

Analysis of Leaf and Soil Nutrients, Microorganisms and Metabolome in the Growth Period of Idesia polycarpa Maxim
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Material
2.3. Observation of Root Morphology Indicators
2.4. Soil and Leaf Sample Collection and Determination of Physical and Chemical Properties
2.5. Determination of Leaf and Soil Microorganisms
2.5.1. DNA Extraction, PCR Amplification, and Library Preparation
2.5.2. Bioinformatics Analysis
2.6. Metabolome Analysis
2.6.1. Metabolite Extraction
2.6.2. The UPLC-MS/MS Analysis
3. Results
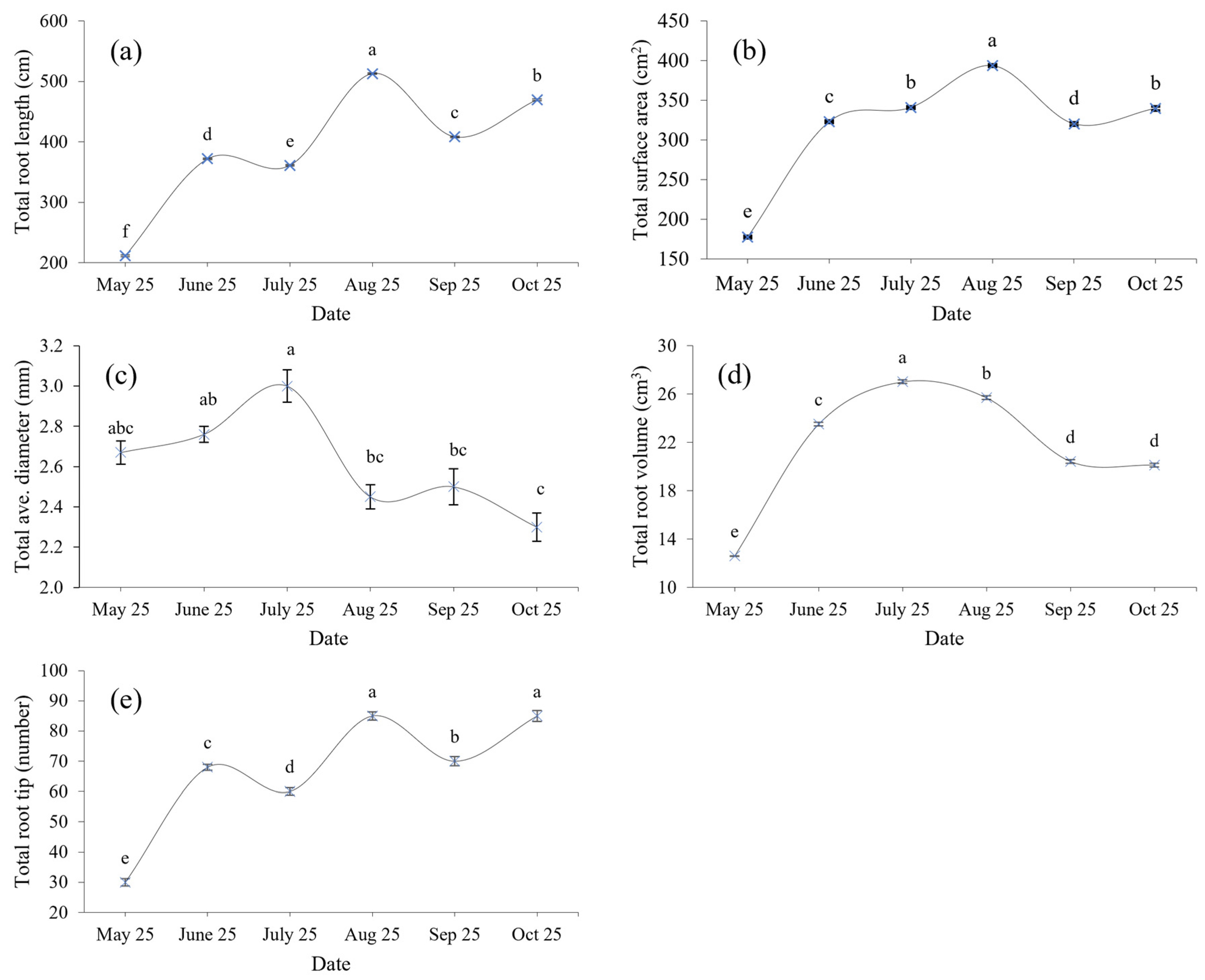
3.1. Characteristics of Fine Roots
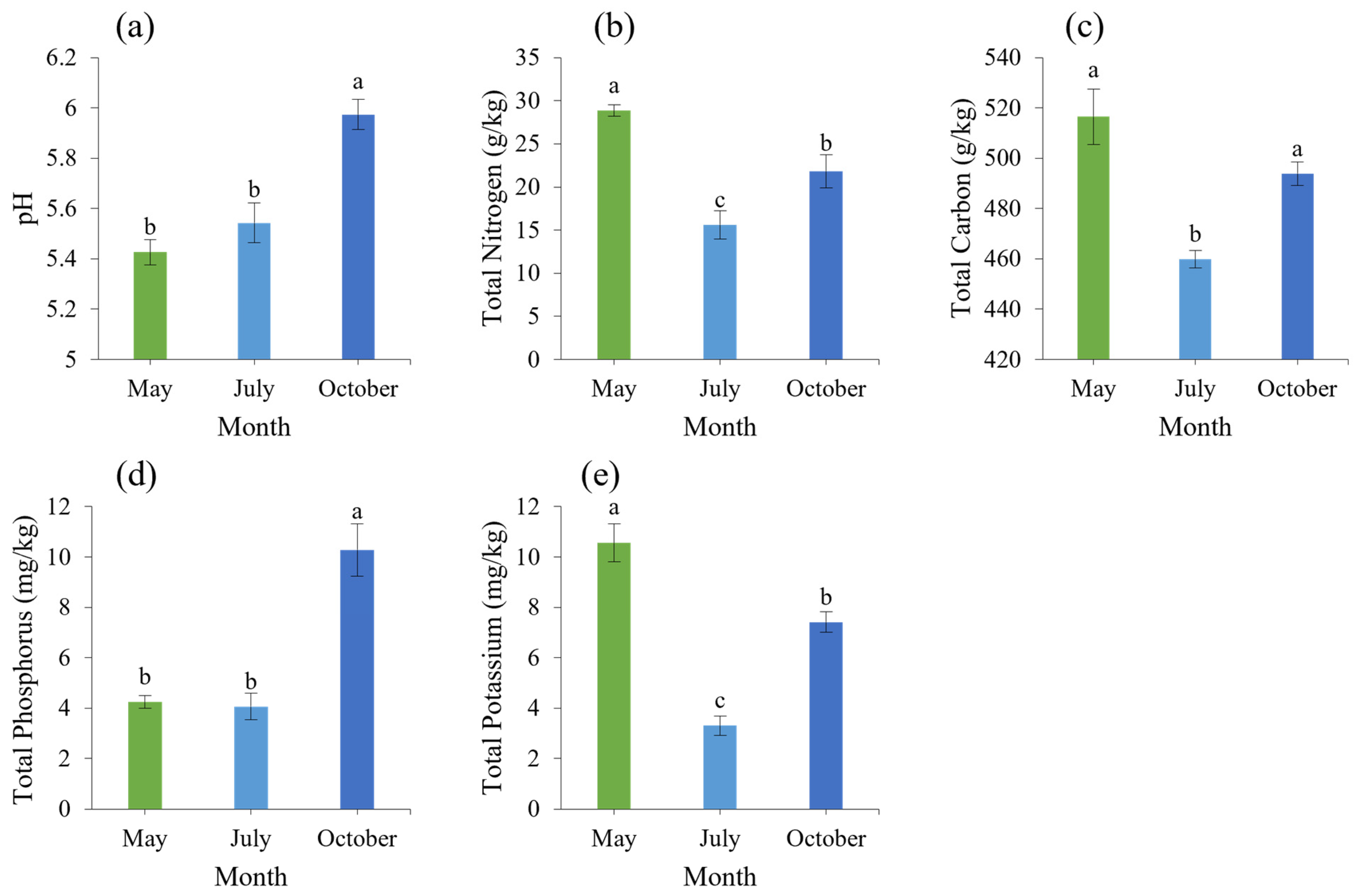
3.2. Soil Physical and Chemical Properties Analysis
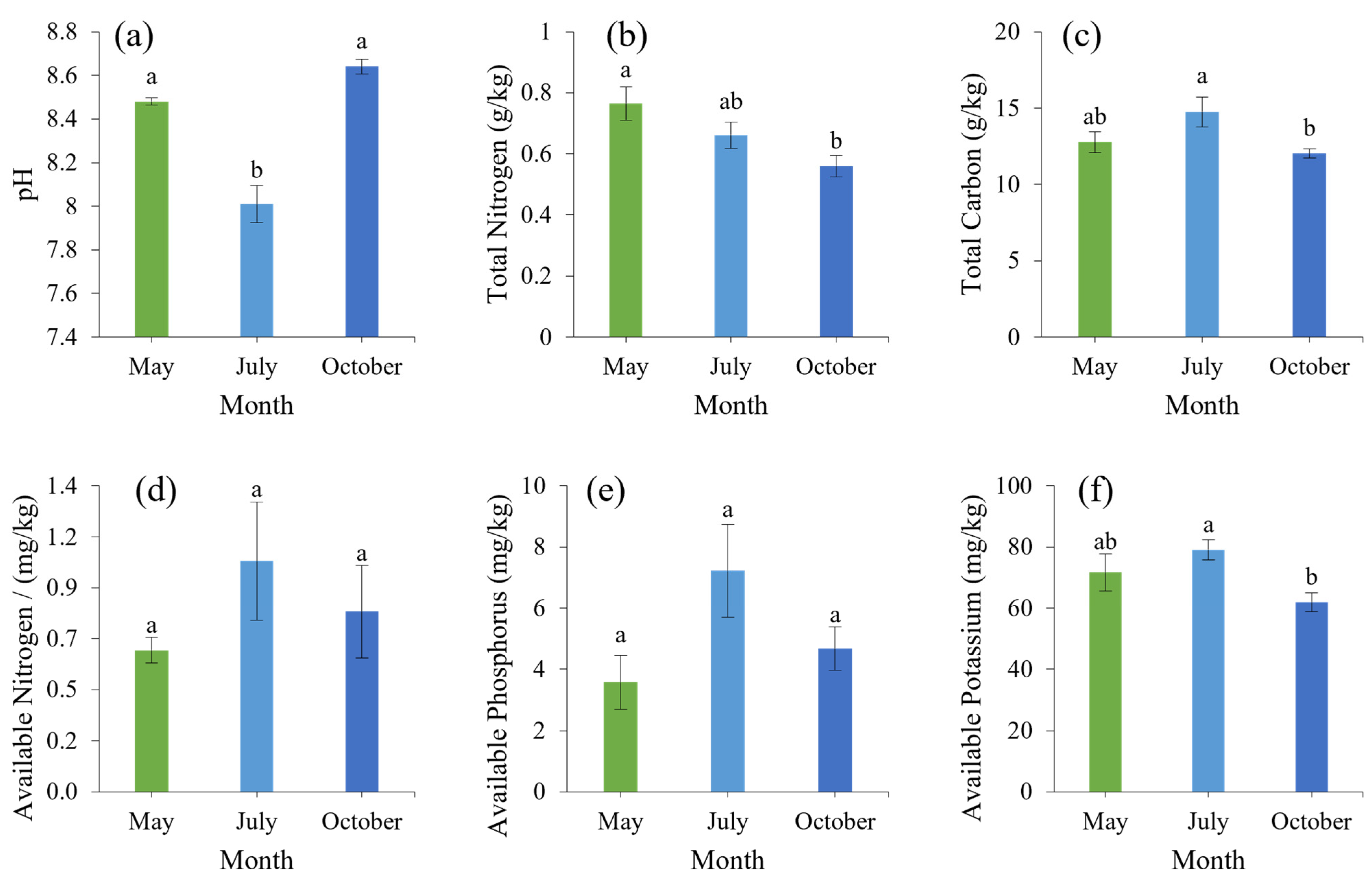
3.3. Leaves Physical and Chemical Properties Analysis
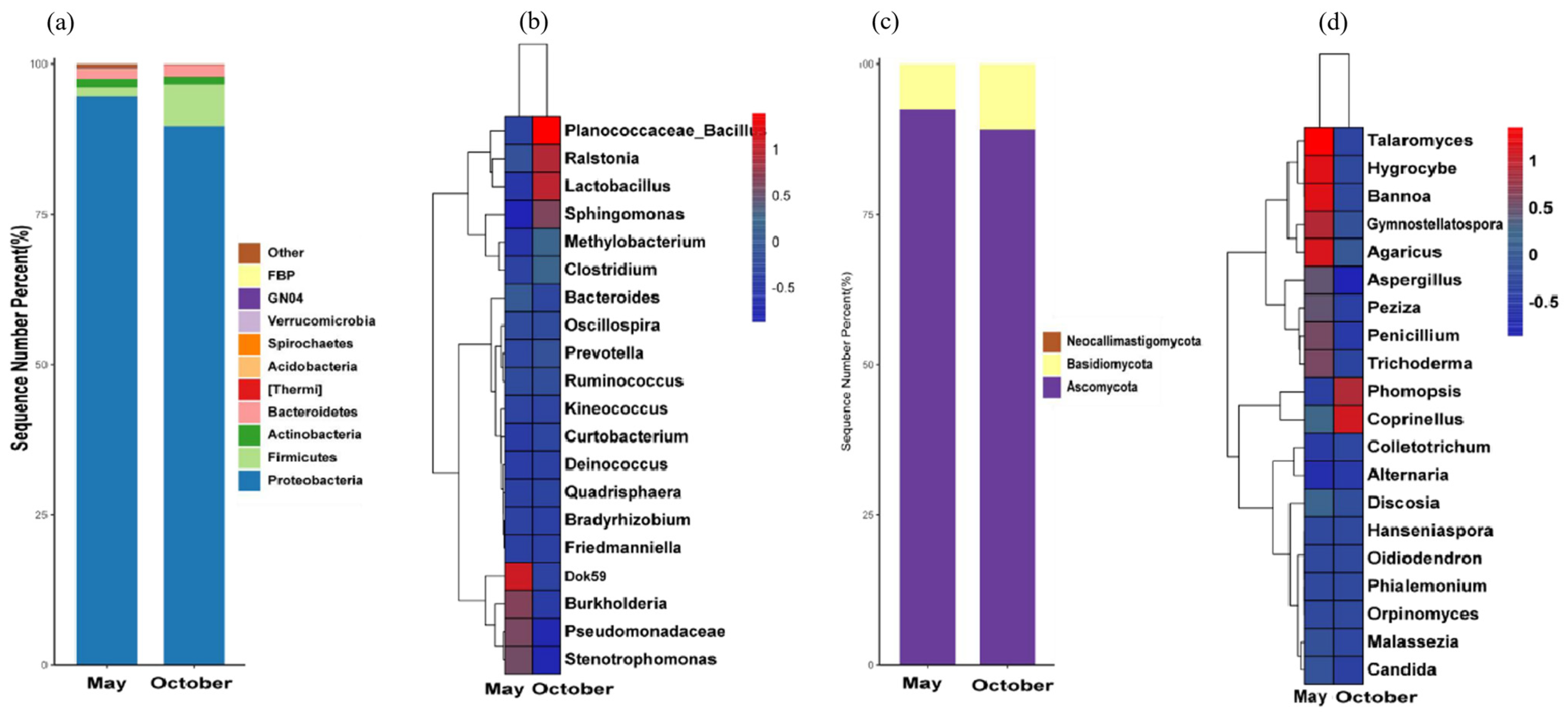
3.4. Analysis of Soil Microbial Diversity in the Rhizosphere
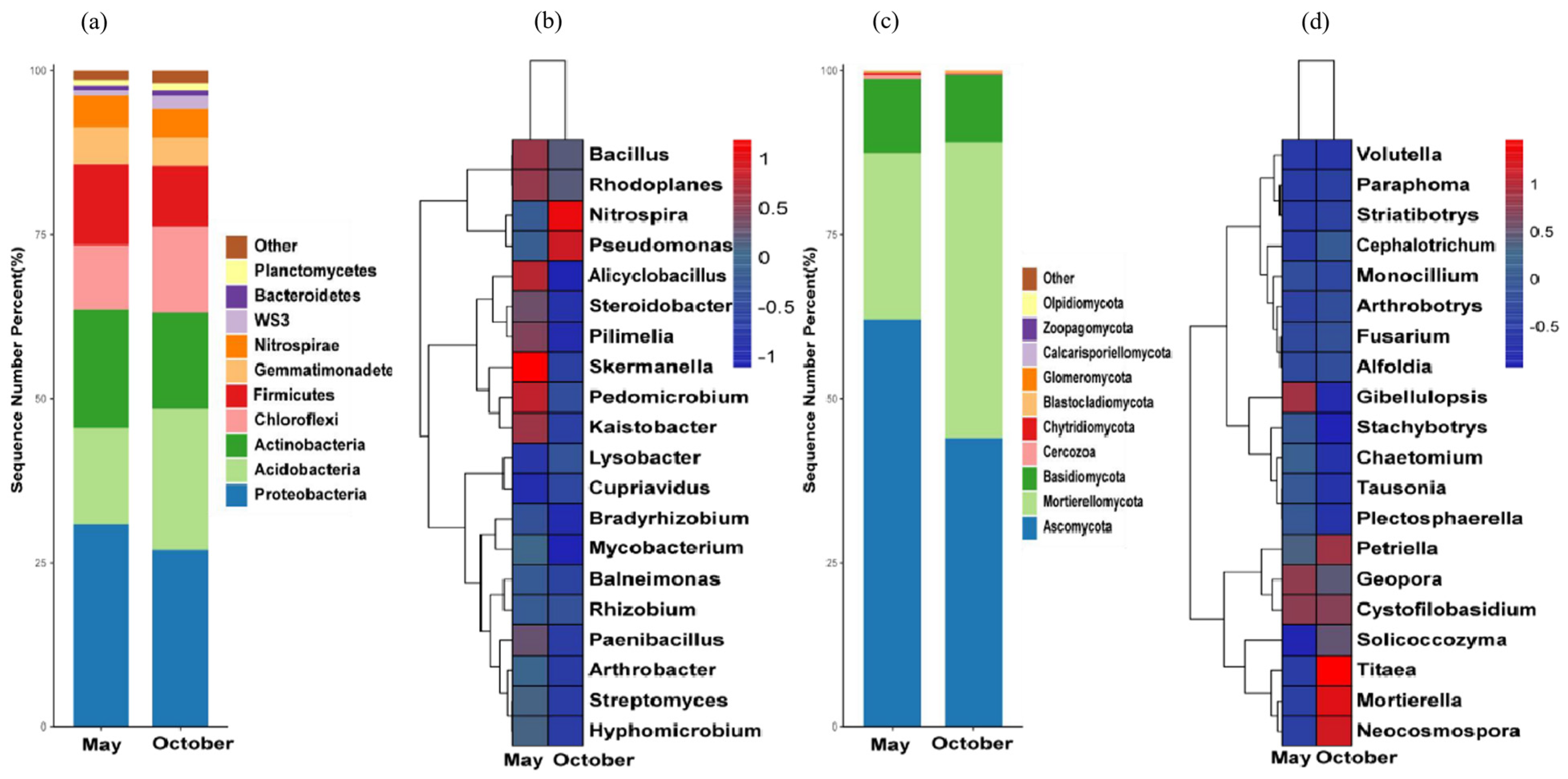
3.4.1. Analysis of Soil Bacterial Diversity in the Rhizosphere
3.4.2. Analysis of Soil Fungal Diversity of Rhizosphere
3.5. Analysis of Leaf Microbial Diversity
3.5.1. Analysis of Leaf Bacteria Diversity
3.5.2. Analysis of Leaf Fungal Diversity
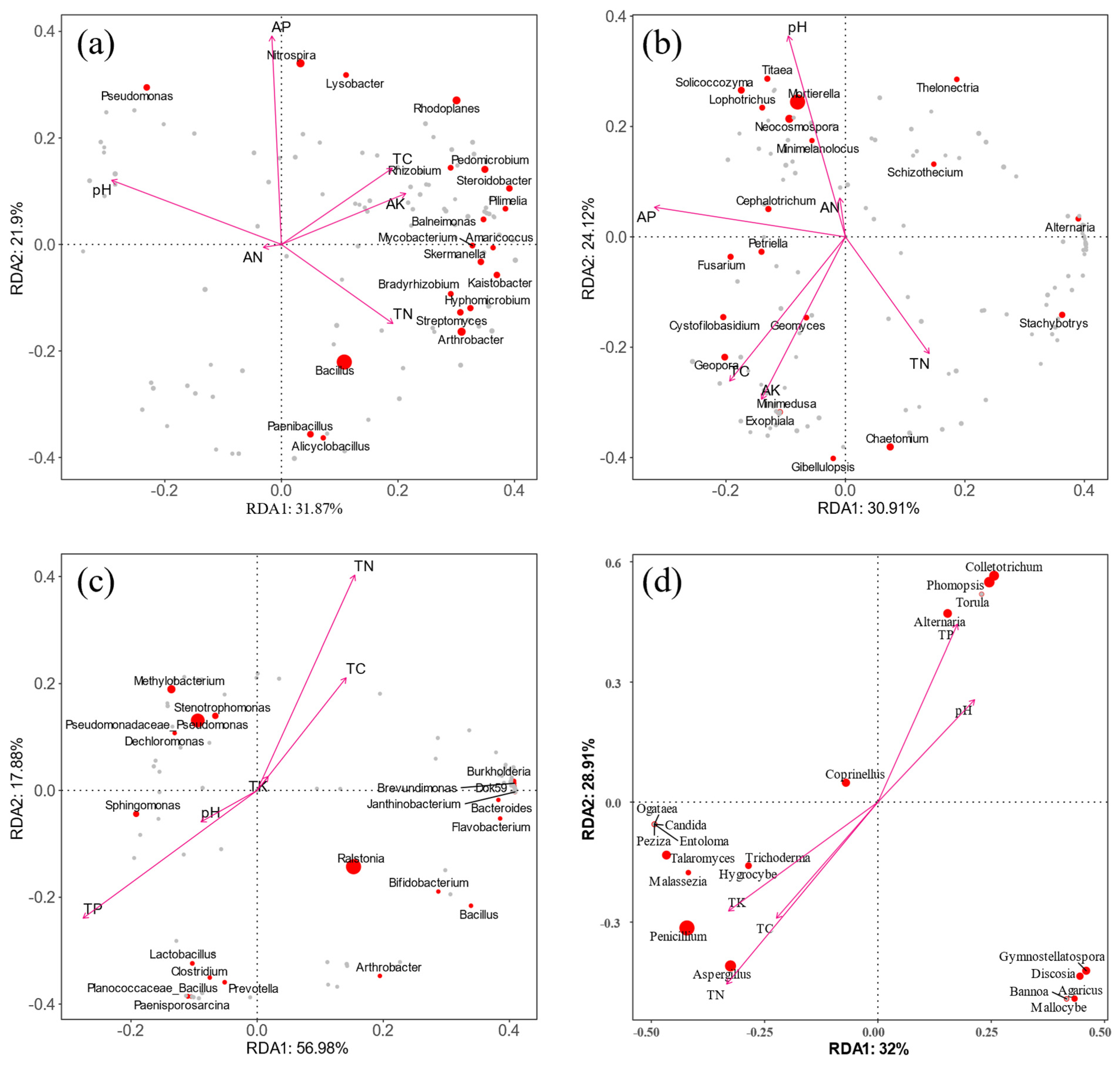
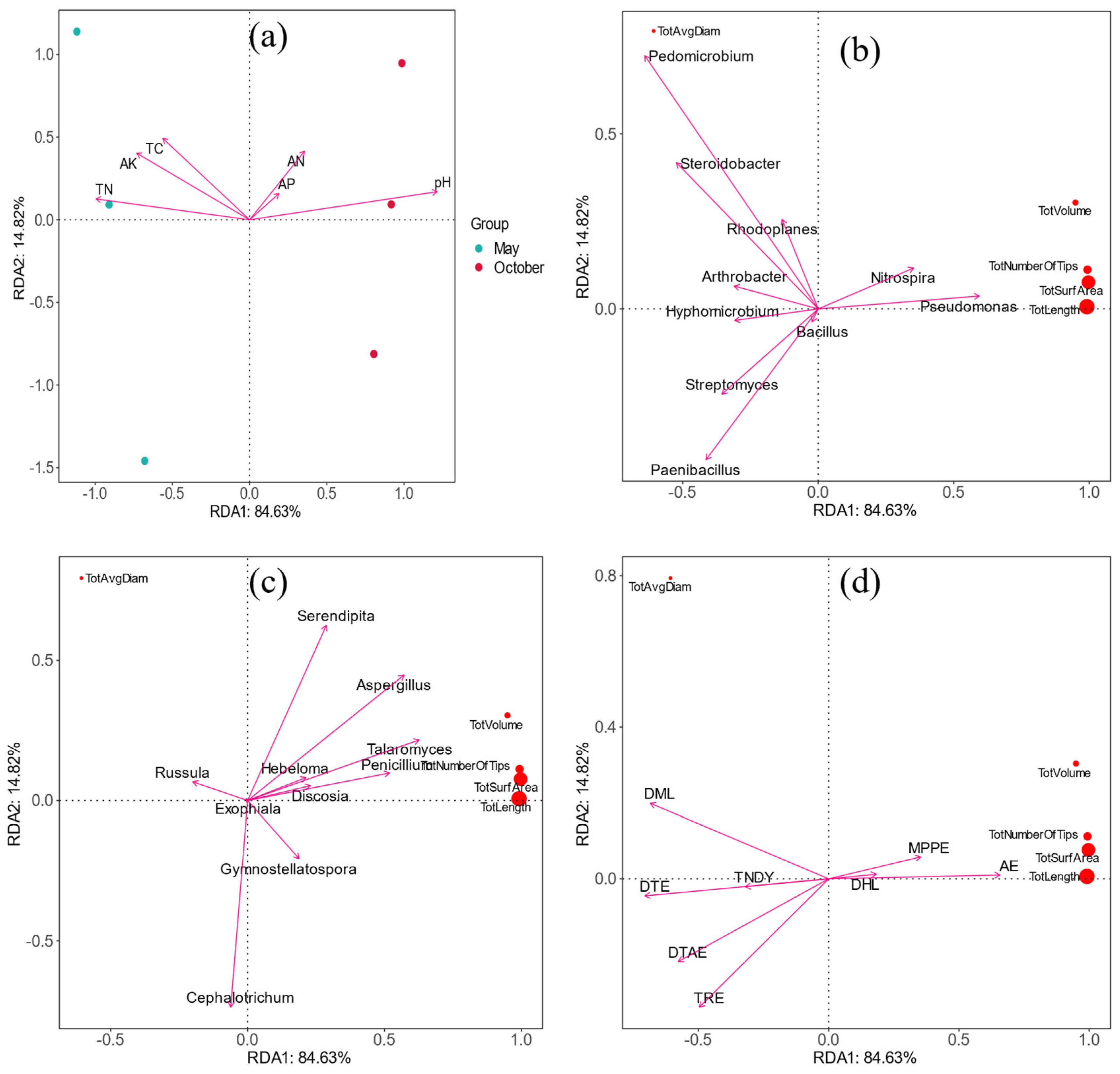
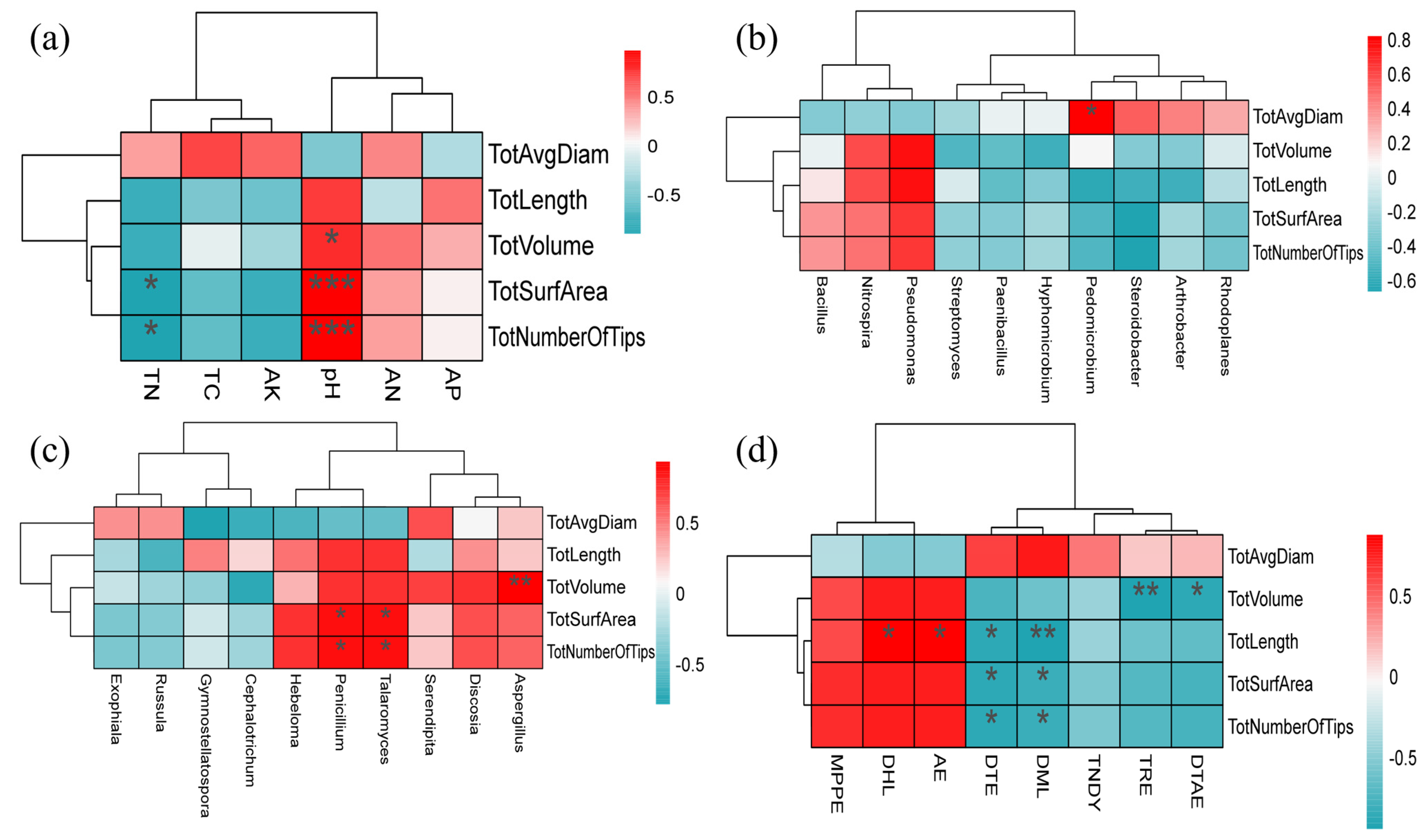
3.6. Correlation Analysis between Microbial Community and Nutrient Factors
