
The Combination of Biochar and Bacillus subtilis Biological Agent Reduced the Relative Abundance of Pathogenic Bacteria in the Rhizosphere Soil of Panax notoginseng


Abstract
1. Introduction
2. Materials and Methods
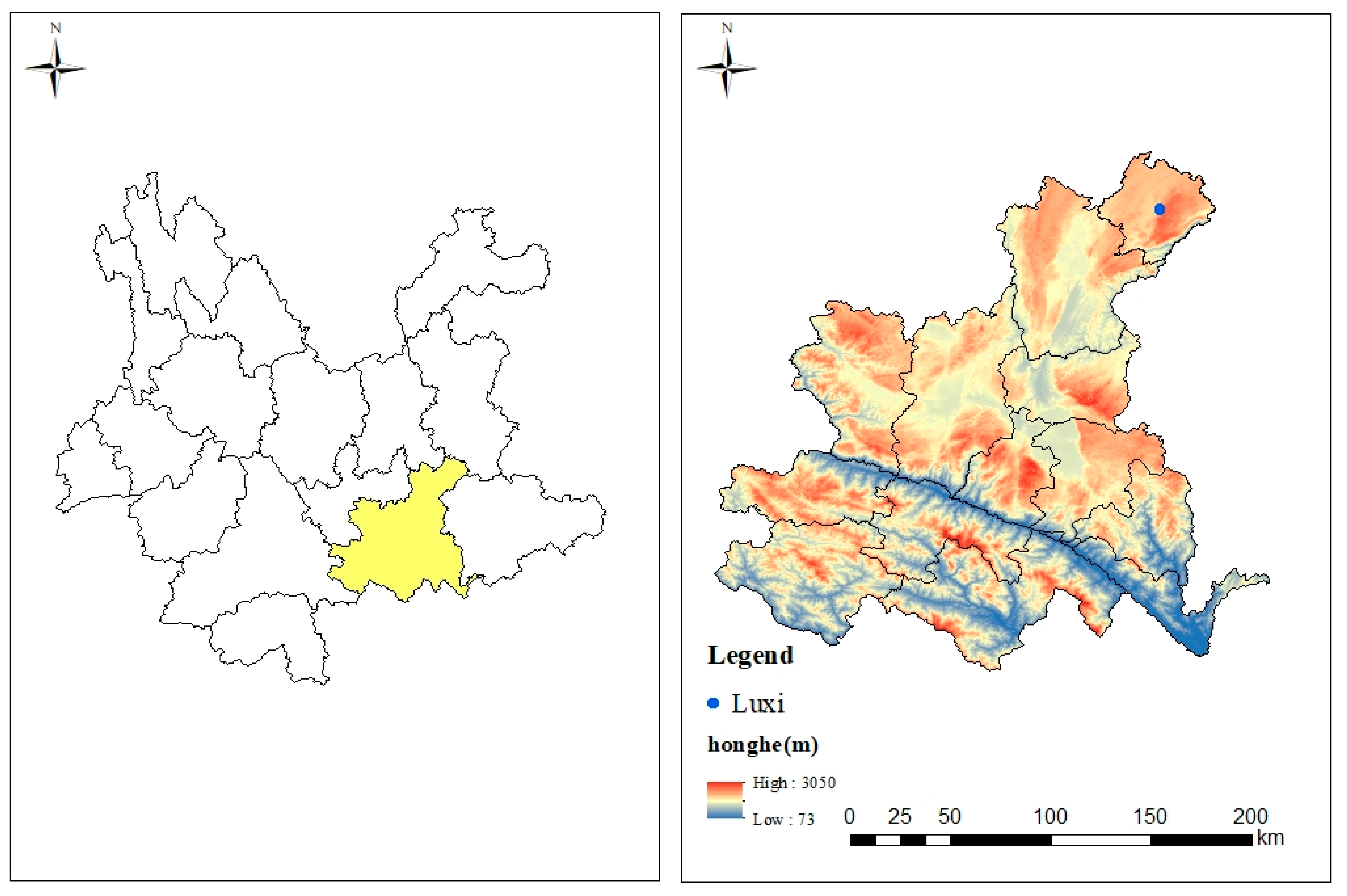
2.1. Experimental Site and Experimental Design
2.2. DNA Extraction and Amplification
2.3. Bioinformatic Analysis
2.4. Statistical Analysis
3. Results
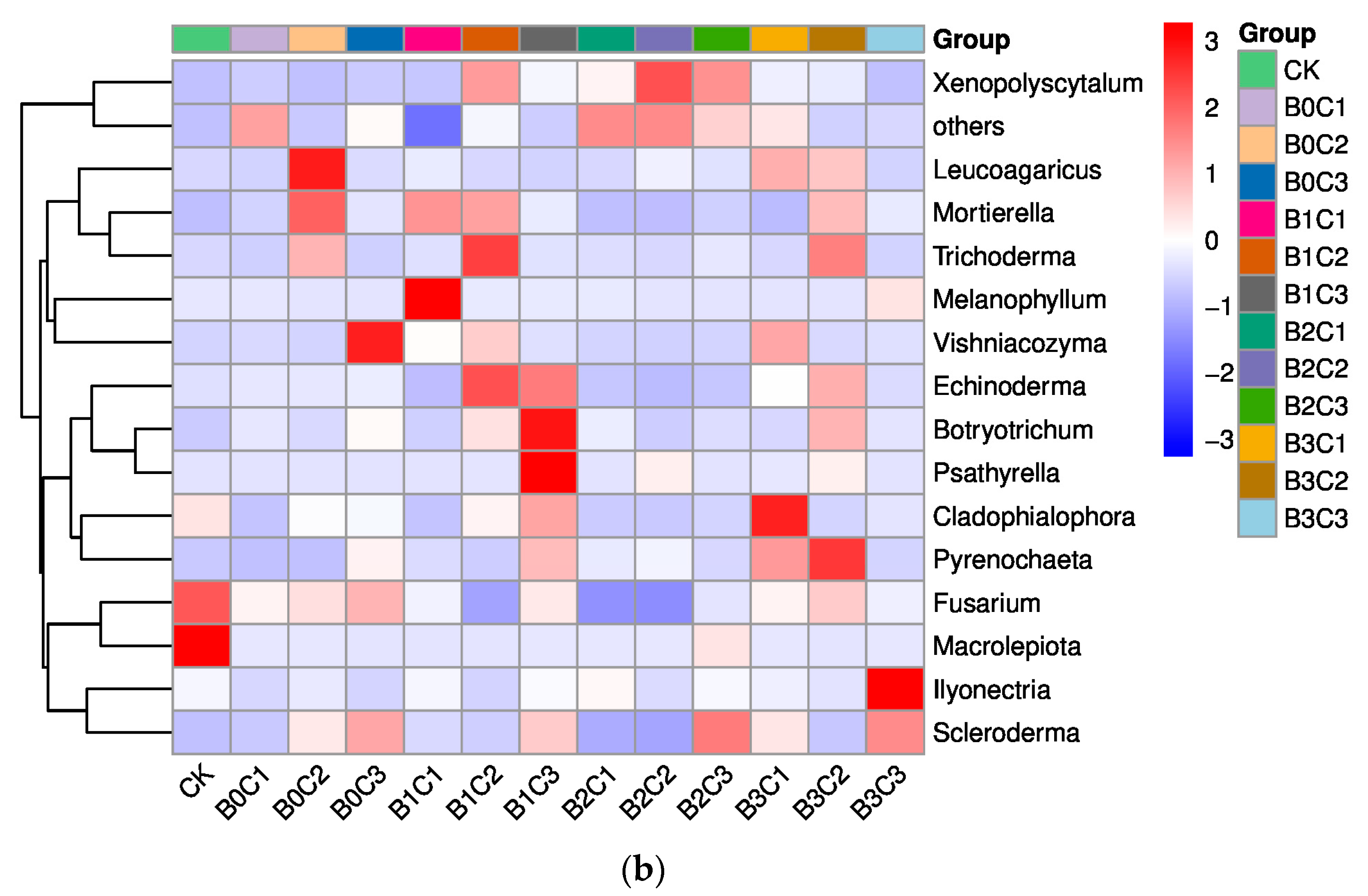
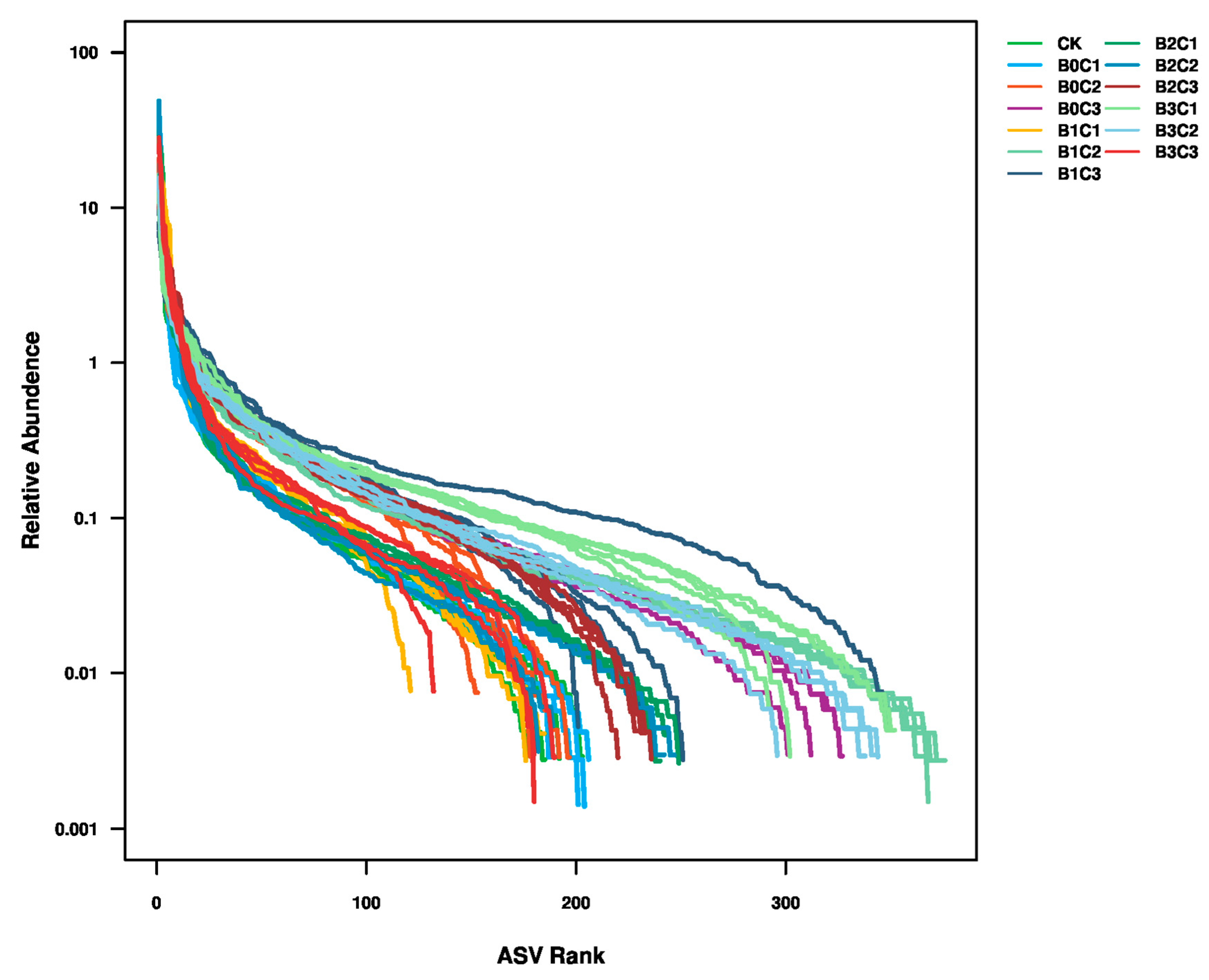
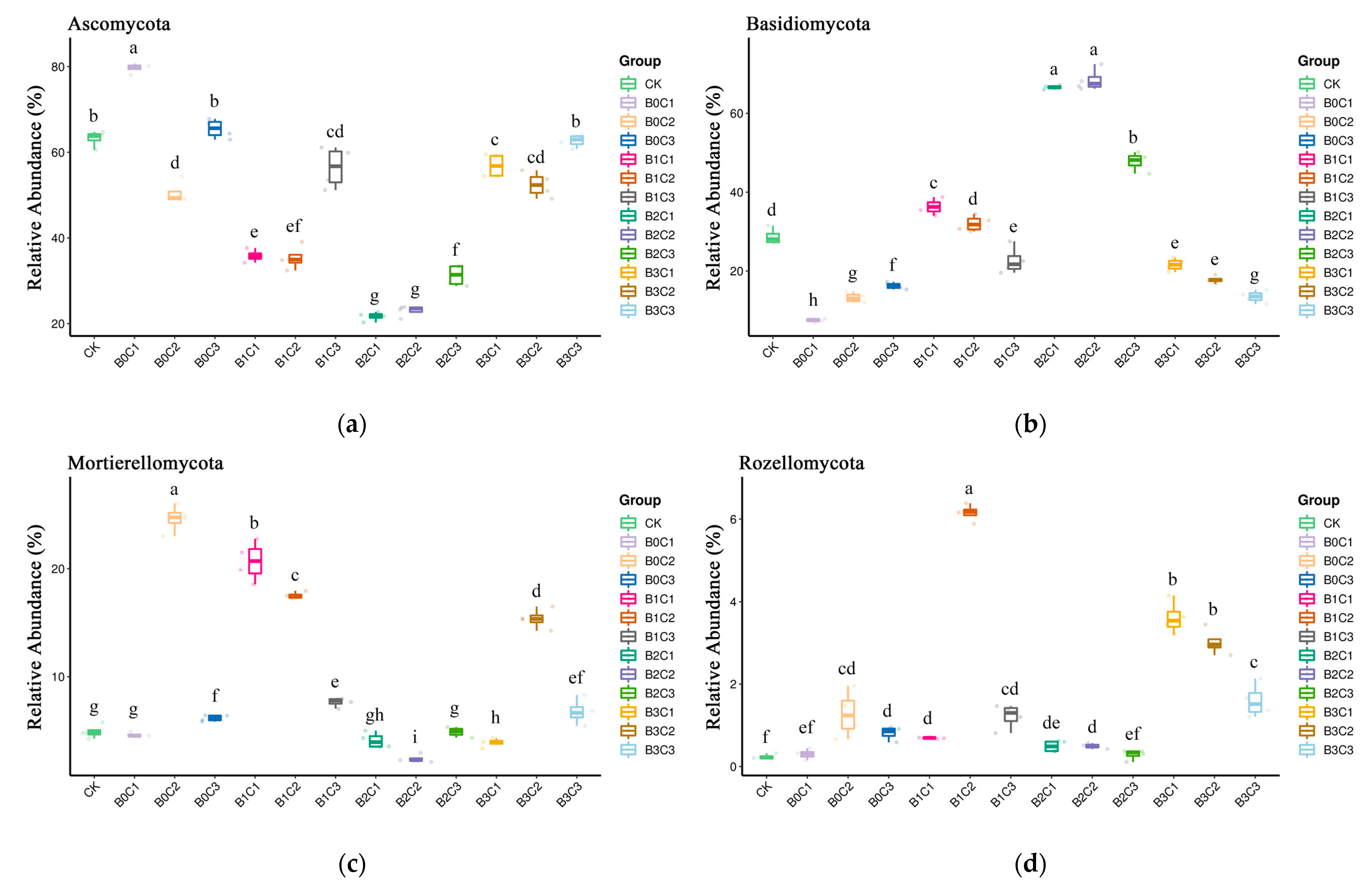
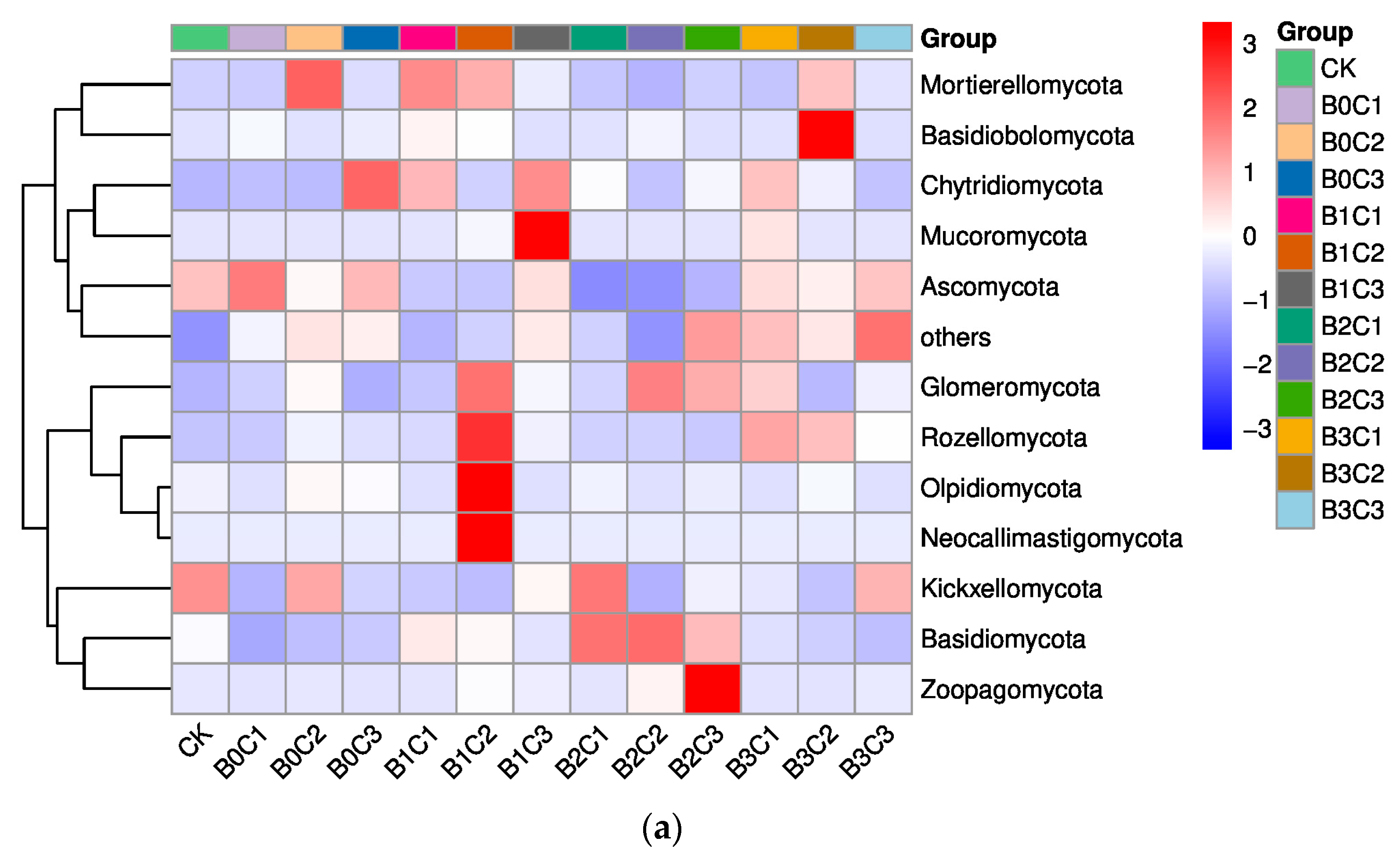
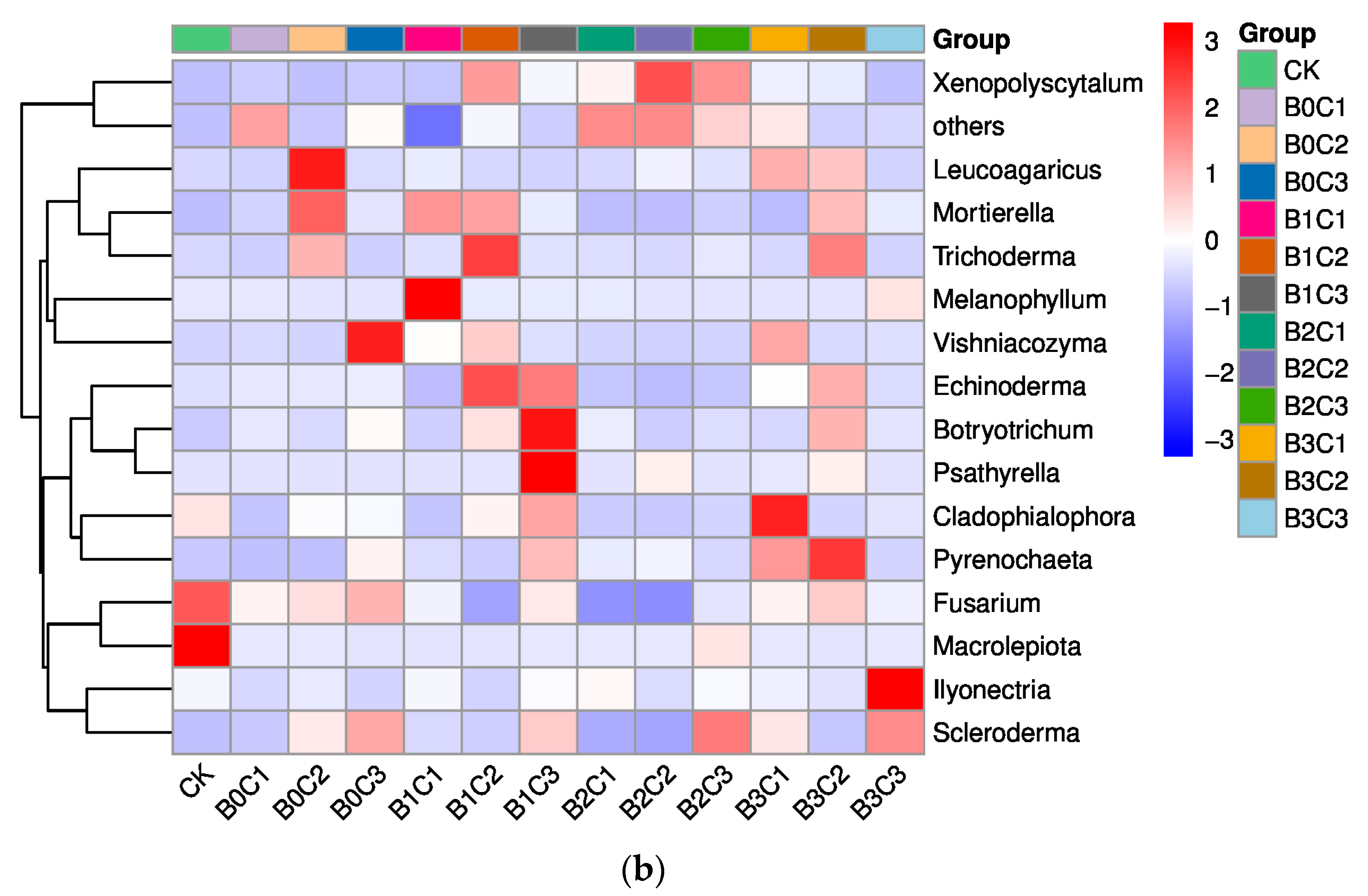
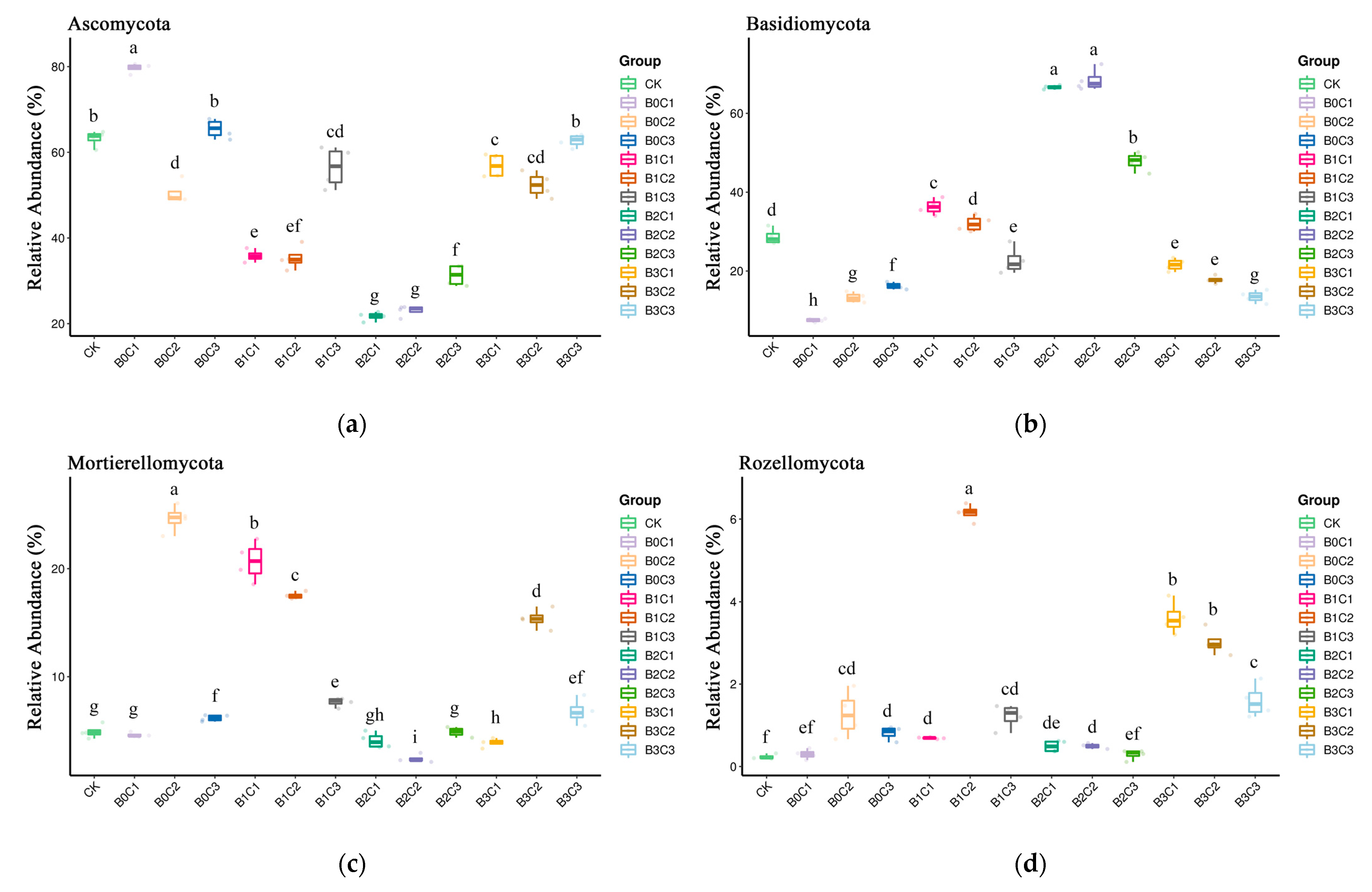
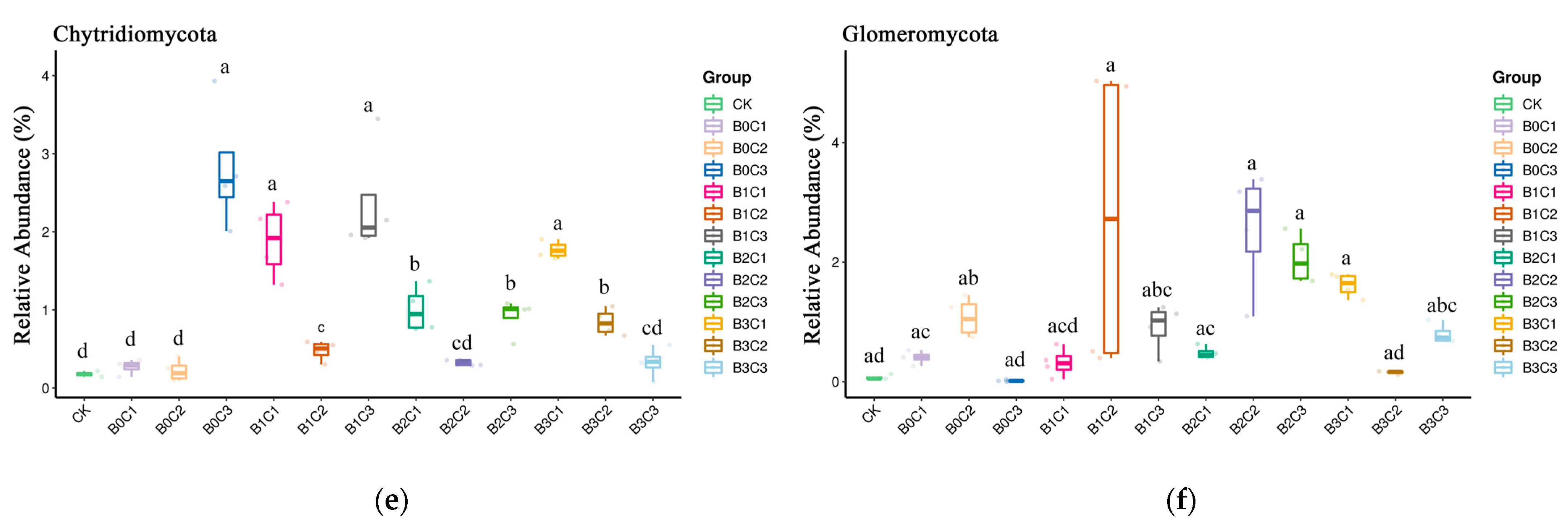
3.1. Community Structure of Fungi in Rhizosphere Soil of Panax notoginseng
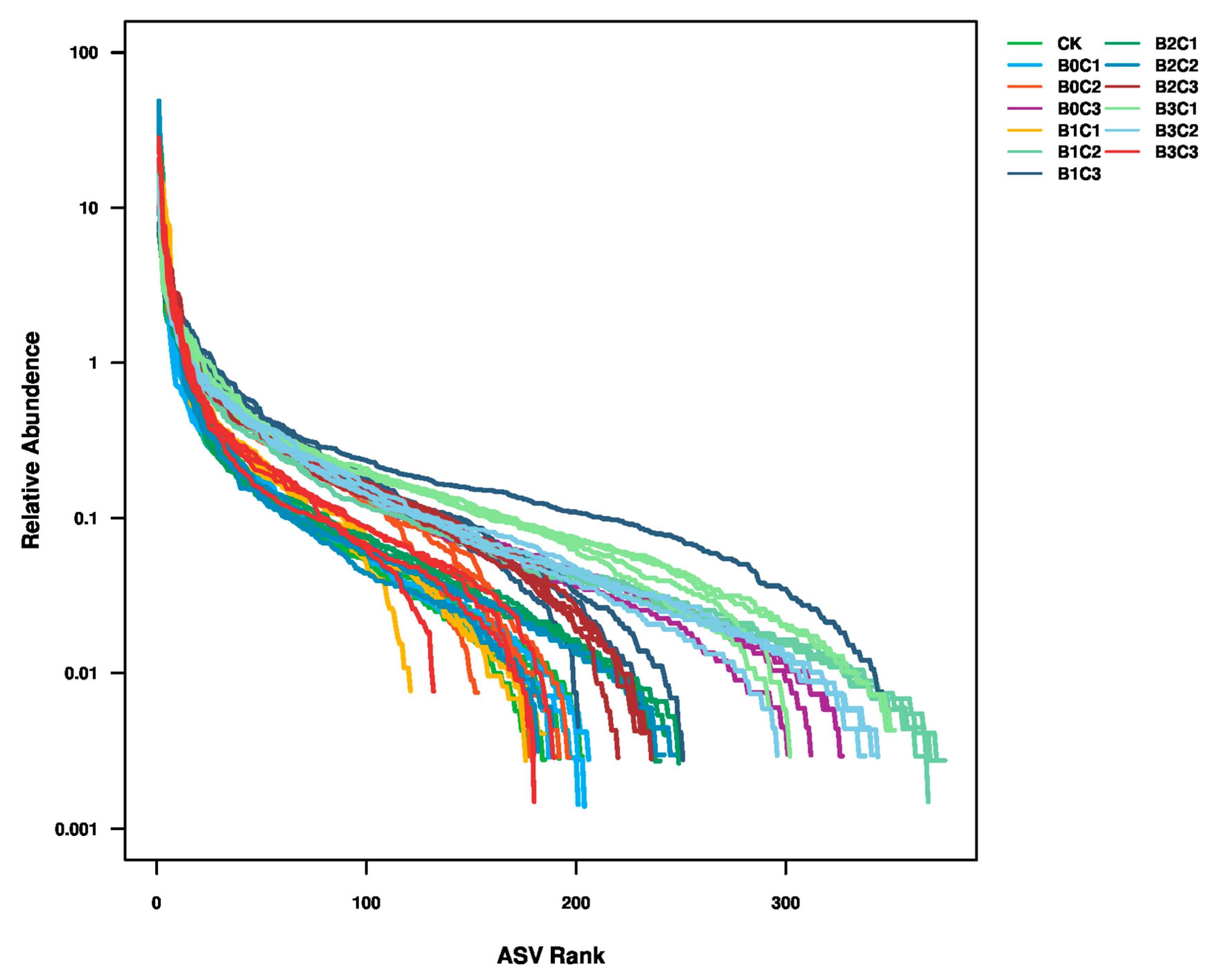
3.2. Fungal Diversity in Rhizosphere Soil
3.2.1. Alpha Diversity Analysis
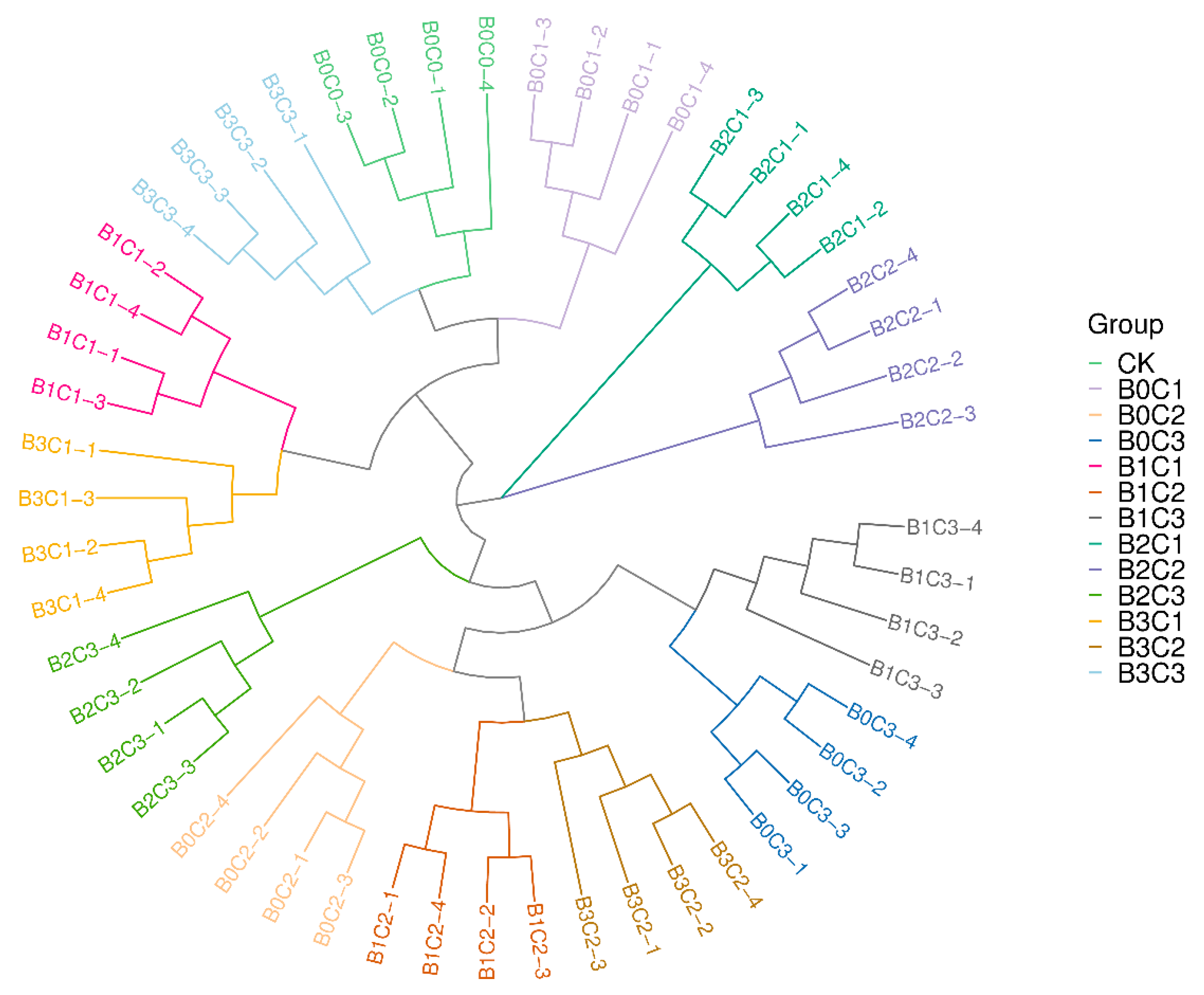
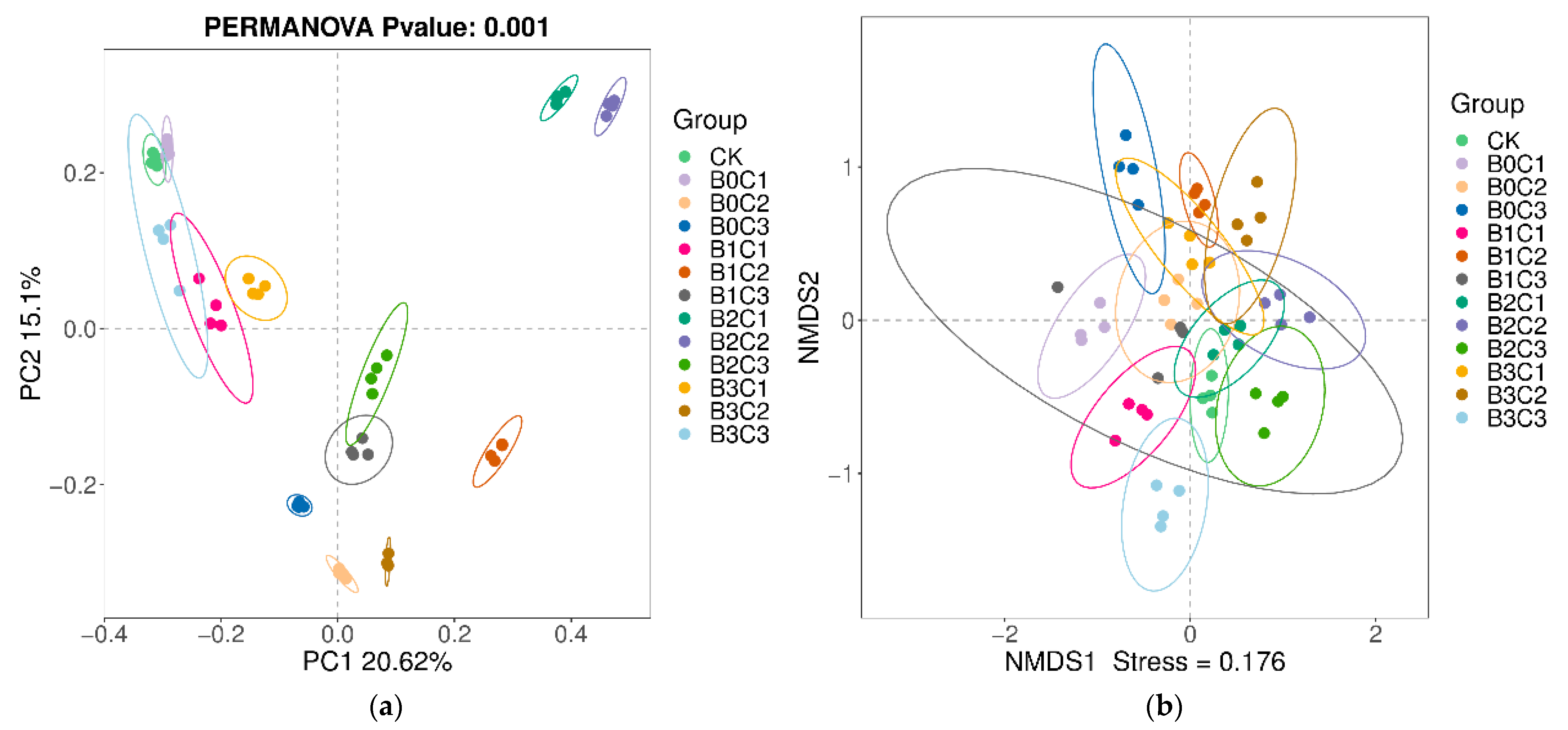
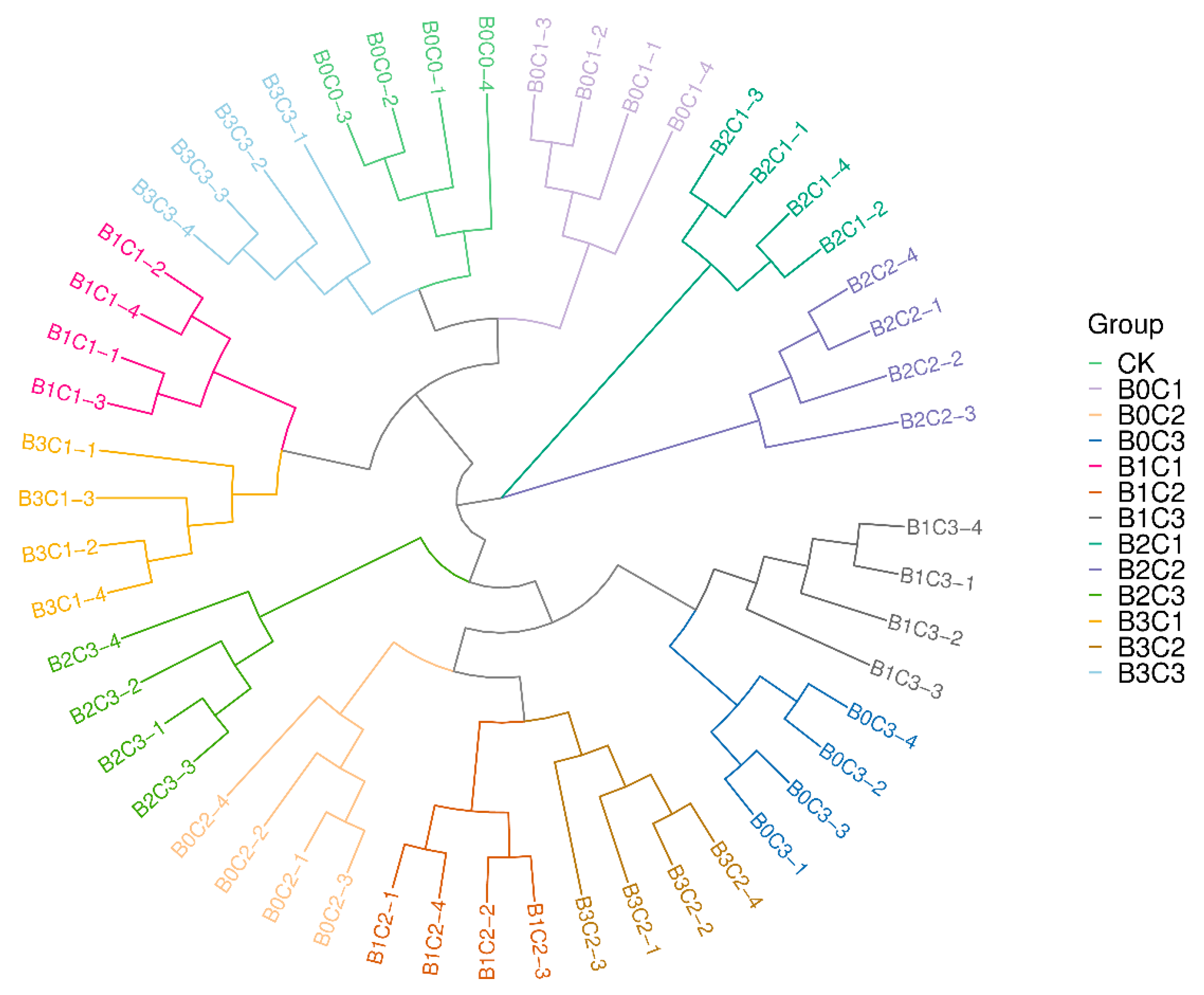
3.2.2. Beta Diversity Analysis
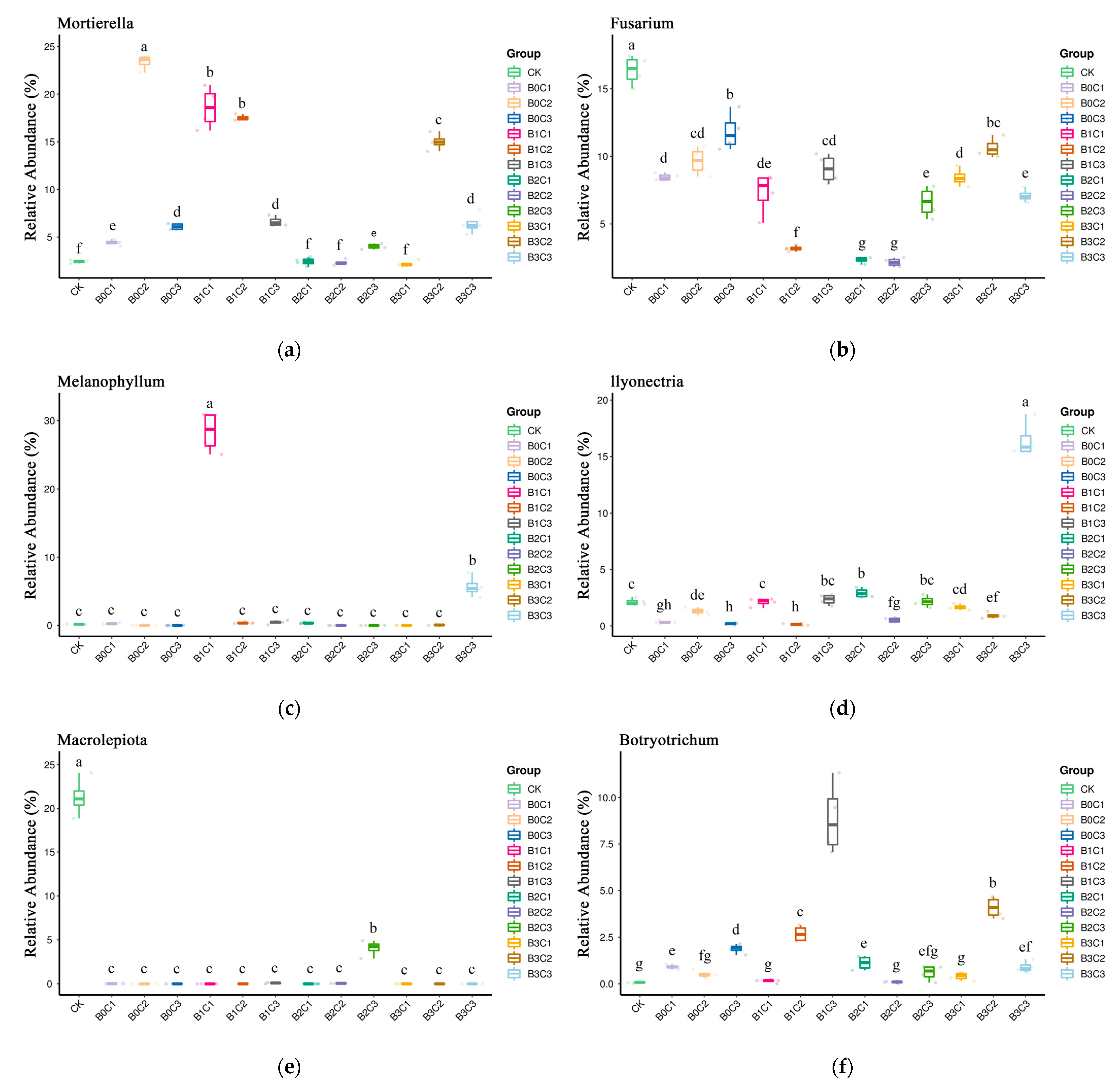
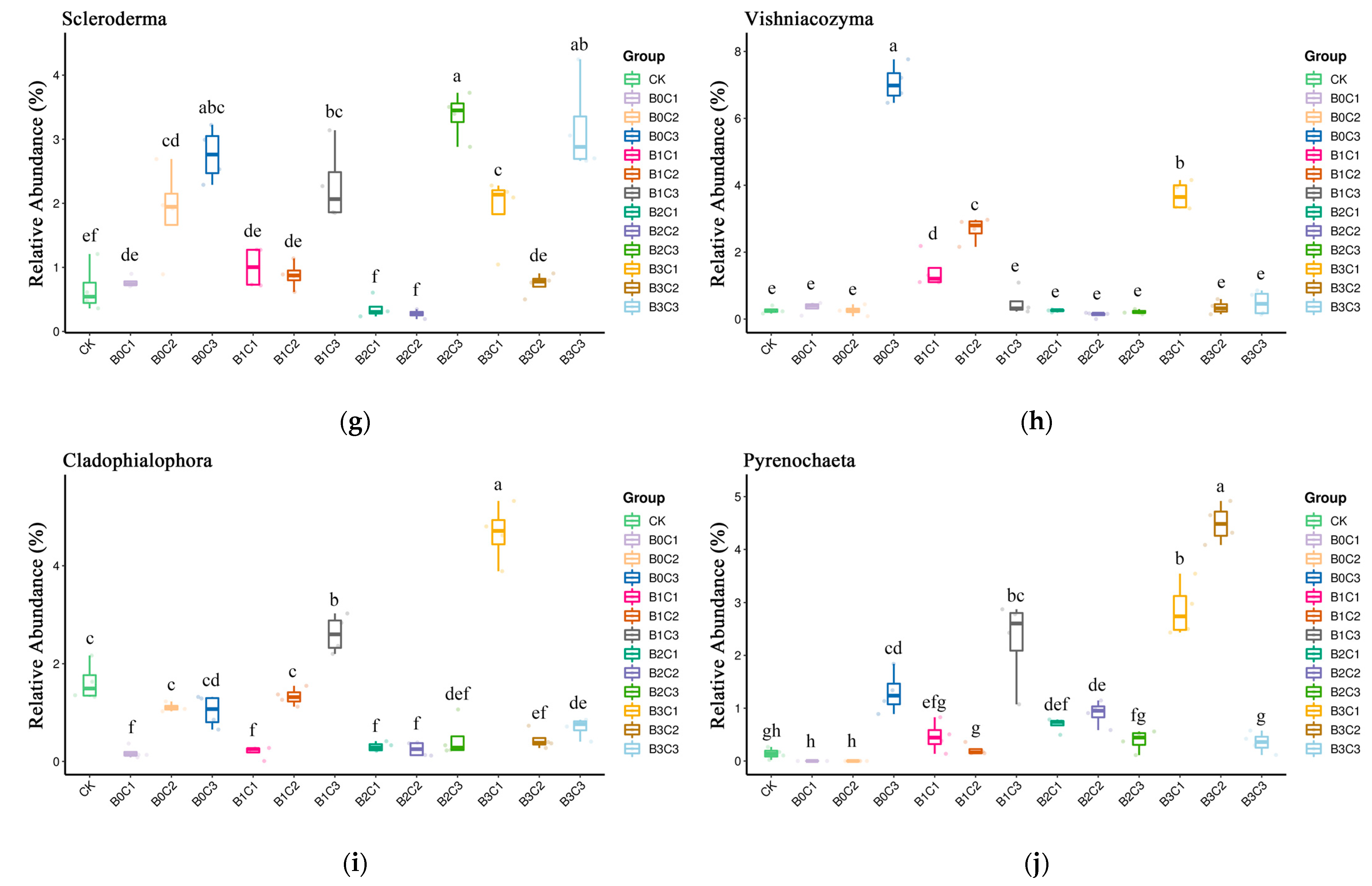
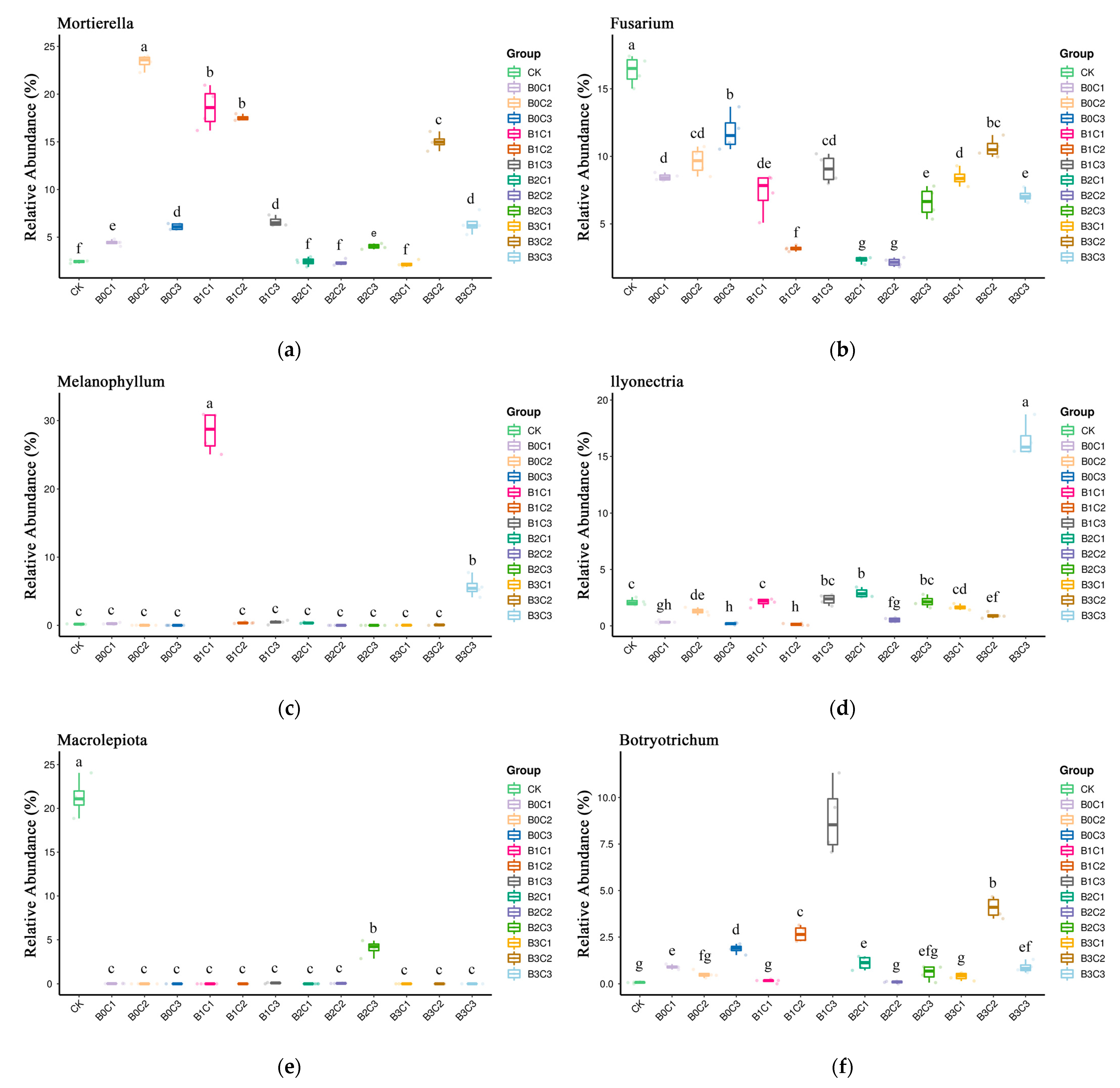
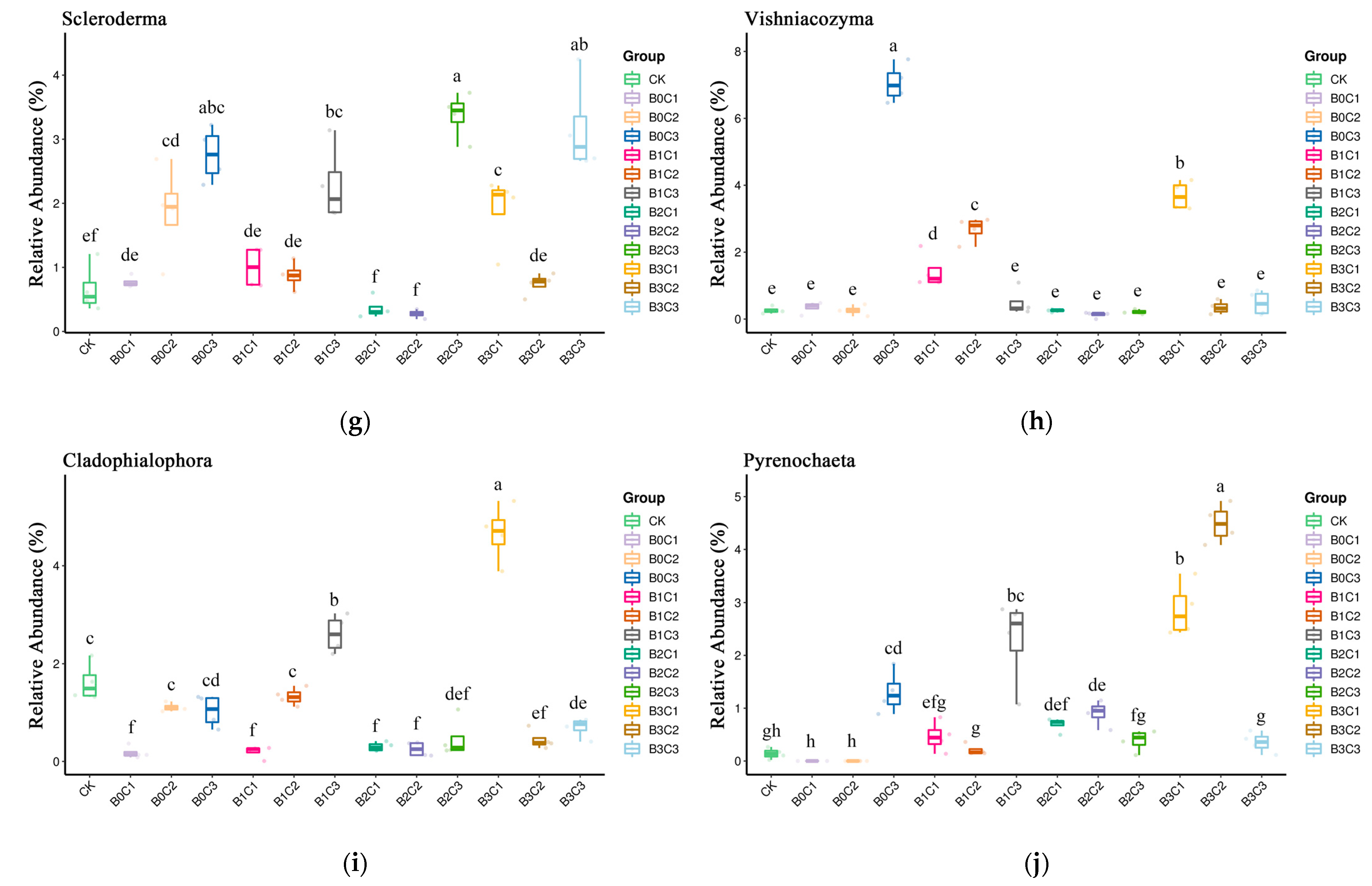
3.3. ANOVA Difference Statistics
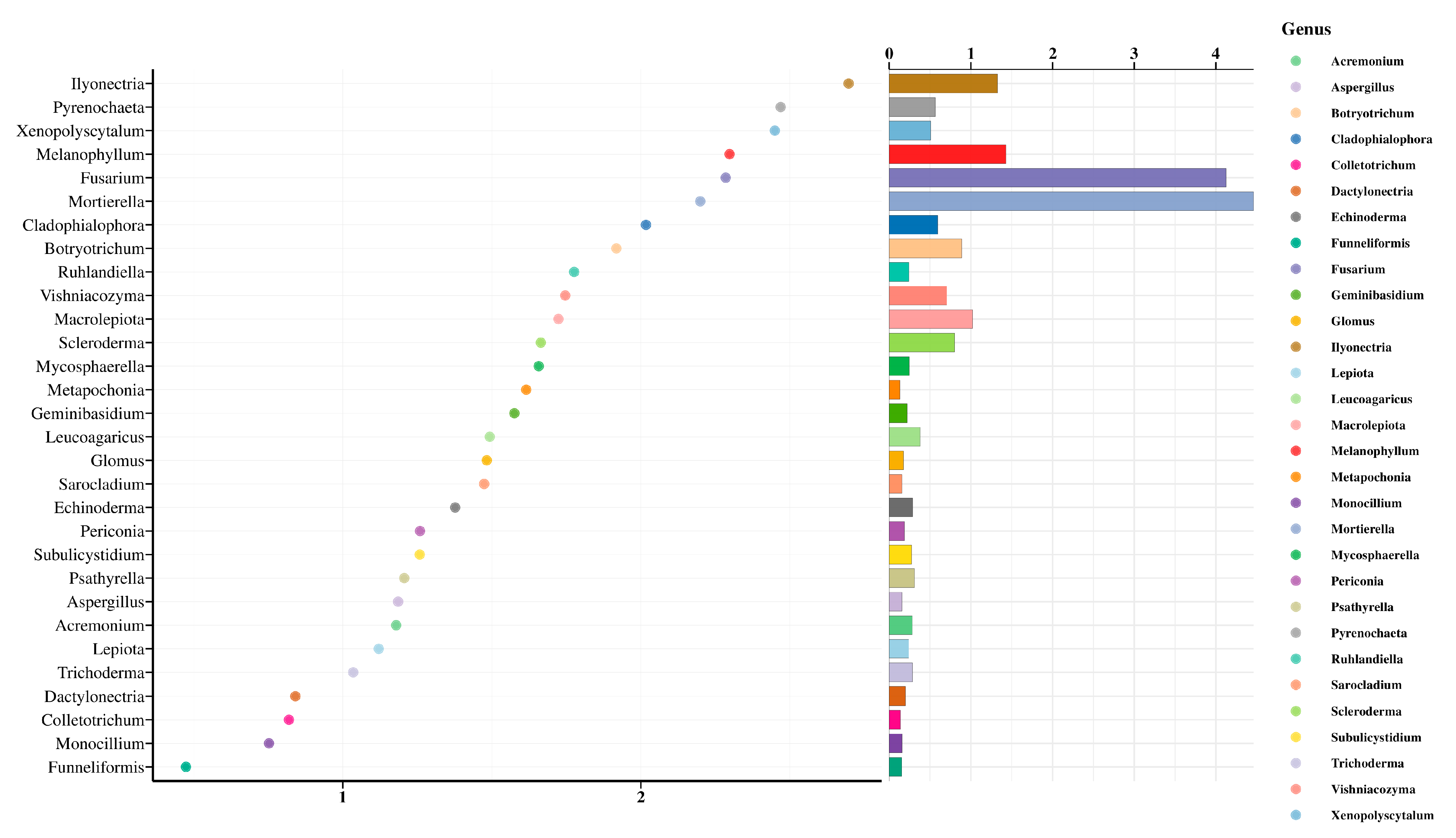
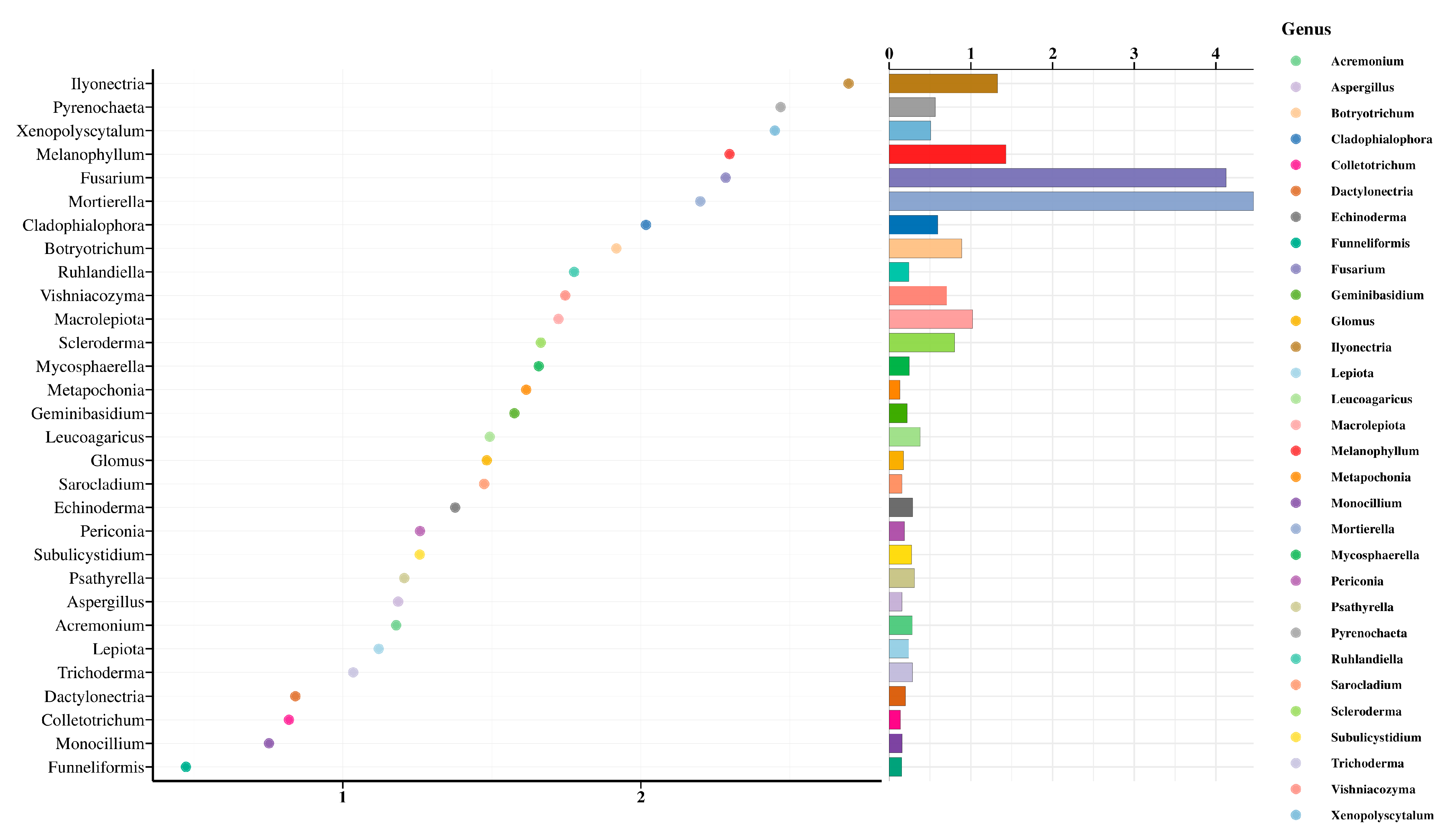
3.4. Random Forest Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- He, C.; Wang, H.; Jin, X.; Jin, B.; Su, S.; Duan, Y.; He, X. Identification of Alternaria species associated with black spot disease on Panax notoginseng in Yunnan and Guangxi. Acta Phytopathol. Sin. 2020, 50, 246–250. [Google Scholar]
- Zhang, L.; Gao, Y.; Dong, L.; Wei, G.; Yang, J.; Chen, J.; Sha, B. Major Diseases of Panax notoginseng And Their Control Strategies. Mod. Tradit. Chin. Med. Mater. Medica-World Sci. Technol. 2017, 19, 1635–1640. [Google Scholar]
