
New Insights into the Production of Assyrtiko Wines from the Volcanic Terroir of Santorini Island Using Lachancea thermotolerans


 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Microbial Strains and Culture Conditions
2.2. Experimental Design and Winemaking Conditions
2.3. Chemical Analyses
2.3.1. Standard Oenological Parameters
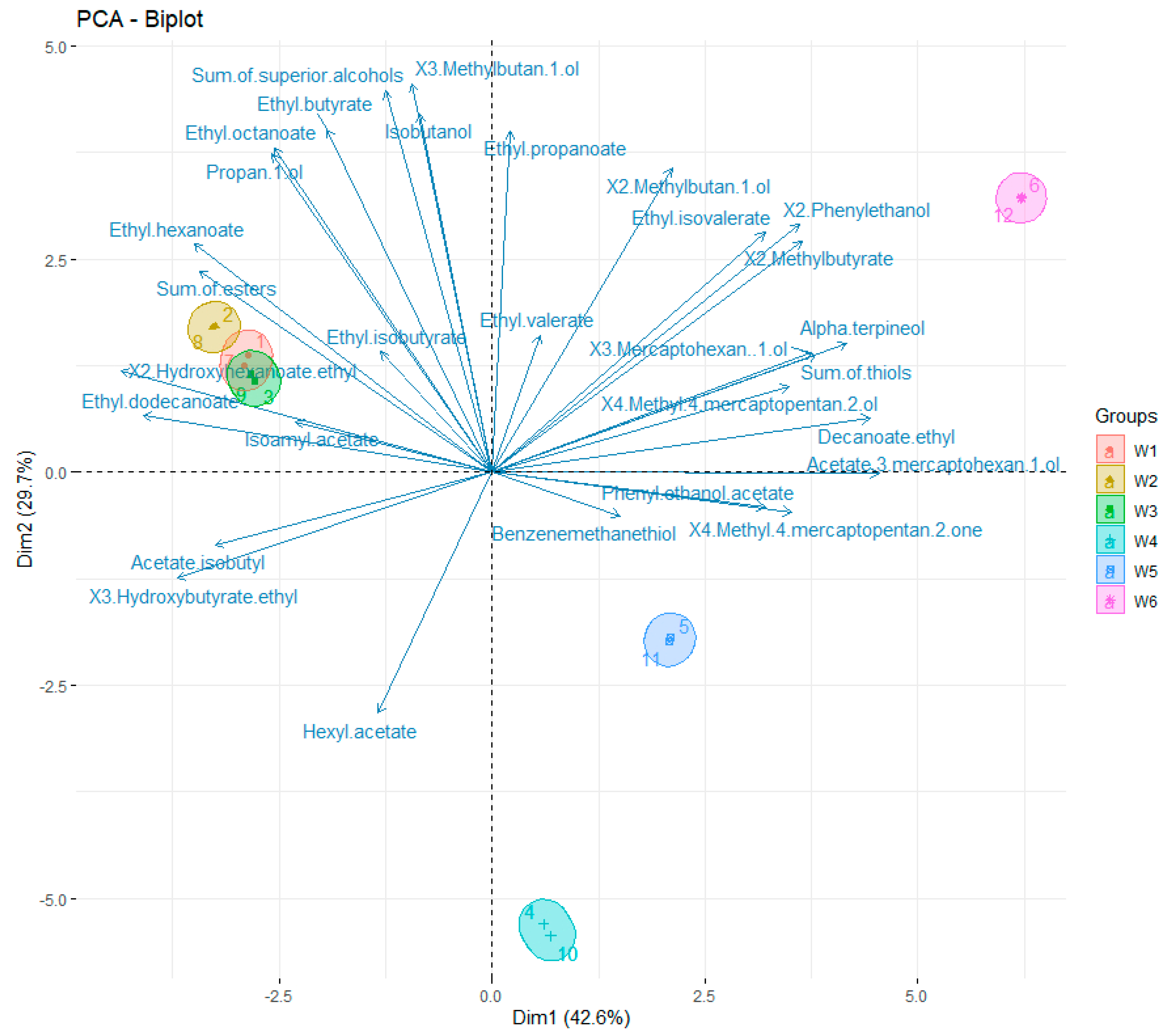
2.3.2. Identification and Quantification of Volatile Compounds
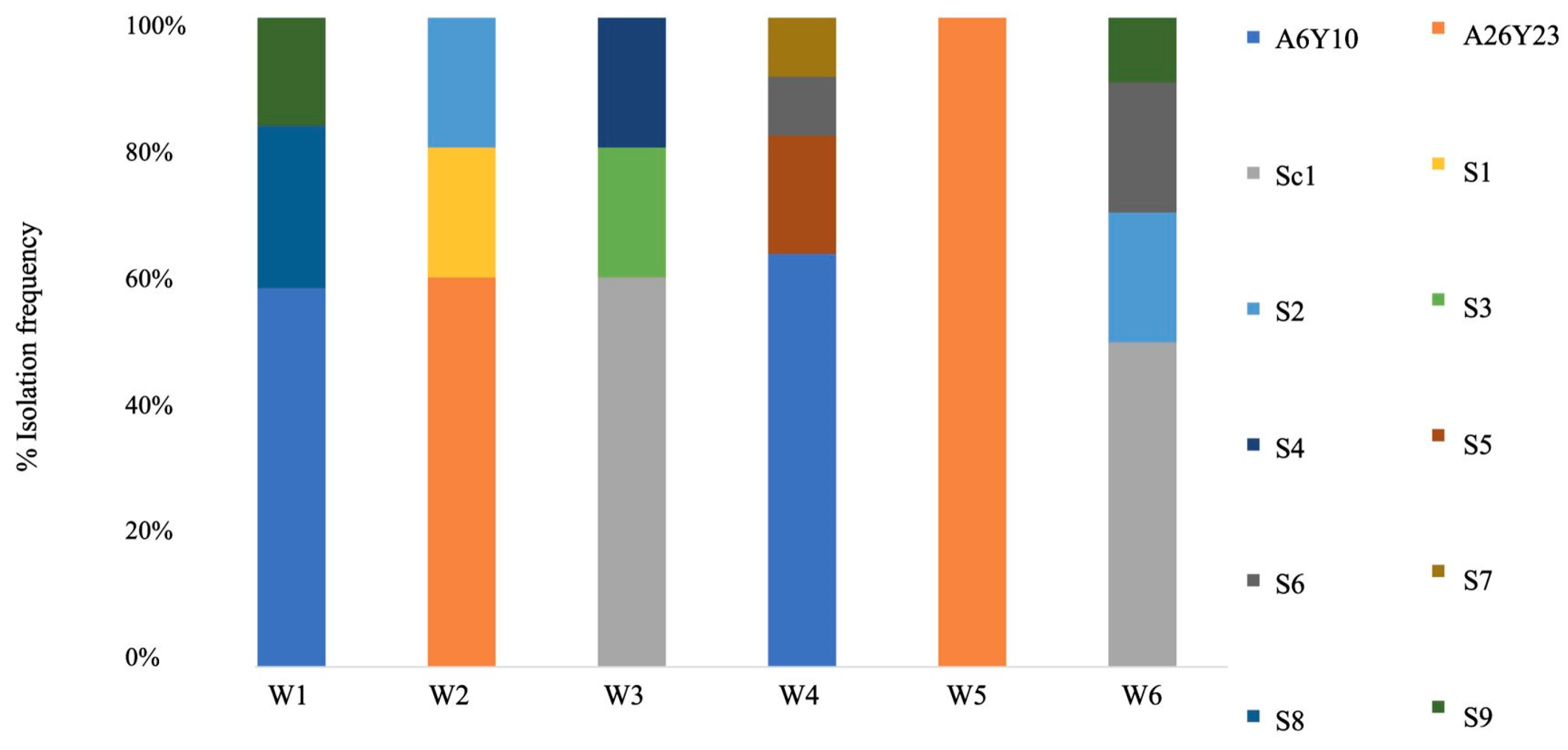
2.4. Microbiological Analyses and Molecular Typing
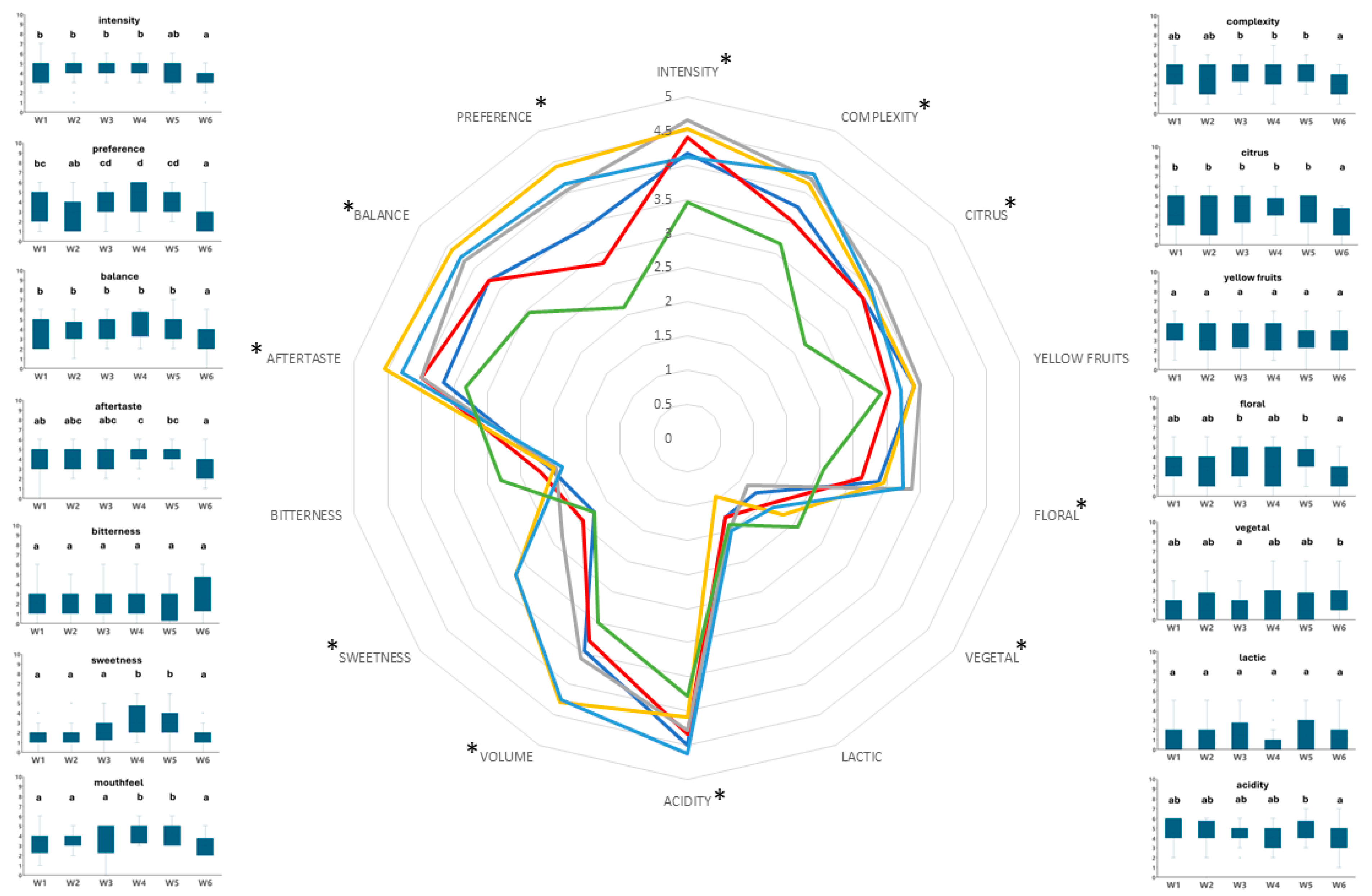
2.5. Sensory Analysis
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Contreras, A.; Curtin, C.; Varela, C. Yeast population dynamics reveal a potential ‘collaboration’ between Metschnikowia pulcherrima and Saccharomyces uvarum for the production of reduced alcohol wines during Shiraz fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 1885–1895. [Google Scholar] [CrossRef] [PubMed]
- Canonico, L.; Comitini, F.; Oro, L.; Ciani, M. Sequential fermentation with selected immobilized non-Saccharomyces yeast for reduction of ethanol content in wine. Front. Microbiol. 2016, 7, 278. [Google Scholar] [CrossRef] [PubMed]
- Rocker, J.; Strub, S.; Ebert, K.; Grossmann, M. Usage of different aerobic non-Saccharomyces yeasts and experimental conditions as a tool for reducing the potential ethanol content in wines. Eur. Food Res. Technol. 2016, 242, 2051–2070. [Google Scholar] [CrossRef]
- Varela, C.; Sengler, F.; Solomon, M.; Curtin, C. Volatile flavour profile of reduced alcohol wines fermented with the non-conventional yeast species Metschnikowia pulcherrima and Saccharomyces uvarum. Food Chem. 2016, 209, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Mestre Furlani, M.V.; Maturano, Y.P.; Combina, M.; Mercado, L.A.; Toro, M.E.; Vazquez, F. Selection of non-Saccharomyces yeasts to be used in grape musts with high alcoholic potential: A strategy to obtain wines with reduced ethanol content. FEMS Yeast Res. 2017, 17, fox010. [Google Scholar] [CrossRef] [PubMed]
- Puertas, B.; Jiménez, M.J.; Cantos-Villar, E.; Cantoral, J.M.; Rodríguez, M.E. Use of Torulaspora delbrueckii and Saccharomyces cerevisiae in semi-industrial sequential inoculation to improve quality of Palomino and Chardonnay wines in warm climates. J. Appl. Microbiol. 2017, 122, 733–746. [Google Scholar] [CrossRef]
- Dutraive, O.; Benito, S.; Fritsch, S.; Beisert, B.; Patz, C.-D.; Rauhut, D. Effect of sequential inoculation with non-Saccharomyces and Saccharomyces yeasts on Riesling wine chemical composition. Fermentation 2019, 5, 79. [Google Scholar] [CrossRef]
- Englezos, V.; Pollon, M.; Rantsiou, K.; Ortiz-Julien, A.; Botto, R.; Río Segade, S.; Giacosa, S.; Rolle, L.; Cocolin, L. Saccharomyces cerevisiae-Starmerella bacillaris strains interaction modulates chemical and volatile profile in red wine mixed fermentations. Food Res. Int. 2019, 122, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.; Esteve-Zarzoso, B.; Cabellos, J.M.; Arroyo, T. Sequential non-Saccharomyces and Saccharomyces cerevisiae fermentations to reduce the alcohol content in wine. Fermentation 2020, 6, 60. [Google Scholar] [CrossRef]
