
Unveiling a Listeria monocytogenes Outbreak in a Rabbit Farm: Clinical Manifestation, Antimicrobial Resistance, Genomic Insights and Environmental Investigation

 , , ,
, , ,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
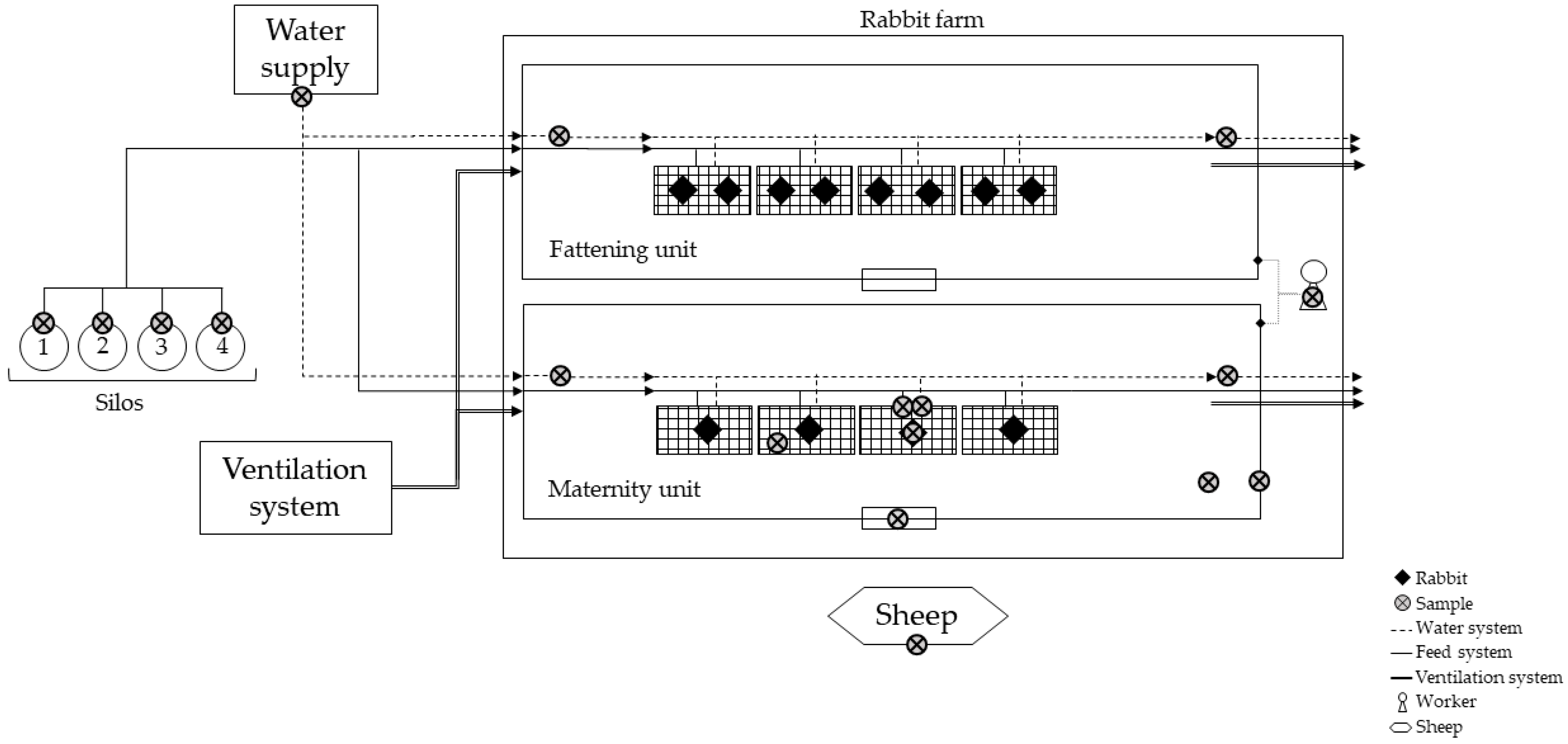
2.1. Rabbit Farm Description
2.2. Sample Collection
2.2.1. Does Samples
2.2.2. Environmental Samples
2.2.3. Nearest Livestock Population
2.3. L. monocytogenes Detection
2.4. PCR Identification of L. monocytogenes
2.5. Antimicrobial Resistance Testing
2.6. WGS Characterization
2.6.1. Genomic DNA Extraction and Whole-Genome Sequencing (WGS)
2.6.2. Bioinformatics Analysis
3. Results
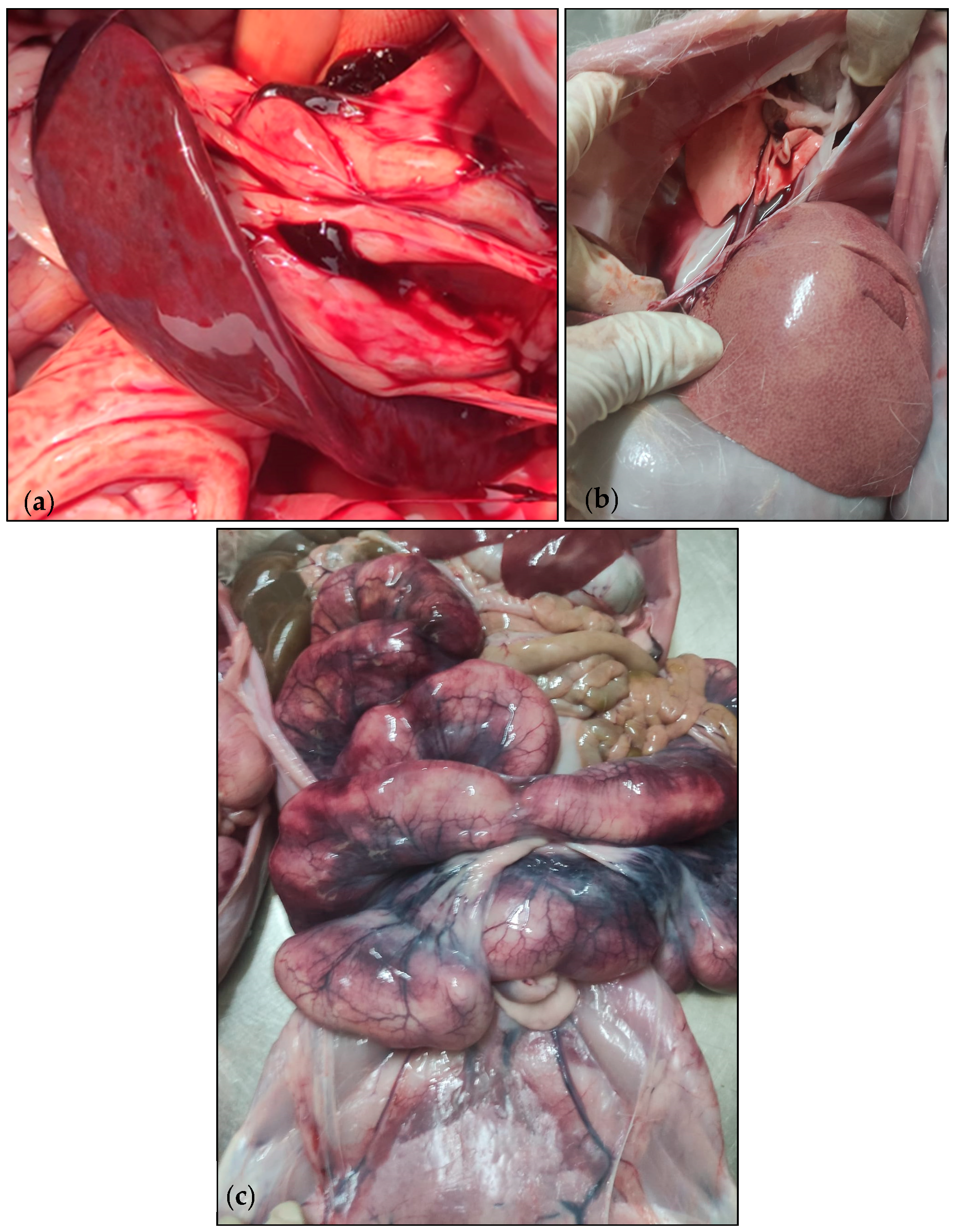
3.1. Case Description
3.2. Detection of L. monocytogenes
3.3. Phenotypic and Genotypic Characterization of L. monocytogenes Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amajoud, N.; Leclercq, A.; Soriano, J.M.; Bracq-Dieye, H.; El Maadoudi, M.; Senhaji, N.S.; Kounnoun, A.; Moura, A.; Lecuit, M.; Abrini, J. Prevalence of Listeria spp. and Characterization of Listeria monocytogenes Isolated from Food Products in Tetouan, Morocco. Food Control 2018, 84, 436–441. [Google Scholar] [CrossRef]
- Dhama, K.; Karthik, K.; Tiwari, R.; Shabbir, M.Z.; Barbuddhe, S.; Malik, S.V.S.; Singh, R.K. Listeriosis in Animals, Its Public Health Significance (Food-Borne Zoonosis) and Advances in Diagnosis and Control: A Comprehensive Review. Vet. Q. 2015, 35, 211–235. [Google Scholar] [CrossRef] [PubMed]
- OIE. Chapter 3.9.6. Listeria monocytogenes. In OIE Terrestrial Manual; OIE: Paris, France, 2018; pp. 1705–1722. [Google Scholar]