- Wang, T.; Guo, R.X.; Zhou, G.H.; Zhou, X.D.; Kou, Z.Z.; Sui, F.; Li, C.; Tang, L.Y.; Wang, Z.J. Traditional uses, botany, phytochemistry, pharmacology and toxicology of Panax notoginseng (Burk.) FH Chen: A review. J. Ethnopharmacol. 2016, 188, 234–258. [Google Scholar] [CrossRef]
- Kim, J.H. Pharmacological and medical applications of Panax ginseng and ginsenosides: A review for use in cardiovascular diseases. J. Ginseng Res. 2018, 42, 264–269. [Google Scholar] [CrossRef]
- Lau, A.J.; Toh, D.F.; Chua, T.K.; Pang, Y.K.; Woo, S.O.; Koh, H.L. Antiplatelet and anticoagulant effects of Panax notoginseng: Comparison of raw and steamed Panax notoginseng with Panax ginseng and Panax quinquefolium. J. Ethnopharmacol. 2009, 125, 380–386. [Google Scholar] [CrossRef]
- Sun, X.; Li, L.; Long, G.; Zhang, G.; Meng, Z.; Yang, S.; Chen, J. The progress and prospect on consecutive monoculture problems of Panax notoginseng. Chin. J. Ecol. 2015, 34, 885–893. [Google Scholar]
- Ye, C.; Fang, H.Y.; Liu, H.J.; Yang, M.; Zhu, S.S. Current status of soil sickness research on Panax notoginseng in Yunnan, China. Allelopath. J. 2019, 47, 1–14. [Google Scholar] [CrossRef]
- Liu, H.J.; Yang, M.; Zhu, S.S. Strategies to solve the problem of Soil sickness of Panax notoginseng (Family: Araliaceae). Allelopath. J. 2019, 47, 37–56. [Google Scholar] [CrossRef]
- Pei, Y.; Yin, M.; Li, Q.H.; Zhang, Y.F.; Zhong, Y.; Chen, X.; Zhang, Y.P.; Huang, B.; Ren, Z. Diversity and Community Structure of Arbuscular Mycorrhizal Fungi (AMF) in the Rhizospheric Soil of Panax notoginseng in Different Ages. Eurasian Soil Sci. 2023, 56, 329–339. [Google Scholar] [CrossRef]
- Tuo, Y.; Wang, Z.Y.; Zheng, Y.; Shi, X.L.; Liu, X.N.; Ding, M.J.; Yang, Q.L. Effect of water and fertilizer regulation on the soil microbial biomass carbon and nitrogen, enzyme activity, and saponin content of Panax notoginseng. Agric. Water Manag. 2023, 278, 108145. [Google Scholar] [CrossRef]
- Tang, J.; Han, H.; Liu, B.; Yang, Q.; Liu, X.; Liu, Y. Effects of irrigation frequency and fertilization amount on active ingredient accumulation and morbidity of Panax notoginseng. Trans. Chin. Soc. Agric. Eng. 2020, 36, 55–63. [Google Scholar]