- Hranilovic, A.; Gambetta, J.M.; Jeffery, D.W.; Grbin, P.R.; Jiranek, V. Lower-alcohol wines produced by Metschnikowia pulcherrima and Saccharomyces cerevisiae co-fermentations: The effect of sequential inoculation timing. Int. J. Food Microbiol. 2020, 329, 108651. [Google Scholar] [CrossRef]
- Lee, S.-B.; Park, H.-D. Isolation and investigation of potential non-Saccharomyces yeasts to improve the volatile terpene compounds in Korean Muscat Bailey A wine. Microorganisms 2020, 8, 1552. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wang, Q.-Q.; Xu, Y.-H.; Li, A.-H.; Tao, Y.-S. Increased glycosidase activities improved the production of wine varietal odorants in mixed fermentation of P. fermentans and high antagonistic S. cerevisiae. Food Chem. 2020, 332, 127426. [Google Scholar] [CrossRef] [PubMed]
- Quincozes, L.; Marcon, Â.R.; Spinelli, F.R.; Gabbardo, M.; Eckhardt, D.P.; Cunha, W.M.d.; Costa, V.B.; Jacques, R.J.S.; Schumacher, R.L. Physicochemical, aromatic and sensory properties of the ‘Riesling Italico’ wines fermented with Saccharomyces and non-Saccharomyces yeasts. Cienc. Rural 2020, 50, e20190622. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, H.; Xue, J.; Tang, C.; Duan, C.; Yan, G. Use of Torulaspora delbrueckii and Hanseniaspora vineae co-fermentation with Saccharomyces cerevisiae to improve aroma profiles and safety quality of Petit Manseng wines. LWT-Food Sci. Technol. 2022, 161, 113360. [Google Scholar] [CrossRef]
- Han, B.; Gao, J.; Han, X.; Deng, H.; Wu, T.; Li, C.; Zhan, J.; Huang, W.; You, Y. Hanseniaspora uvarum FS35 degrades putrescine in wine through the direct oxidative deamination pathway of copper amine oxidase 1. Food Res. Int. 2022, 162, 111923. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yan, J.; Zhang, W.; Zhang, Y.; Dong, Z.; Luo, H.; Liu, M.; Su, J. Comparison of potential Wickerhamomyces anomalus to improve the quality of Cabernet Sauvignon wines by mixed fermentation with Saccharomyces cerevisiae. LWT-Food Sci. Technol. 2023, 173, 114285. [Google Scholar] [CrossRef]
- Xia, H.; Zhang, Z.; Sun, L.; Zhang, Q.; Zhang, J. effects of mixed fermentation on the aroma compounds of ‘Italian Riesling’ dry white wine in eastern foothill of Helan mountain. Fermentation 2023, 9, 303. [Google Scholar] [CrossRef]
- Maccaferri, S.; Klinder, A.; Brigidi, P.; Cavina, P.; Costabile, A. Potential probiotic Kluyveromyces marxianus B0399 modulates the immune response in Caco-2 cells and peripheral blood mononuclear cells and impacts the human gut microbiota in an in vitro colonic model system. Appl. Environ. Microbiol. 2012, 78, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef]
- Smith, I.M.; Baker, A.; Arneborg, N.; Jespersen, L. Non-Saccharomyces yeasts protect against epithelial cell barrier disruption induced by Salmonella enterica subsp. enterica serovar Typhimurium. Lett. Appl. Microbiol. 2015, 61, 491–497. [Google Scholar] [CrossRef]