- Abd El-Ghany, W.A. Listeriosis in Rabbits (Oryctolagus cuniculus): A Significant Bacterial with an Emerging Zoonosis. J. Adv. Vet. Res. 2022, 13, 150–156. [Google Scholar]
- Colagiorgi, A.; Bruini, I.; Di Ciccio, P.A.; Zanardi, E.; Ghidini, S.; Ianieri, A. Listeria monocytogenes Biofilms in the Wonderland of Food Industry. Pathogens 2017, 6, 41. [Google Scholar] [CrossRef] [PubMed]
- McLauchlin, J. Listeriosis. In Zoonoses: Biology, Clinical Practice, and Public Health Control; Oxford University Press: Oxford, UK, 2011; Volume 1, pp. 117–127. [Google Scholar]
- Wartha, S.; Huber, S.; Kraemer, I.; Alter, T.; Messelhäußer, U. Presence of Listeria at Primary Production and Processing of Food of Non-Animal Origin (FNAO) in Bavaria, Germany. J. Food Prot. 2023, 86, 100015. [Google Scholar] [CrossRef] [PubMed]
- EFSA. The European Union One Health 2022 Zoonoses Report. EFSA J. 2023, 21, e8442. [Google Scholar] [CrossRef]
- Mena, C.; Almeida, G.; Carneiro, L.; Teixeira, P.; Hogg, T.; Gibbs, P.A. Incidence of Listeria monocytogenes in Different Food Products Commercialized in Portugal. Food Microbiol. 2004, 21, 213–216. [Google Scholar] [CrossRef]
- Magalhães, R.; Almeida, G.; Ferreira, V.; Santos, I.; Silva, J.; Mendes, M.M.; Pita, J.; Mariano, G.; Mâncio, I.; Sousa, M.M.; et al. Cheese-Related Listeriosis Outbreak, Portugal, March 2009 to February 2012. Eurosurveillance 2015, 20, 21104. [Google Scholar] [CrossRef]
- Praça, J.; Furtado, R.; Coelho, A.; Correia, C.B.; Borges, V.; Gomes, J.P.; Pista, A.; Batista, R. Listeria monocytogenes, Escherichia coli and Coagulase Positive Staphylococci in Cured Raw Milk Cheese from Alentejo Region, Portugal. Microorganisms 2023, 11, 322. [Google Scholar] [CrossRef]