- Zhang, Y.; Liang, J.P.; Tang, Z.Y.; Yang, Q.L. Rain Shelter Cultivation Reduces Root Rot Incidence of Panax notoginseng by Altering Root Exudates and Bacterial Communities under Micro-Irrigation and Fertilization. Agron.-Basel 2023, 13, 1257. [Google Scholar] [CrossRef]
- Li, J.; Yang, Q.L.; Shi, Z.T.; Zang, Z.N.; Liu, X.G. Effects of deficit irrigation and organic fertilizer on yield, saponin and disease incidence in Panax notoginseng under shaded conditions. Agric. Water Manag. 2021, 256, 107056. [Google Scholar] [CrossRef]
- Tan, Y.; Cui, Y.; Li, H.; Kuang, A.; Li, X.; Wei, Y.; Ji, X. Rhizospheric soil and root endogenous fungal diversity and composition in response to continuous Panax notoginseng cropping practices. Microbiol. Res. 2017, 194, 10–19. [Google Scholar] [CrossRef]
- Liu, H.J.; Yang, X.Y.; Miao, Z.Q.; Li, S.D.; Chen, Y.H.; Liu, G.; Hao, L.Y.; Zhang, Z.L. Characteristics of soil microflora of Panax notoginseng in different continuous cropping years. Allelopath. J. 2018, 44, 145–157. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Boyce, A.N. Role of Plant Growth Promoting Rhizobacteria in Agricultural Sustainability-A Review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Ryu, C.-M.; Farag, M.A.; Kloepper, J.W.; Kloepper, J.W. Invisible Signals from the Underground: Bacterial Volatiles Elicit Plant GrowthPromotion and Induce Systemic Resistance. Plant Pathol. J. 2005, 21, 7–12. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Hashem, A.; Abd Allah, E.F. Bacillus: A Biological Tool for Crop Improvement through Bio-Molecular Changes in Adverse Environments. Front. Physiol. 2017, 8, 293128. [Google Scholar] [CrossRef] [PubMed]
- Han, L.F.; Sun, K.; Yang, Y.; Xia, X.H.; Li, F.B.; Yang, Z.F.; Xing, B.S. Biochar’s stability and effect on the content, composition and turnover of soil organic carbon. Geoderma 2020, 364, 114184. [Google Scholar] [CrossRef]
