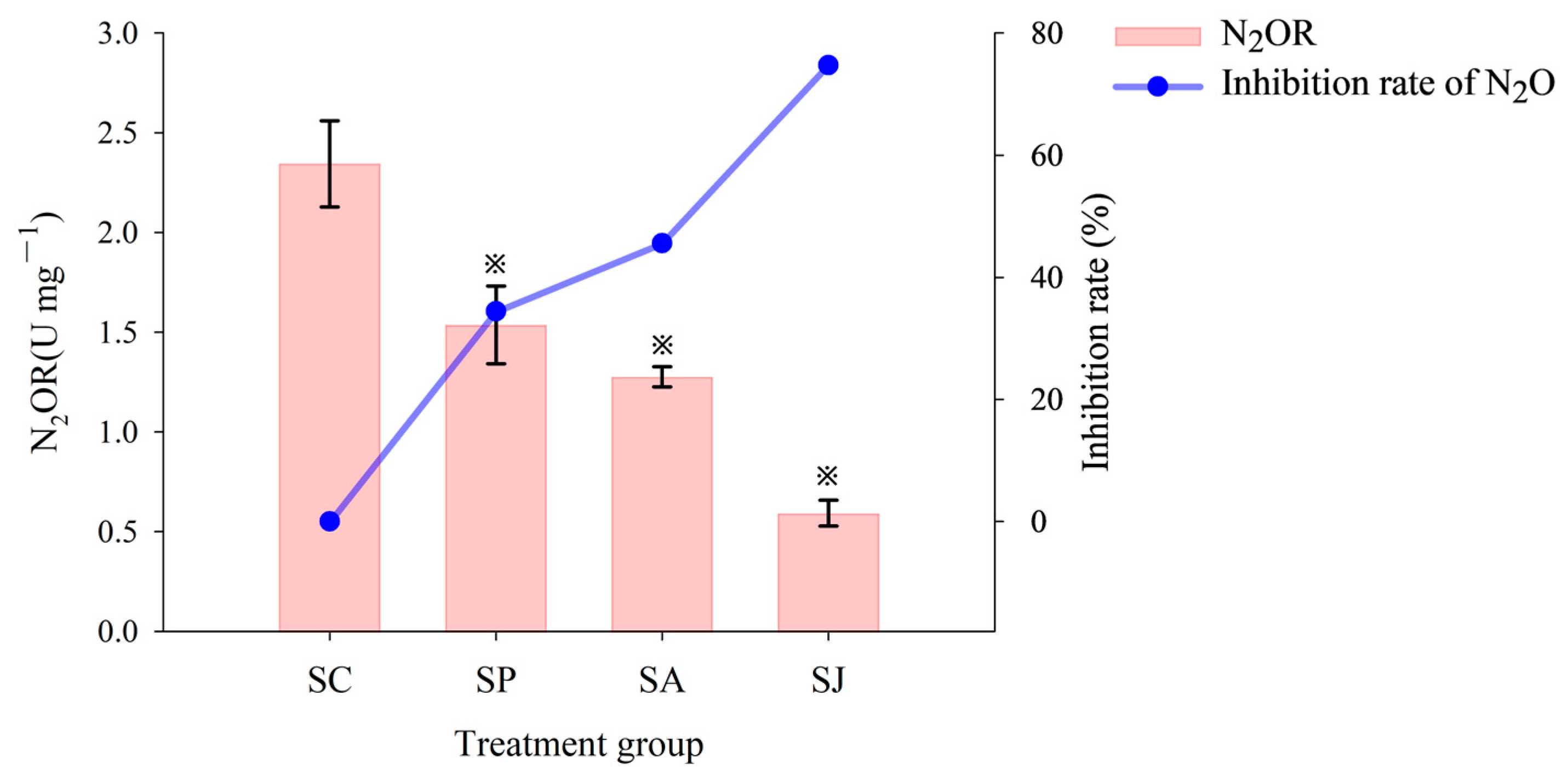
3.1.3. N2O Accumulation and Reduction
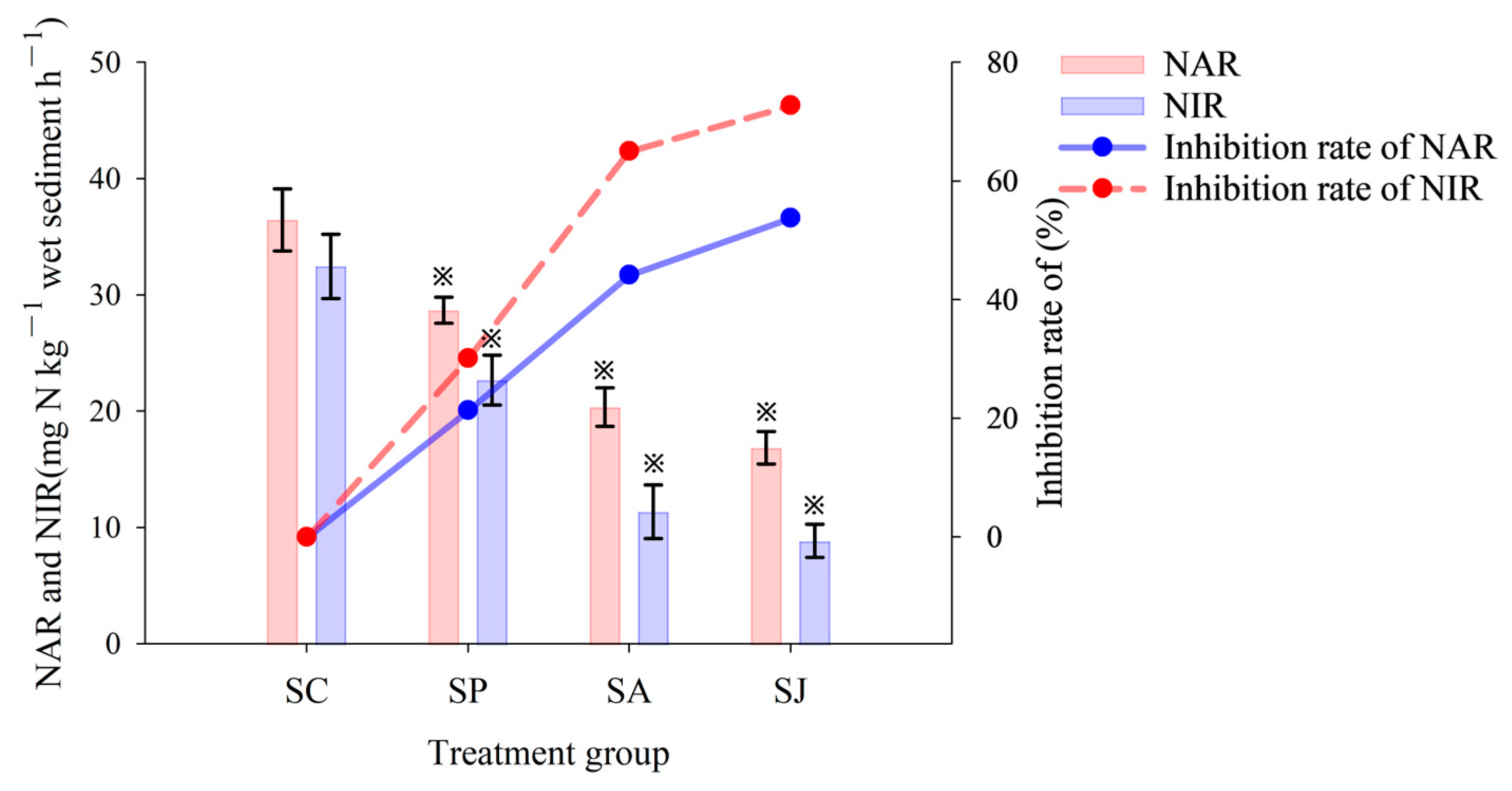
3.2. Effect of Single and Combined Contamination of Phe and AgNPs on Denitrifying Reductase Activities
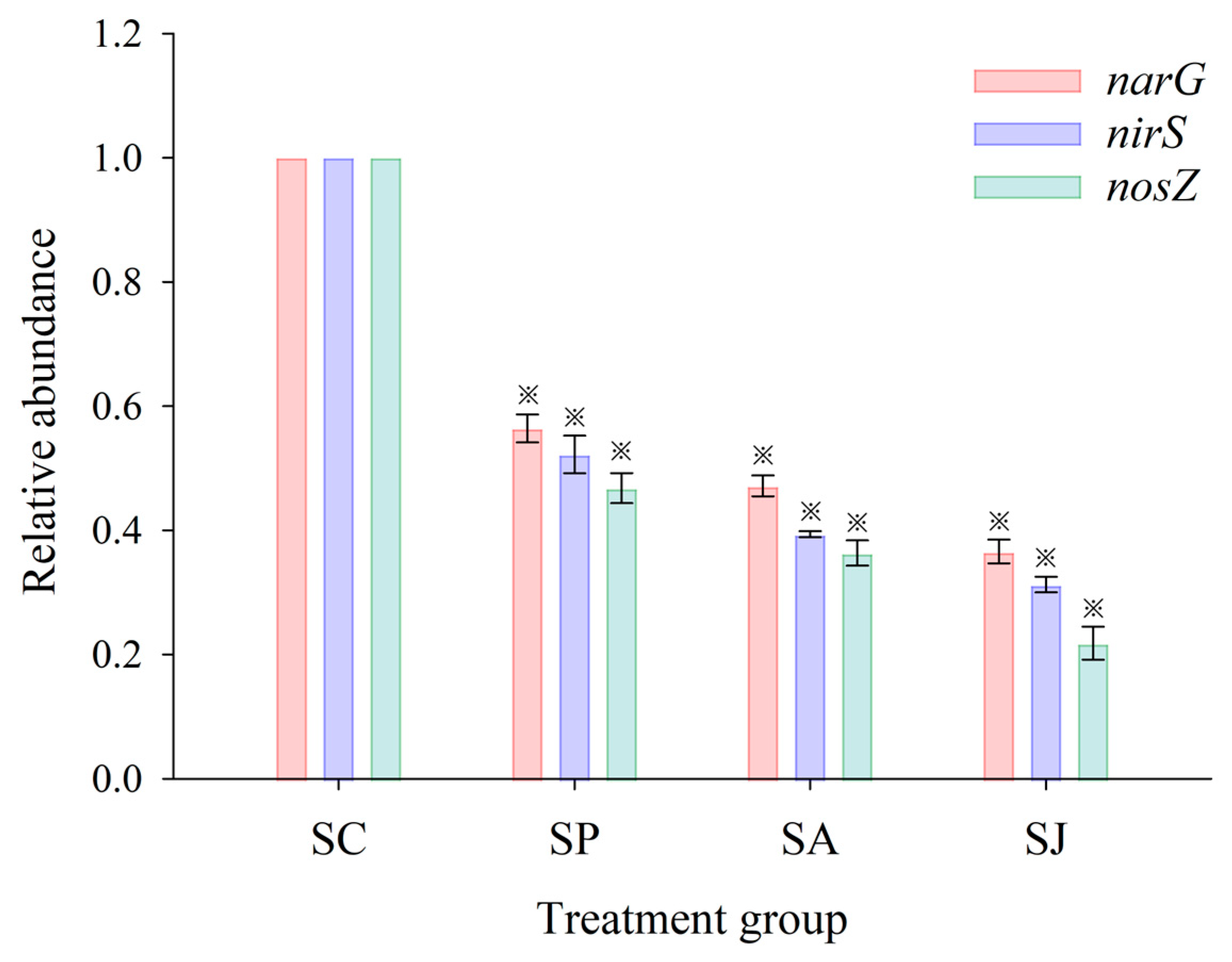
3.3. Effect of Single and Combined Contamination of Phe and AgNPs on Total Bacterial Count and Relative Abundance of Denitrifying Genes
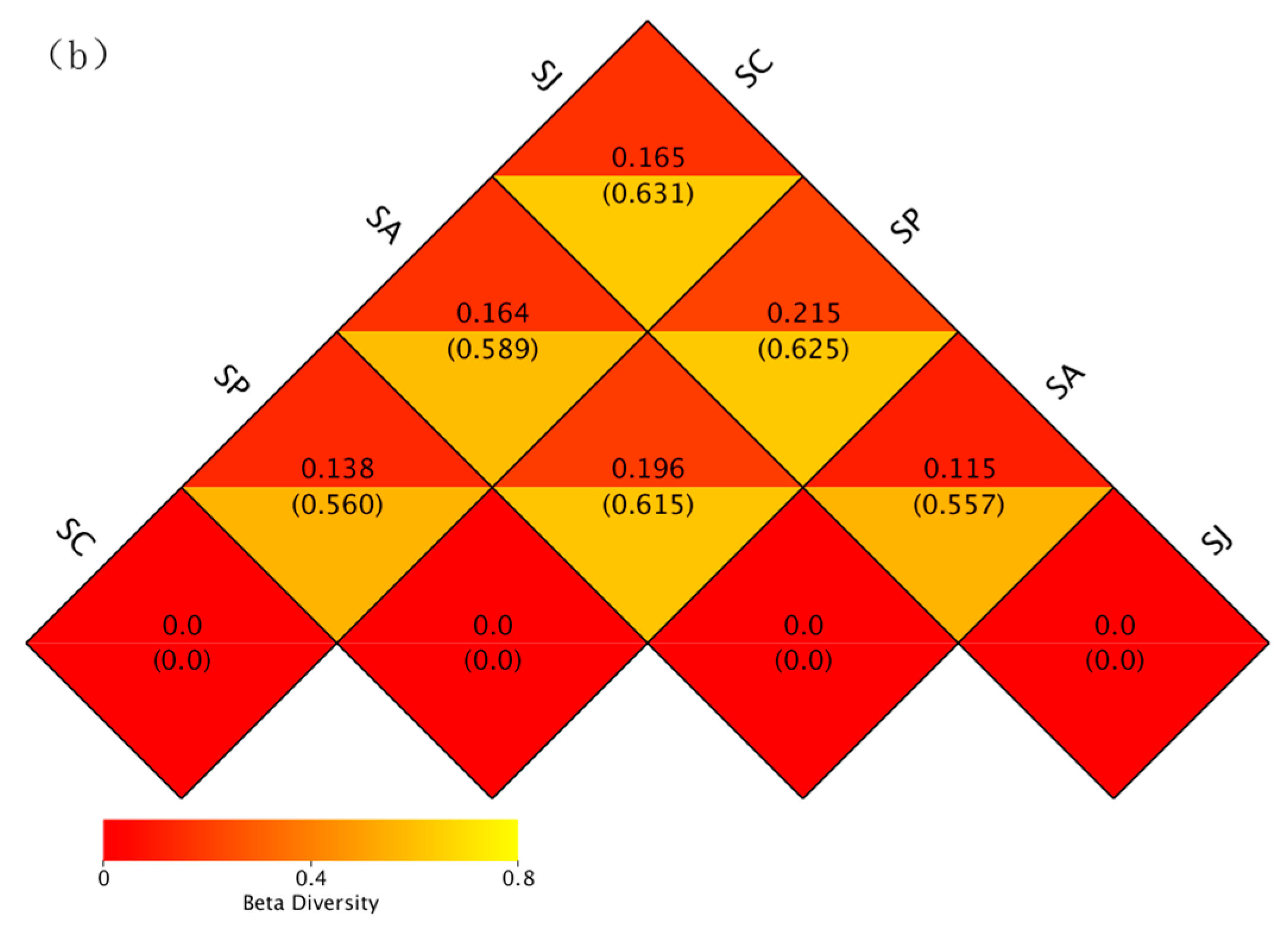
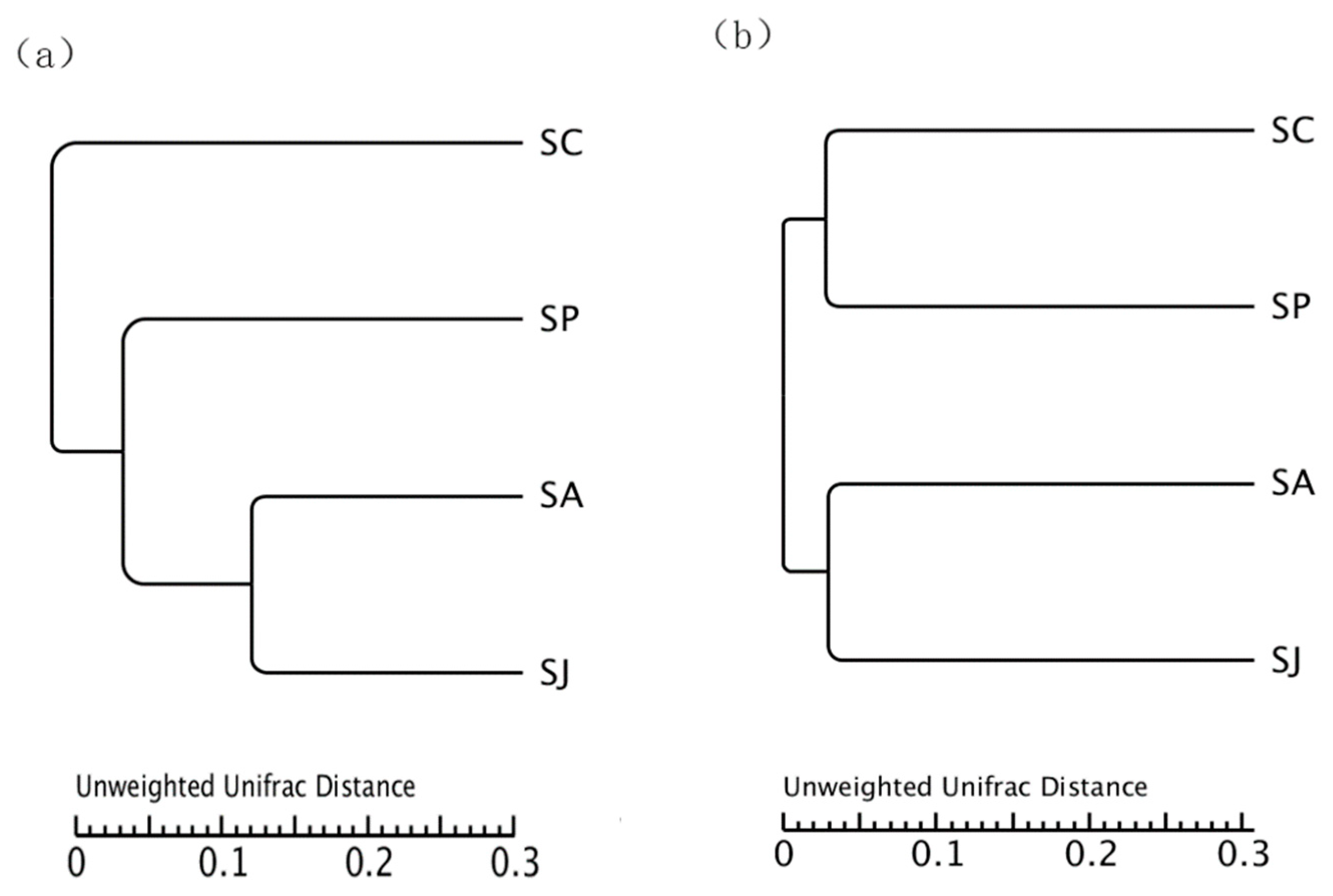
3.4. Community Diversity and Similarity of Denitrifying Bacteria after Single and Combined Exposure to Phe and AgNPs
3.4.1. Alpha Diversity
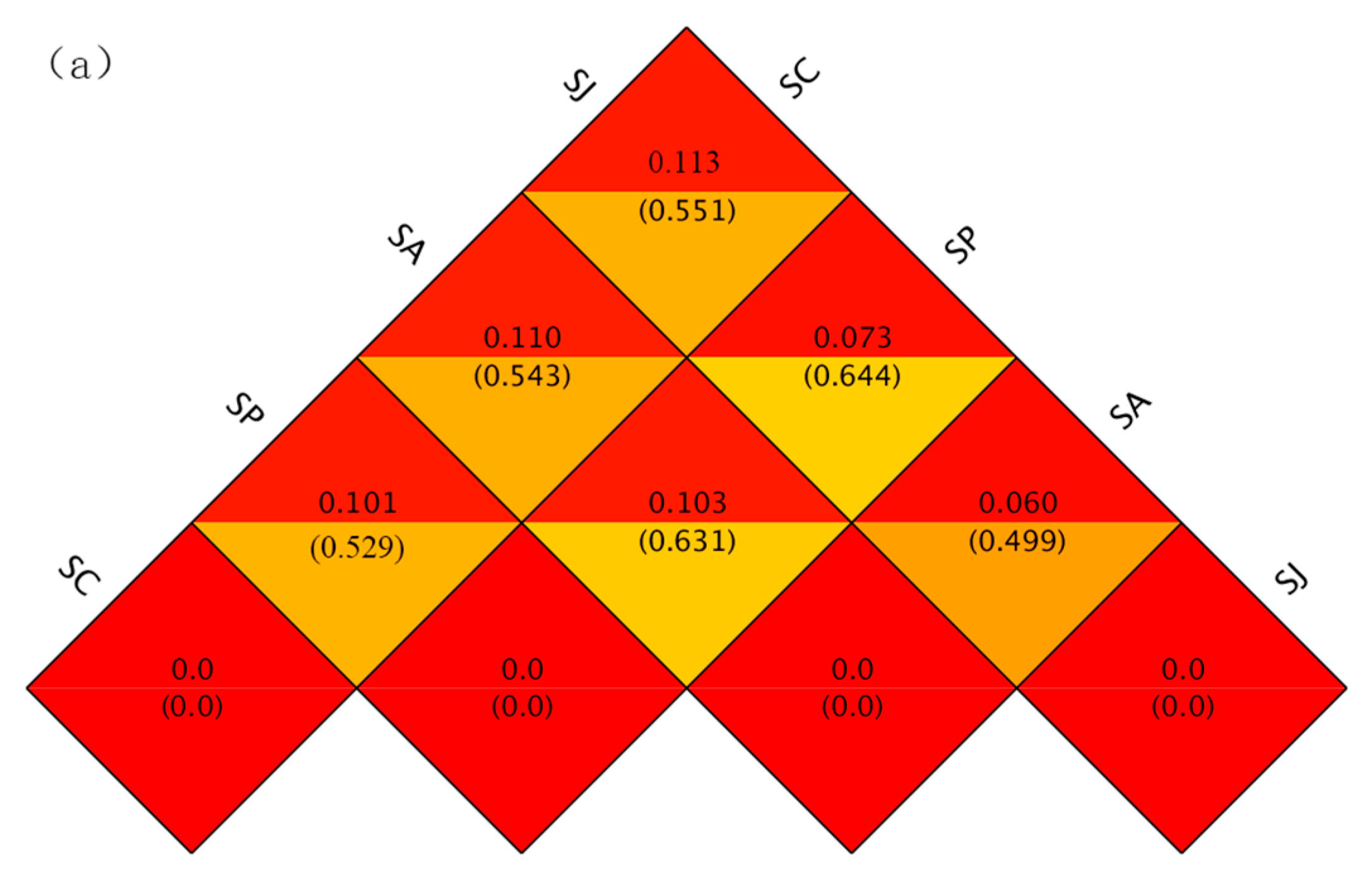
3.4.2. Beta Diversity
3.4.3. UPGMA Cluster Analysis
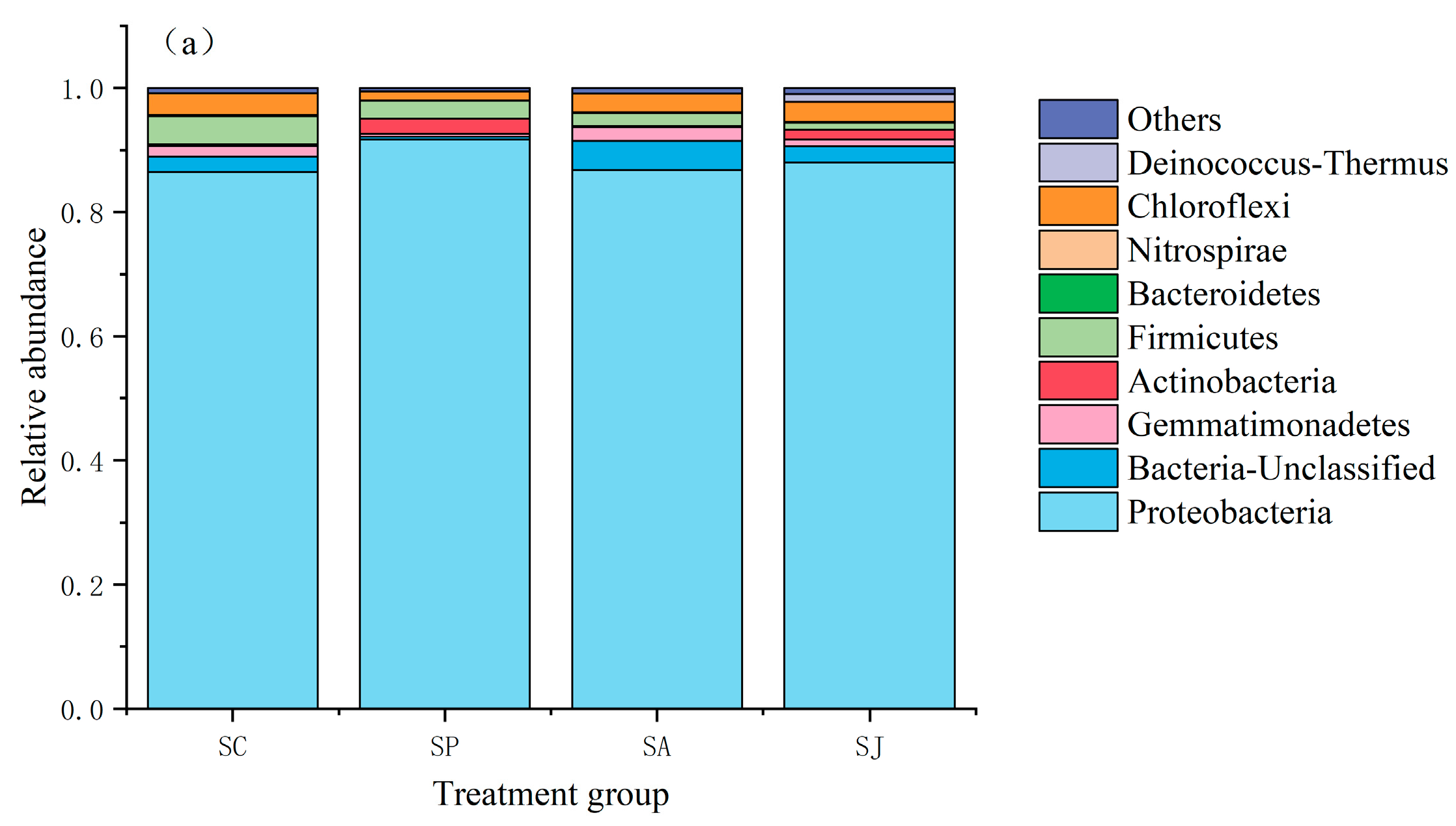
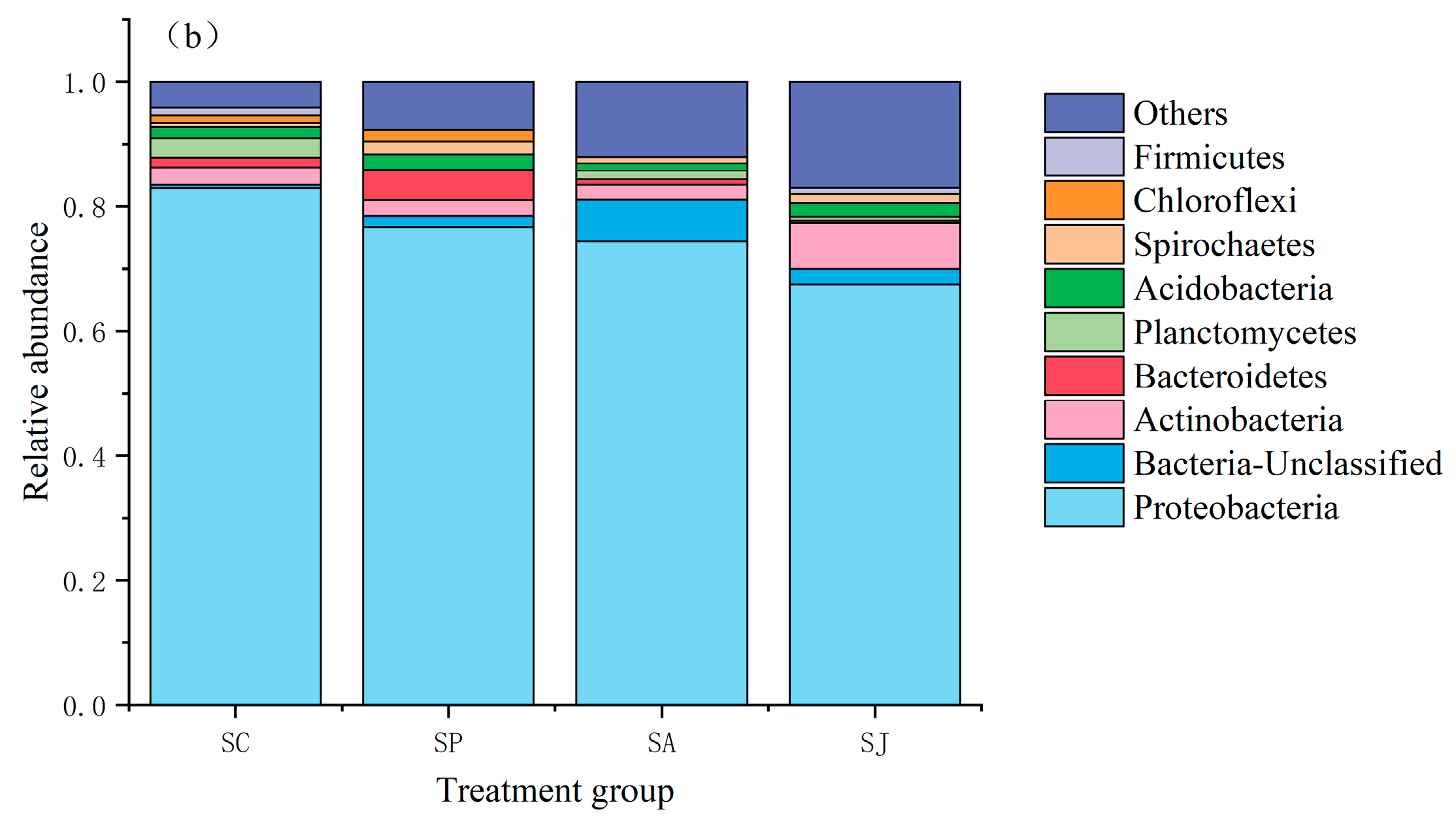
3.5. Community Structure of Denitrifying Bacteria after Single and Combined Contamination of Phenanthrene and AgNPs
3.5.1. Changes in Bacterial Community Structure at Phylum Level in Different Treatment Groups
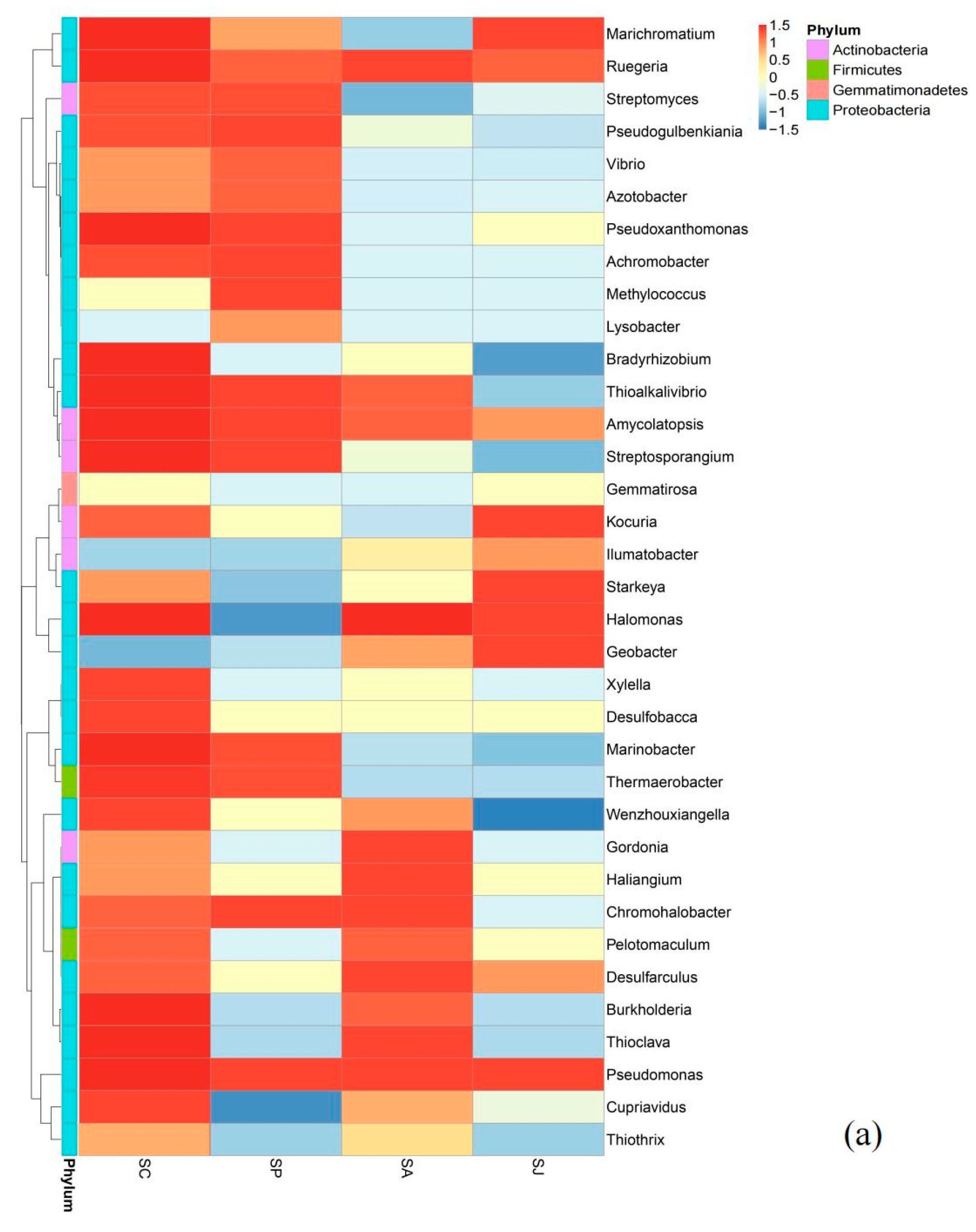
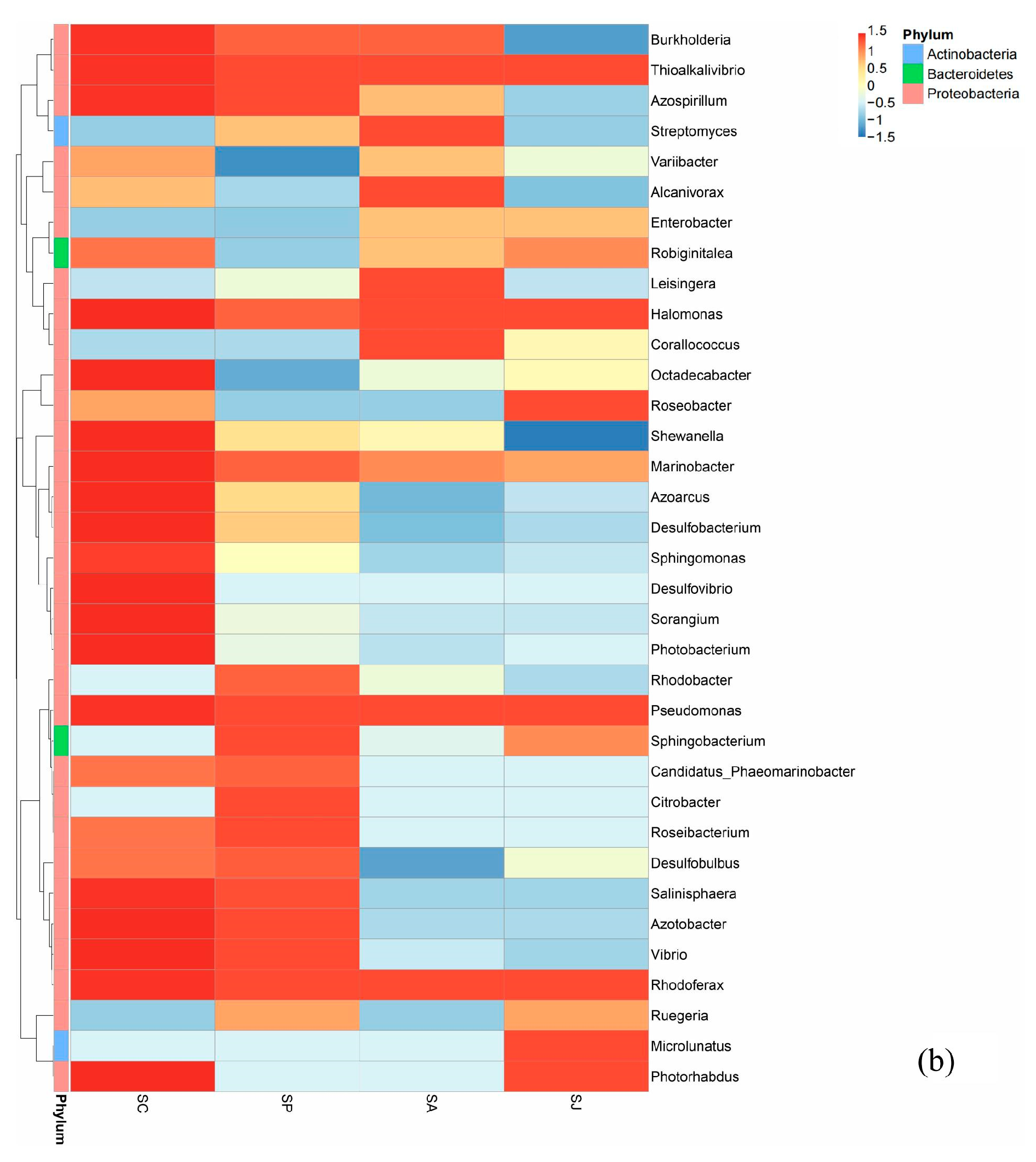
3.5.2. Changes in Bacterial Community Structure at the Genus Level in Different Treatment Groups
3.6. Environmental Implications
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hsiao, S.Y.; Hsu, T.C.; Liu, J.W.; Xie, X.; Zhang, Y.; Lin, J.; Wang, H.; Yang, J.Y.; Hsu, S.C.; Dai, M. Nitrification and its oxygen consumption along the turbid Chang Jiang River plume. Biogeosciences 2014, 11, 2083–2098. [Google Scholar] [CrossRef]
- Rabalais, N.N. Nitrogen in aquatic ecosystems. Ambio 2002, 31, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Liu, T.; Xia, X.; Xia, N. Effect of particle size and composition of suspended sediment on denitrification in river water. Sci. Total Environ. 2016, 541, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Harter, J.; Weigold, P.; El-Hadidi, M.; Huson, D.H.; Kappler, A.; Behrens, S. Soil biochar amendment shapes the composition of N2O-reducing microbial communities. Sci. Total Environ. 2016, 562, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Crutzen, P.J. The influence of nitrogen oxides on the atmospheric ozone content. Q. J. R. Meteorol. Soc. 1970, 96, 320–325. [Google Scholar] [CrossRef]
- Dickinson, R.E.; Cicerone, R.J. Future global warming from atmospheric trace gases. Nature. 1986, 319, 109. [Google Scholar] [CrossRef]
- Backhaus, T.; Blanck, H.; Faust, M. Hazard and Risk Assessment of Chemical Mixtures under REACH: State of the Art, Gaps and Options for Improvement; Swedish Chemicals Agency: Stockholms Lan, Swedish, 2010. [Google Scholar]
- McGillicuddy, E.; Murray, I.; Kavanagh, S.; Morrison, L.; Fogarty, A.; Cormican, M.; Dockery, P.; Prendergast, M.; Rowan, N.; Morris, D. Silver nanoparticles in the environment: Sources, detection and ecotoxicology. Sci. Total Environ. 2017, 575, 231–246. [Google Scholar] [CrossRef]
- Verma, P.; Maheshwari, S.K. Applications of silver nanoparticles in diverse sectors. Int. J. Nano Dimens. 2019, 10, 18–36. [Google Scholar]