- Vilar, M.J.; Yus, E.; Sanjuán, M.L.; Diéguez, F.J.; Rodríguez-Otero, J.L. Prevalence of and Risk Factors for Listeria Species on Dairy Farms. J. Dairy. Sci. 2007, 90, 5083–5088. [Google Scholar] [CrossRef]
- WOAH. Chapter 1.3. Diseases, Infections and Infestations by WOAH. In Terrestrial Animal Health Code; World Organisation for Animal Health: Paris, France, 2023; pp. 1–3. [Google Scholar]
- Ibrahim, G.A.; Ibrahim, H.N. Bacteriological, Clinic-Pathological Studies of Listeria monocytogenes in Rabbits and Detection of Some Virulence Genes by Polymerase Chain Reaction (PCR). Environ. Health 2016, 2, 250–264. [Google Scholar]
- Gelbíčová, T.; Florianová, M.; Tomáštíková, Z.; Pospíšilová, L.; Koláčková, I.; Karpíšková, R. Prediction of Persistence of Listeria monocytogenes ST451 in a Rabbit Meat Processing Plant in the Czech Republic. J. Food Prot. 2019, 82, 1350–1356. [Google Scholar] [CrossRef]
- Pasquali, F.; Palma, F.; Guillier, L.; Lucchi, A.; De Cesare, A.; Manfreda, G. Listeria monocytogenes Sequence Types 121 and 14 Repeatedly Isolated within One Year of Sampling in a Rabbit Meat Processing Plant: Persistence and Ecophysiology. Front. Microbiol. 2018, 9, 340797. [Google Scholar] [CrossRef] [PubMed]
- De Cesare, A.; Parisi, A.; Mioni, R.; Comin, D.; Lucchi, A.; Manfreda, G. Listeria monocytogenes Circulating in Rabbit Meat Products and Slaughterhouses in Italy: Prevalence Data and Comparison among Typing Results. Foodborne Pathog. Dis. 2017, 14, 167–176. [Google Scholar] [CrossRef]
- Nash, M.L.; Hungerford, L.L.; Nash, T.G.; Zinn, G.M. Epidemiology and Economics of Clinical Listeriosis in a Sheep Flock. Prev. Vet. Med. 1995, 24, 147–156. [Google Scholar] [CrossRef]