- Palansooriya, K.N.; Wong, J.T.F.; Hashimoto, Y.; Huang, L.B.; Rinklebe, J.; Chang, S.X.; Bolan, N.; Wang, H.L.; Ok, Y.S. Response of microbial communities to biochar-amended soils: A critical review. Biochar 2019, 1, 3–22. [Google Scholar] [CrossRef]
- Blanco-Canqui, H. Biochar and Soil Physical Properties. Soil Sci. Soc. Am. J. 2017, 81, 687–711. [Google Scholar] [CrossRef]
- Ameloot, N.; Graber, E.R.; Verheijen, F.G.A.; De Neve, S. Interactions between biochar stability and soil organisms: Review and research needs. Eur. J. Soil Sci. 2013, 64, 379–390. [Google Scholar] [CrossRef]
- Prendergast-Miller, M.T.; Duvall, M.; Sohi, S.P. Biochar-root interactions are mediated by biochar nutrient content and impacts on soil nutrient availability. Eur. J. Soil Sci. 2014, 65, 173–185. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Chandra, J.; Retuerto, M.; Sikaroodi, M.; Brown, R.E.; Jurevic, R.; Salata, R.A.; Lederman, M.M.; Gillevet, P.M.; Ghannoum, M.A. Oral Mycobiome Analysis of HIV-Infected Patients: Identification of Pichia as an Antagonist of Opportunistic Fungi. PLoS Pathog. 2014, 10, e1003996. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Li, H.; Shi, M.J.; Ren, K.Y.; Zhang, L.; Ye, W.D.; Zhang, W.T.; Cheng, Y.Y.; Xia, X.Q. Visual Omics: A web-based platform for omics data analysis and visualization with rich graph-tuning capabilities. Bioinformatics 2023, 39, btac777. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Polme, S.; Koljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef]
- Mi, C.Y.; Yang, R.H.; Rao, J.F.; Yang, S.Z.; Wei, F.G.; Li, O.; Hu, X.F. Unveiling of Dominant Fungal Pathogens Associated With Rusty Root Rot of Panax notoginseng Based on Multiple Methods. Plant Dis. 2017, 101, 2046–2052. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, Y.; Zhang, X.; Cao, N.; Li, B.; Liang, J.; Yang, Q. Effects of Combined Application of Biological Agent and Fertilizer on Fungal Community Structure in Rhizosphere Soil of Panax notoginseng. Agronomy 2023, 13, 2093. [Google Scholar] [CrossRef]
