
Molecular Role of HIV-1 Human Receptors (CCL5–CCR5 Axis) in neuroAIDS: A Systematic Review

 ,
,  ,
, 
 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Design
2.2. Search Strategy
2.3. Data Extraction
3. Results
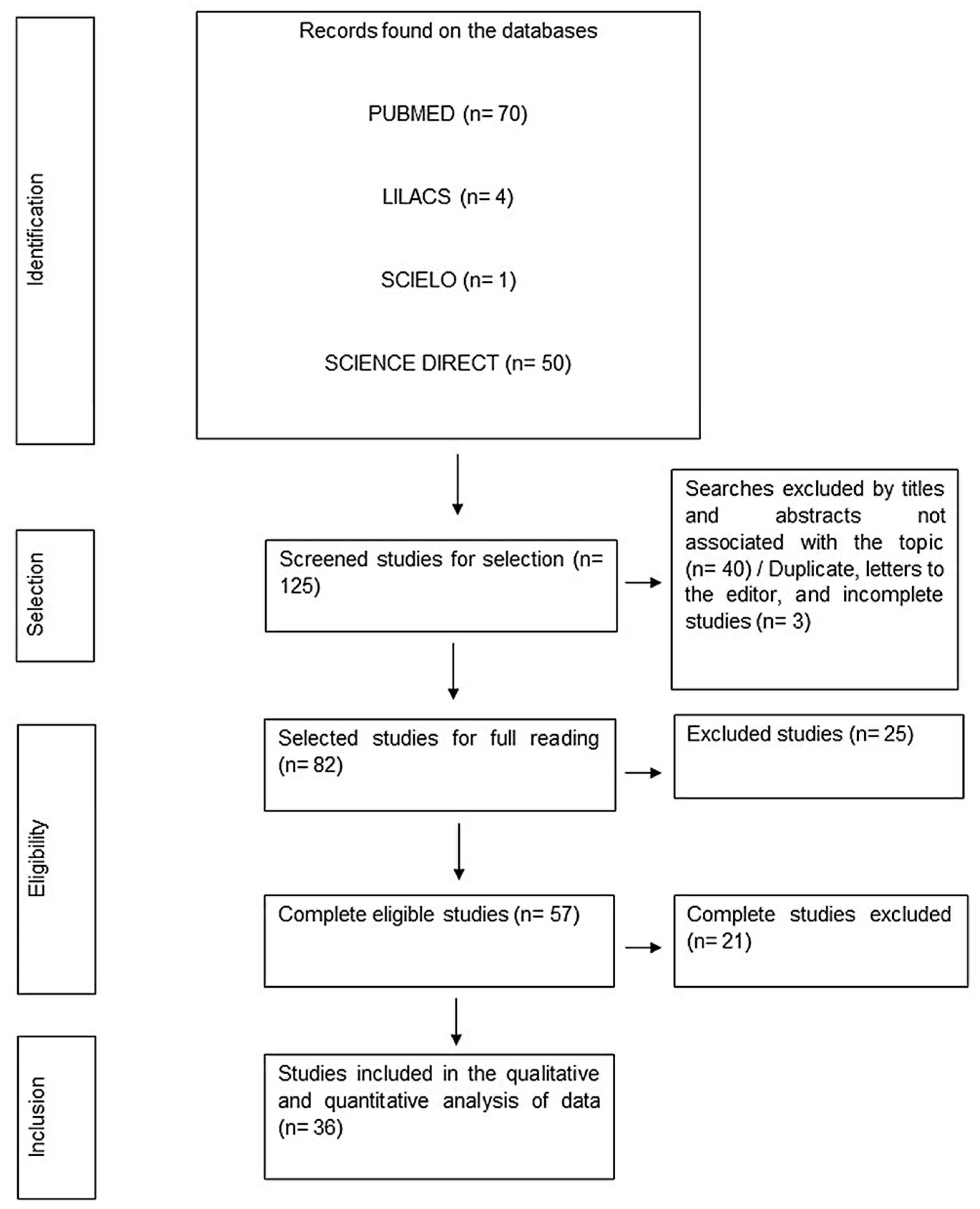
3.1. Literature Search
3.2. Characteristics of the Studies
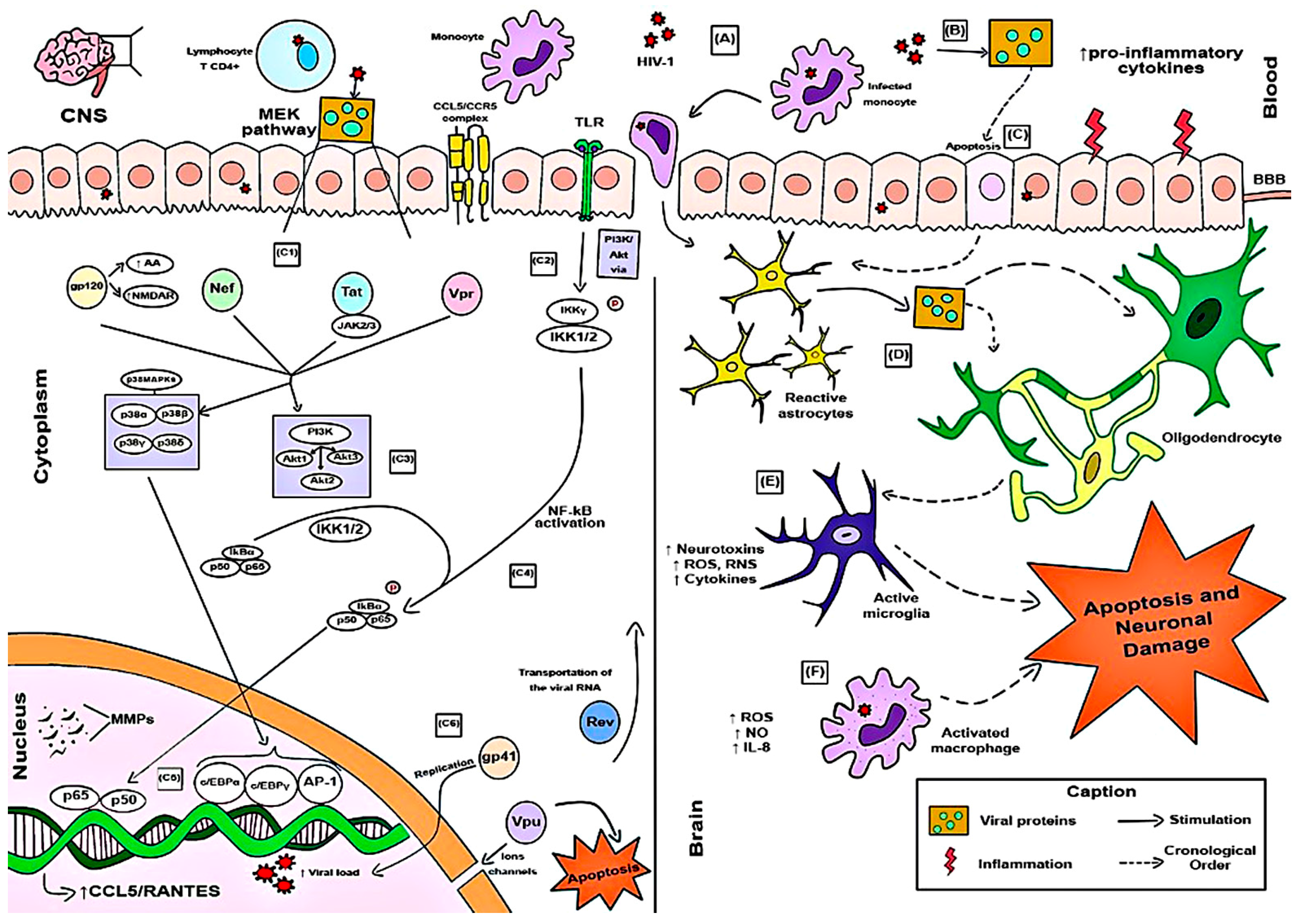
3.3. Effects of HIV-1 Infection on Cellular and Molecular Pathways from the Perspective of the CCL5–CCR5 Immune Axis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Al-Jabri, A.A. Mechanisms of host resistance against HIV infection and progression to AIDS. Sultan Qaboos Univ. Med. J. 2007, 7, 82. [Google Scholar]
- Banks, W.; Ercal, N.; Otamis Price, T. The Blood-Brain Barrier in NeuroAIDS. Curr. HIV Res. 2006, 4, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Balcom, E.F.; Roda, W.C.; Cohen, E.A.; Li, M.Y.; Power, C. HIV-1 persistence in the central nervous system: Viral and host determinants during antiretroviral therapy. Curr. Opin. Virol. 2019, 38, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Sacktor, N.; McDermott, M.P.; Marder, K.; Schifitto, G.; Selnes, O.A.; McArthur, J.C.; Stern, Y.; Albert, S.; Palumbo, D.; Kieburtz, K. HIV-associated cognitive impairment before and after the advent of combination therapy. J. Neurovirol. 2002, 8, 136–142. [Google Scholar] [CrossRef]
- Rojas-Celis, V.; Valiente-Echeverría, F.; Soto-Rifo, R.; Toro-Ascuy, D. New Challenges of HIV-1 Infection: How HIV-1 Attacks and Resides in the Central Nervous System. Cells 2019, 8, 1245. [Google Scholar] [CrossRef]
- Soares, D.C.; Filho, L.C.F.; Souza dos Reis, H.; Rodrigues, Y.C.; Freitas, F.B.; de Oliveira Souza, C.; Damacena, G.N.; Véras, N.M.C.; Gaspar, P.C.; Benzaken, A.S.; et al. Assessment of the Accuracy, Usability and Acceptability of a Rapid Test for the Simultaneous Diagnosis of Syphilis and HIV Infection in a Real-Life Scenario in the Amazon Region, Brazil. Diagnostics 2023, 13, 810. [Google Scholar] [CrossRef]
- Balasubramaniam, M.; Pandhare, J.; Dash, C. Immune control of HIV. J. Life Sci. Westlake Village Calif. 2019, 1, 4. [Google Scholar] [CrossRef]
- Dore, G.J.; Correll, P.K.; Li, Y.; Kaldor, J.M.; Cooper, D.A.; Brew, B.J. Changes to AIDS dementia complex in the era of highly active antiretroviral therapy. AIDS 1999, 13, 1249–1253. [Google Scholar] [CrossRef]
- Brignol, S.; Kerr, L.; Amorim, L.D.; Dourado, I. Fatores associados a infecção por HIV numa amostra respondent-driven sampling de homens que fazem sexo com homens, Salvador. Rev. Bras. Epidemiol. 2016, 19, 256–271. [Google Scholar] [CrossRef]
- Bruxelle, J.-F.; Trattnig, N.; Mureithi, M.W.; Landais, E.; Pantophlet, R. HIV-1 entry and prospects for protecting against infection. Microorganisms 2021, 9, 228. [Google Scholar] [CrossRef]
- Ripamonti, E.; Clerici, M. The association of memory disorders and chronic HIV disease in the antiretroviral therapy era: A systematic literature review. HIV Med. 2020, 21, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Muñoz, L.; Barroso, R.; Dyrhaug, S.Y.; Navarro, G.; Lucas, P.; Soriano, S.F.; Vega, B.; Costas, C.; Muñoz-Fernández, M.Á.; Santiago, C. CCR5/CD4/CXCR4 oligomerization prevents HIV-1 gp120IIIB binding to the cell surface. Proc. Natl. Acad. Sci. USA 2014, 111, E1960–E1969. [Google Scholar] [CrossRef] [PubMed]
- Prathipati, P.K.; Mandal, S.; Destache, C.J. A review of CCR5 antibodies against HIV: Current and future aspects. Ther. Deliv. 2019, 10, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Brelot, A.; Chakrabarti, L.A. CCR5 Revisited: How Mechanisms of HIV Entry Govern AIDS Pathogenesis. J. Mol. Biol. 2018, 430, 2557–2589. [Google Scholar] [CrossRef] [PubMed]
- Barmania, F.; Pepper, M.S. C-C chemokine receptor type five (CCR5): An emerging target for the control of HIV infection. Appl. Transl. Genom. 2013, 2, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Mutuiri, S.P.M.; Kutima, H.L.; Mwapagha, L.M.; Munyao, J.K.; Nyamache, A.K.; Wanjiru, I.; Khamadi, S.A. RANTES Gene Polymorphisms Associated with HIV-1 Infections in Kenyan Population. Dis. Markers 2016, 2016, 1–6. [Google Scholar] [CrossRef]
- Bhusal, R.P.; Foster, S.R.; Stone, M.J. Structural basis of chemokine and receptor interactions: Key regulators of leukocyte recruitment in inflammatory responses. Protein Sci. 2020, 29, 420–432. [Google Scholar] [CrossRef]
- Kumar, R.; Bhatia, M.; Pai, K. Role of Chemokines in the Pathogenesis of Visceral Leishmaniasis. Curr. Med. Chem. 2022, 29, 5441–5461. [Google Scholar] [CrossRef]
- Zanoni, M.; Palesch, D.; Pinacchio, C.; Statzu, M.; Tharp, G.K.; Paiardini, M.; Chahroudi, A.; Bosinger, S.E.; Yoon, J.; Cox, B.; et al. Innate, non-cytolytic CD8+ T cell-mediated suppression of HIV replication by MHC-independent inhibition of virus transcription. PLoS Pathog. 2020, 16, e1008821. [Google Scholar] [CrossRef]
- Tamamis, P.; Floudas, C.A. Elucidating a Key Anti-HIV-1 and Cancer-Associated Axis: The Structure of CCL5 (Rantes) in Complex with CCR5. Sci. Rep. 2014, 4, 5447. [Google Scholar] [CrossRef]
- Nabi, B.; Rehman, S.; Pottoo, F.H.; Baboota, S.; Ali, J. Dissecting the therapeutic relevance of gene therapy in NeuroAIDS: An evolving epidemic. Curr. Gene Ther. 2020, 20, 174–183. [Google Scholar] [CrossRef]
- Eggers, C.; Arendt, G.; Hahn, K.; Husstedt, I.W.; Maschke, M.; Neuen-Jacob, E.; Obermann, M.; Rosenkranz, T.; Schielke, E.; Straube, E. HIV-1-associated neurocognitive disorder: Epidemiology, pathogenesis, diagnosis, and treatment. J. Neurol. 2017, 264, 1715–1727. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.M.D.C.; Pimenta, C.A.D.M.; Nobre, M.R.C. The PICO strategy for the research question construction and evidence search. Rev. Lat. Am. Enferm. 2007, 15, 508–511. [Google Scholar] [CrossRef] [PubMed]
- Aromataris, E.; Munn, Z. Joanna Briggs Institute reviewer’s manual. Joanna Briggs Inst. 2017, 2017. [Google Scholar] [CrossRef]