- Department of Primary Industries and Regional Development’s Agriculture and Food Division Infertility and Abortion in Ewes. Available online: https://www.agric.wa.gov.au/livestock-biosecurity/infertility-and-abortion-ewes (accessed on 29 January 2024).
- EU. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes. Off. J. Eur. Union 2010, 276, 33–79. [Google Scholar]
- ISO 11290-1; Microbiology of the Food Chain—Horizontal Method for the Detection and Enumeration of Listeria monocytogenes and of Listeria spp.—Part 1: Detection Method. ISO: Geneva, Switzerland, 2017.
- Paziak-Domańska, B.; Bogusławska, E.; Więckowska-Szakiel, M.; Kotłowski, R.; Różalska, B.; Chmiela, M.; Kur, J.; Dąbrowski, W.; Rudnicka, W. Evaluation of the API Test, Phosphatidylinositol-Specific Phospholipase C Activity and PCR Method in Identification of Listeria monocytogenes in Meat Foods. FEMS Microbiol. Lett. 1999, 171, 209–214. [Google Scholar] [CrossRef]
- Soni, D.K.; Dubey, S.K. Phylogenetic Analysis of the Listeria monocytogenes Based on Sequencing of 16S RRNA and HlyA Genes. Mol. Biol. Rep. 2014, 41, 8219–8229. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing Antimicrobial Susceptibility Testing EUCAST Disk Diffusion Method. Available online: https://www.eucast.org/ast_of_bacteria/disk_diffusion_methodology (accessed on 16 January 2024).
- European Committee on Antimicrobial Susceptibility Testing Breakpoint Tables for Interpretation of MICs and Zone Diameters. Available online: https://www.eucast.org/clinical_breakpoints (accessed on 16 January 2024).