- Villalba, M.L.; Susana Sáez, J.; del Monaco, S.; Lopes, C.A.; Sangorrín, M.P. TdKT, a new killer toxin produced by Torulaspora delbrueckii effective against wine spoilage yeasts. Int. J. Food Microbiol. 2016, 217, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Simonin, S.; Alexandre, H.; Nikolantonaki, M.; Coelho, C.; Tourdot-Maréchal, R. Inoculation of Torulaspora delbrueckii as a bio-protection agent in winemaking. Food Res. Int. 2018, 107, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Simonin, S.; Roullier-Gall, C.; Ballester, J.; Schmitt-Kopplin, P.; Quintanilla-Casas, B.; Vichi, S.; Peyron, D.; Alexandre, H.; Tourdot-Maréchal, R. Bio-protection as an alternative to sulphites: Impact on chemical and microbial characteristics of red wines. Front. Microbiol. 2020, 11, 1308. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Agarbati, A.; Canonico, L.; Galli, E.; Ciani, M. Purification and characterization of WA18, a new mycocin produced by Wickerhamomyces anomalus active in wine against Brettanomyces bruxellensis spoilage yeasts. Microorganisms 2020, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Windholtz, S.; Redon, P.; Lacampagne, S.; Farris, L.; Lytra, G.; Cameleyre, M.; Barbe, J.-C.; Coulon, J.; Thibon, C.; Masneuf-Pomarède, I. Non-Saccharomyces yeasts as bioprotection in the composition of red wine and in the reduction of sulfur dioxide. LWT-Food Sci. Technol. 2021, 149, 111781. [Google Scholar] [CrossRef]
- Van Leeuwen, C.; Destrac-Irvine, A. Modified grape composition under climate change conditions requires adaptations in the vineyard. OENO One 2017, 51, 147–154. [Google Scholar] [CrossRef]
- Fazio, N.A.; Russo, N.; Foti, P.; Pino, A.; Caggia, C.; Randazzo, C.L. Inside current winemaking challenges: Exploiting the potential of conventional and unconventional yeasts. Microorganisms 2023, 11, 1338. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.; Navascués, E.; Calderón, F.; Santos, A.; Marquina, D.; Benito, S. An integrative view of the role of Lachancea thermotolerans in wine technology. Foods 2021, 10, 2878. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.; Hofmann, T.; Laier, M.; Lochbühler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on quality and composition of Riesling wines fermented by sequential inoculation with non-Saccharomyces and Saccharomyces cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Balikci, E.K.; Tanguler, H.; Jolly, N.P.; Erten, H. Influence of Lachancea thermotolerans on cv. Emir wine fermentation. Yeast 2016, 33, 313–321. [Google Scholar] [CrossRef]
- Whitener, M.B.; Stanstrup, J.; Carlin, S.; Divol, B.; Du Toit, M.; Vrhovsek, U. Effect of non-Saccharomyces yeasts on the volatile chemical profile of Shiraz wine. Aust. J. Grape Wine Res. 2017, 23, 179–192. [Google Scholar] [CrossRef]
- Castrillo, D.; Rabuñal, E.; Neira, N.; Blanco, P. Oenological potential of non-Saccharomyces yeasts to mitigate effects of climate change in winemaking: Impact on aroma and sensory profiles of Treixadura wines. FEMS Yeast Res. 2019, 19, foz065. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Bañuelos, M.A.; Vaquero, C.; Loira, I.; Cuerda, R.; Palomero, F.; González, C.; Suárez-Lepe, J.A.; Wang, J.; Han, S.; et al. Lachancea thermotolerans as a tool to improve pH in red wines from warm regions. Eur. Food Res. Technol. 2019, 245, 885–894. [Google Scholar] [CrossRef]