- Munn, Z.; Aromataris, E.; Tufanaru, C.; Stern, C.; Porritt, K.; Farrow, J.; Lockwood, C.; Stephenson, M.; Moola, S.; Lizarondo, L.; et al. The development of software to support multiple systematic review types: The Joanna Briggs Institute System for the Unified Management, Assessment and Review of Information (JBI SUMARI). Int. J. Evid. Based Healthc. 2019, 17, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.J.A.; Silva, C.S.; Da Silva Vieira, M.C.; Dos Santos, P.A.S.; Frota, C.C.; Lima, K.V.B.; Lima, L.N.G.C. The Relationship between TLR3 Rs3775291 Polymorphism and Infectious Diseases: A Meta-Analysis of Case-Control Studies. Genes 2023, 14, 1311. [Google Scholar] [CrossRef] [PubMed]
- Gonek, M.; McLane, V.D.; Stevens, D.L.; Lippold, K.; Akbarali, H.I.; Knapp, P.E.; Dewey, W.L.; Hauser, K.F.; Paris, J.J. CCR5 mediates HIV-1 Tat-induced neuroinflammation and influences morphine tolerance, dependence, and reward. Brain Behav. Immun. 2018, 69, 124–138. [Google Scholar] [CrossRef] [PubMed]
- Dever, S.M.; Costin, B.N.; Xu, R.; El-Hage, N.; Balinang, J.; Samoshkin, A.; O’Brien, M.A.; McRae, M.; Diatchenko, L.; Knapp, P.E.; et al. Differential expression of the alternatively-spliced OPRM1 isoform MOR-1K in HIV-infected subjects. AIDS 2014, 28, 19–30. [Google Scholar] [CrossRef]
- Sabri, F.; Tresoldi, E.; Di Stefano, M.; Polo, S.; Monaco, M.C.; Verani, A.; Fiore, J.R.; Lusso, P.; Major, E.; Chiodi, F.; et al. Nonproductive Human Immunodeficiency Virus Type 1 Infection of Human Fetal Astrocytes: Independence from CD4 and Major Chemokine Receptors. Virology 1999, 264, 370–384. [Google Scholar] [CrossRef]
- Mayer, V.; Schmidtmayerová, H. Encephalopathy in AIDS--increased formation of beta-chemokines in monocytes after HIV-1 virus infection: Mechanisms of CNS involvement. Bratislavské Lekárske Listy 1997, 98, 330–334. [Google Scholar]
- Albright, A.V.; Shieh, J.T.C.; Itoh, T.; Lee, B.; Pleasure, D.; O’Connor, M.J.; Doms, R.W.; González-Scarano, F. Microglia Express CCR5, CXCR4, and CCR3, but of These, CCR5 Is the Principal Coreceptor for Human Immunodeficiency Virus Type 1 Dementia Isolates. J. Virol. 1999, 73, 205–213. [Google Scholar] [CrossRef]
- Yuan, Y.; Arnatt, C.K.; El-Hage, N.; Dever, S.M.; Jacob, J.C.; Selley, D.E.; Hauser, K.F.; Zhang, Y. A bivalent ligand targeting the putative mu opioid receptor and chemokine receptor CCR5 heterodimer: Binding affinity versus functional activities. Med. Chem. Commun. 2013, 4, 847. [Google Scholar] [CrossRef]
- Maung, R.; Medders, K.E.; Sejbuk, N.E.; Desai, M.K.; Russo, R.; Kaul, M. Genetic Knockouts Suggest a Critical Role for HIV Co-Receptors in Models of HIV Gp120-Induced Brain Injury. J. Neuroimmune Pharmacol 2012, 7, 306–318. [Google Scholar] [CrossRef]
- Zhou, D.; Masliah, E.; Spector, S.A. Autophagy Is Increased in Postmortem Brains of Persons With HIV-1-Associated Encephalitis. J. Infect. Dis. 2011, 203, 1647–1657. [Google Scholar] [CrossRef] [PubMed]
- Gorantla, S.; Makarov, E.; Roy, D.; Finke-Dwyer, J.; Murrin, L.C.; Gendelman, H.E.; Poluektova, L. Immunoregulation of a CB2 Receptor Agonist in a Murine Model of NeuroAIDS. J. Neuroimmune Pharmacol. 2010, 5, 456–468. [Google Scholar] [CrossRef] [PubMed]
- Polianova, M.T.; Ruscetti, F.W.; Pert, C.B.; Ruff, M.R. Chemokine receptor-5 (CCR5) is a receptor for the HIV entry inhibitor peptide T (DAPTA). Antivir. Res. 2005, 67, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Rao, V.R.; Ruiz, A.P.; Prasad, V.R. Viral and cellular factors underlying neuropathogenesis in HIV associated neurocognitive disorders (HAND). AIDS Res. Ther. 2014, 11, 13. [Google Scholar] [CrossRef]
- Shah, A.; Singh, D.P.; Buch, S.; Kumar, A. HIV-1 envelope protein Gp120 up regulates CCL5 production in astrocytes which can be circumvented by gp120-specific siRNA and inhibitors of NF-kappa B pathway. Biochem. Biophys. Res. Commun. 2011, 414, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Si, Q.; Kim, M.-O.; Zhao, M.-L.; Landau, N.R.; Goldstein, H.; Lee, S.C. Vpr- and Nef-Dependent Induction of RANTES/CCL5 in Microglial Cells. Virology 2002, 301, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shah, A.; Gangwani, M.R.; Silverstein, P.S.; Fu, M.; Kumar, A. HIV-1 Nef Induces CCL5 production in astrocytes through P38-MAPK and PI3K/Akt pathway and utilizes NF-kB, CEBP and AP-1 transcription factors. Sci. Rep. 2014, 4, 4450. [Google Scholar] [CrossRef]
- Gumbs, S.B.H.; Berdenis van Berlekom, A.; Kübler, R.; Schipper, P.J.; Gharu, L.; Boks, M.P.; Ormel, P.R.; Wensing, A.M.J.; de Witte, L.D.; Nijhuis, M. Characterization of HIV-1 Infection in Microglia-Containing Human Cerebral Organoids. Viruses 2022, 14, 829. [Google Scholar] [CrossRef]
- Rottman, J.B.; Ganley, K.P.; Williams, K.; Wu, L.; Mackay, C.R.; Ringler, D.J. Cellular localization of the chemokine receptor CCR5. Correlation to cellular targets of HIV-1 infection. Am. J. Pathol. 1997, 151, 1341–1351. [Google Scholar]
- Chang, T.L.-Y.; Gordon, C.J.; Roscic-Mrkic, B.; Power, C.; Proudfoot, A.E.I.; Moore, J.P.; Trkola, A. Interaction of the CC-Chemokine RANTES with Glycosaminoglycans Activates a p44/p42 Mitogen-Activated Protein Kinase-Dependent Signaling Pathway and Enhances Human Immunodeficiency Virus Type 1 Infectivity. J. Virol. 2002, 76, 2245–2254. [Google Scholar] [CrossRef] [PubMed]
- Vago, L.; Nebuloni, M.; Bonetto, S.; Pellegrinelli, A.; Zerbi, P.; Ferri, A.; Lavri, E.; Capra, M.; Grassi, M.P.; Costanzi, G. Rantes distribution and cellular localization in the brain of HIV-infected patients. Clin. Neuropathol. 2001, 20, 139–145. [Google Scholar]
- Martin-Blondel, G.; Brassat, D.; Bauer, J.; Lassmann, H.; Liblau, R.S. CCR5 blockade for neuroinflammatory diseases—Beyond control of HIV. Nat. Rev. Neurol. 2016, 12, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Gorry, P.R.; Taylor, J.; Holm, G.H.; Mehle, A.; Morgan, T.; Cayabyab, M.; Farzan, M.; Wang, H.; Bell, J.E.; Kunstman, K.; et al. Increased CCR5 Affinity and Reduced CCR5/CD4 Dependence of a Neurovirulent Primary Human Immunodeficiency Virus Type 1 Isolate. J. Virol. 2002, 76, 6277–6292. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Smit, T.K.; Morgello, S.; Tourtellotte, W.; Gelman, B.; Brew, B.J.; Saksena, N.K. Env Gp120 Sequence Analysis of HIV Type 1 Strains from Diverse Areas of the Brain Shows Preponderance of CCR5 Usage. AIDS Res. Hum. Retroviruses 2006, 22, 177–181. [Google Scholar] [CrossRef]