- Llarena, A.; Ribeiro-Gonçalves, B.F.; Nuno Silva, D.; Halkilahti, J.; Machado, M.P.; Da Silva, M.S.; Jaakkonen, A.; Isidro, J.; Hämäläinen, C.; Joenperä, J.; et al. INNUENDO: A Cross-sectoral Platform for the Integration of Genomics in the Surveillance of Food-borne Pathogens. EFSA Support. Publ. 2018, 15, 1498E. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes De Novo Assembler. Curr. Protoc. Bioinform. 2020, 70, e102. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B. Aligning Short Sequencing Reads with Bowtie. Curr. Protoc. Bioinform. 2010, 32, 11.7.1–11.7.14. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast Metagenomic Sequence Classification Using Exact Alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef]
- Silva, M.; Machado, M.P.; Silva, D.N.; Rossi, M.; Moran-Gilad, J.; Santos, S.; Ramirez, M.; Carriço, J.A. ChewBBACA: A Complete Suite for Gene-by-Gene Schema Creation and Strain Identification. Microb. Genom. 2018, 4, e000166. [Google Scholar] [CrossRef] [PubMed]
- Moura, A.; Criscuolo, A.; Pouseele, H.; Maury, M.M.; Leclercq, A.; Tarr, C.; Björkman, J.T.; Dallman, T.; Reimer, A.; Enouf, V.; et al. Whole Genome-Based Population Biology and Epidemiological Surveillance of Listeria monocytogenes. Nat. Microbiol. 2016, 2, 16185. [Google Scholar] [CrossRef] [PubMed]
- Mamede, R.; Vila-Cerqueira, P.; Silva, M.; Carriço, J.A.; Ramirez, M. Chewie Nomenclature Server (Chewie-NS): A Deployable Nomenclature Server for Easy Sharing of Core and Whole Genome MLST Schemas. Nucleic Acids Res. 2021, 49, D660–D666. [Google Scholar] [CrossRef]
- Mixão, V.; Pinto, M.; Sobral, D.; Di Pasquale, A.; Gomes, J.P.; Borges, V. ReporTree: A Surveillance-Oriented Tool to Strengthen the Linkage between Pathogen Genetic Clusters and Epidemiological Data. Genome Med. 2023, 15, 43. [Google Scholar] [CrossRef]
- Zhou, Z.; Alikhan, N.-F.; Sergeant, M.J.; Luhmann, N.; Vaz, C.; Francisco, A.P.; Carriço, J.A.; Achtman, M. GrapeTree: Visualization of Core Genomic Relationships among 100,000 Bacterial Pathogens. Genome Res. 2018, 28, 1395–1404. [Google Scholar] [CrossRef]
- Van Walle, I.; Björkman, J.T.; Cormican, M.; Dallman, T.; Mossong, J.; Moura, A.; Pietzka, A.; Ruppitsch, W.; Takkinen, J. Retrospective Validation of Whole Genome Sequencing-Enhanced Surveillance of Listeriosis in Europe, 2010 to 2015. Eurosurveillance 2018, 23, 1700798. [Google Scholar] [CrossRef]
- Cruz, H.; Pinheiro, M.; Borges, V. ReporType: A Flexible Bioinformatics Tool for Targeted Loci Screening and Typing of Infectious Agents. Int. J. Mol. Sci. 2024, 25, 3172. [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E.; et al. AMRFinderPlus and the Reference Gene Catalog Facilitate Examination of the Genomic Links among Antimicrobial Resistance, Stress Response, and Virulence. Sci. Rep. 2021, 11, 12728. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2019, 48, D517–D525. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.-M. ARG-ANNOT, a New Bioinformatic Tool to Discover Antibiotic Resistance Genes in Bacterial Genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Jolley, K.A.; Maiden, M.C. BIGSdb: Scalable Analysis of Bacterial Genome Variation at the Population Level. BMC Bioinform. 2010, 11, 595. [Google Scholar] [CrossRef] [PubMed]
- Naemi, A.-O.; Dey, H.; Kiran, N.; Sandvik, S.T.; Slettemeås, J.S.; Nesse, L.L.; Simm, R. NarAB Is an ABC-Type Transporter That Confers Resistance to the Polyether Ionophores Narasin, Salinomycin, and Maduramicin, but Not Monensin. Front. Microbiol. 2020, 11, 506049. [Google Scholar] [CrossRef] [PubMed]