- Gottschalk, F.; Sonderer, T.; Scholz, R.W.; Nowack, B. Modeled environmental concentrations of engineered nanomaterials (TiO2, ZnO, Ag, CNT, fullerenes) for different regions. Environ. Sci. Technol. 2009, 43, 9216–9222. [Google Scholar] [CrossRef]
- Griffitt, R.J.; Hyndman, K.; Denslow, N.D.; Barber, D.S. Comparison of molecular and histological changes in zebrafish gills exposed to metallic nanoparticles. Toxicol Sci. 2009, 107, 404–415. [Google Scholar] [CrossRef]
- Wang, X.; Yuan, K.; Chen, B.; Lin, L.; Huang, B.; Luan, T. Monthly variation and vertical distribution of parent and alkyl polycyclic aromatic hydrocarbons in estuarine water column: Role of suspended particulate matter. Environ. Pollut. 2016, 216, 599–607. [Google Scholar] [CrossRef]
- Cao, C.; Huang, J.; Yan, C.N.; Zhang, X.X.; Ma, Y.X. Impacts of Ag and Ag2S nanoparticles on the nitrogen removal within vertical flow constructed wetlands treating secondary effluent. Sci. Total Environ. 2021, 777, 145171. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.X.; Deng, H.; Qiao, M.; Mu, Y.J.; Zhu, Y.G. Effect of pyrene on denitrification activity and abundance and composition of denitrifying community in an agricultural soil. Environ. Pollut. 2011, 159, 1886–1895. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.H.; Wang, M.X.; Zuo, X.H.; Li, Z.L.; Luo, F.; Zhou, Z.F. Effects of PAHs Pollution on the Community Structure of Denitrifiers in a Typical Oilfield. Huanjing Kexue/Environ. Sci. 2016, 37, 4750–4759. [Google Scholar]
- Zhou, Z.F.; Yao, Y.H.; Wang, M.X.; Zuo, X.H. Co-effects of pyrene and nitrate on the activity and abundance of soil denitrifiers under anaerobic condition. Arch. Microbiol. 2017, 199, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Wuebbles, D. Nitrous oxide: No laughing matter. Science 2009, 326, 56–57. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.; Liu, Y.; Li, X.; Hou, J.; Bai, Z.; Wang, L.; Zhao, J. New insights into denitrification and phosphorus removal with degradation of polycyclic aromatic hydrocarbons in two-sludge system. Bioresour. Technol. 2022, 346, 126610. [Google Scholar] [CrossRef]
- VandeVoort, A.R.; Skipper, H.; Arai, Y. Macroscopic assessment of nanosilver toxicity to soil denitrification kinetics. J. Environ. Qual. 2014, 43, 1424–1430. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Hou, L.; Liu, M.; Newell, S.E.; Yin, G.; Yu, C.; Zhang, H.; Li, X.; Gao, D.; Gao, J.; et al. Effects of silver nanoparticles on nitrification and associated nitrous oxide production in aquatic environments. Sci. Adv. 2017, 3, 1603229. [Google Scholar] [CrossRef]
- Liu, S.; Wang, C.; Hou, J.; Wang, P.; Miao, L.; Fan, X.; You, G.; Xu, Y. Effects of Ag and Ag2S nanoparticles on denitrification in sediments. Water Res. 2018, 137, 28–36. [Google Scholar] [CrossRef]
- VandeVoort, A.R.; Arai, Y. Effect of silver nanoparticles on soil denitrification kinetics. Ind. Biotechnol. 2012, 8, 358–364. [Google Scholar] [CrossRef]