- Abdel-Haq, N.; Hao, H.-N.; Lyman, W.D. Infection of Fetal Human Astrocytes by M-Tropic HIV Is Not Inhibited by Prostaglandin-Mediated CCR5 Downregulation. Pediatr. Res. 1999, 45, 155A. [Google Scholar] [CrossRef]
- Eugenin, E.A.; Osiecki, K.; Lopez, L.; Goldstein, H.; Calderon, T.M.; Berman, J.W. CCL2/Monocyte Chemoattractant Protein-1 Mediates Enhanced Transmigration of Human Immunodeficiency Virus (HIV)-Infected Leukocytes across the Blood–Brain Barrier: A Potential Mechanism of HIV–CNS Invasion and NeuroAIDS. J. Neurosci. 2006, 26, 1098–1106. [Google Scholar] [CrossRef]
- Kaul, M.; Ma, Q.; Medders, K.E.; Desai, M.K.; Lipton, S.A. HIV-1 coreceptors CCR5 and CXCR4 both mediate neuronal cell death but CCR5 paradoxically can also contribute to protection. Cell Death Differ. 2007, 14, 296–305. [Google Scholar] [CrossRef]
- EL-HAGE, N.; DEVER, S.M.; PODHAIZER, E.M.; ARNATT, C.K.; ZHANG, Y.; HAUSER, K.F. A novel bivalent HIV-1 entry inhibitor reveals fundamental differences in CCR5 -μ- opioid receptor interactions in human astroglia and microglia. AIDS 2013, 27, 2181–2190. [Google Scholar] [CrossRef]
- Maung, R.; Hoefer, M.M.; Sanchez, A.B.; Sejbuk, N.E.; Medders, K.E.; Desai, M.K.; Catalan, I.C.; Dowling, C.C.; de Rozieres, C.M.; Garden, G.A.; et al. CCR5 Knockout Prevents Neuronal Injury and Behavioral Impairment Induced in a Transgenic Mouse Model by a CXCR4-using HIV-1 Glycoprotein 120. J. Immunol. 2014, 193, 1895–1910. [Google Scholar] [CrossRef]
- Nickoloff-Bybel, E.A.; Festa, L.; Meucci, O.; Gaskill, P.J. Co-receptor signaling in the pathogenesis of neuroHIV. Retrovirology 2021, 18, 24. [Google Scholar] [CrossRef] [PubMed]
- Sorce, S.; Myburgh, R.; Krause, K.-H. The chemokine receptor CCR5 in the central nervous system. Prog. Neurobiol. 2011, 93, 297–311. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Chen, Y.; Farzan, M.; Choe, H.; Ohagen, A.; Gartner, S.; Busciglio, J.; Yang, X.; Hofmann, W.; Newman, W.; et al. CCR3 and CCR5 are co-receptors for HIV-1 infection of microglia. Nature 1997, 385, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Hahn, Y.K.; Podhaizer, E.M.; McLane, V.D.; Zou, S.; Hauser, K.F.; Knapp, P.E. A central role for glial CCR5 in directing the neuropathological interactions of HIV-1 Tat and opiates. J. Neuroinflamm. 2018, 15, 285. [Google Scholar] [CrossRef] [PubMed]
- Riviere-Cazaux, C.; Cornell, J.; Shen, Y.; Zhou, M. The role of CCR5 in HIV-associated neurocognitive disorders. Heliyon 2022, 8, e09950. [Google Scholar] [CrossRef] [PubMed]
- Bauer, O.; Milenkovic, V.M.; Hilbert, S.; Sarubin, N.; Weigl, J.; Bahr, L.-M.; Wetter, T.C.; Heckel, B.; Wetzel, C.H.; Rupprecht, R.; et al. Association of Chemokine (C-C Motif) Receptor 5 and Ligand 5 with Recovery from Major Depressive Disorder and Related Neurocognitive Impairment. Neuroimmunomodulation 2021, 27, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Nookala, A.R.; Shah, A.; Noel, R.J.; Kumar, A. HIV-1 Tat-Mediated Induction of CCL5 in Astrocytes Involves NF-κB, AP-1, C/EBPα and C/EBPγ Transcription Factors and JAK, PI3K/Akt and p38 MAPK Signaling Pathways. PLoS ONE 2013, 8, e78855. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Greenhill, S.; Huang, S.; Silva, T.K.; Sano, Y.; Wu, S.; Cai, Y.; Nagaoka, Y.; Sehgal, M.; Cai, D.J.; et al. CCR5 is a suppressor for cortical plasticity and hippocampal learning and memory. eLife 2016, 5, e20985. [Google Scholar] [CrossRef] [PubMed]
- Ruhanya, V.; Jacobs, G.B.; Naidoo, S.; Paul, R.H.; Joska, J.A.; Seedat, S.; Nyandoro, G.; Engelbrecht, S.; Glashoff, R.H. Impact of Plasma IP-10/CXCL10 and RANTES/CCL5 Levels on Neurocognitive Function in HIV Treatment-Naive Patients. AIDS Res. Hum. Retroviruses 2021, 37, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Altfeld, M.; Gale Jr, M. Innate immunity against HIV-1 infection. Nat. Immunol. 2015, 16, 554–562. [Google Scholar] [CrossRef]
- Lehner, T.; Wang, Y.; Whittall, T.; Seidl, T. Innate Immunity and HIV-1 Infection. Adv. Dent. Res. 2011, 23, 19–22. [Google Scholar] [CrossRef]
- Motwani, L.; Asif, N.; Patel, A.; Vedantam, D.; Poman, D.S. Neuropathy in Human Immunodeficiency Virus: A Review of the Underlying Pathogenesis and Treatment. Cureus 2022, 14, e25905. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Balinang, J.M.; Paris, J.J.; Hauser, K.F.; Fuss, B.; Knapp, P.E. Effects of HIV-1 Tat on oligodendrocyte viability are mediated by Ca MKII β–GSK 3β interactions. J. Neurochem. 2019, 149, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Holroyd, K.B.; Vishnevetsky, A.; Srinivasan, M.; Saylor, D. Neurologic complications of acute HIV infection. Curr. Treat. Options Infect. Dis. 2020, 12, 227–242. [Google Scholar] [CrossRef]
- Lawrence, D.M.; Major, E.O. HIV-1 and the brain: Connections between HIV-1-associated dementia, neuropathology and neuroimmunology. Microbes Infect. 2002, 4, 301–308. [Google Scholar] [CrossRef]
- Albright, A.V.; Soldan, S.S.; González-Scarano, F. Pathogenesis of human immunodeficiency virus-induced neurological disease. J. Neurovirol. 2003, 9, 222–227. [Google Scholar] [CrossRef]
- Antinori, A.; Arendt, G.; Becker, J.T.; Brew, B.J.; Byrd, D.A.; Cherner, M.; Clifford, D.B.; Cinque, P.; Epstein, L.G.; Goodkin, K.; et al. Updated research nosology for HIV-associated neurocognitive disorders. Neurology 2007, 69, 1789–1799. [Google Scholar] [CrossRef]
- Brown, M.J.; Zhang, J.; Miller, M.C.; Amoatika, D.; Addo, P.N.O.; Kaur, A.; Bagasra, O.; Ingram, L.A. Health disparities in mortality among individuals with HIV-associated dementia in South Carolina. AIDS Care 2023, 36, 291–295. [Google Scholar] [CrossRef]
- Cornea, A.; Lata, I.; Simu, M.; Rosca, E.C. Assessment and Diagnosis of HIV-Associated Dementia. Viruses 2023, 15, 378. [Google Scholar] [CrossRef]
- Wiley, C.A.; Schrier, R.D.; Nelson, J.A.; Lampert, P.W.; Oldstone, M.B. Cellular localization of human immunodeficiency virus infection within the brains of acquired immune deficiency syndrome patients. Proc. Natl. Acad. Sci. USA 1986, 83, 7089–7093. [Google Scholar] [CrossRef] [PubMed]