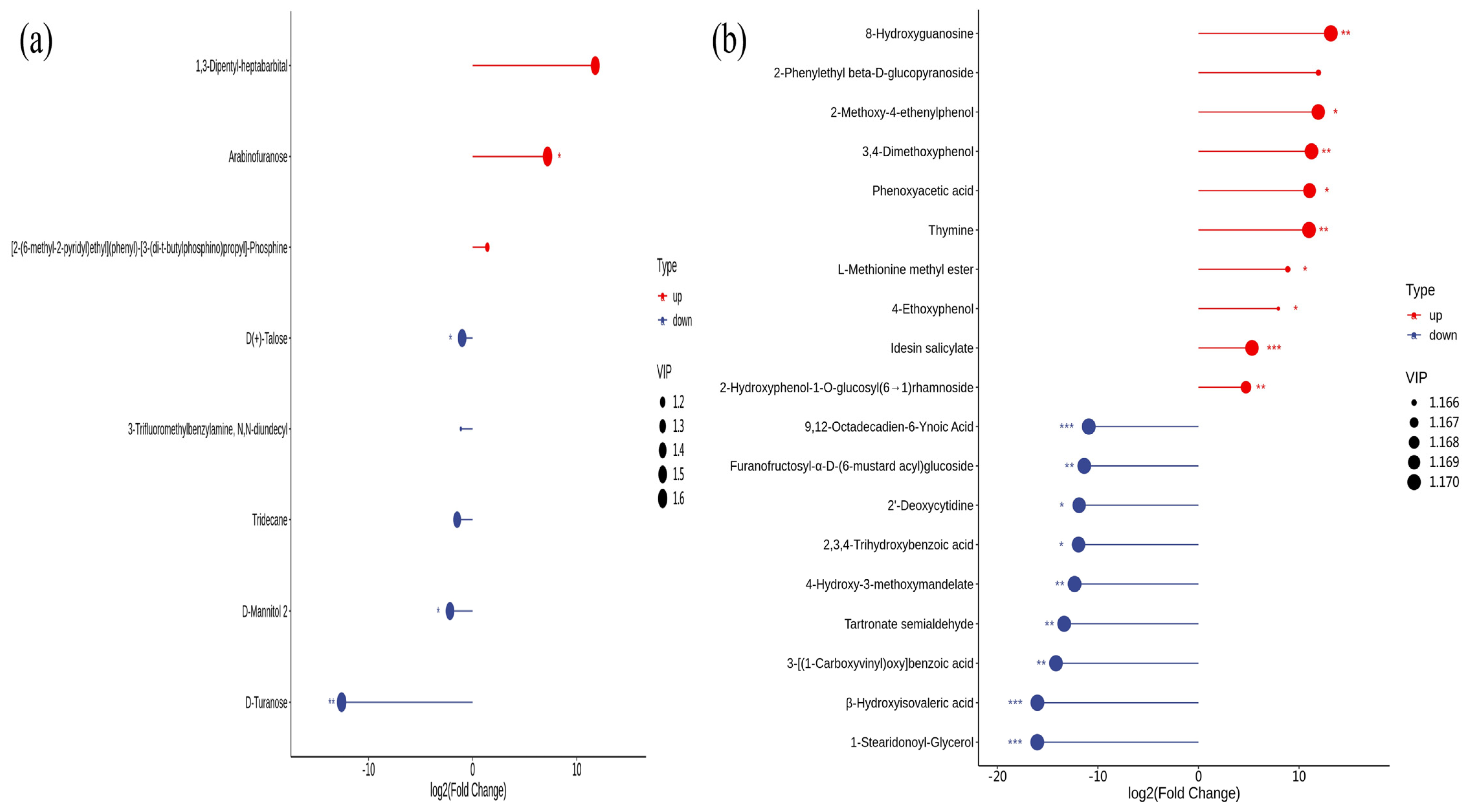
3.7. Metabolomic Analysis of Rhizosphere Soil and Leaves of Young Plants at Different Stages
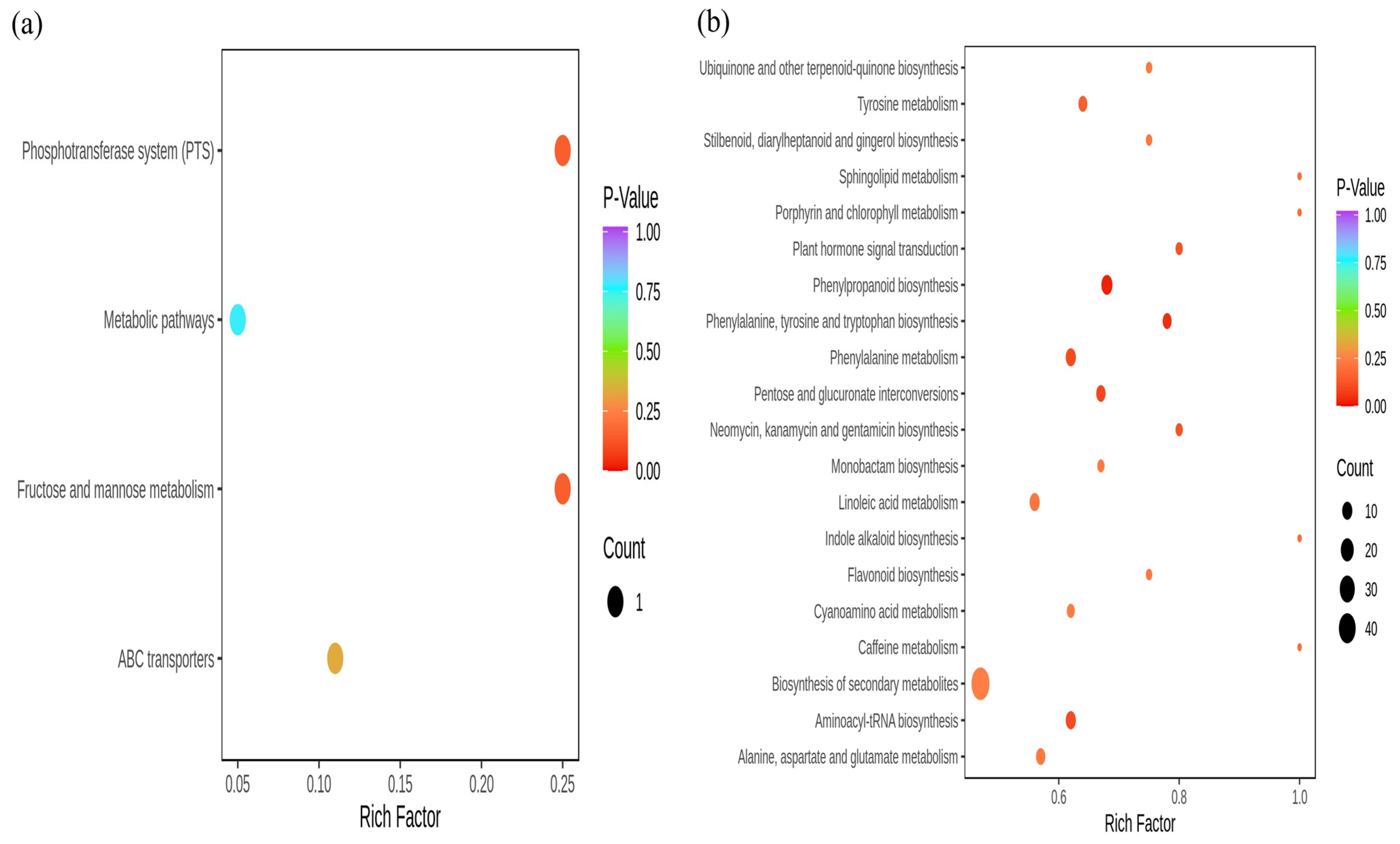
3.8. KEGG Analysis
3.9. Correlation Analysis between Root Morphology and Soil Environment
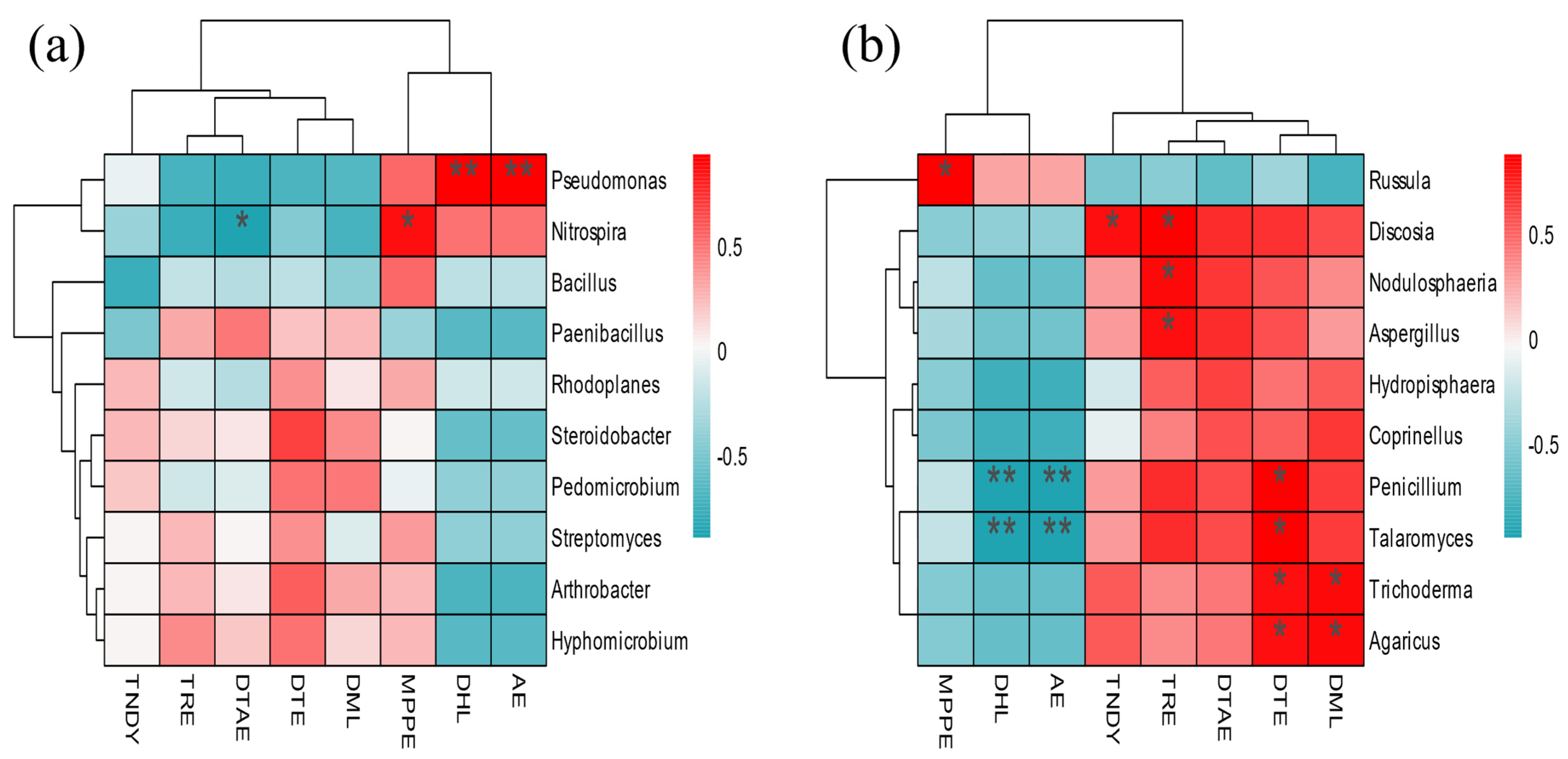
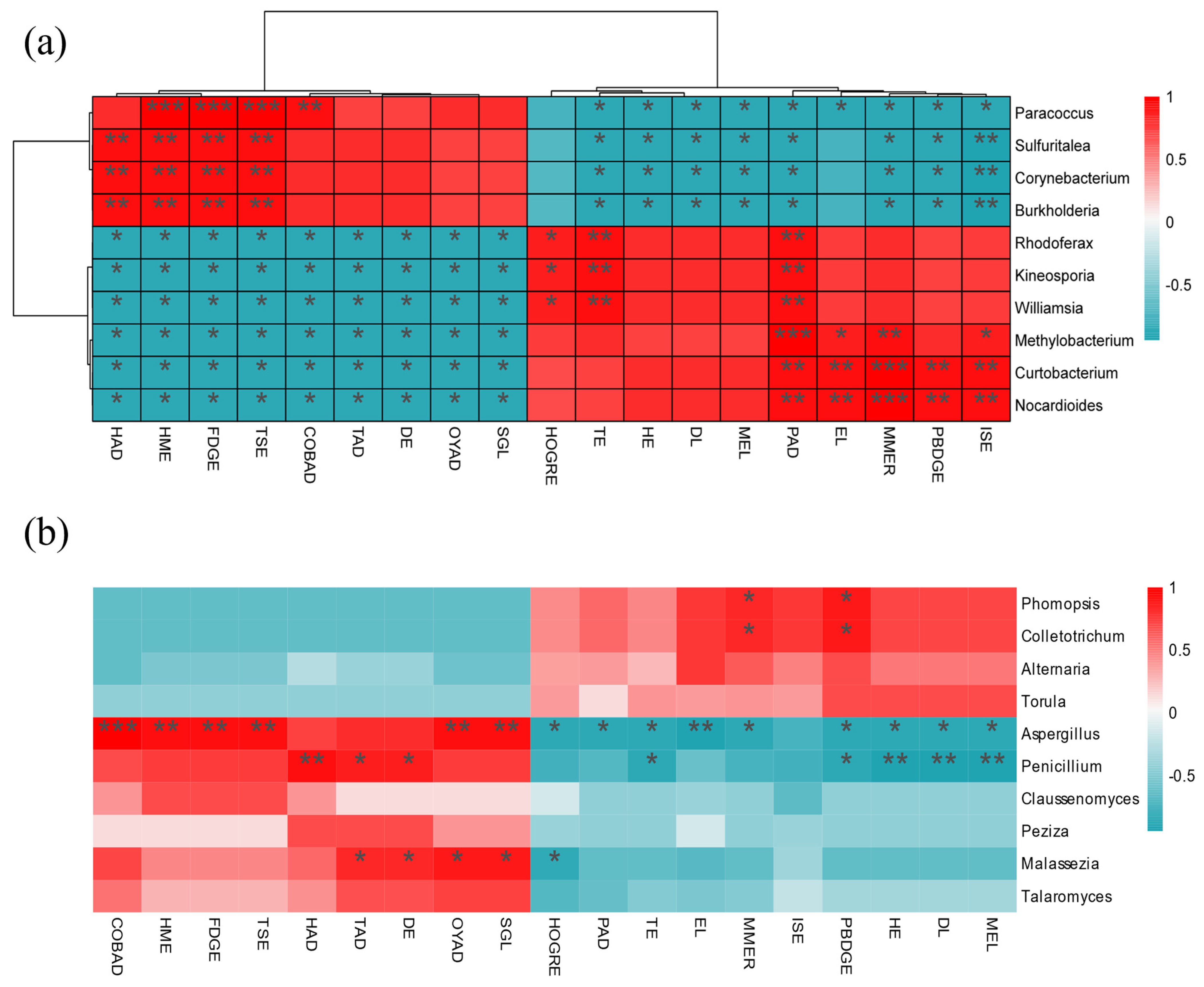
3.10. Metabolome and Microbial Association Analysis
4. Discussion
4.1. Root Characteristics and Leaf–Soil Nutrients
4.2. Leaf-Soil Microbial and Metabolome
4.3. Association of Dominant Microorganisms with Root Morphology, Nutrients, and Differential Metabolites
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, J.S.; Wu, J.; Fang, L.S.; Wang, Y.M.; Geng, X.D.; Liu, Z. Effects of temperature and photoperiod on the branching after sprouted from buds in Idesia polycarpa. J. Northwest For. Univ. 2019, 131, 111–117. [Google Scholar] [CrossRef]
- Li, Y.; Peng, T.; Huang, L.; Zhang, S.Y.; He, Y.Y.; Tang, L. The evaluation of lipids raw material resources with the fatty acid profile and morphological characteristics of Idesia polycarpa Maxim. var. vestita Diels fruit in harvesting. Ind. Crops Prod. 2019, 129, 114–122. [Google Scholar] [CrossRef]
- Bai, B.; Lu, C. Plant population regeneration limitation—from seed production to sapling establishment. Acta Ecol. Sin. 2011, 31, 6624–6632. (In Chinese) [Google Scholar]