- Xu, J.; Bao, S.P.; Xiang, D.F.; Xue, L.; Tang, W.; Fang, T. Effects of silver nanoparticles on denitrification and anammox in sediments of hypertrophic and mesotrophic lakes. Sci. Total Environ. 2023, 858, 159933. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Miao, L.Z.; Li, B.L.; Shan, S.J.; Li, D.P.; Hou, J. Long-term effects of Ag NPs on denitrification in sediment: Importance of Ag NPs exposure ways in aquatic ecosystems. Water Res. 2023, 242, 120283. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.X.; Yin, Y.; Wang, S.H. Effect of nanomaterials on ecotoxicity of phenanthrene in Carassius auratus. Environ. Chem. 2013, 32, 1342–1347. [Google Scholar]
- Schwab, F.; Bucheli, T.D.; Camenzuli, L.; Magrez, A.; Knauer, K.; Sigg, L.; Nowack, B. Diuron sorbed to carbon nanotubes exhibits enhanced toxicity to Chlorella vulgaris. Environ. Sci. Technol. 2013, 47, 7012–7019. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.F.; Bai, J.; Li, K.R.; Zhao, Y.G.; Tian, Y.Z. Impacts of phenanthrene on denitrification activity and transcription of related functional genes in estuarine and marine sediments. J. Ocean Univ. China. 2020, 19, 124–134. [Google Scholar] [CrossRef]
- Sun, P.F.; Li, K.R.; Yi, S.K.; Li, H.; Chen, X. Effects of silver nanoparticles on denitrification and associated N2O release in estuarine and marine sediments. J. Ocean Univ. China. 2022, 21, 131–140. [Google Scholar] [CrossRef]
- Wigginton, N.S.; Haus, K.L.; Hochella, M.F., Jr. Aquatic environmental nanoparticles. J. Environ. Monit. 2007, 9, 1306–1316. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.X.; Ma, Q.M.; Yang, X.N.; Li, Y.J. Characteristic parameters of organic hydrocarbons in the surface sediments of Jiaozhou bay. Mar. Environ. Res. 2017, 36, 216–221. [Google Scholar]
- Fang, H.; Liang, H.; Guosen, Z. Distribution and Sources of Polycyclic Aromatic Hydrocarbons in Surface Sediments from the East China Sea. Earth Environ. 2018, 46, 50–58. [Google Scholar]
- Zhang, Z.; Lo, I.M.C.; Zheng, G.; Woon, K.S.; Rao, P. Effect of autotrophic denitrification on nitrate migration in sulfide-rich marine sediments. J. Soils Sediments. 2015, 15, 1019–1028. [Google Scholar] [CrossRef]
- Sørensen, J. Denitrification rates in a marine sediment as measured by the acetylene inhibition technique. Appl. Environ. Microbiol. 1978, 36, 139–143. [Google Scholar] [CrossRef]
- Joye, S.B.; Smith, S.V.; Hollibaugh, J.T.; Paerl, H.W. Estimating denitrification rates in estuarine sediments: A comparison of stoichiometric and acetylene based methods. Biogeochemistry 1996, 33, 197–215. [Google Scholar] [CrossRef]
- Wigginton, N.S.; de Titta, A.; Piccapietra, F.; Dobias, J.; Nesatyy, V.J.; Suter, M.J.F.; Bernier-Latmani, R. Binding of silver nanoparticles to bacterial proteins depends on surface modifications and inhibits enzymatic activity. Environ. Sci. Technol. 2010, 44, 2163–2168. [Google Scholar] [CrossRef]