- Budka, H. Human immunodeficiency virus (HIV) envelope and core proteins in CNS tissues of patients with the acquired immune deficiency syndrome (AIDS). Acta Neuropathol. 1990, 79, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Bissel, S.J.; Wiley, C.A. Human Immunodeficiency Virus Infection of the Brain: Pitfalls in Evaluating Infected/Affected Cell Populations. Brain Pathol. 2006, 14, 97–108. [Google Scholar] [CrossRef]
- Shaik, M.M.; Peng, H.; Lu, J.; Rits-Volloch, S.; Xu, C.; Liao, M.; Chen, B. Structural basis of coreceptor recognition by HIV-1 envelope spike. Nature 2019, 565, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Palomino, D.C.T.; Marti, L.C. Chemokines and immunity. Einstein 2015, 13, 469–473. [Google Scholar] [CrossRef]
- Pittaluga, A. Ccl5–glutamate cross-talk in astrocyte-neuron communication in multiple sclerosis. Front. Immunol. 2017, 8, 1079. [Google Scholar] [CrossRef]
- Dutta, D.; Kundu, M.; Mondal, S.; Roy, A.; Ruehl, S.; Hall, D.A.; Pahan, K. RANTES-induced invasion of Th17 cells into substantia nigra potentiates dopaminergic cell loss in MPTP mouse model of Parkinson’s disease. Neurobiol. Dis. 2019, 132, 104575. [Google Scholar] [CrossRef]
- Nath, A.; Anderson, C.; Jones, M.; Maragos, W.; Booze, R.; Mactutus, C.; Bell, J.; Hauser, K.F.; Mattson, M. Neurotoxicity and dysfunction of dopaminergic systems associated with AIDS dementia. J. Psychopharmacol. 2000, 14, 222–227. [Google Scholar] [CrossRef]
- Haughey, N.J.; Mattson, M.P. Calcium dysregulation and neuronal apoptosis by the HIV-1 proteins Tat and Gp120. J. Acquir. Immune Defic. Syndr. 2002, 31, S55–S61. [Google Scholar] [CrossRef] [PubMed]
- Koutsilieri, E.; Sopper, S.; Scheller, C.; Ter Meulen, V.; Riederer, P. Involvement of dopamine in the progression of AIDS Dementia Complex. J. Neural Transm. 2002, 109, 399–410. [Google Scholar] [CrossRef] [PubMed]
- McLaren, P.J.; Fellay, J. HIV-1 and human genetic variation. Nat. Rev. Genet. 2021, 22, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Langer, S.; Zhang, Z.; Herbert, K.M.; Yoh, S.; König, R.; Chanda, S.K. Sensor Sensibility—HIV-1 and the Innate Immune Response. Cells 2020, 9, 254. [Google Scholar] [CrossRef] [PubMed]
- Carrington, M. Genetics of HIV-1infection: Chemokine receptor CCR5 polymorphism and its consequences. Hum. Mol. Genet. 1999, 8, 1939–1945. [Google Scholar] [CrossRef]
- Vega, J.A.; Villegas-Ospina, S.; Aguilar-Jiménez, W.; Rugeles, M.T.; Bedoya, G.; Zapata, W. Haplotypes in CCR5-CCR2, CCL3 and CCL5 are associated with natural resistance to HIV-1 infection in a Colombian cohort. Biomedica 2017, 37, 267–273. [Google Scholar] [CrossRef]
- Silva, M.J.A.; Marinho, R.L.; Dos Santos, P.A.S.; Dos Santos, C.S.; Ribeiro, L.R.; Rodrigues, Y.C.; Lima, K.V.B.; Lima, L.N.G.C. The Association between CCL5/RANTES SNPs and Susceptibility to HIV-1 Infection: A Meta-Analysis. Viruses 2023, 15, 1958. [Google Scholar] [CrossRef] [PubMed]
- Santana, D.S.; Silva, M.J.A.; de Marin, A.B.R.; Costa, V.L.D.S.; Sousa, G.S.M.; de Sousa, J.G.; Silva, D.C.; da Cruz, E.C.; Lima, L.N.G.C. The Influence Between C-C Chemokine Receptor 5 Genetic Polymorphisms and the Type-1 Human Immunodeficiency Virus: A 20-Year Review. AIDS Res. Hum. Retroviruses 2023, 39, 13–32. [Google Scholar] [CrossRef]
- Zhao, J.; She, S.; Xie, L.; Chen, X.; Mo, C.; Huang, L.; Tang, W.; Chen, X. The Effects of RANTES Polymorphisms on Susceptibility to HIV-1 Infection and Disease Progression: Evidence from an Updated Meta-Analysis. AIDS Res. Hum. Retroviruses 2016, 32, 517–528. [Google Scholar] [CrossRef]
- Ioannidis, J.P.A. Effects of CCR5-Δ 32, CCR2-64I, and SDF-1 3′A Alleles on HIV-1 Disease Progression: An International Meta-Analysis of Individual-Patient Data. Ann. Intern. Med. 2001, 135, 782. [Google Scholar] [CrossRef]
- Jang, D.H.; Choi, B.-S.; Kim, S.S. The effects of RANTES/CCR5 promoter polymorphisms on HIV disease progression in HIV-infected Koreans. Int. J. Immunogenet. 2008, 35, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.J.A.; Ribeiro, L.R.; Gouveia, M.I.M.; Marcelino, B.D.R.; dos Santos, C.S.; Lima, K.V.B.; Lima, L.N.G.C. Hyperinflammatory Response in COVID-19: A Systematic Review. Viruses 2023, 15, 553. [Google Scholar] [CrossRef] [PubMed]
- Oppermann, M. Chemokine receptor CCR5: Insights into structure, function, and regulation. Cell. Signal. 2004, 16, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Van Der Ryst, E. Maraviroc—A CCR5 Antagonist for the Treatment of HIV-1 Infection. Front. Immunol. 2015, 6, 277. [Google Scholar] [CrossRef] [PubMed]
- Milanini, B.; Valcour, V. Differentiating HIV-Associated Neurocognitive Disorders from Alzheimer’s Disease: An Emerging Issue in Geriatric Neuro HIV. Curr. HIV AIDS Rep. 2017, 14, 123. [Google Scholar] [CrossRef] [PubMed]
- Hartley, O.; Offord, R.E. Engineering chemokines to develop optimized HIV inhibitors. Curr. Protein Pept. Sci. 2005, 6, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Vangelista, L.; Secchi, M.; Lusso, P. Rational design of novel HIV-1 entry inhibitors by RANTES engineering. Vaccine 2008, 26, 3008–3015. [Google Scholar] [CrossRef]
- Secchi, M.; Vangelista, L. Rational Engineering of a Sub-Picomolar HIV-1 Blocker. Viruses 2022, 14, 2415. [Google Scholar] [CrossRef]
- Rodríguez-Frade, J.M.; Vila-Coro, A.J.; Martín, A.; Nieto, M.; Sánchez-Madrid, F.; Proudfoot, A.E.; Wells, T.N.; Martínez, A.C.; Mellado, M. Similarities and Differences in RANTES-and (AOP)-RANTES–triggered Signals: Implications for Chemotaxis. J. Cell Biol. 1999, 144, 755–765. [Google Scholar] [CrossRef]
- Appay, V.; Rowland-Jones, S.L. RANTES: A versatile and controversial chemokine. Trends Immunol. 2001, 22, 83–87. [Google Scholar] [CrossRef]
- Kranjc, M.K.; Novak, M.; Pestell, R.G.; Lah, T.T. Cytokine CCL5 and receptor CCR5 axis in glioblastoma multiforme. Radiol. Oncol. 2019, 53, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Lan, T.; Wei, Y.; Wei, X. CCL5/CCR5 axis in human diseases and related treatments. Genes Dis. 2022, 9, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Secchi, M.; Vassena, L.; Morin, S.; Schols, D.; Vangelista, L. Combination of the CCL5-Derived Peptide R4.0 with Different HIV-1 Blockers Reveals Wide Target Compatibility and Synergic Cobinding to CCR5. Antimicrob. Agents Chemother. 2014, 58, 6215–6223. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| N° | Title | Author and Year of Publication/Database/(JBI Score) | Methodology | Results |