- Sun, H.; Guan, H.; Wang, X.; Wang, T.; Li, H.; Peng, W.; Liu, B.; Fan, F. Effects of Biochar on the Soil Properties and Fungal Community Structure under Continuous Cropping of Panax notoginseng. Biotechnol. Bull. 2023, 39, 221–231. [Google Scholar]
- Bao, L.M.; Liu, Y.Y.; Ding, Y.F.; Shang, J.J.; Wei, Y.L.; Tan, Y.; Zi, F.T. Interactions Between Phenolic Acids and Microorganisms in Rhizospheric Soil From Continuous Cropping of Panax notoginseng. Front. Microbiol. 2022, 13, 791603. [Google Scholar] [CrossRef] [PubMed]
- Pandey, D.; Daverey, A.; Arunachalam, K. Biochar: Production, properties and emerging role as a support for enzyme immobilization. J. Clean. Prod. 2020, 255, 120267. [Google Scholar] [CrossRef]
- Wu, L.P.; Wei, C.B.; Zhang, S.R.; Wang, Y.D.; Kuzyakov, Y.; Ding, X.D. MgO-modified biochar increases phosphate retention and rice yields in saline-alkaline soil. J. Clean. Prod. 2019, 235, 901–909. [Google Scholar] [CrossRef]
- Topoliantz, S.; Ponge, J.F.; Ballof, S. Manioc peel and charcoal: A potential organic amendment for sustainable soil fertility in the tropics. Biol. Fertil. Soils 2005, 41, 15–21. [Google Scholar] [CrossRef]
- Laird, D.; Fleming, P.; Wang, B.Q.; Horton, R.; Karlen, D. Biochar impact on nutrient leaching from a Midwestern agricultural soil. Geoderma 2010, 158, 436–442. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Ghassemi-Golezani, K. Improving electrochemical characteristics of plant roots by biochar is an efficient mechanism in increasing cations uptake by plants. Chemosphere 2023, 313, 137365. [Google Scholar] [CrossRef]
- Gaskin, J.W.; Steiner, C.; Harris, K.; Das, K.C.; Bibens, B. Effect of low-temperature pyrolysis conditions on biochar for agricultural use. Trans. ASABE 2008, 51, 2061–2069. [Google Scholar] [CrossRef]