- Blanco, P.; Rabuñal, E.; Neira, N.; Castrillo, D. Dynamic of Lachancea thermotolerans population in monoculture and mixed fermentations: Impact on wine characteristics. Beverages 2020, 6, 36. [Google Scholar] [CrossRef]
- Romani, C.; Lencioni, L.; Bartolini, A.B.; Ciani, M.; Mannazzu, I.; Domizio, P. Pilot scale fermentations of Sangiovese: An overview on the impact of Saccharomyces and non-Saccharomyces wine yeasts. Fermentation 2020, 6, 63. [Google Scholar] [CrossRef]
- Fairbairn, S.; Engelbrecht, L.; Setati, M.E.; du Toit, M.; Bauer, F.F.; Divol, B.; Rossouw, D. Combinatorial analysis of population dynamics, metabolite levels and malolactic fermentation in Saccharomyces cerevisiae/Lachancea thermotolerans mixed fermentations. Food Microbiology 2021, 96, 103712. [Google Scholar] [CrossRef] [PubMed]
- Hranilovic, A.; Albertin, W.; Capone, D.L.; Gallo, A.; Grbin, P.R.; Danner, L.; Bastian, S.E.; Masneuf-Pomarede, I.; Coulon, J.; Bely, M.; et al. Impact of Lachancea thermotolerans on chemical composition and sensory profiles of Merlot wines. Food Chem. 2021, 349, 129015. [Google Scholar] [CrossRef] [PubMed]
- Korenika, A.-M.J.; Tomaz, I.; Preiner, D.; Lavrić, M.; Šimić, B.; Jeromel, A. Influence of L. thermotolerans and S. cerevisiae commercial yeast sequential inoculation on aroma composition of red wines (Cv Trnjak, Babic, Blatina and Frankovka). Fermentation 2021, 7, 4. [Google Scholar] [CrossRef]
- Snyder, E.C.; Jiranek, V.; Hranilovic, A. Impact of Lachancea thermotolerans strain and lactic acid concentration on Oenococcus oeni and malolactic fermentation in wine. OENO One 2021, 55, 365–380. [Google Scholar] [CrossRef]
- Ženišová, K.; Cabicarová, T.; Sidari, R.; Kolek, E.; Pangallo, D.; Szemes, T.; Kuchta, T. Effects of co-fermentation with Lachancea thermotolerans or Metschnikowia pulcherrima on concentration of aroma compounds in Pinot Blanc wine. J. Food Nutr. Res. 2021, 60, 87–91. [Google Scholar]
- Vaquero, C.; Loira, I.; Heras, J.M.; Carrau, F.; González, C.; Morata, A. Biocompatibility in ternary fermentations with Lachancea thermotolerans, other non-Saccharomyces and Saccharomyces cerevisiae to control pH and improve the sensory profile of wines from warm areas. Front. Microbiol. 2021, 12, 832. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Guzzon, R.; Ongaro, M.; Paolini, M.; Nardin, T.; Malacarne, M.; Roman, T.; Larcher, R. Biological acidification of “Vino Santo di Gambellara” by mixed fermentation of L. thermotolerans and S. cerevisiae. Role of nitrogen in the evolution of fermentation and aroma profile. OENO One 2023, 57, 205–217. [Google Scholar] [CrossRef]
- Mucalo, A.; Budic-Leto, I.; Zdunic, G. Effect of sequential fermentation with Lachancea thermotolerans/S. cerevisiae on aromatic and flavonoid profiles of Plavac Mali wine. Foods 2023, 12, 1912. [Google Scholar] [CrossRef] [PubMed]
- Tzamourani, A.P.; Taliadouros, V.; Paraskevopoulos, I.; Dimopoulou, M. Developing a novel selection method for alcoholic fermentation starters by exploring wine yeast microbiota from Greece. Front. Microbiol. 2023, 14, 1301325. [Google Scholar] [CrossRef] [PubMed]