|---|---|---|---|---|
| 1 | CCR5 mediates HIV-1 Tat-induced neuroinflammation and influences morphine tolerance, dependence, and reward. | Gonek et al. (2018) [28]/PUBMED/JBI (9/9) | Experimental study | Tat and opioids that act on the mu opioid receptor (MORs) seem to promote CCR5 and CCR2 signaling, causing neuroinflammation and striatal neurotoxicity to increase. The reduced therapeutic effectiveness and enhanced rewarding characteristics of morphine that we have discovered herein may be caused by these effects. |
| 2 | Differential expression of the alternatively spliced OPRM1 isoform μ-opioid receptor-1K in HIV-infected individuals. | Dever et al. (2014) [29]/PUBMED/JBI (10/11) | Cohort | HIVE was linked to enhanced expression of MCP-1, MCP-2, and CCL5/RANTES. |
| 3 | Nonproductive human immunodeficiency virus type 1 infection of human fetal astrocytes: independence from CD4 and major chemokine receptors. | Sabri et al. (1999) [30]/PUBMED and Science Direct/JBI (8/9) | Experimental study | On the cell surface of human fetal astrocytes, there is no indication of any of the key HIV-1 coreceptors, such as CXCR4, CCR5, CCR3, and CCR2b, or the CD4 molecule. However, RT-PCR was able to identify the mRNA transcripts for CXCR4, CCR5, Bonzo/STRL33/TYMSTR, and APJ. Furthermore, results demonstrate that primary HIV-1 isolates can infect astrocytes by a mechanism unrelated to CD4 or important chemokine receptors (such CCL5/RANTES). Additionally, astrocytes have the potential to be latent HIV-1 carriers and, upon activation, may play a role in the transmission of the infection to nearby cells like macrophages or microglia. |
| 4 | Encephalopathy in AIDS—increased formation of beta-chemokines in monocytes after HIV-1 virus infection: mechanisms of CNS involvement. | Mayer; Schmidtmayerová (1997) [31]/PUBMED/JBI (10/11) | Cohort | When HIV illness is advanced, dementia is linked to an increased viral load in the brain. The most plausible pathogenetic mechanism causing brain dysfunction and damage is neurotoxicity linked to the activity of macrophages and microglial cells that are HIV-infected. Our research has shown that HIV infection of macrophages significantly increases the production of the chemokines MIP-1-alpha, MIP-1-beta, and RANTES/CCL5 (subgroup C-C). Due to their chemoattractant and activating qualities, these substances may contribute to leukocytosis and inflammation, which in turn raises the population of HIV-susceptible cells, facilitates their infection, and ultimately increases intrathecal virus spread. |
| 5 | Microglia express CCR5, CXCR4, and CCR3, but of these, CCR5 is the principal coreceptor for human immunodeficiency virus type 1 dementia isolates. | Albright et al. (1999) [32]/PUBMED/JBI (9/9) | Experimental study | In HIV dementia (HIVD) patients, CCR5 was expressed at greater levels by microglia than CCR3 or CXCR4. It revealed that only CCR5 and CXCR4 of these three chemokine receptors could transduce a signal in microglia in response to their respective ligands, MIP-1 and SDF-1. Since the anti-CCR5 antibody 2D7 was able to significantly reduce microglial infection by both wild-type and single-round luciferase pseudotype reporter viruses, we also discovered that CCR5 is the main coreceptor used for infection of human adult microglia by the HIV type 1 dementia isolates HIV-1DS-br, HIV-1RC-br, and HIV-1YU-2. |
| 6 | A Bivalent Ligand Targeting the Putative Mu Opioid Receptor and Chemokine Receptor CCR5 Heterodimers: Binding Affinity versus Functional Activities. | Yuan et al. (2013) [33]/PUBMED/JBI (9/9) | Experimental study | While the CCR5 and MOR may coexist as heterodimers in human astrocytes, the CCR5 receptor may exist as a monomer or form homodimers in monoclonal receptor-expressed assays. The CCR5 binding pocket in the heterodimer may accommodate the bivalent ligand, preferably to cause a blockage of viral entry. |
| 7 | Genetic knockouts suggest a critical role for HIV co-receptors in models of HIV gp120-induced brain injury. | Maung et al. (2012) [34]/PUBMED/JBI (11/11) | Review | Apoptosis and neuronal damage are brought on by HIV-1 proteins like gp120, which drive macrophages to generate neurotoxins. Additionally, the two main HIV co-receptors, the chemokine receptors CCR5 and CXCR4, have a wide range of physiological uses and are expressed in cells other than immune cells, such as those in the brain. Transgenic mice with HIV gp120 expression in the brain resemble NeuroAIDS brains in various pathogenic ways. Among CCR5-expressing and CCR5-deficient animals, there does not appear to be a difference in peripheral production of the chemokines CCL5/RANTES, CCL3/MIP-1, or CCL4/MIP-1ß. However, in contrast to wild-type controls, CCR5 Knockout (KO) mice exhibit improved humoral or cell-mediated immune responses, depending on the antigen trigger. |
| 8 | Autophagy is increased in postmortem brains of persons with HIV-1-associated encephalitis. | Zhou; Masliah; Spector (2011) [35]/PUBMED/JBI (9/10) | Case-control | In neural cells exposed to either CXCR4- or CCR5-tropic HIV-1 gp120, levels of autophagic proteins and autophagosomes were elevated. The etiology of neuroAIDS is significantly influenced by the dysregulation of autophagy during HIV infection. |
| 9 | Immunoregulation of a CB2 receptor agonist in a murine model of neuroAIDS. | Gorantla et al. (2012) [36]/PUBMED/JBI (9/9) | Experimental study | In neural cells exposed to either CXCR4- or CCR5-tropic HIV-1 gp120, levels of autophagic proteins and autophagosomes were elevated. CCR5 expression on CD4+ cells was considerably decreased with Gp1a therapy in both immunodeficient mice reconstituted with human peripheral blood lymphocytes (hu-PBL) and HIVE mice, according to flow cytometric studies. Between the treatment and control groups, the levels of CCR5 expression on CD8+ cells were not substantially different. |