- Schug, H.; Isaacson, C.W.; Sigg, L.; Ammann, A.A.; Schirmer, K. Effect of TiO2 nanoparticles and uv radiation on extracellular enzyme activity of intact heterotrophic biofilms. Environ. Sci. Technol. 2014, 48, 11620–11628. [Google Scholar] [CrossRef] [PubMed]
- Kristjansson, J.K.; Hollocher, T.C. First practical assay for soluble nitrous oxide reductase of denitrifying bacteria and a partial kinetic characterization. J. Biol. Chem. 1980, 255, 704–707. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, Y. Reduction of N2O and NO generation in anaerobic−aerobic (low dissolved oxygen) biological wastewater treatment process by using sludge alkaline fermentation liquid. Environ. Sci. Technol. 2011, 45, 2137–2143. [Google Scholar] [CrossRef]
- López-Gutiérrez, J.C.; Henry, S.; Hallet, S.; Martin-Laurent, F.; Catroux, G.; Philippot, L. Quantification of a novel group of nitrate-reducing bacteria in the environment by real-time PCR. J. Microbiol. Methods. 2004, 57, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Hammes, F.A.; Egli, T. New method for assimilable organic carbon determination using flow-cytometric enumeration and a natural microbial consortium as inoculum. Environ. Sci. Technol. 2005, 39, 3289–3294. [Google Scholar] [CrossRef]
- Throbäck, I.N.; Enwall, K.; Jarvis, Å.; Hallin, S. Reassessing PCR primers targeting nirS, nirK and nosZ genes for community surveys of denitrifying bacteria with DGGE. FEMS Microbiol. Ecol. 2004, 49, 401–417. [Google Scholar] [CrossRef]
- Henry, S.; Bru, D.; Stres, B.; Hallet, S.; Philippot, L. Quantitative detection of the nosZ gene, encoding nitrous oxide reductase, and comparison of the abundances of 16S rRNA, narG, nirK, and nosZ genes in soils. Appl. Environ. Microbiol. 2006, 72, 5181–5189. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods. 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Lee, P.H. Shortcut nitrification/denitrification through limited-oxygen supply with two extreme COD/N-and-ammonia active landfill leachates. Chen. En. J. 2021, 404, 126511. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics. 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods. 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods. 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Fan, C.; Liu, G.; Zhang, L.; Shang, J.; Gu, X. Seasonal variation of potential denitrification rates of surface sediment from Meiliang Bay, Taihu Lake, China. J. Environ. Sci-China. 2010, 22, 961–967. [Google Scholar] [CrossRef]
- Magalhães, C.M.; Machado, A.; Matos, P.; Bordalo, A.A. Impact of copper on the diversity, abundance and transcription of nitrite and nitrous oxide reductase genes in an urban European estuary. FEMS Microbiol. Ecol. 2011, 77, 274–284. [Google Scholar] [CrossRef]
- Khalil, M.A.K.; Rasmussen, R.A. Nitrous oxide: Trends and global mass balance over the last 3000 years. Ann. Glaciol. 1988, 10, 73–79. [Google Scholar] [CrossRef]