- Liu, C.; Lu, Q.; Wang, S.C.; Chen, M.Y.; Xing, S.H. Effects of microhabitat on the growth of Phellodendron amurense saplings. Chin. J. Ecol. 2023, 42, 1–14. Available online: http://kns.cnki.net/kcms/detail/21.1148.q.20230509.1650.020.html (accessed on 2 April 2024).
- Curran, P.M.; Murray, P.M. Soil disturbance, amelioration and rehabilitation affect forest growth, health, soil carbon and chemistry on five long-term soil productivity (LTSP) sites in southeastern British Columbia. For. Ecol. Manag. 2023, 546, 121362. [Google Scholar] [CrossRef]
- Cornelia, H.; Sladjana, S.; Klara, M.K.; Christoph, S.; Jaane, K.; Eliasbeth, A.C.L.; Michael, B.; Friederike, L. Seasonally driven internal P and N nutrient (re)cycling strategies of beech saplings are element specific. Environ. Exp. Bot. 2022, 199, 104894. [Google Scholar] [CrossRef]
- Zheng, C.X. Nitrogen Absorption, Allocation and Utilization of Young Dwarf Rootstock ‘FUJI’ Apple Trees; Northwest A&F University: Yangling, China, 2014. (In Chinese) [Google Scholar]
- Krunoslav, S.; Davor, P.; Tomislav, K.; Lazarevic, B.; Vukmirovic, A.; Sango, M.; Skvorc, Z. Growth of common beech (Fagus sylvatica L.) saplings under the impact of suboptimal mineral nutrition. Sumar. List 2022, 146, 175–185. [Google Scholar] [CrossRef]