| 10 | Chemokine receptor-5 (CCR5) is a receptor for the HIV entry inhibitor peptide T (DAPTA). | Polianova; Ruscetti; Ruff (2005) [37]/PUBMED/JBI (9/9) | Experimental study | According to their results, DAPTA (a CCR5 antagonist) reduces the binding of the viral envelope to CCR5 in R5-tropic HIV isolates. This inhibition most likely occurs through competitive binding of peptide T (DAPTA), a non-toxic experimental antiviral entry inhibitor derived from HIV-1, to CCR5. The degradation of gp120’s interaction to the chemokine receptor CCR5 is what gives DAPTA its specific antiviral effect for HIV-1 R5-tropic viruses. |
| 11 | Viral and cellular factors underlying neuropathogenesis in HIV associated neurocognitive disorders (HAND). | Rao; Ruiz; Prasad (2014) [38]/PUBMED/JBI (11/11) | Review | Although neurons do not support HIV-1 infection or replication, they do express several cell-surface receptors (such as CCR5, CXCR4, NMDAR, etc.) that make them vulnerable to insults from viral proteins (such as Tat, gp120), inflammatory cytokines (such as TNF-alpha, IL-1), and small metabolites (such as nitric oxide, arachidonic acid, etc.) secreted by immune cells in the brain. The N-methyl-D-aspartate receptors (NMDAR), the low-density lipoprotein receptor-related protein (LRP), the chemokine receptors CCR5 and CXCR4, and the dopamine transporter are the key mediators of this susceptibility. HIV-1 gp120 can bind to either CCR5 or CXCR4 and cause neuroblastoma cell death. Gp120 can directly bind to CCR5 or CXCR4, starting a signaling cascade driven by p38-MAPK that causes neuronal death. The natural ligands of both CCR5 (such as CCL5, CCL3) and CXCR4 (such as CXCL12) were discovered to be neuroprotective against gp120 neurotoxicity. |
| 12 | HIV-1 Envelope Protein Gp120 Up-Regulates CCL5 Production in Astrocytes Which Can Be Circumvented by Inhibitors of NF-ΚB Pathway. | Shah et al. (2011) [39]/PUBMED/JBI (10/11) | Cohort | The function of CCL5 in promoting CCL2 production, which can facilitate leukocytes infected with HIV-1 migrating through the BBB. The gp120-induced mRNA expression of CCL5 is time-dependent. Increased CCL5 levels may encourage monocyte migration through the BBB, aggravating neuroinflammatory illness. Additionally, CCL5 has a protective function in the setting of HIV-1 infection at doses of 10 nanomoles (nM) (equal to 78.76 ng/mL), promoting neuronal survival under pro-apoptotic conditions. However, the maximal levels of CCL5 induction by gp120 in the current investigation were only 3 ng/mL, preventing the concentration from being protective. CCL5 rise mediated by Gp120 can only be understood as responsible for a neuroinflammatory response. A particular NF-kB inhibitor and siRNA might partially stop the elevated CCL5 synthesis. |
| 13 | Vpr- and Nef-Dependent Induction of RANTES/CCL5 in Microglial Cells. | Si et al. (2002) [40]/PUBMED/JBI (9/9) | Experimental study | HIVD patients’ CSF fluids have been discovered to have elevated levels of RANTES/CCL5. These brains exhibit both HIV-1 and inflammatory mediators, which can promote RANTES/CCL5 synthesis. In astrocytes, microglia, and macrophages, CCL5/RANTES can be activated by cytokines and viral substances (recombinant HIV-1 proteins). CCL5 is produced with a delayed kinetics associated with increased viral replication, and necessitates HIV-1 infection. CCL5 secretion is unaffected by X4-viruses, and is not inhibited by cytokine antagonists or antibodies. The viral accessory protein Vpr, in addition to Nef, is required for the synthesis of RANTES/CCL5, indicating a unique function for Vpr in chemokine induction in primary macrophage-type cells. Additionally, the p38 MAP kinase lowers chemokine expression in microglia. |
| 14 | HIV-1 Nef Induces CCL5 Production in Astrocytes through P38-MAPK and PI3K/Akt Pathway and Utilizes NF-KB, CEBP and AP-1 Transcription Factors. | Liu et al. (2014) [41]/PUBMED/JBI (7/9) | Experimental study | Nef astrocytes exhibit a substantial induction of CCL5. CCL5 expression by HIV-1 proteins has shown that NF-kB plays a significant role. Through the p38-MAPK and PI3K/Akt pathways, HIV-1 Nef stimulates the synthesis of CCL5 in astrocytes while making use of the transcription factors NF-kB, CEBP, and AP-1. |
| 15 | Characterization of HIV-1 Infection in Microglia-Containing Human Cerebral Organoids. | Gumbs et al. (2022) [42]/PUBMED/JBI (9/9) | Experimental study | The CCR5 co-receptor supports productive HIV infection in cerebral organoids and isolated organoid-derived microglia (oMG), both of which have been infected with replication-competent HIV reporter viruses. The co-expression of markers unique to microglia and the HIV CD4 and CCR5 receptors was necessary for HIV infection susceptibility. |
| 16 | Cellular localization of the chemokine receptor CCR5. Correlation to cellular targets of HIV-1 infection. | Rottman et al. (1997) [43]/PUBMED/JBI (9/9) | Experimental study | CCR5 is expressed by astrocytes, microglia, and neurons. When there is chronic inflammation, there are more CCR5-immunoreactive cells present, and this immunoreactivity is positively correlated with the severity of the inflammation as measured by histopathology. |
| 17 | Interaction of the CC-Chemokine RANTES with Glycosaminoglycans Activates a p44/p42 Mitogen-Activated Protein Kinase-Dependent Signaling Pathway and Enhances Human Immunodeficiency Virus Type 1 Infectivity. | Chang et al. (2002) [44]/PUBMED and Science Direct/JBI (11/11) | Cohort | It implies that RANTES can activate signaling pathways through an alternate receptor or receptors that are GAG dependent, but which are presumably not known, particular GPCR chemokine receptors, at both low, physiologically appropriate concentrations and at greater, potentially supraphysiological concentrations. At a post entry stage of the viral life cycle, the subsequent activation of MAPK at high RANTES concentrations can improve the efficacy of HIV-1 replication. Protein tyrosine kinases (PTK)- and MAPK-dependent signaling pathway is activated by the interaction of CCL5/RANTES with cell surface, glycosaminoglycans (GAGs). |
| 18 | Rantes distribution and cellular localization in the brain of HIV-infected patients. | Vago et al. (2001) [45]/PUBMED/JBI (10/10) | Case-control | RANTES was highly expressed in instances of inflamed brain lesions (22/24 HIV-positive patients and 2/7 HIV-negative patients). Microglial cells and lymphocytes were found to be positive in both diffuse and nodular areas. Only individuals who tested positive for HIV had positive astrocytes. RANTES was never negative in multinucleated giant cells. |