- Philippot, L.; Hallin, S. Finding the missing link between diversity and activity using denitrifying bacteria as a model functional community. Curr. Opin. Microbiol. 2005, 8, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Ramos, S.M.; Alvarez-Bernal, D.; Montes-Molina, J.A.; Van Cleemput, O.; Dendooven, L. Emission of nitrous oxide from hydrocarbon contaminated soil amended with waste water sludge and earthworms. Appl. Soil Ecol. 2009, 41, 69–76. [Google Scholar] [CrossRef]
- Alburquerque, J.A.; Sánchez-Monedero, M.A.; Roig, A.; Cayuela, M.L. High concentrations of polycyclic aromatic hydrocarbons (naphthalene, phenanthrene and pyrene) failed to explain biochar’s capacity to reduce soil nitrous oxide emissions. Environ. Pollut. 2015, 196, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Warshawsky, D. Metabolic activation of polycyclic and heterocyclic aromatic hydrocarbons and DNA damage: A review. Toxicol. Appl. Pharmacol. 2005, 206, 73–93. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Xia, Q.; Yan, J.; Herreno-Saenz, D.; Wu, Y.S.; Tang, I.W.; Fu, P.P. Photoirradiation of polycyclic aromatic hydrocarbons with UVA light–a pathway leading to the generation of reactive oxygen species, lipid peroxidation, and DNA damage. Int. J. Environ. Res. Public Health. 2006, 3, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Sotiriou, G.A.; Pratsinis, S.E. Antibacterial activity of nanosilver ions and particles. Environ. Sci. Technol. 2010, 44, 5649–5654. [Google Scholar] [CrossRef] [PubMed]
- You, C.; Han, C.; Wang, X.; Zheng, Y.; Li, Q.; Hu, X.; Sun, H. The progress of silver nanoparticles in the antibacterial mechanism, clinical application and cytotoxicity. Mol. Biol. Rep. 2012, 39, 9193–9201. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Tak, Y.K.; Song, J.M. Does the antibacterial activity of silver nanoparticles depend on the shape of the nanoparticle? A study of the gram-negative bacterium Escherichia coli. Appl. Environ. Microbiol. 2007, 73, 1712–1720. [Google Scholar] [CrossRef] [PubMed]
- Zou, Q.Y. Composite Toxic Effects and Mechanisms of AgNPs and Phenol on Pure Denitrifying Bacteria. Master’s Thesis, Harbin Institute of Technology, Harbin, China, 2015. [Google Scholar]
- Guo, L.Y.; Shi, F.; Yang, L.Y. Advances in functional genes and molecular ecology in denitrifiers. Microbiol. China 2011, 38, 583–590. [Google Scholar]
- Beddow, J.; Stölpe, B.; Cole, P.A.; Lead, J.R.; Sapp, M.; Lyons, B.P.; McKew, B.; Steinke, M.; Benyahia, F.; Colbeck, I.; et al. Estuarine sediment hydrocarbon-degrading microbial communities demonstrate resilience to nanosilver. Int. Biodeter. Biodegr. 2014, 96, 206–215. [Google Scholar] [CrossRef]
- Sikkema, J.; de Bont, J.A.; Poolman, B. Interactions of cyclic hydrocarbons with biological membranes. J. Biol. Chem. 1994, 269, 8022–8028. [Google Scholar] [CrossRef] [PubMed]