- International Organisation of Vine and Wine. Compendium of International Methods of Wine and Must Analysis; OIV: Paris, France, 2020. [Google Scholar]
- Cozzolino, D.; Parker, M.; Dambergs, R.G.; Herderich, M.; Gishen, M. Chemometrics and visible-near infrared spectroscopic monitoring of red wine fermentation in a pilot scale. Biotechnol. Bioeng. 2006, 95, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulou, M.; Goulioti, E.; Troianou, V.; Toumpeki, C.; Paramithiotis, S.; Gosselin, Y.; Dorignac, E.; Papadopoulos, G.; Kotseridis, Y. Effect of Saccharomyces cerevisiae and Saccharomyces pastorianus co-inoculation on alcoholic fermentation behavior and aromatic profile of Sauvignon Blanc wine. Fermentation 2022, 8, 539. [Google Scholar] [CrossRef]
- Tominaga, T.; Furrer, A.; Henry, R.; Dubourdieu, D. Identification of new volatile thiols in the aroma of Vitis vinifera L. var. Sauvignon blanc wines. Flavour Fragr. J. 1998, 13, 159–162. [Google Scholar] [CrossRef]
- Heymann, H.; Ebeler, S.E. Sensory and Instrumental Evaluation of Alcoholic Beverages; Elsevier Inc.: London, UK, 2017. [Google Scholar]
- Gentilesco, G.; Coletta, A.; Tarricone, L.; Alba, V. Bioclimatic characterization relating to temperature and subsequent future scenarios of vine growing across the Apulia region in southern Italy. Agriculture 2023, 13, 644. [Google Scholar] [CrossRef]
- Wagner, M.; Stanbury, P.; Dietrich, T.; Döring, J.; Ewert, J.; Foerster, C.; Freund, M.; Friedel, M.; Kammann, C.; Koch, M.; et al. Developing a sustainability vision for the global wine industry. Sustainability 2023, 15, 10487. [Google Scholar] [CrossRef]
- Alsafadi, K.; Bi, S.; Bashir, B.; Alsalman, A.; Srivastava, A.K. Future scenarios of bioclimatic viticulture indices in the eastern Mediterranean: Insights into sustainable vineyard management in a changing climate. Sustainability 2023, 15, 11740. [Google Scholar] [CrossRef]
- Omazić, B.; Prtenjak, M.T.; Bubola, M.; Meštrić, J.; Karoglan, M.; Prša, I. Application of statistical models in the detection of grapevine phenology changes. Agric. For. Meteorol. 2023, 341, 109682. [Google Scholar] [CrossRef]
- Castillo, N.; Cavazos, T.; Pavia, E.G. Impact of climate change in Mexican winegrape regions. Int. J. Climatol. 2023, 43, 6621–6642. [Google Scholar] [CrossRef]
- Massano, L.; Fosser, G.; Gaetani, M.; Bois, B. Assessment of climate impact on grape productivity: A new application for bioclimatic indices in Italy. Sci. Total Environ. 2023, 905, 167134. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Tejera, O.; Bonada, M.; Petrie, P.R.; Nieto, H.; Bellvert, J.; Sadras, V.O. Viticulture adaptation to global warming: Modelling gas exchange, water status and leaf temperature to probe for practices manipulating water supply, canopy reflectance and radiation load. Agric. For. Meteorol. 2023, 331, 109351. [Google Scholar] [CrossRef]
- Sun, Q.; Granco, G.; Groves, L.; Voong, J.; Van Zyl, S. Viticultural manipulation and new technologies to address environmental challenges caused by climate change. Climate 2023, 11, 83. [Google Scholar] [CrossRef]