| 19 | CCR5 blockade for neuroinflammatory diseases--beyond control of HIV. | Martin-Blondel et al. (2016) [46]/PUBMED/JBI (10/11) | Review | HIV infection of CNS-resident cells is facilitated by CCR5, which is expressed in the CNS of HIV patients. Because CCR5 encourages the migration of immune cells into the CNS, it may have a role in the neurocognitive dysfunction linked to HIV. In experimental animals, the maraviroc monotherapy (a CCR5 antagonist drug) decreased microglial activation, major pro-inflammatory mediator expression, and levels of indicators of neuronal damage. The addition of maraviroc medication to antiretroviral therapy may lead to improvements in neurocognitive state, according to small trials. |
| 20 | Increased CCR5 Affinity and Reduced CCR5/CD4 Dependence of a Neurovirulent Primary Human Immunodeficiency Virus Type 1 Isolate. | Gorry et al. (2002) [47]/PUBMED/JBI (11/11) | Cohort | In some CNS illness patients, HIV-1 variants with elevated CCR5 affinity and decreased dependency on CCR5/CD4 might be present. These viruses would result from adaptive evolution aimed at infecting target cells that express relatively little CD4. Our research further indicates that a subgroup of neurotropic R5-viruses with higher CCR5 affinity and neurotropic R5X4-viruses may be involved in CNS neurodegenerative pathways. |
| 21 | Env gp120 sequence analysis of HIV type 1 strains from diverse areas of the brain shows preponderance of CCR5 usage. | Shah et al. (2006) [48]/Science Direct/JBI (9/10) | Case-control | CCR5, CXCR4, and CCR3 are expressed by microglia in the brain. It was postulated that HIV-1 neurodegenerative symptoms may be caused by enhanced CCR5 affinity, a pathogenic viral phenotype. HIV-1 from peripheral nerves is mostly CCR5 dependent and causes neuroinflammation and neuronal death. |
| 22 | Infection of Fetal Human Astrocytes by M-Tropic HIV Is Not Inhibited by Prostaglandin-Mediated CCR5 Downregulation. | Abdel-Haq; Hao; Lyman (1999) [49]/PUBMED/JBI (10/11) | Cohort | The expression of CCR5 can be reduced in human astrocytes by prostaglandins (PGE1, PGE2, and PGF1α), however they are unable to prevent infection by a monotropic strain of HIV. This implies that other coreceptors, which have not yet been discovered, are involved in the CNS infection of HIV and the development of neuroAIDS. |
| 23 | CCL2/Monocyte Chemoattractant Protein-1 Mediates Enhanced Transmigration of Human Immunodeficiency Virus (HIV)-Infected Leukocytes across the Blood–Brain Barrier: A Potential Mechanism of HIV–CNS Invasion and NeuroAIDS. | Eugenin et al. (2006) [50]/PUBMED/JBI (8/9) | Experimental study | Few monocytes infected with the X4-virus or lymphocytes infected with HIV contribute to increased monocyte transmigration in any manner. However, when R5-viruses are utilized, the extent of monocyte transmigration is much greater, demonstrating that viral tropism is critical in promoting monocyte transmigration and subsequent BBB rupture. On HIV-infected leukocytes, CXCR3 and CCR5 are retained or expressed at higher levels. |
| 24 | HIV-1 coreceptors CCR5 and CXCR4 both mediate neuronal cell death but CCR5 paradoxically can also contribute to protection. | Kaul et al. (2005) [51]/PUBMED/JBI (9/9) | Experimental study | RANTES and MIP-1β, which activate an Akt-dependent signaling pathway, provide neuroprotection. Contrarily, gp120, which prefers CCR5, causes neuronal death. Depending on the precise situation, the properties of the ligand, and the ensuing signaling cascade, CCR5 can either benefit or hurt the neurological system. Heterologous desensitization of CXCR4 receptors on neurons underlies the neuroprotective effects of CCR5-binding MIP-1β and RANTES. |
| 25 | A novel bivalent HIV-1 entry inhibitor reveals fundamental differences in CCR5-μ-opioid receptor interactions between human astroglia and microglia. | El-Hage et al. (2013) [52]/PUBMED/JBI (9/9) | Experimental study | It is believed that gp120/gp41-CCR5 interactions will occur when CD4 is present on microglia but not astrocytes. In addition to CCR5, it was shown that human microglia express MOR at extremely low relative levels as compared to astrocytes. When MOR is activated, CCR5 is upregulated in human host cells, which promotes HIV-1 infection and replication. |
| 26 | CCR5 knockout prevents neuronal injury and behavioral impairment induced in a transgenic mouse model by a CXCR4-using HIV-1 glycoprotein 120. | Maung et al. (2014)/PUBMED [53]/JBI (7/9) | Experimental study | Innate immune response elements might be triggered by gp120 without the need for the CCR5 or neuronal damage. CCL2, CCL5, and CXCL10 are a few of the up-regulated factors that have been linked to HIV neuropathogenesis. Neurotoxicity in HIVD is CCR5 dependent. |
| 27 | Co-receptor signaling in the pathogenesis of neuroHIV. | Nickoloff-Bybel et al. (2021) [54]/PUBMED/JBI (11/11) | Review | Patients with HIVE have some viral proteins in their brains, including gp120, which may have a neurotoxic effect. It was discovered a connection between CSF viral levels and neurological impairment, further pointing to a connection between viral persistence and neuroHIV. Interactions between host CNS cells and the infected cells that make up a persistent reservoir of HIV in the brain are a major contributor to neuropathogenesis in people on cART. By binding to both CXCR4 and CCR5 on neurons, the viral proteins that are shed, such as gp120, actively cause neurotoxicity. By secreting neuroinflammatory mediators including inflammatory cytokines and chemokines, infected and activated macrophages and microglia induce neuronal damage. The interaction of HIV virions with CXCR4 and CCR5 can activate other myeloid populations and trigger signaling pathways that might directly cause neuroinflammation. |
| 28 | The chemokine receptor CCR5 in the central nervous system. | Sorce et al. (2011) [55]/Science Direct/JBI (10/11) | Review | Through preventing interactions between the HIV gp-120 and the host CD4+ T cell and macrophage CCR5 receptors, CCR5 antagonists reduce HIV adherence to target cells in the host. |