- Palma, F.; Pasquali, F.; Lucchi, A.; De Cesare, A.; Manfreda, G. Whole Genome Sequencing for Typing and Characterisation of Listeria monocytogenes Isolated in a Rabbit Meat Processing Plant. Ital. J. Food Saf. 2017, 6, 6879. [Google Scholar] [CrossRef] [PubMed]
- Schoder, D.; Guldimann, C.; Märtlbauer, E. Asymptomatic Carriage of Listeria monocytogenes by Animals and Humans and Its Impact on the Food Chain. Foods 2022, 11, 3472. [Google Scholar] [CrossRef]
- Matle, I.; Mbatha, K.R.; Madoroba, E. A Review of Listeria monocytogenes from Meat and Meat Products: Epidemiology, Virulence Factors, Antimicrobial Resistance and Diagnosis. Onderstepoort J. Vet. Res. 2020, 87, 1–20. [Google Scholar] [CrossRef]
- Suyemoto, M.M.; Spears, P.A.; Hamrick, T.S.; Barnes, J.A.; Havell, E.A.; Orndorff, P.E. Factors Associated with the Acquisition and Severity of Gestational Listeriosis. PLoS ONE 2010, 5, e13000. [Google Scholar] [CrossRef] [PubMed]
- Kylie, J.; Brash, M.; Whiteman, A.; Tapscott, B.; Slavic, D.; Weese, J.S.; Turner, P. V Biosecurity Practices and Causes of Enteritis on Ontario Meat Rabbit Farms. Can. Vet. J. 2017, 58, 571–578. [Google Scholar]
- Huss, A.; Cochrane, R.; Jones, C.; Atungulu, G.G. Physical and Chemical Methods for the Reduction of Biological Hazards in Animal Feeds. In Food and Feed Safety Systems and Analysis; Elsevier: Amsterdam, The Netherlands, 2018; pp. 83–95. [Google Scholar]
- Olson, E.G.; Grenda, T.; Ghosh, A.; Ricke, S.C. Microbial Pathogen Contamination of Animal Feed. In Present Knowledge in Food Safety; Elsevier: Amsterdam, The Netherlands, 2023; pp. 378–393. [Google Scholar]
- Munoz, L.R.; Pacheco, W.J.; Hauck, R.; Macklin, K.S. Evaluation of Commercially Manufactured Animal Feeds to Determine Presence of Salmonella, Escherichia coli, and Clostridium perfringens. J. Appl. Poult. Res. 2021, 30, 100142. [Google Scholar] [CrossRef]
- Moura, A.; Leclercq, A.; Vales, G.; Tessaud-Rita, N.; Bracq-Dieye, H.; Thouvenot, P.; Madec, Y.; Carlier, C.; Lecuit, M. Phenotypic and Genotypic Antimicrobial Resistance of Listeria monocytogenes: An Observational Study in France. Lancet Reg. Health 2024, 37, 100843. [Google Scholar] [CrossRef] [PubMed]
- Kolář, M.; Urbánek, K.; Látal, T. Antibiotic Selective Pressure and Development of Bacterial Resistance. Int. J. Antimicrob. Agents 2001, 17, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Luque-Sastre, L.; Arroyo, C.; Fox, E.M.; McMahon, B.J.; Bai, L.; Li, F.; Fanning, S. Antimicrobial Resistance in Listeria Species. Microbiol. Spectr. 2018, 6, 10-1128. [Google Scholar] [CrossRef]
- World Health Organization. Listeria monocytogenes in Ready-to-Eat (RTE) Foods: Attribution, Characterization and Monitoring—Meeting Report; Microbiological Risk Assessment Series No. 38; Food and Agriculture Organization of the United Nations: Rome, Italy, 2022. [Google Scholar]
- Jennison, A.V.; Masson, J.J.; Fang, N.-X.; Graham, R.M.; Bradbury, M.I.; Fegan, N.; Gobius, K.S.; Graham, T.M.; Guglielmino, C.J.; Brown, J.L.; et al. Analysis of the Listeria monocytogenes Population Structure among Isolates from 1931 to 2015 in Australia. Front. Microbiol. 2017, 8, 603. [Google Scholar] [CrossRef] [PubMed]
- Finazzi, G.; Filipello, V.; Gori, M.; Scaltriti, E.; Bracchi, C.; Menozzi, I.; Tanzi, E.; Bolzoni, L. A Listeria monocytogenes ST325 Clone Is Widespread in the Lombardy Region Dairy Processing Plants. Eur. J. Public Health 2020, 30, v614. [Google Scholar] [CrossRef]