- Santos, I.V.; Renaud-Gentié, C.; Roux, P.; Levasseur, A.; Bulle, C.; Deschênes, L.; Boulay, A.-M. Prospective life cycle assessment of viticulture under climate change scenarios, application on two case studies in France. Sci. Total Environ. 2023, 880, 163288. [Google Scholar] [CrossRef] [PubMed]
- Lakatos, L.; Mitre, Z. Effect of drought on the future sugar content of wine grape varieties till 2100: Possible adaptation in the Hungarian Eger wine region. Biomolecules 2023, 13, 1143. [Google Scholar] [CrossRef]
- Bohnert, G.; Martin, B. Impacts of climate change induced drought and adaptation strategies in wine-growing in the Rhine Valley (France, Germany, Switzerland). Total Environ. Res. Themes 2023, 8, 100081. [Google Scholar] [CrossRef]
- Payan, C.; Gancel, A.-L.; Jourdes, M.; Christmann, M.; Teissedre, P.-L. Wine acidification methods: A review. OENO One 2023, 57, 113–126. [Google Scholar] [CrossRef]
- Vicente, J.; Baran, Y.; Navascues, E.; Santos, A.; Calderon, F.; Marquina, D.; Rauhut, D.; Benito, S. Biological management of acidity in wine industry: A review. Int. J. Food Microbiol. 2022, 375, 109726. [Google Scholar] [CrossRef]
- Vicente, J.; Kelanne, N.; Rodrigo-Burgos, L.; Navascués, E.; Calderón, F.; Santos, A.; Marquina, D.; Yang, B.; Benito, S. Influence of different Lachancea thermotolerans strains in the wine profile in the era of climate challenge. FEMS Yeast Res. 2023, 23, foac062. [Google Scholar] [CrossRef] [PubMed]
- Kapsopoulou, K.; Mourtzini, A.; Anthoulas, M.; Nerantzis, E. Biological acidification during grape must fermentation using mixed cultures of Kluyveromyces thermotolerans and Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2007, 23, 735–739. [Google Scholar] [CrossRef]
- Mendoza, L.M.; Manca de Nadra, M.C.; Bru, E.; Farías, M.E. Influence of wine related physicochemical factors on the growth and metabolism of non-Saccharomyces and Saccharomyces in mixed culture. J. Ind. Microbiol. Biotechnol. 2009, 36, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, L.M.; Manca de Nadra, M.C.; Farías, M.E. Kinetics and metabolic behaviour of a composite culture of Kloeckera apiculata and Saccharomyces cerevisiae wine related strains. Biotechnol. Lett. 2007, 29, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Beco, L.; Comitini, F. Fermentation behaviour and metabolic interactions of multistarter wine yeast fermentations. Int. J. Food Microbiol. 2006, 108, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Benito, S. The impacts of Lachancea thermotolerans yeast strains on winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 6775–6790. [Google Scholar] [CrossRef] [PubMed]
- Gabler, A.M.; Ludwig, A.; Frank, O.; Dawid, C. NMR-based tastant polymer interaction studies and the influence on the taste perception of red wine. J. Agric. Food Chem. 2023, 71, 18454–18465. [Google Scholar] [CrossRef] [PubMed]
- Ferrero-del-Teso, S.; Arapitsas, P.; Jeffery, D.W.; Ferreira, C.; Mattivi, F.; Fernandez-Zurbano, P.; Saenz-Navajas, M.-P. Exploring UPLC-QTOF-MS-based targeted and untargeted approaches for understanding wine mouthfeel: A sensometabolomic approach. Food Chem. 2024, 437, 137726. [Google Scholar] [CrossRef]