| 29 | CCR3 and CCR5 are co-receptors for HIV-1 infection of microglia. | He et al. (1997) [56]/PUBMED/JBI (11/11) | Cohort | CCR3 and CCR5 are both utilized to allow viral entrance in microglial cells. Subsets of HIV-1 isolates that are M-tropic primarily utilize CCR3, with CCR5 serving as a secondary receptor. |
| 30 | A central role for glial CCR5 in directing the neuropathological interactions of HIV-1 Tat and opiates. | Kim et al. (2018) [57]/PUBMED/JBI (10/11) | Cohort | Striatum neurons were shielded against co-exposure to HIV-1 Tat and morphine by loss of glial but not neuronal CCR5. Loss of CCR5 prevented the interaction between Tat and morphine and restored the toxic effects of Tat in the cultures while morphine was still present. In mechanisms unrelated to its crucial involvement in HIV infection, CCR5 is implicated, at least in part through changing the equilibrium between proBDNF and mBDNF levels. The varied morphine/opioid toxicity in the presence or absence of CCR5 also raises questions about the complicated interactions between CCR5 and MOR, which may entail heterologous interactions. |
| 31 | The role of CCR5 in HIV-associated neurocognitive disorders. | Riviere-Cazaux et al. (2022) [58]/PUBMED/JBI (11/11) | Review | In addition to impairing axonal regeneration following neuronal injury, CCR5 activation can result in neuronal cAMP response element-binding protein (CREB) and mitogen-activated protein kinase (MAPK) inactivation. The gp120 V3 domain’s direct association with CCR5 may also impair synaptic plasticity, which would inhibit memory without triggering neuroinflammation. |
| 32 | Association of Chemokine (C-C Motif) Receptor 5 and Ligand 5 with Recovery from Major Depressive Disorder and Related Neurocognitive Impairment. | Bauer et al. (2021) [59]/PUBMED/JBI (10/11) | Cohort | Even before starting therapy, CCR5 and CCL5 levels were much lower in the responder group than in the nonresponder group. Regarding neurocognitive impairment in MDD patients, it was discovered that after five weeks of treatment, a greater misperception of the emotion “anger” was linked to a more pronounced change in CCR5, and that a faster perception of the emotion “disgust” was linked to a greater decrease in CCL5 over the same period. |
| 33 | HIV-1 Tat-Mediated Induction of CCL5 in Astrocytes Involves NF-κB, AP-1, C/EBPα and C/EBPγ Transcription Factors and JAK, PI3K/Akt and p38 MAPK Signaling Pathways. | Nookala et al. (2013) [60]/PUBMED/JBI (9/9) | Experimental study | Peak mRNA and protein levels for CCL5 expression were seen in Tat at 1 h and 48 h after transfection, respectively, indicating a time-dependent increase in the expression of the gene. CCL5 was up-regulated by AP-1, C/EBP-alpha, and C/EBP-gamma. |
| 34 | CCR5 is a suppressor for cortical plasticity and hippocampal learning and memory. | Zhou et al. (2016) [61]/PUBMED/JBI (8/9) | Experimental study | Since CCR5 is a potent memory and plasticity suppressor, HIV-related cognitive impairments may be exacerbated by viral proteins that overactivate CCR5. |
| 35 | Impact of Plasma IP-10/CXCL10 and RANTES/CCL5 Levels on Neurocognitive Function in HIV Treatment-Naive Patients. | Ruhanya et al. (2021) [62]/PUBMED/JBI (11/11) | Cohort | The most reliable indicators of the global deficit score (GDS) associated with neurocognitive impairment were IP-10 and RANTES. Both cytokines had an inverse relationship with CD4+ T cell counts and were positively connected with lymphocyte proviral load and plasma viral load. With the greatest closeness, IP-10 and RANTES formed their own cluster. |
| 36 | New Challenges of HIV-1 Infection: How HIV-1 Attacks and Resides in the Central Nervous System. | Rojas-Celis et al. (2019) [5]/PUBMED/JBI (11/11) | Review | Since macrophages are CD4+ cells and express the CXCR4 and CCR5 coreceptors, they are as vulnerable to HIV-1 infection as microglia are. However, the most common coreceptor in macrophages is CCR5, as data show that CXCR4-mediated entry of HIV-1 particles into macrophages does not confer any potential for infection. CCR5 is more strongly linked than CCR3 to viral entrance and the subsequent onset of dementia. It is feasible to find HIV-1 as infectious virions, viral proteins, and even nucleic acids in cultured astrocytes even though they do not exhibit CD4 or CCR5. Despite not expressing CD4 or CCR5, oligodendrocytes do express CXCR4. CXCR4, CCR5, and CCR3 are surface-expressed receptors in adult neurons. Inflammatory cytokines, ROS, and RNS that surround neurons provide an environment that is pro-inflammatory and reactive, which harms the neurons. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, M.J.A.; Marinho, R.L.; Rodrigues, Y.C.; Brasil, T.P.; Dos Santos, P.A.S.; Silva, C.S.; Sardinha, D.M.; Lima, K.V.B.; Lima, L.N.G.C. Molecular Role of HIV-1 Human Receptors (CCL5–CCR5 Axis) in neuroAIDS: A Systematic Review. Microorganisms 2024, 12, 782. https://doi.org/10.3390/microorganisms12040782
Silva MJA, Marinho RL, Rodrigues YC, Brasil TP, Dos Santos PAS, Silva CS, Sardinha DM, Lima KVB, Lima LNGC. Molecular Role of HIV-1 Human Receptors (CCL5–CCR5 Axis) in neuroAIDS: A Systematic Review. Microorganisms. 2024; 12(4):782. https://doi.org/10.3390/microorganisms12040782
Chicago/Turabian StyleSilva, Marcos Jessé Abrahão, Rebecca Lobato Marinho, Yan Corrêa Rodrigues, Thiago Pinto Brasil, Pabllo Antonny Silva Dos Santos, Caroliny Soares Silva, Daniele Melo Sardinha, Karla Valéria Batista Lima, and Luana Nepomuceno Gondim Costa Lima. 2024. "Molecular Role of HIV-1 Human Receptors (CCL5–CCR5 Axis) in neuroAIDS: A Systematic Review" Microorganisms 12, no. 4: 782. https://doi.org/10.3390/microorganisms12040782
APA StyleSilva, M. J. A., Marinho, R. L., Rodrigues, Y. C., Brasil, T. P., Dos Santos, P. A. S., Silva, C. S., Sardinha, D. M., Lima, K. V. B., & Lima, L. N. G. C. (2024). Molecular Role of HIV-1 Human Receptors (CCL5–CCR5 Axis) in neuroAIDS: A Systematic Review. Microorganisms, 12(4), 782. https://doi.org/10.3390/microorganisms12040782