- Haase, J.K.; Didelot, X.; Lecuit, M.; Korkeala, H.; Achtman, M. The Ubiquitous Nature of Listeria monocytogenes Clones: A Large-scale Multilocus Sequence Typing Study. Environ. Microbiol. 2014, 16, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, B.; Anand, S. Environmental Persistence of Listeria monocytogenes and Its Implications in Dairy Processing Plants. Compr. Rev. Food Sci. Food Saf. 2023, 22, 4573–4599. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sample ID | Type of Sample | Sample Type |
|---|---|---|
| 3572 | Water | Well water |
| 3573 | Water | Water from the initial dispenser line |
| 3574 | Water | Water from the final dispenser line |
| 3575 | Water | Water from the initial dispenser line |
| 3576 | Water | Water from the final dispenser line |
| 3577 | Surface | Feeders |
| 3578 | Surface | Drinkers |
| 3579 | Surface | Hands of workers |
| 3580 | Surface | Walls |
| 3581 | Surface | Cages |
| 3582 | Surface | Window panels |
| 3583 | Surface | Footwear of workers |
| 3584 | Surface | Flooring |
| 3585 | Feed | Silo number 1 |
| 3586 | Feed | Silo number 2 |
| 3587 | Feed | Silo number 3 |
| 3588 | Feed | Silo number 4 |
| 3597 | Biological | Sheep’s faeces |
| 3598 | Biological | Sheep’s nasal swab |
| Sample ID | Local | Source | Sample Type | Detection of L. monocytogenes |
|---|---|---|---|---|
| 3572 | Outside of the farm | Environmental | Well water | − |
| 3585 | Outside of the farm | Environmental | Silo number 1 | − |
| 3586 | Outside of the farm | Environmental | Silo number 2 | − |
| 3587 | Outside of the farm | Environmental | Silo number 3 | − |
| 3588 | Outside of the farm | Environmental | Silo number 4 | − |
| 3597 | Outside of the farm | Environmental | Sheep’s faeces | + |
| 3598 | Outside of the farm | Environmental | Sheep’s nasal swab | − |
| 3579 | Between units | Environmental | Hands of workers | − |
| 3583 | Between units | Environmental | Footwear of workers | − |
| 3575 | Fattening unit | Environmental | Water from the initial dispenser line | − |
| 3576 | Fattening unit | Environmental | Water from the final dispenser line | + |
| 3573 | Maternity unit | Environmental | Water from the initial dispenser line | − |
| 3574 | Maternity unit | Environmental | Water from the final dispenser line | − |
| 3577 | Maternity unit | Environmental | Feeders | + |
| 3578 | Maternity unit | Environmental | Drinkers | − |
| 3580 | Maternity unit | Environmental | Walls | + |
| 3581 | Maternity unit | Environmental | Cages | + |
| 3582 | Maternity unit | Environmental | Window panels | − |
| 3584 | Maternity unit | Environmental | Flooring | + |
| 3589 | Maternity unit | Animal | Pool of uterus | + |
| 3590 | Maternity unit | Animal | Pool of uterus | + |
| 3591 | Maternity unit | Animal | Pool of liver | + |
| 3592 | Maternity unit | Animal | Pool of lung | + |
| 3593 | Maternity unit | Animal | Pool of spleen | + |
| 3594 | Maternity unit | Animal | Pool of stillborns | + |
| 3595 | Maternity unit | Animal | Pool of stillborns | + |
| 3596 | Maternity unit | Animal | Pool of stillborns | + |
| Strain ID | Local | Source | Sample | AST A | ARG | Lineage | CC | ST | |
|---|---|---|---|---|---|---|---|---|---|
| Shared | Differentially Present | ||||||||
| 3597F/2 | Outside of the farm | Environmental | Sheep faeces | Susceptible | tetM, fosX, lin, norB, lmrB, sul, mprF | None | II | 31 | 325 |
| 3576F/1 | Fattening unit | Environmental | Water from the final dispenser line | SXT | tetM, fosX, lin, norB, lmrB, sul, mprF | dfrD, mphB, lnuG, NarAB operon | II | 31 | 325 |