- Li, Y.C.; Li, Y.F.; Chang, S.X.; Yang, Y.F.; Fu, S.L.; Jiang, P.K.; Luo, Y.; Yang, M.; Chen, Z.H.; Hu, S.D.; et al. Biochar reduces soil heterotrophic respiration in a subtropical plantation through increasing soil organic carbon recalcitrancy and decreasing carbon degrading microbial activity. Soil Biol. Biochem. 2018, 122, 173–185. [Google Scholar] [CrossRef]
- Palansooriya, K.N.; Ok, Y.S.; Awad, Y.M.; Lee, S.S.; Sung, J.K.; Koutsospyros, A.; Moon, D.H. Impacts of biochar application on upland agriculture: A review. J. Environ. Manag. 2019, 234, 52–64. [Google Scholar] [CrossRef]
- Muhammad, N.; Dai, Z.M.; Xiao, K.C.; Meng, J.; Brookes, P.C.; Liu, X.M.; Wang, H.Z.; Wu, J.J.; Xu, J.M. Changes in microbial community structure due to biochars generated from different feedstocks and their relationships with soil chemical properties. Geoderma 2014, 226, 270–278. [Google Scholar] [CrossRef]
- Choppala, G.; Bolan, N.; Kunhikrishnan, A.; Bush, R. Differential effect of biochar upon reduction-induced mobility and bioavailability of arsenate and chromate. Chemosphere 2016, 144, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.M.; Yang, Z.F.; He, P.F.; Munir, S.; Wu, Y.X.; Ho, H.H.; He, Y.Q. Deciphering the bacterial and fungal communities in clubroot-affected cabbage rhizosphere treated with Bacillus Subtilis XF-1. Agric. Ecosyst. Environ. 2018, 256, 12–22. [Google Scholar] [CrossRef]
- Dong, L.; Xu, J.; Feng, G.; Li, X.; Chen, S. Soil bacterial and fungal community dynamics in relation to Panax notoginseng death rate in a continuous cropping system. Sci. Rep. 2016, 6, 31802. [Google Scholar] [CrossRef] [PubMed]
- Yuejuan, L.; Zhongshun, M.A.O.; Shusheng, Z.H.U.; Zhongjian, C.; Fugang, W.E.I.; Zhaobo, Y.I.N.; Youyong, Z.H.U.; Xiahong, H.E. The pathogen of Panax notoginseng root rust spot. Mycosystema 2015, 34, 177–185. [Google Scholar]
- Gebreyohannes, G.; Nyerere, A.; Bii, C.; Sbhatu, D.B. Challenges of intervention, treatment, and antibiotic resistance of biofilm-forming microorganisms. Heliyon 2019, 5, e02192. [Google Scholar] [CrossRef]