- Ali, L.; Wu, T.; Chen, K.; Ullah, S.; Xu, Y.; Yang, M. Analysis of physiological response and differential protein expression of Paramichelia baillonii saplings under phosphorus deficiency. Physiol. Plant 2024, 176, e14225. [Google Scholar] [CrossRef]
- Oktavia, D.; Park, J.W.; Jin, G.Z. Biochar-biofertilizer combinations enhance growth and nutrient uptake in silver maple grown in an urban soil. PLoS ONE 2023, 18, e0288291. [Google Scholar] [CrossRef]
- Marenco, A.R.; Magalhaes, S.D.N.; Gouvea, S.D.R.P.; Antezana-Vera, S.A. Juvenile tree growth correlates with photosynthesis and leaf phosphorus content in central Amazonia. Rev. Ceres 2015, 62, 175–183. [Google Scholar] [CrossRef][Green Version]
- Jimenez, N.M.; Pinto, R.J.; Ripoll, A.M.; Sanchez-Miranda, A.; Navarro, F.B. Restoring silvopastures with oak saplings: Effects of mulch and diameter class on survival, growth, and annual leaf-nutrient patterns. Agrofor. Syst. 2014, 88, 935–946. [Google Scholar] [CrossRef]
- Gou, Q.; Lv, Y.; Zhang, T.; Li, J.Y.; Zhao, H.J.; Liu, J.L. Dynamics and influencing factors of endophytic bacterial community in leaves of Lycium barbarum during different growth periods. Chin. J. Ecol. 2020, 39, 2593–2601. [Google Scholar] [CrossRef]
- Wang, Z.H.; Yu, W.T.; Wang, P.J.; Fan, X.J.; Liu, C.G.; Cai, C.P.; Ye, N.W. Transcriptome analysis at different flowering developmental stages of tea plant. J. Fujian Agric. For. Univ. (Nat. Sci. Ed.) 2022, 51, 46–52. [Google Scholar] [CrossRef]