- Jones, P.R.; Gawel, R.; Francis, I.L.; Waters, E.J. The influence of interactions between major white wine components on the aroma, flavour and texture of model white wine. Food Qual. Prefer. 2008, 19, 596–607. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Code | Inoculum |
|---|---|
| W1 | L. thermotolerans strain Lt1—S. cerevisiae strain A6Y10 |
| W2 | L. thermotolerans strain Lt1—S. cerevisiae strain A26Y23 |
| W3 | L. thermotolerans strain Lt1—S. cerevisiae strain Sc1 |
| W4 | S. cerevisiae strain A6Y10 |
| W5 | S. cerevisiae strain A26Y23 |
| W6 | S. cerevisiae strain Sc1 |
| W1 | W2 | W3 | W4 | W5 | W6 | |
|---|---|---|---|---|---|---|
| Fermentation duration (d) | 12 (0) | 12 (0) | 12 (0) | 18 (0) | 18 (0) | 8 (0) |
| TAV (% v/v) | 13.7 (0.1) ab | 13.7 (0.2) ab | 13.6 (0.2) ab | 13.8 (0.1) b | 13.8 (0.1) b | 13.5 (0.2) a |
| Glucose + Fructose (g/L) | 0.7 (0.01) c | 0.7 (0.03) c | 0.8 (0.04) d | 1.7 (0.01) e | 0.4 (0.04) a | 0.5 (0.03) b |
| Total acidity (g/L) | 7.26 (0.14) c | 7.14 (0.21) b c | 7.23 (0.32) c | 6.48 (0.16) a | 6.90 (0.12) b | 6.48 (0.22) a |
| Volatile acidity (g/L) | 0.43 (0.01) b | 0.41 (0.01) b | 0.41 (0.01) b | 0.60 (0.03) c | 0.61 (0.02) c | 0.20 (0.02) a |
| pH | 2.9 (0.01) a | 2.9 (0.05) a | 2.9 (0.07) a | 3.0 (0.04) a | 3.0 (0.08) a | 2.9 (0.09) a |
| L-lactic acid (g/L) | 1.0 (0.04) b | 1.0 (0.06) b | 1.1 (0.02) c | 0 (0.0) a | 0 (0.0) a | 0 (0.0) a |
| L-malic acid (g/L) | 1.0 (0.05) a | 1.0 (0.03) a | 1.0 (0.02) a | 1.2 (0.05) c | 1.3 (0.02) d | 1.1 (0.05) b |
| Total SO2 (mg/L) | 87 (3.1) a | 99 (4.7) b | 103 (6.8) b | 117 (5.5) c | 107 (6.7) b | 105 (4.9) b |
| Free SO2 (mg/L) | 17 (1.2) a | 23 (2.1) bc | 22 (2.2) ab | 28 (4.6) cd | 22 (3.5) ab | 31 (3.3) d |
| W1 | W2 | W3 | W4 | W5 | W6 | |
|---|---|---|---|---|---|---|
| Higher alcohols (mg/L) | ||||||
| 2-Methylbutan-1-ol | 25.7 (0.42) b | 25.2 (0.49) b | 24.9 (0.21) b | 15.1 (0.14) a | 29.6 (0.28) c | 33.5 (0.21) d |
| 3-Methylbutan-1-ol | 221 (1.4) cd | 242 (7.6) e | 218 (2.7) c | 107 (0.6) a | 152 (2.7) b | 228 (4.1) d |
| Isobutanol | 29.7 (0.49) b | 32.6 (0.21) c | 33.0 (0.49) c | 22.2 (0.42) a | 21.75 (0.48) a | 33.1 (0.43) c |
| Propan-1-ol | 81.1 (0.63) d | 92.8 (0.64) e | 82.8 (1.48) d | 23.0 (0.63) a | 55.3 (0.62) b | 60.9 (0.42) c |
| Thiols (ng/L) | ||||||
| 4-methyl-4-mercaptopentan-2-one | 0 (0.0) a | 3.05 (0.212) b | 3.00 (0.141) b | 3.30 (0.141) b | 6.95 (0.212) d | 6.05 (0.221) c |
| 3-mercaptohexan-1-ol | 704 (2.6) c | 676 (8.3) b | 674 (7.1) b | 1245 (10.2) d | 500 (3.2) a | 2755 (20.0) e |
| Acetate 3-mercaptohexan-1-ol | 56.3 (1.62) a | 58.7 (0.56) a | 66.0 (0.99) d | 82.8 (1.20) b | 78.5 (1.34) c | 108 (3.3) e |
| 4-methyl-4-mercaptopentan-2-ol | 18.8 (0.28) a | 20.8 (1.10) b | 21.8 (0.49) b | 17.8 (0.63) a | 32.1 (1.06) d | 29.1 (0.77) c |
| Benzenemethanethiol | 2.20 (0.141) e | 0 (0.0) a | 0.475 (0.035) b | 1.18 (0.021) cd | 1.08 (0.169) c | 1.35 (0.063) d |
| Ethyl esters (mg/L) | ||||||
| Ethyl propanoate | 333 (5.6) c | 289 (14.8) b | 288 (18.4) b | 182 (17.7) a | 282 (9.2) b | 329 (7.1) c |
| Ethyl isobutyrate | 109 (5.6) c | 105 (4.9) bc | 97.5 (2.12) b | 104 (2.1) bc | 43.5 (4.94) a | 104 (1.4) bc |
| Ethyl butyrate | 461 (1.1) cd | 467 (3.4) d | 485 (4.4) e | 398 (1.3) a | 425 (6.1) b | 454 (2.0) c |