- Gui, M.; Chen, Q.; Ma, T.; Zheng, M.; Ni, J. Effects of heavy metals on aerobic denitrification by strain Pseudomonas stutzeri PCN-1. Appl. Microbiol. Biotechnol. 2017, 101, 1717–1727. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Wang, H.; Zhang, W.C.; Xie, Z.C.; Fang, T. An investigation into the effects of silver nanoparticles on natural microbial communities in two freshwater sediments. Environ. Pollut. 2016, 219, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.C.; Jones, K.C. Bioremediation of soil contaminated with polynuclear aromatic hydrocarbons (PAHs): A review. Environ. Pollut 1993, 81, 229–249. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.H.; Tang, W.H.; Li, S.Y.; Hong, J. Isolation, identification and degradation ability of phenanthrene-degrading bacteria from contaminated soils. Res. Environ. Sci. 2009, 22, 675–681. [Google Scholar]
- Zhao, B.; Wang, H.; Li, R.; Mao, X. Thalassospira xianhensis sp. nov., a polycyclic aromatic hydrocarbon-degrading marine bacterium. Int. J. Syst. Evol. Micr. 2010, 60, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Sobolev, D.; Begonia, M. Effects of heavy metal contamination upon soil microbes: Lead-induced changes in general and denitrifying microbial communities as evidenced by molecular markers. Int. J. Environ. Res. Public Health 2008, 5, 450–456. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Wu, Q.H.; Huang, S.; Ye, J.X.; Zhang, H.J.; Zhang, R.D. Effect of polycyclic aromatic hydrocarbons on the vertical distribution of denitrifying genes in river sediments. Huan Jing Ke Xue 2012, 33, 3592–3597. [Google Scholar] [PubMed]
- Zheng, X.; Su, Y.; Chen, Y.; Wan, R.; Liu, K.; Li, M.; Yin, D. Zinc oxide nanoparticles cause inhibition of microbial denitrification by affecting transcriptional regulation and enzyme activity. Environ. Sci. Technol. 2014, 48, 13800–13807. [Google Scholar] [CrossRef]
- Yu, W.; Liu, R.; Wang, J.; Xu, F.; Shen, Z. Source apportionment of PAHs in surface sediments using positive matrix factorization combined with GIS for the estuarine area of the Yangtze River, China. Chemosphere 2015, 134, 263–271. [Google Scholar] [CrossRef]
- Li, B.; Zhang, X.; Guo, F.; Wu, W.; Zhang, T. Characterization of tetracycline resistant bacterial community in saline activated sludge using batch stress incubation with high-throughput sequencing analysis. Water Res. 2013, 47, 4207–4216. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Hu, Z.; Li, P.; Gajaraj, S. Governing factors affecting the impacts of silver nanoparticles on wastewater treatment. Sci. Total Environ. 2016, 572, 852–873. [Google Scholar] [CrossRef] [PubMed]
- Bamborough, L.; Cummings, S.P. The impact of increasing heavy metal stress on the diversity and structure of the bacterial and actinobacterial communities of metallophytic grassland soil. Biol. Fertil. Soils. 2009, 45, 273–280. [Google Scholar] [CrossRef]
- Xie, Y.; Fan, J.; Zhu, W.; Amombo, E.; Chen, L. Effect of heavy metals pollution on soil microbial diversity and bermudagrass genetic variation. Front. Plant Sci. 2016, 7, 755. [Google Scholar] [CrossRef] [PubMed]