- Merino-Martín, L.; Griffiths, R.I.; Gweon, H.S.; Furget-Bretagnon, C.; Oliver, A.; Mao, Z.; Le Bissonnais, Y.; Stokes, A. Rhizosphere bacteria are more strongly related to plant root traits than fungi in temperate montane forests: Insights from closed and open forest patches along an elevational gradient. Plant Soil 2020, 450, 183–200. [Google Scholar] [CrossRef]
- Xiao, Y.F. Multi-Omics Analysis of the Integrated Transcriptome Metabolome-Rhizosphere Microbiome Regulatory Model during the Development to Senescence of Rhododendron; Guizhou Normal University: Guizhou, China, 2023. [Google Scholar]
- Lian, T.X.; Huang, Y.Y.; Xie, Y.A.; Huo, X.; Shahid, M.Q.; Tian, L.; Jin, J. Rice SST Variation Shapes the Rhizosphere Bacterial Community, Conferring Tolerance to Salt Stress through Regulating Soil Metabolites. mSystems 2020, 5, 10-1128. [Google Scholar] [CrossRef]
- Yasin, D.; Mahdi, M.S.M.; Ali, D.I.; Hossein, R. Effect of macronutrients and micronutrients on biochemical properties in Paulownia shantung. Plant Cell Tissue Organ Cult. (PCTOC) 2022, 151, 483–495. [Google Scholar] [CrossRef]
- Feng, J.; Sohel, R.; Liu, Z.; Wang, Y.M.; Cai, Q.F.; Geng, X.D.; Zhou, H.N.; Zhang, T.; Wang, S.S.; Xue, X.Y.; et al. Diversity analysis of leaf nutrient endophytes and metabolites in dioecious Idesia polycarpa Maxim. leaves during reproductive stages. Life 2022, 12, 2041. [Google Scholar] [CrossRef]