- Guzmán-Soto, I.; McTiernan, C.; Gonzalez-Gomez, M.; Ross, A.; Gupta, K.; Suuronen, E.J.; Mah, T.F.; Griffith, M.; Alarcon, E.I. Mimicking biofilm formation and development: Recent progress in in vitro and in vivo biofilm models. iScience 2021, 24, 102443. [Google Scholar] [CrossRef]
- Sauer, K.; Stoodley, P.; Goeres, D.M.; Hall-Stoodley, L.; Burmolle, M.; Stewart, P.S.; Bjarnsholt, T. The biofilm life cycle: Expanding the conceptual model of biofilm formation. Nat. Rev. Microbiol. 2022, 20, 608–620. [Google Scholar] [CrossRef]
- Chen, W.M.; Wu, Z.S.; Liu, C.H.; Zhang, Z.Y.; Liu, X.C. Biochar combined with Bacillus subtilis SL-44 as an eco-friendly strategy to improve soil fertility, reduce Fusarium wilt, and promote radish growth. Ecotox. Environ. Safe. 2023, 251, 114509. [Google Scholar] [CrossRef]
- Tao, S.Y.; Wu, Z.S.; Wei, M.M.; Liu, X.C.; He, Y.H.; Ye, B.C. Bacillus subtilis SL-13 biochar formulation promotes pepper plant growth and soil improvement. Can. J. Microbiol. 2019, 65, 333–342. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Applied Amount of Biochar (kg·hm−2) | Applied Amount of Biological Agents (kg·hm−2) |
|---|---|---|
| CK | 0 | 0 |
| B0C1 | 0 | 10 |
| B0C2 | 0 | 15 |
| B0C3 | 0 | 25 |
| B1C1 | 80 | 10 |
| B1C2 | 80 | 15 |
| B1C3 | 80 | 25 |
| B2C1 | 110 | 10 |
| B2C2 | 110 | 15 |
| B2C3 | 110 | 25 |
| B3C1 | 140 | 10 |
| B3C2 | 140 | 15 |
| B3C3 | 140 | 25 |
| Treatment | ASVs | Chao1 | Shannon | Simpson | ACE | Good_Coverage |
|---|---|---|---|---|---|---|
| CK | 189.00 ± 9.87c | 189.06 ± 9.85c | 4.08 ± 0.06f | 0.85 ± 0.01h | 188.73 ± 10.06c | 0.99 |
| B0C1 | 199.73 ± 7.01c | 199.95 ± 7.13c | 3.81 ± 0.08g | 0.79 ± 0.01i | 199.79 ± 6.69c | 0.99 |
| B0C2 | 180.00 ± 17.07c | 180.03 ± 17.10c | 5.58 ± 0.12c | 0.94 ± 0.01e | 179.82 ± 16.88c | 0.99 |
| B0C3 | 316.98 ± 10.64b | 316.98 ± 10.64b | 6.16 ± 0.10b | 0.96 ± 0.00b | 316.60 ± 10.04c | 0.99 |
| B1C1 | 166.18 ± 26.29c | 166.18 ± 26.29c | 4.68 ± 0.11d | 0.92 ± 0.01f | 166.55 ± 26.53c | 0.99 |