| Ethyl 2-methylbutyrate | 10.9 (0.14) ab | 11.8 (1.13) b | 11.4 (0.63) b | 9.7 (0.35) a | 11.2 (0.34) ab | 20.6 (0.92) c |
| Ethyl isovalerate | 25.9 (1.55) bc | 24.3 (0.99) bc | 24.8 (0.21) bc | 22.5 (0.63) b | 19.4 (0.78) a | 53.5 (2.12) d |
| Ethyl valerate | 3.60 (0.565) ab | 3.90 (0.141) ab | 4.40 (0.565) bc | 3.30 (0.424) a | 5.10 (0.142) c | 4.30 (0.422) abc |
| Ethyl hexanoate | 1188 (16.2) d | 1254 (21.2) e | 1252 (32.5) e | 927 (12.7) a | 1092 (14.8) c | 1034 (31.8) b |
| Ethyl octanoate | 2892 (16.2) e | 2716 (26.8) d | 2679 (26.2) d | 1631 (34.6) a | 1972 (23.3) b | 2365 (41.7) c |
| Ethyl 3-hydroxybutyrate | 220 (1.4) b | 211 (11.3) b | 250 (11.4) c | 264 (4.9) c | 0 (0.0) a | 0 (0.0) a |
| Ethyl 2-hydroxyhexanoate | 649 (20.5) c | 720 (2.8) d | 624 (20.5) c | 262 (7.8) b | 0 (0.0) a | 0 (0.0) a |
| Ethyl decanoate | 95.2 (1.06) c | 51.8 (1.13) a | 64.6 (1.98) b | 120 (4.2) d | 310 (2.8) e | 402 (5.6) f |
| Ethyl dodecanoate | 105 (5.6) c | 96.5 (6.36) c | 83.1 (4.38) b | 77.1 (2.96) b | 17.3 (1.81) a | 22.9 (2.99) a |
| 2-phenyl ethanol | 9.05 (0.212) b | 9.20 (0.141) b | 10.5 (0.78) c | 5.45 (0.212) a | 11.5 (0.78) c | 26.0 (0.49) d |
| Acetate esters (mg/L) | ||||||
| Isobutyl acetate | 50.3 (0.99) d | 46.1 (1.20) c | 51.7 (0.99) d | 38.8 (1.13) b | 55.4 (0.84) e | 23.6 (0.56) a |
| Isoamyl acetate | 2607 (13.4) b | 2712 (35.3) c | 2690 (32.5) c | 1467 (26.8) a | 3343 (33.9) d | 1487 (19.1) a |
| Hexyl acetate | 122 (2.1) b | 122 (2.0) b | 133 (2.2) c | 136 (2.2) c | 195 (1.4) d | 67.0 (1.42) a |
| Phenylethyl acetate | 31.4 (0.84) b | 11.6 (0.56) a | 41.3 (1.83) c | 32.9 (0.14) b | 135 (3.5) e | 82.6 (1.98) d |
| Terpenoids (mg/L) | ||||||
| Alpha-terpineol | 43.8 (1.69) b | 41.9 (0.14) b | 37.9 (1.48) a | 42.5 (0.78) b | 52.4 (1.99) c | 58.1 (1.27) d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tzamourani, A.; Paramithiotis, S.; Favier, M.; Coulon, J.; Moine, V.; Paraskevopoulos, I.; Dimopoulou, M. New Insights into the Production of Assyrtiko Wines from the Volcanic Terroir of Santorini Island Using Lachancea thermotolerans. Microorganisms 2024, 12, 786. https://doi.org/10.3390/microorganisms12040786
Tzamourani A, Paramithiotis S, Favier M, Coulon J, Moine V, Paraskevopoulos I, Dimopoulou M. New Insights into the Production of Assyrtiko Wines from the Volcanic Terroir of Santorini Island Using Lachancea thermotolerans. Microorganisms. 2024; 12(4):786. https://doi.org/10.3390/microorganisms12040786
Chicago/Turabian StyleTzamourani, Aikaterini, Spiros Paramithiotis, Marion Favier, Joana Coulon, Virginie Moine, Ioannis Paraskevopoulos, and Maria Dimopoulou. 2024. "New Insights into the Production of Assyrtiko Wines from the Volcanic Terroir of Santorini Island Using Lachancea thermotolerans" Microorganisms 12, no. 4: 786. https://doi.org/10.3390/microorganisms12040786
APA StyleTzamourani, A., Paramithiotis, S., Favier, M., Coulon, J., Moine, V., Paraskevopoulos, I., & Dimopoulou, M. (2024). New Insights into the Production of Assyrtiko Wines from the Volcanic Terroir of Santorini Island Using Lachancea thermotolerans. Microorganisms, 12(4), 786. https://doi.org/10.3390/microorganisms12040786