- Wang, H.M.; Li, Z.; Cai, Q.F.; Wang, Y.M.; Geng, X.D.; Fang, L.S.; Yao, S.Y.; Li, H.Y.; Liu, Z. Mutant Bisexual and Wild Male Flowers Were Compared by Integrated Proteome and Transcriptome Analyses to Provide Insight into the Sex Conversion of Idesia polycarpa Maxim. Forests 2023, 14, 1737. [Google Scholar] [CrossRef]
- Li, Z.; Yang, Y.; Feng, J.; Rana, S.; Wang, S.; Wang, H.; Zhang, T.; Wang, Y.; Guo, G.; Cai, Q.; et al. Evaluation of Fine Root Morphology and Rhizosphere Environmental Characteristics of the Dioecious Idesia polycarpa Maxim. Forests 2024, 15, 234. [Google Scholar] [CrossRef]
- Rana, S.; Liu, Z. Study on the pattern of vegetative growth in young dioecious trees of Idesia polycarpa maxim. Trees 2021, 35, 69–80. [Google Scholar] [CrossRef]
- Wang, H.; Rana, S.; Li, Z.; Geng, X.; Wang, Y.; Cai, Q.; Li, S.; Sun, J.; Liu, Z. Morphological and anatomical changes during floral bud development of the trioecious Idesia polycarpa Maxim. Braz. J. Bot 2022, 45, 679–688. [Google Scholar] [CrossRef]
- Rana, S.; Jemim, R.S.; Li, Z.; Geng, X.; Wang, Y.; Cai, Q.; Liu, Z. Study of the pattern of reproductive allocation and fruit development in young dioecious trees of Idesia polycarpa Maxim. S. Afr. J. Bot. 2022, 146, 472–480. [Google Scholar] [CrossRef]
- YC/T222-2007; Determination of pH in Tobacco and Tobacco Products. National Tobacco Standardization Technical Committee: Beijing, China, 2007.