- Samarajeewa, A.D.; Velicogna, J.R.; Princz, J.I.; Subasinghe, R.M.; Scroggins, R.P.; Beaudette, L.A. Effect of silver nano-particles on soil microbial growth, activity and community diversity in a sandy loam soil. Environ. Pollut. 2017, 220, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Wertz, S.; Degrange, V.; Prosser, J.I.; Poly, F.; Commeaux, C.; Freitag, T.; Guillaumaud, N.; Le Roux, X. Maintenance of soil functioning following erosion of microbial diversity. Environ. Microbiol. 2006, 8, 2162–2169. [Google Scholar] [CrossRef]
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Legendre, P.; De Cáceres, M. Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning. Ecol. Lett. 2013, 16, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative β diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef]
- Idris, R.; Trifonova, R.; Puschenreiter, M.; Wenzel, W.W.; Sessitsch, A. Bacterial communities associated with flowering plants of the Ni hyperaccumulator Thlaspi goesingense. Appl Env. Microbiol. 2004, 70, 2667–2677. [Google Scholar] [CrossRef]
- Spain, A.M.; Krumholz, L.R.; Elshahed, M.S. Abundance, composition, diversity and novelty of soil Proteobacteria. ISME J. 2009, 3, 992. [Google Scholar] [CrossRef] [PubMed]
- Azarbad, H.; Niklinska, M.; Laskowski, R.; van Straalen, N.M.; van Gestel, C.A.; Zhou, J.; He, Z.; Wen, C.; Röling, W.F. Microbial community composition and functions are resilient to metal pollution along two forest soil gradients. FEMS Microbiol Ecol. 2015, 91, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Li, M.; Li, F.; Li, H.; Gao, Z. Distinct distribution patterns of prokaryotes between sediment and water in the Yellow River estuary. Appl. Microbiol. Biotechnol. 2016, 100, 9683–9697. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.F.; Chen, J.Y.; Chen, G.L.; Guan, D.M. Study on the Quantity and Diversity of Denitrifying Bacteria in Sediments of Liaohe Estuary. Acta Oceanol. Sin. Chin. Ed. 2011, 33, 94–102. [Google Scholar]
- Li, W.; Luo, D.; Adyel, T.M.; Wu, J.; Miao, L.; Kong, M.; Hou, J. Dynamic responses of carbon metabolism of sediment microbial communities to Ag nanoparticles: Effects of the single and repeated exposure scenarios. Sci. Total Environ. 2023, 870, 161891. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Xue, L.; Xiang, D.; Xian, B.; Tang, W.; Fang, T. Silver nanoparticles alter the bacterial assembly and antibiotic resistome in biofilm during colonization. Environ. Science-Nano 2023, 10, 656–671. [Google Scholar] [CrossRef]
- Rees, A.P.; Faraggiana, E.; Tait, K.; Celussi, M.; Dafnomilli, E.; Manna, V.; Manning, A.; Pitta, P.; Tsiola, A.; Živanović, S. The presence of silver nanoparticles reduces demand for dissolved phosphorus to the benefit of biological nitrogen fixation in the coastal eastern Mediterranean Sea. Front. Mar. Sci. 2022, 9, 907269. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, X.; Wang, Y.; Duan, L.; Liu, X.; Zhang, X.; Dong, L. Deep relationships between bacterial community and polycyclic aromatic hydrocarbons in soil profiles near typical coking plants. Environ. Sci. Pollut. R. 2023, 30, 64486–64498. [Google Scholar] [CrossRef]