| B1C2 | 370.00 ± 4.06a | 370.19 ± 4.02a | 6.00 ± 0.02b | 0.95 ± 0.00d | 370.14 ± 3.71a | 0.99 |
| B1C3 | 259.48 ± 54.81bc | 259.50 ± 54.80bc | 6.53 ± 0.41ab | 0.98 ± 0.00a | 259.42 ± 54.78bc | 0.99 |
| B2C1 | 242.75 ± 3.90c | 242.79 ± 3.92c | 4.37 ± 0.08e | 0.86 ± 0.01h | 242.33 ± 4.71c | 0.99 |
| B2C2 | 215.00 ± 29.66c | 215.01 ± 29.66c | 3.96 ± 0.19fg | 0.77 ± 0.02i | 214.95 ± 29.56c | 0.99 |
| B2C3 | 229.50 ± 6.02c | 229.51 ± 6.00c | 5.93 ± 0.04b | 0.96 ± 0.00b | 229.31 ± 6.31c | 0.99 |
| B3C1 | 324.00 ± 27.24b | 324.00 ± 27.24b | 6.62 ± 0.09a | 0.97 ± 0.00a | 323.91 ± 27.39b | 0.99 |
| B3C2 | 330.25 ± 19.90b | 330.27 ± 19.91b | 6.08 ± 0.07b | 0.96 ± 0.00c | 330.69 ± 20.17b | 0.99 |
| B3C3 | 170.50 ± 22.60c | 170.50 ± 22.60c | 4.67 ± 0.13a | 0.90 ± 0.01g | 170.91 ± 22.88c | 0.99 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Liu, Y.; Li, S.; Yang, Q. The Combination of Biochar and Bacillus subtilis Biological Agent Reduced the Relative Abundance of Pathogenic Bacteria in the Rhizosphere Soil of Panax notoginseng. Microorganisms 2024, 12, 783. https://doi.org/10.3390/microorganisms12040783
Zhou Y, Liu Y, Li S, Yang Q. The Combination of Biochar and Bacillus subtilis Biological Agent Reduced the Relative Abundance of Pathogenic Bacteria in the Rhizosphere Soil of Panax notoginseng. Microorganisms. 2024; 12(4):783. https://doi.org/10.3390/microorganisms12040783
Chicago/Turabian StyleZhou, Yingjie, Yanwei Liu, Siwen Li, and Qiliang Yang. 2024. "The Combination of Biochar and Bacillus subtilis Biological Agent Reduced the Relative Abundance of Pathogenic Bacteria in the Rhizosphere Soil of Panax notoginseng" Microorganisms 12, no. 4: 783. https://doi.org/10.3390/microorganisms12040783
APA StyleZhou, Y., Liu, Y., Li, S., & Yang, Q. (2024). The Combination of Biochar and Bacillus subtilis Biological Agent Reduced the Relative Abundance of Pathogenic Bacteria in the Rhizosphere Soil of Panax notoginseng. Microorganisms, 12(4), 783. https://doi.org/10.3390/microorganisms12040783