- Ru, K. Methods of Soil Agrochemical Analysis; China Agricultural Science and Technology Press: Beijing, China, 1999; pp. 166–187. [Google Scholar]
- Shang, S.S.; Yu, X.F.; Qi, Y. Comparison of methods for the determination of hydrolyzable nitrogen in soil and precautions. Agric. Technol. 2022, 42, 97–99. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. Dada2: High-resolution sample inference from illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Teng, Z.N.; Lyu, J.H.; Chen, Y.K.; Zhang, J.H.; Ye, N.H. Effects of stress-induced ABA on root architecture development: Positive and negative actions. Crop J. 2023, 11, 1072–1079. [Google Scholar] [CrossRef]
- Bai, H.X.; Zhang, R.H.; Zhang, X.H.; Wang, L.; Lv, S.; Li, J.W. Populus euphratica seedling root growth characteristics under different water treatmen. For. Eesources Manag. 2015, 11, 61–66. [Google Scholar] [CrossRef]
- Kong, M.; Kang, J.; Han, C.L.; Gu, Y.J.; Siddique, H.M.K.; Li, F.M. Nitrogen, phosphorus, and potassium resorption responses of alfalfa to increasing soil water and P availability in a semi-arid environment. Agronomy 2020, 2, 310. [Google Scholar] [CrossRef]
- Rania, A.B.A.; Hayfa, J.K.; Fakher, A.; Mejda, D.R. A three-year study of comparative effects of four organic amendments on soil health dynamics, tomato production, and rhizosphere microbial community. Commun. Soil Sci. Plant Anal. 2023, 17, 2439–2458. [Google Scholar] [CrossRef]
- Cai, Z.P.; Wang, X.M.; Chen, B.L.; Jiang, P.A.; Shen, J.D.; Liu, M. Annual biomass and nutrient accumulation of Korla fragrant pear. J. Plant Nutr. Fertil. 2013, 19, 656–663. [Google Scholar] [CrossRef]
- Saleha, F.; Sook, I.P.; Myung, J.C. Impact of controlled nitrogen application in water solution on seedling growth, tissue and soil nutrient concentrations in vegetative propagation of strawberry. Hortic. Environ. Biotechnol. 2023, 64, 41–50. [Google Scholar] [CrossRef]
- Albert, G.; Yang, W.R.; Conlan, X.A.; Kong, L.X.; CaHill, D.M.; Wang, Y.C. Boosting plant photosynthesis with carbon dots: A critical review of performance and prospects. Small 2023, 19, e2300671. [Google Scholar] [CrossRef]
- Wang, H.; Niu, D.K.; Hu, D.N.; Guo, X.M. Effects of different fertilizer treatments on phosphorus status of soils and leaves in Camellia oleifera forest. Non-Wood For. Res. 2014, 32, 52–57. [Google Scholar] [CrossRef]
- Li, H.L. Effects of Exogenous MeJA on Photosynthetic Carbon Metabolism and Physiological Characteristics of Strawberry Seedlings under High Temperature Stress; Shandong Agricultural University: Jinan, China, 2023. [Google Scholar]
- Pang, F.H.; Li, X.Q.; Duan, L.Y.; Chen, Y.; Ji, M.F.; Zhang, H.; Han, H.; Chen, Z.J. High-throughput sequencing combined with metabolomics to analyze the effect of heavy metal contamination on farmland soil microbial community and function. Environ. Sci. 2022, 43, 4333–4341. [Google Scholar] [CrossRef]
- Wang, Y.N.; Zeng, X.B.; Ma, Q.; Zhang, Y.; Yu, W.T.; Zheng, Z.; Zhang, N.; Xu, L.Y. Differential responses of canonical nitrifiers and comammox Nitrospira to long-term fertilization in an Alfisol of Northeast China. Front. Microbiol. 2023, 14, 1095937. [Google Scholar] [CrossRef] [PubMed]
- Fu, K.M.; Zhang, X.M.; Fan, Y.; Bian, Y.H.; Qiu, F.G.; Cao, X.Q. The enrichment characterization of Nitrospira under high DO conditions. Environ. Technol. 2023, 21–22. [Google Scholar] [CrossRef]
- Hafiza, F.; Faizah, U.; Muhammed, T.; Nida, S.; Shahid, S.; Asma, S.H. Biological control potential of endophytic fungi with amelioration of systemic resistance in sunflower and GC–MS metabolic profiling of Talaromyces assiutensis. Curr. Microbiol. 2023, 80, 61. [Google Scholar] [CrossRef]
- Xiong, W.; Li, R.; Ren, Y.; Liu, C.; Zhao, Q.Y.; Wu, H.S.; Jousset, A.; Shem, Q.R. Distinct roles for soil fungal and bacterial communities associated with the suppression of vanilla Fusarium wilt disease. Soil Biol. Biochem. 2017, 107, 198–207. [Google Scholar] [CrossRef]
- Su, J.; Li, S.M. Solation and identification of soil-borne pathogenic fungi and their biocontrol bacteria. Grassl. Turf. 2019, 39, 79–84+91. [Google Scholar] [CrossRef]
- Liu, L.L.; Li, H.L.; Meng, Z.S.; Peng, J.Y.; Li, X.D.; Peng, C.; Lu, L.; Xu, X. Differences in phyllosphere microbial communities structure between female and male Populus cathayana. Acta Microbiol. Sin. 2017, 60, 556–569. [Google Scholar] [CrossRef]
- Ren, H.Y.; Xu, Q.; Qi, X.J.; Yu, Z.P.; Zheng, X.L.; Zhang, S.W.; Wang, Z.S. Differences of bacterial and fungal communities in the tree and rhizosphere of the healthy and twig blight-diseased bayberry. Chin. J. Appl. Ecol. 2021, 32, 3107–3118. [Google Scholar] [CrossRef]
- Weisener, C.; Lee, J.; Chaganti, R.S.; Reid, T.; Falk, N.; Drouillard, K. Investigating sources and sinks of N2O expression from freshwater microbial communities in urban watershed sediments. Chemosphere 2017, 188, 697–705. [Google Scholar] [CrossRef]
- Ma, B.G.; Wei, X.H.; Meng, X.J.; Sun, Z.X. Advancements in study on Burkholderia for plant disease biocontrol. Agric. Res. Appl. 2023, 36, 1–8. [Google Scholar] [CrossRef]
- Amna, S.F.; Anne, J.D.; Shazia, K.; Oscar, P.K. Draft genome sequences of Bacillus velezensis strains AF_3B and OS2, Bacillus amyloliquefaciens strain BS9, Bacillus halotolerans strain A1, and Bacillus sp. strain BS3, producing biosurfactants with antimicrobial potential. Microbiol. Resour. Announc. 2022, 11, e0048222. [Google Scholar] [CrossRef]
- Zhang, T. A Master Thesis Submitted to University of Electronic Science and Technology of China; University of Electronic Science and Technology of China: Chengdu, China, 2022. [Google Scholar] [CrossRef]
- Veeresh; Amaresh, Y.S.; Aswathanarayana, D.S.; Gowdar, S.B.; Nidagundi, J.M. Efficacy of fungicides and commercially available organic products against blight of cotton caused by Alternaria macrospora. Int. J. Environ. Clim. Chang. 2023, 13, 3612–3617. [Google Scholar] [CrossRef]