- Haritash, A.K.; Kaushik, C.P. Biodegradation aspects of polycyclic aromatic hydrocarbons (PAHs): A review. J. Hazard. Mater. 2009, 169, 1–15. [Google Scholar] [CrossRef]
- Krzyszczak, A.; Dybowski, M.P.; Zarzycki, R.; Kobyłecki, R.; Oleszczuk, P.; Czech, B. Long-term physical and chemical aging of biochar affected the amount and bioavailability of PAHs and their derivatives. J. Hazard. Mater. 2022, 440, 129795. [Google Scholar] [CrossRef]
- Jin, B.; Niu, J.; Liu, Y.; Zhao, J.; Yin, Z. Effects of polycyclic aromatic hydrocarbons on sludge performance for denitrification and phosphorus removal. Chem. Eng. J. 2020, 397, 125552. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Y.P.; Liu, S.Y.; Guo, J.S.; Fang, F.; Yan, P. The effect of silver nanoparticles on aerobic denitrifying bacteria during biological nitrogen removal: A new perspective based on morphological effects. Chem. Eng. J. 2023, 471, 144538. [Google Scholar] [CrossRef]
- Zhang, Z.Z.; Cheng, Y.F.; Xu, L.Z.J.; Bai, Y.H.; Jin, R.C. Anammox granules show strong resistance to engineered silver nanoparticles during long-term exposure. Bioresour. Technol. 2018, 259, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Premnath, N.; Mohanrasu, K.; Rao, R.G.R.; Dinesh, G.H.; Prakash, G.S.; Ananthi, V.; Ponnuchamy, K.; Muthusamy, G.; Arun, A.A. Crucial Review on Polycyclic Aromatic Hydrocarbons—Environmental Occurrence and Strategies for Microbial Degradation. Chemosphere 2021, 280, 130608. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sample | OTUs | Shannon | Simpson | Chao1 | Ace | Coverage |
|---|---|---|---|---|---|---|---|
| nirS | SC | 454 | 2.54 | 0.309 | 509 | 519 | 0.999 |
| SP | 405 | 1.60 | 0.680 | 409 | 425 | 0.999 | |
| SA | 442 | 1.89 | 0.637 | 501 | 508 | 0.999 | |
| SJ | 399 | 2.28 | 0.552 | 448 | 460 | 0.999 | |
| nosZ | SC | 1038 | 3.39 | 0.774 | 1962 | 1538 | 0.996 |
| SP | 964 | 3.14 | 0.793 | 873 | 931 | 0.995 | |
| SA | 836 | 2.98 | 0.837 | 1195 | 1239 | 0.997 | |
| SJ | 721 | 3.25 | 0.776 | 1064 | 1009 | 0.997 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, P.; Bai, J.; Lian, J.; Tan, Y.; Chen, X. Single and Combined Effects of Phenanthrene and Silver Nanoparticles on Denitrification Processes in Coastal Marine Sediments. Microorganisms 2024, 12, 745. https://doi.org/10.3390/microorganisms12040745
Sun P, Bai J, Lian J, Tan Y, Chen X. Single and Combined Effects of Phenanthrene and Silver Nanoparticles on Denitrification Processes in Coastal Marine Sediments. Microorganisms. 2024; 12(4):745. https://doi.org/10.3390/microorganisms12040745
Chicago/Turabian StyleSun, Pengfei, Jie Bai, Jie Lian, Yongyu Tan, and Xi Chen. 2024. "Single and Combined Effects of Phenanthrene and Silver Nanoparticles on Denitrification Processes in Coastal Marine Sediments" Microorganisms 12, no. 4: 745. https://doi.org/10.3390/microorganisms12040745