| 3577C/1 | Maternity unit | Environmental | Feeders | SXT | tetM, fosX, lin, norB, lmrB, sul, mprF | dfrD, mphB, lnuG, NarAB operon | II | 31 | 325 |
| 3580C/1 | Maternity unit | Environmental | Walls | SXT | tetM, fosX, lin, norB, lmrB, sul, mprF | dfrD, mphB, lnuG, NarAB operon | II | 31 | 325 |
| 3581C/3 | Maternity unit | Environmental | Cages | Susceptible | tetM, fosX, lin, norB, lmrB, sul, mprF | None | II | 31 | 325 |
| 3584F/2 | Maternity unit | Environmental | Flooring | Susceptible | tetM, fosX, lin, norB, lmrB, sul, mprF | None | II | 31 | 325 |
| 3589C/1 | Maternity unit | Animal | Uterus | Susceptible | tetM, fosX, lin, norB, lmrB, sul, mprF | None | II | 31 | 325 |
| 3590F/2 | Maternity unit | Animal | Uterus | SXT | tetM, fosX, lin, norB, lmrB, sul, mprF | dfrD, mphB, lnuG, NarAB operon | II | 31 | 325 |
| 3591C/1 | Maternity unit | Animal | Liver | SXT | tetM, fosX, lin, norB, lmrB, sul, mprF | dfrD, mphB, lnuG, NarAB operon | II | 31 | 325 |
| 3591C/2 | Maternity unit | Animal | Liver | SXT | tetM, fosX, lin, norB, lmrB, sul, mprF | dfrD, mphB, lnuG, NarAB operon | II | 31 | 325 |
| 3592C/1 | Maternity unit | Animal | Lung | SXT | tetM, fosX, lin, norB, lmrB, sul, mprF | dfrD, mphB, lnuG, NarAB operon | II | 31 | 325 |
| 3592C/2 | Maternity unit | Animal | Lung | SXT | tetM, fosX, lin, norB, lmrB, sul, mprF | dfrD, mphB, lnuG, NarAB operon | II | 31 | 325 |
| 3593C/1 | Maternity unit | Animal | Spleen | SXT | tetM, fosX, lin, norB, lmrB, sul, mprF | dfrD, mphB, lnuG, NarAB operon | II | 31 | 325 |
| 3593C/5 | Maternity unit | Animal | Spleen | SXT | tetM, fosX, lin, norB, lmrB, sul, mprF | dfrD, mphB, lnuG, NarAB operon | II | 31 | 325 |
| 3594C/1 | Maternity unit | Animal | Stillborns | SXT | tetM, fosX, lin, norB, lmrB, sul, mprF | dfrD, mphB, lnuG, NarAB operon | II | 31 | 325 |
| 3595C/1 | Maternity unit | Animal | Stillborns | SXT | ND | ND | ND | ND | ND |
| 3595C/2 | Maternity unit | Animal | Stillborns | SXT | ND | ND | ND | ND | ND |
| 3596C/1 | Maternity unit | Animal | Stillborns | SXT | ND | ND | ND | ND | ND |
| 3596C/2 | Maternity unit | Animal | Stillborns | SXT | tetM, fosX, lin, norB, lmrB, sul, mprF | dfrD, mphB, lnuG, NarAB operon | II | 31 | 325 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, I.C.; Ribeiro-Almeida, M.; Silveira, L.; Prata, J.C.; de Carvalho, A.P.; Roque, C.; Gomes, J.P.; Borges, V.; Pista, Â.; Martins da Costa, P. Unveiling a Listeria monocytogenes Outbreak in a Rabbit Farm: Clinical Manifestation, Antimicrobial Resistance, Genomic Insights and Environmental Investigation. Microorganisms 2024, 12, 785. https://doi.org/10.3390/microorganisms12040785
Rodrigues IC, Ribeiro-Almeida M, Silveira L, Prata JC, de Carvalho AP, Roque C, Gomes JP, Borges V, Pista Â, Martins da Costa P. Unveiling a Listeria monocytogenes Outbreak in a Rabbit Farm: Clinical Manifestation, Antimicrobial Resistance, Genomic Insights and Environmental Investigation. Microorganisms. 2024; 12(4):785. https://doi.org/10.3390/microorganisms12040785
Chicago/Turabian StyleRodrigues, Inês C., Marisa Ribeiro-Almeida, Leonor Silveira, Joana C. Prata, André Pinto de Carvalho, Carla Roque, João Paulo Gomes, Vítor Borges, Ângela Pista, and Paulo Martins da Costa. 2024. "Unveiling a Listeria monocytogenes Outbreak in a Rabbit Farm: Clinical Manifestation, Antimicrobial Resistance, Genomic Insights and Environmental Investigation" Microorganisms 12, no. 4: 785. https://doi.org/10.3390/microorganisms12040785