- He, Y.X.; Ma, Y.Y.; He, Z.Y.; He, S.Q.; Zhang, S.P.; Li, Y.; Yuan, W.J. First Report of Stem Rot Caused by Botryosphaeria dothidea and Phomopsis velata on Forsythia suspensa in China. Plant Dis. 2022, 10, 2247. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.H.; Zhu, X.Z.; Wang, S.; Liu, Z.; Feng, J.; Li, Z. Research progress of main diseases insect pests and control technology of Idesia polycarpa. J. Henan For. Sci. Technol. 2021, 41, 21–25+38. [Google Scholar] [CrossRef]
- Yang, M.; Li, S.; Cao, H.X.; Huang, T.; Li, Y.; Xie, J.W.; Zhao, G.M. Microbial mechanism of Helianthus Tuberosus root exudates for improving saline-alkali soil. Bull. Soil Water Conserv. 2021, 41, 71–81. [Google Scholar] [CrossRef]
- Zhou, X.; Guo, Z.; Chen, C.; Jia, Z.J. Soil microbial community structure and diversity are largely influenced by soil pH and nutrient quality in 78-year-old tree plantations. Biogeosciences 2017, 14, 2101–2111. [Google Scholar] [CrossRef]
- Liao, L.B.; Shi, F.S.; Zhang, N.N.; Chen, X.X.; Bu, X.X.; Sun, F.Y. Effects of different planting years on rhizosphere soil physiochemical properties and microbial community of Zanthoxylum bungeanum. Bull. Bot. Res. 2022, 42, 466–474. [Google Scholar] [CrossRef]
- La, J.L.; Lin, H.S.; Lin, Q.; Wu, X.H.; Wei, M.K. Isolation of a bacteria with resistance to copper and its biological characters. J. Anhui Agric. Sci. 2011, 39, 22521–22524. [Google Scholar] [CrossRef]
- Wang, Z. Microbial Community Diversity of Carbon Sequestration and Its Influencing Factors in the Tibetan Alpine Meadows; China University of Geosciences: Beijing, China, 2019. [Google Scholar] [CrossRef]
- Yu, H.X.; Liang, H.L.; Wang, Z.X.; Yang, X.Y.; Li, W.H. Isolation, identification and growth-promoting effects of culturable nitrogen-fixing bacteria and ammonifying bacteria in rhizosphere soil of Mikania micrantha. Acta Microbiol. Sin. 2022, 62, 1851–1863. [Google Scholar] [CrossRef]
- Gui, X.W.; Zhang, Y.; Song, Q.N.; Chen, H.H.; Yang, Q.P.; Luan, F.G. Isolation and identification of the mineral potassium-solubilizing bacteria in Phyllostachys edulis forest. J. For. Environ. 2018, 38, 499–505. [Google Scholar] [CrossRef]
- Zhao, Y.K.; Zhang, W.S.; Wang, Y.N.; Li, K.X.; Jia, H.Z.; Li, X. Research progress in physiology and molecular biology of plant response to high pH. Chin. J. Eco-Agric. 2008, 16, 783–787. [Google Scholar]
- Luo, W.; Shi, L.H. Isolation and identification of a potassium-releasing strain of Pseudomonas sp. J. Anhui Agric. Sci. 2016, 44, 17–19. [Google Scholar] [CrossRef]
- Luo, L.Y.; Zhang, Z.; Wang, P.; Han, Y.Q.; Jin, D.C.; Tan, X.Q.; Zhang, D.Y.; Hamid, M.-R.; Lu, X.Y.; Liu, Y. Variations in phyllosphere microbial community along with the development of angular leaf-spot of cucumber. AMB Express 2019, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Shi, H.X.; Liu, L.L.; Zhang, X.; Yang, P.; Chen, H.; Feng, Y.; Chen, X.O. Structure and diversity of bacterial community in different niches of garden plant Ligustrum lucidum Ait. Acta Microbiol. Sin. 2022, 62, 686–702. [Google Scholar] [CrossRef]
- Yan, H.P. Dynamic Changes of Endogenous Hormones during Flowering and Fruiting of Idesia polycarpa Maxim; Henan Agricultural University: Zhengzhou, China, 2019. [Google Scholar] [CrossRef]
- Qin, H. Leaf Functional Traits and Rhizosphere Bacterial Community of Urophysa Rockii in Thereintroduced Process; Sichuan Agricultural University: Yaan, China, 2022. [Google Scholar]
- Zhan, H.X.; Liu, X.L.; Zhang, Y.R.; Jiang, X. Analysis of soil bacterial community composition of Codonopsis pilosula in different production Area. Mol. Plant Breed. 2023, 21, 4510–4517. [Google Scholar]
- Xiao, Y.H.; Chen, R.; Chen, L.; Yang, B.; Jiang, L.H.; Fang, J. Endophytic fungus Talaromyces sp. MR1 promotes the growth and cadmium uptake of Arabidopsis thaliana L. under cadmium stress. Curr. Microbiol. 2023, 80, 346. [Google Scholar] [CrossRef] [PubMed]
- Sofia, S.; Helgeneusa, C.; Teresa, L.; Vitor, R.; Nuno, R.; Albert, P.J.; Teresa, L.N.; Paula, B. Potential of the endophyte Penicillium commune in the control of olive anthracnose via induction of antifungal volatiles in host plant. Biol. Control 2023, 187, 105373. [Google Scholar]
- Qi, X.H. Study on Continuously Cropping Autotoxicity of Rehmannia Glutinosa Libosch; Fujian Agriculture and Forestry University: Fuzhou, China, 2008. [Google Scholar]
- Wang, P.; Meng, R.Y.; Mao, R.Z.; Yang, K.; Su, Z.W.; Wang, Z.Q.; Yang, S.Q.; Huang, H.P.; He, X.H. Soil physical and chemical properties, microorganisms and metabolites in different culture environments of Gastrodia elata. Chin. J. Exp. Tradit. Med. Formulae 2021, 27, 164–174. [Google Scholar] [CrossRef]
- Bi, B.Y.; Wang, K.; Zhang, H.; Wang, Y.; Fei, H.Y.; Pan, R.P.; Han, F.P. Plants use rhizosphere metabolites to regulate soil microbial diversity. Land Degrad. Dev. 2021, 32, 5267–5280. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Wang, S.; Rana, S.; Wang, Y.; Liu, Z.; Cai, Q.; Geng, X.; Yuan, Q.; Yang, Y.; Miao, C.; et al. Analysis of Leaf and Soil Nutrients, Microorganisms and Metabolome in the Growth Period of Idesia polycarpa Maxim. Microorganisms 2024, 12, 746. https://doi.org/10.3390/microorganisms12040746
Zhang T, Wang S, Rana S, Wang Y, Liu Z, Cai Q, Geng X, Yuan Q, Yang Y, Miao C, et al. Analysis of Leaf and Soil Nutrients, Microorganisms and Metabolome in the Growth Period of Idesia polycarpa Maxim. Microorganisms. 2024; 12(4):746. https://doi.org/10.3390/microorganisms12040746
Chicago/Turabian StyleZhang, Tao, Shasha Wang, Sohel Rana, Yanmei Wang, Zhen Liu, Qifei Cai, Xiaodong Geng, Qiupeng Yuan, Yi Yang, Chao Miao, and et al. 2024. "Analysis of Leaf and Soil Nutrients, Microorganisms and Metabolome in the Growth Period of Idesia polycarpa Maxim" Microorganisms 12, no. 4: 746. https://doi.org/10.3390/microorganisms12040746
APA StyleZhang, T., Wang, S., Rana, S., Wang, Y., Liu, Z., Cai, Q., Geng, X., Yuan, Q., Yang, Y., Miao, C., Xue, X., Dai, L., & Li, Z. (2024). Analysis of Leaf and Soil Nutrients, Microorganisms and Metabolome in the Growth Period of Idesia polycarpa Maxim. Microorganisms, 12(4), 746. https://doi.org/10.3390/microorganisms12040746